

Abstract
Background
Previous studies suggest that sensory information is “received” but not “perceived” under general anesthesia. Whether and to what extent the brain continues to process sensory inputs in a drug-induced unconscious state remain unclear.
Methods
107 rats were randomly assigned to 12 different anesthesia and odor exposure paradigms. The immunoreactivities of the immediate early gene products c-Fos and Egr1 as neural activity markers were combined with behavioral tests to assess the integrity and relationship of cellular and behavioral responsiveness to olfactory stimuli under a surgical plane of ketamine-xylazine general anesthesia.
Results
The olfactory sensory processing centers can distinguish the presence or absence of experimental odorants even when animals were fully anesthetized. In the anesthetized state, the c-Fos immunoreactivity in the higher olfactory cortices revealed a difference between novel and familiar odorants similar to that seen in the awake state, suggesting that the anesthetized brain functions beyond simply receiving external stimulation. Re-exposing animals to odorants previously experienced only under anesthesia resulted in c-Fos immunoreactivity similar to that elicited by familiar odorants, indicating that previous registration had occurred in the anesthetized brain. Despite the “cellular memory,” however, odor discrimination and forced-choice odor-recognition tests showed absence of behavioral recall of the registered sensations, except for a longer latency in odor recognition tests.
Conclusions
Histologically distinguishable registration of sensory process continues to occur at cellular level under ketamine-xylazine general anesthesia despite the absence of behavioral recognition, consistent with the notion that general anesthesia causes disintegration of information processing without completely blocking cellular communications.
INTRODUCTION
It remains a mystery whether and to what extent the brain in a drug-induced unconscious state, such as that under general anesthesia, continues to process and ultimately perceive sensory information. Electroencephalography measurements under deep general anesthesia have shown that cerebral cortices continue to respond to external stimuli even during burst-suppression,1,2 suggesting that information is still received in the cortices. However, empirically, it is believed that an unconscious brain cannot process and interpret external stimuli and that memories cannot be formed under, or immediately after, general anesthesia.3 Human experiences seem to suggest that a surgical plane of general anesthesia produces a brain physiological state that is behaviorally incompatible with conscious information processing, learning, or memory formation. At the cellular level, however, these assertions have not been rigorously tested experimentally. The global functioning of the brain in the clinical state of general anesthesia remains poorly understood.
Two schools of thought have prevailed in recent years on how general anesthesia might lead to unconsciousness. The first, developed based on the studies of somatosensory processes under general anesthesia, proposes that general anesthetics block a thalamocortical “gate” by suppressing or preventing the transfer of somatosensory information from the periphery to the cerebral cortex.4 The second theory contends that it is not the thalamocortical switch but the disintegration of information processing, particularly the disruption between the feed-forward and feedback loops and suppression of the feedback from the prefrontal cortex and parietal association cortex to the occipital cortex, that is responsible for the loss of consciousness (LOC).5 Experimental supports in favor of the second theory include the observations that electroencephalography, electrophysiology, and brain imaging measurements continue to show cortical activation even under anesthesia. It has been suggested that general anesthesia is a state in which “information is received but not perceived.”6
In this study, we investigated whether, and to what extent, anesthetized brains can actually process external stimuli. We chose to study the olfactory sensory system, in which thalamus and thalamocortical connection are not directly involved. Three brain regions—the olfactory bulb (OB), the anterior piriform cortex (APC), and the orbitofrontal cortex (OFC)—are essential for olfactory processing. The OB is the primary olfactory region for receiving sensory inputs and then integrating and projecting the inputs to the secondary olfactory processing regions, chiefly the APC where odorant identity is encoded.7 From there, the information is relayed to the tertiary processing centers in the OFC. In this study, we focused on the global responsiveness of APC and OFC to sensory stimulations, as these regions are part of the high-order cortices responsible for perceiving olfactory information. By quantifying the histological and behavioral registration of well-defined olfactory events in rats under a surgical plane of ketamine-xylazine general anesthesia, we determined if general anesthesia suppresses the level of input to the high-order olfactory processing centers and, more importantly, analyzed whether the discrimination and recognition of odorant novelty are preserved under general anesthesia.
MATERIALS AND METHODS
Experimental design principles
The experimental design of the current study took advantage of two unique manifestations of olfactory processing in rodents. The first was the ability of immediate early gene (IEG) products to quantify the cellular registration of olfactory stimulations.8,9 The second was the well-known behavioral characteristics of rats to exhibit interest in, and consequently longer exploration of, novel odorants as compared to familiar odorants. We created various binary olfactory conditions to compare histological registration by IEGs and behavioral recall of the very same olfactory event associated with fully awake and fully anesthetized states. IEGs are a class of rapidly and transiently activated genes. The immunoreactivity of the IEG products depends on the characteristics of neural activities.10 Two extensively studied IEGs, egr1 and c-fos, were used in this study for their unique properties as markers for neural activation11,12 and information integration,13–15 respectively. Specifically, c-Fos immunoreactivity is known to rapidly respond to novel sensory stimulations, with messenger RNA and protein levels detectable almost immediately and peaking at ~30 min and 2 h after stimulation onset, respectively.16 Animals kept in an odor-invariant environment maintain a low and constant basal c-Fos level. Therefore, c-Fos immunoreactivity can be used as a proxy for mapping neuronal registration of novel stimuli.8,17,18 Quantitative comparisons were made for specific changes in behavior and in c-Fos immunoreactivity for discrimination between familiar and novel odor stimulations, and for recognition among familiar, novel, and previously anesthesia-paired odorants.
Animals
All animal-handling procedures were approved by the Institutional Animal Care and Use Committee of the University of Pittsburgh, Pittsburgh, PA. Age-matched, seven-week old, male Sprague-Dawley rats were obtained from Harlan Laboratories and allowed to acclimate to a double-barrier housing facility for a week before any experiments. Animals were individually housed in airflow-regulated cages to minimize incidental odor contamination and were given food and water ad libitum. Animals were not deprived of background odor during housing but utmost caution was used to maintain the odor environment constant; even the personal hygiene products used by the experimenters and a dedicated animal caregiver were regulated and kept invariant throughout the experimental periods. During the experiments, the animal body temperature was carefully maintained between 36–38°C to prevent hypothermia, especially under general anesthesia. We did not measure other physiological parameters in the experimental groups but instead closely monitored the physiological state of all animals using the same criteria for normal physiology conditions as those found in other related studies in our laboratory. Introducing invasive physiological measurements in the experimental groups could introduce other confounding effects to the analyses, particularly IEG variations. Sharing the same physiological monitoring devices among different odor exposure groups can compromise both the novelty of olfactory cues and the integrity of the odor environment.
Experimental groups
One hundred and seven rats were randomly assigned to 12 experimental groups in three different experimental sets, as defined and color-coded in Table 1. The group notations in the table refer to the training/testing protocols. For example, “Odor/Odor” refers to training with the experimental odorants and testing with the same odorants (hence a familiar odor condition), “Air/Odor” refers to training without experimental odorants (Air) and testing with a novel odorant, and so on. The number of rats (n) in each group ranged from 6 to 24, determined by the power analysis on the basis of pilot studies. The complex design of 12 experimental groups provides important controls to correlate c-Fos immunoreactivity to novel odorant registration and to rule out any unforeseen effects from animal handling or anesthesia procedure alone. Two different odorant training protocols, termed repetitive or single session as depicted in Figure 1, were used. The first experimental set, consisting of Groups 1–5 (38 rats), used the repetitive training protocol and was experimented over a period of 8 days to establish the time dependence of the animals’ habituation to both the experimental environment and experimental odorants. Group 1 received minimal experimental manipulation to serve as the naïve control for histology. One day before the first training session, rats in Groups 2–5 were transferred to a clean training/testing cage and allowed to acclimate to the experimental setup for 30 min, during which the animals could freely interact with an empty, capless, 1.5-mL Eppendorf tube (Eppendorf, Hauffauge, NY) placed in the cage. On the following 6 training days, rats were again transferred to a clean training/testing cage and acclimated to the environment having an identical empty Eppendorf tube for 20 min. After the 20 min, the videotaping was initiated and the empty Eppendorf tube was replaced with either a new empty tube (Group 2, 4, and 5) or an odorant-containing tube (Group 3). The animals were allowed to freely interact with the Eppendorf tubes for additional 20 min. The same procedure was repeated on the last (testing) day when animals in Groups 2 and 3 received the odorant-containing tube, so that the odorants were novel to Group 2 and familiar to Group 3. On the testing day, Groups 4 and 5 were anesthetized and then presented with either odorants (Group 4, novel odor under anesthesia) or a new empty tube (Group 5, anesthesia control).
Table 1.
Experimental Groups
| Group | Notation and Color Code* | Description | Training | Testing | n |
|---|---|---|---|---|---|
| 1 | Naïve | Naïve control | - | - | 8 |
| 2 | Air/Odor | Novel odor, awake | Air | Odor | 7 |
| 3 | Odor/Odor | Familiar odor, awake | Odor | Odor | 7 |
| 4 | Air/Odor+Anes | Novel odor, anesthetized | Air | Odor+Anes | 8 |
| 5 | Air/Air+Anes | Anesthesia control | Air | Air+Anes | 8 |
|
| |||||
| 6 | Air/Air | Sham control | Air | Air | 6 |
| 7 | Air/Odor | Novel odor, awake | Air | Odor | 6 |
| 8 | Odor/Odor | Familiar odor, awake | Odor | Odor | 7 |
| 9 | Air+Anes/Odor | Novel odor, after anesthesia | Air+Anes | Odor | 6 |
| 10 | Odor+Anes/Odor | Anesthesia-paired odor | Odor+Anes | Odor | 6 |
|
| |||||
| 11 |
|
Novel/Familiar recognition | Odor1 | Odor1 & Odor2 | 14 |
| 12 |
|
Novel/Anes-paired recognition | Odor1+Anes | Odor1 & Odor2 | 24 |
Figure 1.
A schematic representation of the two experimental protocols used in the studies. In the repeated training protocol, animals were handled and acclimated to the experimental setup without experimental odorants for 30 min on Day 0 (dotted bar), followed by 20 min per day training with and without experimental odorants (hashed bars) for 6 days, and then tested on Day 7 with the experimental odorants (solid bar). In the single-session training protocol, animals were allowed to acclimate to the experimental setup and environment for 3 days without exposure to experimental odorants (dotted bars), followed by a single training session with or without experimental odorants and with or without general anesthesia (hashed bar), and tested 24-h later with or without the experimental odorants and with or without general anesthesia (solid bar). Various combinations of these conditions resulted in different experimental groups as defined in Table 1.
The second experimental set consisted of Groups 6–10 (31 rats), for which the single training protocol depicted in Figure 1 was used. This set of experiments was designed based on the results of the first set showing that odor habituation occurred in rats after a single 20-min exposure and that this habituation lasted for more than 24 h. Five groups (Group 6–10) of different training and testing conditions (see Table 1) were investigated for detailed histological quantification and comparison, and for the odor habituation behavioral tests. The animals were acclimated to the experimental environment 20 min per day for 3 days without exposing to the experimental odorants. They were then subjected to behavioral training or testing procedure (detailed in the “Behavioral Testing” section below) with either air (no odor) or a diluted novel odorant mixture, followed by a 20-min exposure to air or the same but undiluted odorant mixture to solidify the training effects.
The third experimental set, consisting of Groups 11 and 12 (38 rats), was designed for the forced-choice odor recognition test.19,20 One week after arriving at the housing facility, these rats underwent a 24-h familiarization period. Each rat was handled for approximately two minutes, after which four spherical wooden beads without experimental odor were introduced into their home cage. The four control beads remained in the home cage overnight so that the animals became familiar with them. For pre-exposure to a specific odorant, the animals were transferred to a clean cage for 20 min having odorant-containing Eppendorf tubes 24 h before the odor recognition tests.
General anesthesia
A surgical plane of general anesthesia was achieved by intraperitoneal injection of ketamine (Ketaset, Fort Dodge, IA; 100 mg/ml) and xylazine (Anased, Lloyd Laboratories, Shenandoah, IA; 100 mg/ml). A high dose of anesthetics was used to fully anesthetize the animals, with subsequent booster injections given based on pilot studies to ensure that animals were maintained at a surgical plane of anesthesia throughout the entire experimental period without any chance of waking up during the odor exposure. Rats were given an average dose of 154.2 mg/kg ketamine and 20.6 mg/kg xylazine over a ~30 min period. We chose injectable ketamine-xylazine anesthesia to avoid unwanted olfactory stimulation from pungent inhalational anesthetic agents. Widely used in veterinary medicine, ketamine-xylazine combination is among the safest and most easily manageable injectable anesthetics. For spontaneously breathing animals used in this study, ketamine anesthesia is considered one of the best choices for its ability to decouple upper airway muscle function from LOC, thereby stabilizing the airway potency during anesthesia.21,22 Anesthesia depth was reconfirmed by strong foot pinches and/or tail clamping before any odor exposures. Animals in the unanesthetized groups were handled in a similar way, including intraperitoneal injection of 0.3 ml saline to mimic the anesthetic injection procedure.
Odorants
In a binary (novel versus familiar) discrimination design, all experiments, except Groups 11 and 12, used the same odorant mixture. Because the odorant exposure is all or none, relatively high odorant concentration was used for training. The mixture was prepared and used in an identical fashion to minimize any variations in odor concentrations at the time of odor exposure among the groups of the same experimental set. Although APC and OFC show no spatial preference and bias to odorants,23–25 OB is spatially segregated.26 To achieve a ubiquitous bulbar activation, equal parts of six different odorants were combined. They were: isoamyl acetate, cineole, octanol, decanal, capronaldehyde, and R-(−)-carvone. Aliquots of 35-μl odorant mixture from the same stock were applied to a small cotton ball, which was then placed in the bottom of a capless 1.5-ml Eppendorf tube. Odor exposure experiments were conducted in a procedure room away from the housing room using a clean, standard rat cage inside a hard-ducted flow hood, which vented directly to the exterior of the building to prevent incidental odor exposure and to preserve odorant novelty. To introduce odorants, an odorant-containing Eppendorf tube was placed in the opposite corner of the cage from the rats and the animals were then allowed to freely interact with the tube. During the odor habituation behavioral testing, the Eppendorf tube was placed inside a cartridge underneath a center port in a specially designed behavioral testing chamber, as described below under the subheading “Behavioral testing.” For anesthetized animals, the distance between the tube and the nose was manually varied to mimic the awake situation with animal movements. During odorant exposure, the cage or the testing chamber was closed with a transparent cover to retain the odorant within the cage. After exposure, all animals were transferred to a clean cage with a wire grid cover and vented for 15 min within the flow hood to remove any residual odor. All anesthetized animals recovered from anesthesia in a clean cage.
For the odor recognition test (Groups 11 and 12), wooden beads of 2.54-cm in diameter with a 5.6-mm hole bore through the middle (Woodworks Ltd., Haltom City, TX) were individually coded and used. The scented beads were produced by being incubated overnight in a sealed 50-ml Falcon tube (BD Biosciences, Bedford, MA) with 100 μl of a pure odorant (diluted 1:10 in mineral oil) injected into a cotton reservoir in the bottom. Four different odorants (Sigma-Aldrich, St. Louis, MO) were separately used: isoamyl acetate, decanal, R-(−)-carvone, and ethyl butyrate. A plastic insert prevented beads from directly contacting the scented oil. Any beads that came into direct contact with the oil were discarded, so that the scent intensity of a given odor was made as close as possible among all beads. Pilot studies revealed that strongly scented beads were avoided or investigated from a distance. Our method produced beads with an odor intensity that was actively and directly investigated by rats. Preferences for the four odorants were tested in pilot studies using different pairing combinations. Isoamyl acetate and decanal were found to be almost equally preferred by rats, judging by the time they spent exploring the beads.
Behavioral testing
Odor Habituation Test
Our odor-habituation test was a modification of the odorant enrichment and discrimination protocol.11 A custom-built 43.2 cm × 43.2 cm black plexiglass floor was constructed to fit inside an open field activity monitoring chamber of the same dimensions (model ENV-515, Med Associates Inc., St Albans, VT). A removable black plexiglass cartridge, 5.08 cm in diameter, was positioned in the center of the floor, and the circumference of the cartridge was defined as the “odorant zone.” A 7.9-mm diameter hole was bored in the center of the cartridge as the only odor inlet into the chamber, allowing the animals indirect access to the odorant during the behavioral tests. The cartridge lay flush to the floor such that the top of the cartridge blended smoothly into the floor and the animals could only smell through the central hole without noticing the cartridge. The odorant source, hidden from the view of animals, was placed inside the cartridge underneath the floor. The black floor and a clear plexiglass cover allowed low light video recording from above. Animals were habituated to the testing environment by being placed in the chamber for 20 min every day for three days before the training and testing. On the training or testing day, after a 15-min habituation period, an empty or odorant-containing tube was placed underneath the central odor port. The rats were allowed to freely interact with the environment for additional 5 min and their behaviors for the entire period were recorded. Familiarity with the odorant was determined by the duration of smelling within the odor zone. Pilot studies found that the rats typically avoided the central and most open area of the chamber when no odor was present. This avoidance is important as it deters any stationary positioning near the odorant zone. Video was acquired and analyzed using Limelight automated tracking software (v3.0; Actimetrics). Nose, body and tail locations were tracked, and time of the nose within the odor zone was quantified for the first 10, 30, and 100 s from initial approach. Parallel to automated analyses, videos were also scored manually for smelling behaviors by two evaluators who were blind of the experimental groups.
Odor Recognition Test
For the forced-choice odor recognition testing, we adapted a previously described protocol19,20 for use with arbitrary odorants. Animals were allowed to freely choose to smell four wooden beads; two beads were scented and the other two were unscented. After a 24-h bead familiarization period, the control beads were removed from the cage. The testing session was conducted in the animals’ home cages placed inside a hard-ducted hood that vented directly to the exterior of the building. Each animal was allowed to habituate to the environment for 10 minutes. After habituation, the 4 familiarized control beads were returned to the cage in a manner similar to the testing procedure. The rat was allowed to freely interact with the control beads for 5 minutes, after which the beads were removed and the animal left alone for additional 5 minutes. Thereafter, the animal was gently restricted to one end of the cage, and two of the control beads were placed back in the cage along with two scented beads. The four beads were placed in a row (Odorant 1, Control, Control, Odorant 2) on the far side of the cage approximately 2.54 cm from the cage wall. The two scented beads were placed apart to avoid ambiguity in the odor recognition. The rat was then released and allowed to freely interact with the four beads for 5 minutes. A video camera was positioned outside the clear plastic cage to record the animal’s interaction with each bead. The left-right positions of the two scented beads were reversed between trials. One of the two odor choices was pre-exposed 24 h earlier under either awake or anesthetized condition. Adding two unscented beads, instead of using only two scented beads, is known to increase the sensitivity and reliability of the test.19
Two evaluators blinded of the experimental groups assessed the videos acquired during the behavioral task. The time spent smelling the beads was assessed for the first 30 and 60 seconds after the animal approached any bead. Latency to approach was also assessed and capped at 30 seconds. Smelling behaviors (bead-directed sniffing, rearing) within 2.54 cm of the bead were scored. Time spent chewing, rolling, or playing with beads did not count towards smelling duration. Smelling duration over all beads was summed and the time spent smelling each bead was represented as a percentage of the total smelling time to normalize for innately shy or inquisitive animals.
Immunohistochemistry
All histology experiments began two hours after the final odor or air exposure on the testing day. IEG protein production usually peaks at 2 h. Animals were sacrificed and perfused with a solution of 4% paraformaldehyde in phosphate buffered saline (PBS; pH 7.0). Brains were post-fixed overnight and cryoprotected in a 30% sucrose/PBS solution with 0.01% sodium azide as a preservative. 50 μm (Groups 1–5) and 30 μm (Groups 6–10) sections were generated using a sliding microtome (Leica Biosystems, Buffalo Grove, IL) equipped with a freezing stage (Physitemp Instruments, Inc., Clifton, NJ). Rabbit polyclonal antibodies against c-Fos (1:5000, Calbiochem, Billerica, MA) and Egr1 (Cell Signaling, Danvers, MA; 1:10,000) diluted in donkey blocking buffer (DBB; PBS with 10% donkey serum and 0.2% Triton X-100) were separately incubated with free-floating sections overnight at 4°C. Sections for 3,3′-Diaminobenzidine (Sigma-Aldrich) labeling were washed with PBS and then incubated with a biotinylated goat secondary antibody (1:1000 in DBB; Dako, Carpinteria, CA) for 2 h at room temperature, followed by an avidin-peroxidase wash (1:1000 in DBB; Sigma-Aldrich) for 1h. Enzymatic reaction with 3,3′-Diaminobenzidine was performed until sections reached optimal darkness (1–3 min). Sections for fluorescent microscopy were washed with PBS after treatment with primary antibodies and incubated with Alexa Fluor 594 and/or 488-conjugated secondary antibodies (Invitrogen, Carlsbad, CA; 1:500) in the dark. Sections were co-stained with the nuclear marker DAPI (4′,6-diamidino-2-phenylindole; Sigma-Aldrich), washed and mounted onto slides using fluoromount-G mounting medium (Southern Biotech, Birmingham, AL).
Quantification
Two different quantification methods were used. In the OB where odorant-induced neural activation is spatially organized, counting of total number of IEG positive cells in the entire bulbar substructures is required. Stereology, which uses statistical approach to provide three-dimensional quantification based on cell counting in equally spaced two-dimensional optical dissectors, is the most suitable method.27–29 In APC and OFC where spatial segregation does not occur, cell density measurement is the most straightforward and reliable approach. Staining was imaged using an Olympus IX-81 microscope equipped with a Prior motorized stage (Prior Scientific, Rockland, MA), a monochrome Hamamatsu ORCA-ER camera (Hamamatsu, Bridgewater, NJ) and a color Retiga 2000R (QImaging, Surrey, BC, Canada) peltier-cooled charge-coupled-device camera. Images were acquired and analyzed using Image-Pro Plus software (Media Cybernetics, Bethesda, MD) with automatic stage control capability. Macros as modules for Image-Pro software for optical fractionator and optical dissectors30 in stereological analyses were obtained from Dr. Michael King’s laboratory at University of Florida College of Medicine, Gainesville, and were used to quantify IEG positive cells in each region.30 OB sections at +7.9 mm and +9.1 mm from bregma and every sixth section in the APC and lateral OFC (from bregma +3.2mm to +3.8mm)31,32 were quantified and used to estimate the positive cell number in the OB, APC, and OFC. The cell density quantification method in APC and OFC was compared to, and cross-validated by, stereology using the relative between-group ratios from the corresponding groups. For fluorescence staining sections, images were acquired under identical settings, screened by a fixed size, and uniformly quantified using the threshold segmentation method.
Statistical analysis
The GraphPad (GraphPad Software, Inc., La Jolla, CA) and SPSS (IBM Corporation, Armonk, NY) programs were used for statistical analysis. Although our experimental design consisted of two independent variables (i.e., brain regions and odorant types), a prior known differences in cell number, cell density, and physical sizes of the brain regions and sub-regions excluded the use of two-way ANOVA. Therefore, one-way ANOVA was used to determine a significant overall difference, followed by least significant difference or Tukey post hoc analysis for inter-group comparison. To analyze the behavioral data with the repetitive training protocol, repeated measures ANOVA with Bonferroni post hoc test was used. To estimate the number of animals needed for the first two sets of experiments (Groups 1–5 and Groups 6–10), the power analysis was performed for a p of 0.05 and a power of 0.8 based on a typical c-Fos within-group counting variation of ~17% in the pilot studies in order to detect at least a 30% difference between groups. For the forced-choice odorant recognition test (Groups 11 and 12), we focused on detecting a different between a true novel odorant and an anesthesia-paired odorant in a two-tailed t test design for an effect size of 0.85 and a power of 0.8. A p value of <0.05 was considered statistically significant.
RESULTS
Novelty of olfactory sensory input can be quantified by c-Fos immunoreactivity
To establish the time dependent odorant responses and to determine if IEG immunoreactivity in the primary olfactory reception region can differentiate novel from familiar odorants, we quantified the total number of c-Fos+ and Egr1+ cells in the entire granule cell layer (GCL) and mitral layer (ML) of OB using stereology. As shown in Figure 2, the animals having repetitive odorant exposures 20 min per day for seven days (familiar odorant group, n = 7) produced significantly lower c-Fos immunoreactivity in the OB than those that were handled identically but only received the experimental odorants on the testing day (novel odorant group, n = 7). Both groups showed elevated c-Fos and Egr1 immunoreactivities in the OB compared to the naïve controls (n = 8). The increase in Egr1 immunoreactivity, however, did not differentiate whether the odorants were novel or familiar, suggesting that the olfactory sensory inputs were the same in the novel and familiar odorant groups. This is consistent with previous findings that the egr1 gene activation reflects synaptic events,33–36 with the Egr1 expression profile correlating with neural activity measured by 2-deoxyglucose (2-DG) labeling and optical imaging.12 In comparison, c-Fos immunoreactivity does not simply echo neural activities, but relies on information integration and consolidation. The contrast between Egr1 and c-Fos supports the notion that the difference seen in the c-Fos immunoreactivity is indeed specific to odor novelty. One-way ANOVA with respect to odor exposure (novel, familiar, or naïve) showed a significant difference among groups for both c-Fos and Egr1 immunoreactivities. The least significant difference post hoc comparison showed that c-Fos immunoreactivity levels were significantly different between novel and familiar odorant groups in GCL and ML (p < 0.001 in both regions), but the difference between familiar odorant group and the naïve control were not statistically significant. Egr1 immunoreactivity showed no difference between novel and familiar odorant groups, but both groups were significantly different from the naïve control (p < 0.05 in GCL and p < 0.01 in ML). Thus, exposures to experimental odorants caused an Egr1 immunoreactivity increase and odor-novelty-specific c-Fos immunoreactivity increase in the primary olfactory reception regions. Egr1 reflected olfactory sensory input levels without discriminating animals’ familiarity with the odorants, whereas c-Fos immunoreactivity differentiated novel from familiar odorant responses.
Figure 2.

Registration of olfactory sensory information by the immediate early gene products, c-Fos and Egr1, in the olfactory bulb (OB). Representative diaminobenzidine staining of c-Fos (A, B) and Egr1 (C) in the granule cell layer (GCL, lightly shaded area in the top row) and mitral layer (ML, the dark band of stained cells immediately to the right of the dashed lines) of the OB of rats exposed to novel (Air/Odor) or familiar odorants (Odor/Odor), or in the naïve control (Group 1). Row B shows the zoom-in views of rectangle areas in row A. Positively stained cells appear as dark brown. Arrowheads in the zoom-in views mark only a few examples of c-Fos positive cells. The green squares, in which cell counting was performed, were generated by optical dissectors for stereology. Comparison of immunoreactivity of c-Fos (D) and Egr1 (E) in the GCL and ML of the OB as quantified by the stereological analysis. White bars, naïve control (Group 1); light blue bars, novel odorants (Air/Odor, Group 2); and light green bars, familiar odorants (Odor/Odor, Group 3). Data were taken from the first experimental set using the repeated training protocol depicted in Figure 1. ‘*’ and ‘***’ indicate significant differences (p < 0.05 and p < 0.001, respectively) between groups (ANOVA with the least significant difference post hoc test). Egr1, early growth response protein 1.
High olfactory cortices continue to register olfactory inputs under general anesthesia
We next investigated the IEG registration of olfactory stimulations under a surgical plane of general anesthesia. The anesthetic dose used to achieve and maintain general anesthesia was not statistically different among anesthetized groups. We focused our analyses on the consolidation of sensory inputs in APC and OFC because identification and differentiation of odorants in these high olfactory cortical regions underlie the cellular basis of olfactory perception.7 It is generally believed that perception is the important sensory attribute that is removable by general anesthesia. We determined if rats in a fully anesthetized state could continue to register novel odorants by robust elevation of c-Fos immunoreactivity as in conscious rats. Figures 3A and 3B depict representative c-Fos and Egr1 immunoreactivity in the APC and OFC, respectively, in response to novel odorants in the awake (Group 2, Air/Odor) and fully anesthetized (Group 4, Air/Odor+Anes) states, as compared to naïve control without experimental odorants (Group 1) and anesthesia control (Group 5). It is clearly evident that novel odorants, when presented to a fully anesthetized animal, continue to elicit robust c-Fos immunoreactivity in APC (Figure 3C) and OFC (Figure 3D), even though the reception of olfactory sensory inputs to these high cortices, as reflected in Egr1 immunoreactivity (Figures 3E and 3F), has been attenuated to a level comparable to that of anesthesia alone without the experimental odor stimulations.
Figure 3.

Registration of olfactory sensory processing by c-Fos immunoreactivity under a surgical plane of general anesthesia. Representative images of c-Fos and Egr1 immunolabeling in the anterior piriform cortex (A) and the orbitofrontal cortex (B) of rats from four of the 12 experimental groups as defined and color-coded in Table 1. Positively stained cells appear as dark brown; not all but only a few examples of c-Fos positive cells are marked by arrowheads. The bar graphs show quantifications of c-Fos (C, D, G, H) and Egr1 (E, F) immunoreactivity by stereology (C, D, E, F) and by c-Fos positive cell density (G, H) measurements in the anterior piriform cortex (C, E, G) and the orbitofrontal cortex (D, F, H). The same notations and color code as in Table 1 are used. ‘*’, ‘**’, and ‘***’ indicate statistic significance p < 0.05, p < 0.01, and p < 0.001, respectively, based on the least significant difference post hoc comparisons. See Table 2 for other statistics. Scale bar = 50 μm. Anes, anesthesia; Egr1, early growth response protein 1.
Detailed quantification of c-Fos and Egr1 immunoreactivities in APC and OFC was also performed for Groups 6–10. Because APC is known to have no spatial preference in response to odorants23,24 and usually receives olfactory inputs from the entire OB without bias,25 the sensory processing in APC, and subsequently in OFC, is best quantified by c-Fos+ cell densities (Figures 3G and 3H) without subdividing these high-order cortical regions. We validated the cell density quantification method by performing stereology in a subset of animals to confirm that the two quantification methods led to identical conclusions. Key comparisons and a complete list of statistics of c-Fos immunoreactivity in APC and OFC from post hoc multiple comparisons are given in Table 2. The comparison between Group 2 (novel odorants, awake, Air/Odor) and Group 4 (novel odorants under general anesthesia, Air/Odor+Anes) shows that the c-Fos immunoreactivity elicited by novel odorants is not significantly different in APC between fully awake and fully anesthetized animals. In both APC and OFC, the novel odorant-induced increase in c-Fos immunoreactivity under anesthesia (Group 4) is significantly higher than that elicited by the anesthesia alone (no experimental odorants) in Group 5 (p = 0.005 in APC and p = 0.008 in OFC). Moreover, there is no significant difference in c-Fos immunoreactivity between Group 7 (novel odorants awake, Air/Odor) and Group 9 (novel odorants awake, one day after anesthesia, Air+Anes/Odor), suggesting that pre-treating animals with ketamine-xylazine anesthesia 24 h before testing did not alter c-Fos response to novel odorants. Thus, despite the attenuated sensory input as reflected in Egr1 immunoreactivity (Figures 3E and 3F), c-Fos in APC and OFC can still “record” novel experimental odorants under a surgical plane of ketamine-xylazine general anesthesia with a robustly increased c-Fos immunoreactivity.
Table 2.
Comparisons of c-Fos Immunoreactivity between Key Groups*
| Groups Compared | Anterior Piriform Cortex | Orbitofrontal Cortex | |||
|---|---|---|---|---|---|
| c-Fos | Naïve | vs. | Air/Odor+Anes | .000 | .000 |
| c-Fos | Naïve | vs. | Air/Air+Anes | .023 | .291 |
| c-Fos | Air/Odor | vs. | Naïve | .000 | .000 |
| c-Fos | Air/Odor | vs. | Air/Odor+Anes | .275 | .000 |
| c-Fos | Air/Odor | vs. | Air/Air+Anes | .000 | .000 |
| c-Fos | Air/Air+Anes | vs. | Air/Odor+Anes | .005 | .008 |
|
| |||||
| c-Fos | Air/Air | vs. | Air/Odor | .001 | .014 |
| c-Fos | Air/Air | vs. | Odor/Odor | .410 | .214 |
| c-Fos | Air/Air | vs. | Air+Anes/Odor | .028 | .151 |
| c-Fos | Air/Air | vs. | Odor+Anes/Odor | .771 | .085 |
| c-Fos | Odor+Anes/Odor | vs. | Air/Odor | .000 | .000 |
| c-Fos | Odor+Anes/Odor | vs. | Air+Anes/Odor | .014 | .003 |
| c-Fos | Odor+Anes/Odor | vs. | Odor/Odor | .264 | .562 |
| c-Fos | Odor/Odor | vs. | Air/Odor | .004 | .000 |
| c-Fos | Odor/Odor | vs. | Air+Anes/Odor | .125 | .009 |
| c-Fos | Air+Anes/Odor | vs. | Air/Odor | .148 | .261 |
Selected statistics from the least significant difference post hoc multiple comparisons among groups within each experimental set as defined and color-coded in Table 1. “Anes” = Anesthesia.
The ability to differentiate novel from familiar odorants by c-Fos is preserved in higher cortices under general anesthesia
To further differentiate whether the c-Fos registration of odorants under general anesthesia represents certain forms of high-order processing such as recognition of odor novelty, we further investigated (a) whether the ability to differentiate the binary condition between novel and familiar odorants by c-Fos, as shown in Figure 2 for the OB, was preserved in the higher cortices, and (b) whether re-exposing animals to “novel” odorants, which had been previously exposed only under a surgical plane of general anesthesia, would fail to elevate c-Fos immunoreactivity as if odorants were of “familiar” type.
As revealed by comparison between Groups 7 (Air/Odor) and 8 (Odor/Odor) in Figures 3G and 3H and Table 2, c-Fos immunoreactivity is significantly different between novel and familiar odorant stimulation in both APC (p = 0.004) and OFC (p < 0.001). Group 8 suggests that a single 20-min training with novel odorants was sufficient to alter the subsequent c-Fos immunoreactivity in conscious rats, as these rats did not respond to the second exposure with elevated c-Fos activation. In fact, c-Fos immunoreactivity in Group 8 is indistinguishable from the sham control (Group 6). Behavioral testing discussed below supported the conclusion that after a 20-min training with the experimental odorants, rats became behaviorally familiar with the odorants and this behavioral familiarity lasted for more than 24 h. Interestingly, Group 9 (Air+Anes/Odor), which received novel odorant stimulations 24 h after general anesthesia, showed significantly higher c-Fos immunoreactivity than Group 8 in OFC, suggesting that the ability of c-Fos immunoreactivity to differentiate characteristic response to novel and familiar odorants is preserved in this high cortical region. Moreover, the distinction is also maintained after a surgical level of general anesthesia. We therefore conclude that c-Fos immunoreactivity in the secondary and tertiary olfactory processing cortices can continue to differentiate and register novel odorant stimulations under general anesthesia.
To further substantiate this conclusion, one of the most stringent tests is to compare Group 10 (Odor+Anes/Odor) with other groups, particularly with Group 8 (Odor/Odor) and Group 9 (Air+Anes/Odor). In Group 10, animals were exposed to novel odorants for 20 min only when they were fully anesthetized. They were then tested 24 h later with the same odorants while awake. If the same odorants can no longer produce elevated c-Fos response after the second exposure because the animals had “smelled” the odorants under general anesthesia, then the results would be more consistent with the “familiar-type” c-Fos immunoreactivity as seen in Group 8.
The data in Figures 3C, 3D, 3G, and 3H and in Table 2 reflect a pattern that is consistent with the conclusion that novel odorants presented to rats under ketamine-xylazine general anesthesia were successfully registered in the higher olfactory cortices and that the same odorants presented to the same rats again while they were awake elicited a reduced c-Fos immunoreactivity similar to that by familiar odorants. Could the reduction in c-Fos immunoreactivity after the second exposure in Group 10 result from any unforeseen effects from anesthesia? The answer is no, because the similarly high level of c-Fos immunoreactivity in Group 7 and Group 9 confirmed the absence of any delayed effects from anesthesia procedures alone on histological registration of odor novelty. Hence, the same extent of c-Fos immunoreactivity reduction in Group 10 as in Group 8 can only be interpreted as an indication that a single 20-min exposure to novel odorants rendered the odorants unable to re-elicit c-Fos elevation, even when the first exposure was given under a surgical plane of general anesthesia (also, compare the c-Fos immunoreactivity changes from Group 7 to Group 8 in the absence of anesthesia, and from Group 9 to Group 10 in the presence of anesthesia). The c-Fos immunoreactivity in Group 10 was essentially the same as that elicited by air as in Group 6 or by the familiar odorants as in Group 8. These results confirmed that the odorant novelty was positively recognized and histologically registered in the high olfactory cortices under a surgical plane of general anesthesia. The data from Group 10 also provide the direct experimental evidence of “unconscious cellular learning,” i.e., novel stimuli can alter the subsequent representation of the same stimuli without the stimuli first entering a conscious state.
Majority of c-Fos+ cells elicited by novel odorants are neurons
We also determined the cell types of the c-Fos+ cells. We performed fluorescence co-localization with double staining against c-Fos and NeuN, a marker for mature neurons. Brain sections from animals in Groups 2 (novel odorants, awake), 4 (novel odorants, anesthetized), and 5 (anesthesia alone without experimental odorants) were stained. Representative images are shown in Figure 4. Quantitative analyses revealed that the vast majority of c-Fos+ cells are also NeuN positive. In APC, 98.7%, 96.3%, and 91.0% of c-Fos+ cells are neurons in Groups 2, 4, and 5, respectively, whereas in OFC, the percentages are 97.5%, 97.3%, and 87.6% for the three groups, respectively. It is important to point out that there is essentially no difference in cell type distribution of c-Fos+ cells elicited by novel odorants, whether the animals were fully anesthetized or not.
Figure 4.

Representative fluorescent micrographs of c-Fos (red) and NeuN (green) double-staining in (A) anterior piriform cortex (APC) and (B) orbitofrontal cortex (OFC). Corresponding NeuN images are shown in (C) and (D), respectively, for better identification of neurons. Scale bar = 50 μm. Anes, anesthesia; NeuN, neuronal nuclear protein.
We further addressed whether different sub-types of neurons were specially involved in odorant-induced c-Fos activation. Specifically, we determined if a different ratio of excitatory and inhibitory neurons was involved in the c-Fos immunoreactivity in the awake and anesthetized states. We double-labeled brain sections from Groups 2, 4, and 5 with antibodies against c-Fos and GAD67, a marker for inhibitory neurons. Across all groups, only 3.2% ± 0.3% of c-Fos+ cells were also positive for GAD67 in the APC. A slightly greater percentage (7.6% ± 0.5%) of c-Fos+/GAD67+ cells were found in the OFC. Again, we found no difference between Groups 2 (novel odorants, awake) and 4 (novel odorants, anesthetized). Interestingly, the anesthesia control group (Group 5) had a significantly higher (p < 0.05) percentage of c-Fos+/GAD67+ cells in the APC than the novel odorant group (Group 7), but significantly fewer c-Fos+/GAD67+ double-labeled cells in the OFC, suggesting that odorants and anesthesia might activate different populations of cells.
Olfactory events registered under general anesthesia cannot be behaviorally recalled
The registration of olfactory input as reflected in the novelty-specific c-Fos immunoreactivity clearly indicates that sensory processing at the cellular level in the higher cortices is not completely ablated by a surgical level of ketamine-xylazine general anesthesia. This raises an important question as to whether the clearly definable registration of information at the cellular level can be recalled by the animals. We carried out behavioral testing with the animals (see Table 1).
An intriguing discovery of this study is the possible dissociation between histological and behavioral manifestations of the same sensory processes. Despite unambiguous histological registration and differentiation of novel from familiar olfactory events under general anesthesia, the behavioral testing did not fully mirror the c-Fos results. Representative traces of snout paths in the testing chamber and the average time that animals spent smelling the odorants during the first 30 s after approaching the odorants are depicted in Figures 5A and 5B. With the repeated training protocol (Groups 2 and 3), we found that rats became behaviorally habituated to the experimental odorants after only a single exposure and showed little interest in exploring the same odorants from the second exposure onward (Figure 5C). Likewise, our single-session training protocol, which eliminates potential side effects from repeated anesthetic exposures in the anesthetized groups, provides the same contrast to differentiate behavioral phenotypes in the memory recall of various olfactory events.
Figure 5.

Odor habituation tests to discriminate behavioral phenotypes towards novel and familiar odorants. Behavioral familiarity with the odorants was assessed by duration of odorant investigation time. (A) Representative images of rat body traces from Groups 6 through 10 during the 15-min acclimation phase without odorants in the central cartridge. (B) Snout traces generated within the first 100 s after initial approach to the odorant zone during the test. (C) Time course of seven-day repeated behavioral tests of odor habituation, attenuation and discrimination showing that odorant habituation occurs after a single exposure. Plotted are odorant investigation times of animals in Groups 2 and 3 over the repeated training and test days. ‘*’ indicates significant difference (p < 0.05, repeated measures ANOVA with Bonferroni post hoc test). (D) Time course of smelling responses on the acclimation, single session training, and testing trials. Smelling duration was determined by the time during which snout was within the odorant zone scored automatically for the first 30 s after initial approach to the zone. (E) Data on the testing day in (D) are summarized in bar graph representation. All symbols and colors are identical to those in Figures 3G and 3H and Table 1 for easy comparison. Data were derived from the same animals as in Figures 3G and 3H. ‘*’ indicates a significant difference (p < 0.01) relative to a novel odorant exposure without anesthesia (blue bar); ‘†’ indicates a significant effect (p < 0.01) relative to a novel odorant exposure given 24 h after anesthesia (purple bar) or to the second exposure to the anesthesia-paired odorants (gray bar). Notice that the gray bar is not different from the blue and purple bars, in sharp contrast to the histology results in Figures 3G and 3H. ANES, anesthesia.
Quantitative analyses of the behavioral data (Figures 5D and 5E) showed a significant difference among Groups 6–10 (one-way ANOVA, p < 0.001). Tukey post-hoc test revealed two distinct behavioral phenotypes. A second exposure to the same odorants (Group 8) produced an attenuated behavioral response relative to a single exposure (significant difference in odor exploration time between Groups 7 and 8; p < 0.01). Importantly, rats in Group 10 (exposed to novel odorants under anesthesia and then tested 24-h later with the same odorants while awake) spent almost the same amount of time exploring the odorants during the test as the rats that were exposed to the odorants for the first time, indicating that these rats could not recall the first exposure under general anesthesia. No significant difference was found between Group 7 (Air/Odor) and Group 9 (Air+Anes/Odor), between Group 7 and Group 10 (Odor+Anes/Odor), and between Group 9 and Group 10, but these three groups were significantly different (p < 0.01) from sham (Group 6) and familiar odorant group (Group 8). In other words, even though the animals in Group 10 exhibited a characteristic of c-Fos immunoreactivity for familiar odorants in the second exposure, they explored the odorants significantly longer than the animals in Group 8, as if the odorants were completely new to them during the second exposure (Figure 5E). When the behavioral data were scored manually for smelling behaviors instead of automatically tracing the nose in the odorant zone, we arrived at the same conclusion no matter whether the data were analyzed for the first 10 s or first 100 s after animals approaching the odorant zone.
In addition to habituation test to differentiate novel from familiar odorants, we also used the established forced-choice odor recognition procedure to test if animals could recognize an arbitrary but pure odorant among four possible choices (Groups 11 and 12). Among the four choices were wooden beads with a truly novel odor and a previously experienced odor (experienced either awake or under general anesthesia 24 h earlier) along with two additional wooden beads of identical appearance but without experimental odors. In a counter-balanced design, we analyzed if subtle preference existed for the truly novel odorant over the anesthesia-paired odorant. In pilot experiments, we first determined animals’ preferences for different odorants and found that isoamyl acetate (banana scent) and decanal (citrus scent) were similarly preferred by rats.
As shown in Figure 6, animals spent significantly more time smelling a novel odorant than a familiar odorant (Figure 6A, p < 0.001) and the other two unscented choices (p < 0.001). Consistent with the habituation behavioral test, these results indicate that the odorant discrimination task is a robust measure of odorant recognition even with two unrelated odorants, and that rats display clear familiarity with the odorant experienced 24 h earlier. However, when the second odor choice was a previously anesthesia-paired odorant, the animals showed no significant difference in choosing the truly novel odorant over the anesthesia-paired odorant in terms of total smelling time (Figure 6B, p = 0.31, although there is a slight trend in favor of the true novel odorant). This suggests that the animals were unable to recall the previous experience with the anesthesia-paired odorant, again consistent with the behavioral results from the habituation test described above. Interestingly, when the latency of first approach to any choice was analyzed in the forced-choice odor recognition test, we found that the latency was significantly longer (p < 0.0001) when anesthesia-paired odorant was among the four choices (Figure 6C). This result provided indirect evidence implicating certain forms of “hidden” memory for the anesthesia-paired odorant, causing the animals to hesitate on their approach.
Figure 6.
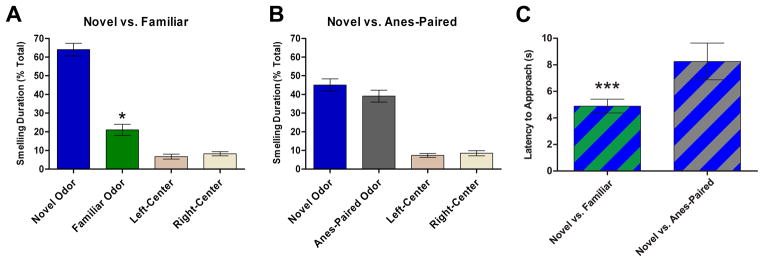
Forced-choice odor recognition tests. After 24-h familiarization with four odorless wooden beads in their home cages, animals were presented with four beads in a row, of which two outer beads were scented and two center beads were odorless. (A) Smelling duration on each bead is scored as a percentage of total smelling time when the two scented beads were a truly novel odor and a familiar odor exposed 24-h earlier while animals were awake. (B) Smelling duration on each bead is scored as a percentage of total smelling time when the two scented beads were a truly novel odor and an anesthesia-paired (Anes-Paired) odor exposed 24-h earlier while the animals were fully anesthetized. (C) Latency of approach to beads is compared between forced choice for (A) and forced choice for (B). The odorants used as novel, familiar, or anesthesia-paired and their positions as outer-left or outer-right were counter-balanced. Data from the third experimental set (Groups 11 and 12). “*” and “***” indicate significant difference p < 0.05 and p < 0.0001, respectively.
DISCUSSION
Sensory processing under general anesthesia
We showed that behavioral LOC and cellular registration of sensory information are distinct and separable processes. An important discovery is that the neural processing involved in recognizing sensory novelty continues to occur in a drug-induced unconscious brain. Equally intriguing is the lack of behavioral recall of this process, except for the approach latency in the odorant recognition test. Although recordings in various sensory cortices have documented that olfactory, visual, somatosensory, auditory, or gustatory inputs are still received and odor habituation and discrimination are measurable in individual neurons under general anesthesia,37–40 recognizing and discriminating the nature of sensory inputs and further consolidating input information under general anesthesia are not expected and have not previously been demonstrated experimentally in the high-order olfactory system. This study is unique in that we created an odorant-learning condition under a surgical plane of general anesthesia and subsequently evaluated and compared cellular registration and behavioral recall in a wakeful state, as demonstrated in Groups 9 and 10. Our observation that ketamine-xylazine anesthesia can reduce sensory input while still allowing cellular registration in the higher olfactory cortices suggests a previously unknown phenomenon that can be termed “anosmiolfaction,” similar to blindsight in the visual system. The latter refers to well-documented observations that blind subjects with severely damaged primary visual cortex can still perceive unseen stimuli involving higher cortical functions.41
The mechanisms of general anesthesia at the systems level remain elusive. Blocking thalamocortical connectivity42 or disintegrating cortical feed-forward and feedback coupling43,44 has been suggested. C-fos activation in response to various well-controlled experimental conditions in this study suggests that sensory inputs are both received and possibly processed under anesthesia, particularly considering that novel odorants experienced under ketamine-xylazine general anesthesia could later only trigger c-Fos responses similar to that by familiar stimuli. Thus, at least for the olfactory system where thalamus involvement is limited and indirect, our results suggest that ketamine-xylazine general anesthesia disrupts the integrity of sensory information processing without necessarily blocking information flow to the higher cortices, thus favoring information disintegration as a systems mechanism of general anesthesia.
Sensory reception versus perception and consolidation
Differentiating reception and perception under general anesthesia is important. Dependence of egr1 and c-fos activation on neural activities has long been established.45 Unlike Egr1, which responds to the presence of sensory input, c-Fos immunoreactivity depends more on input novelty than intensity, suggesting a top-down regulation. Indeed, patterns of 2-DG labeling and c-Fos immunoreactivity often do not match.46 Centrifugal afferent lesions can significantly reduce bulbar c-Fos immunoreactivity without affecting 2-DG response.47 Likewise, unilateral locus ceruleus lesions produce a widespread cortical c-Fos reduction ipsilateral to the lesion without changes in electroencephalography.48 However, if top-down input is maintained, even with unilateral naris closure to block external input, odor stimulation in rodents can still produce robust bulbar c-Fos immunoreactivity both contralateral and ipsilateral to the closure.49 Thus, the ability of c-Fos immunoreactivity in APC and OFC to detect and discriminate the presence and novelty of experimental odorants under ketamine-xylazine general anesthesia, even when basic sensory input was suppressed as reflected by the decreased Egr1 immunoreactivity (Figures 3E and 3F), indicates a process beyond that of simple sensory reception.
Does the c-Fos registration of olfactory events under general anesthesia represent a form of memory consolidation? While many studies have already established the causal relationship between c-Fos activation and memory formation,50,51 the disagreement between c-Fos immunoreactivity and behavioral data (compare the gray bars in Figures 3G and 3H to that in Figure 5E) requires further explanation. Several possibilities can be considered: (1) The c-Fos immunoreactivity in our study might merely mark the onset, not the completion, of memory consolidation. In this case, olfactory information is preserved but anesthesia causes anterograde amnesia, leading to animals’ inability to recall an incomplete memory. This scenario is rather unlikely because if consolidation were incomplete, the second exposure to the same odor in the conscious state would conceivably lead to a different level of c-Fos immunoreactivity for more complete consolidation. (2) Long-term olfactory memories might not store in the OB, APC, or OFC. Consequently, the lack of correlation between odorant-induced c-Fos immunoreactivity in these territories and the behavioral recall upon reintroducing the anesthesia-paired odorants may simply indicate a breakdown in information processing distal to OFC. (3) The histological and behavioral discrepancy may also suggest that “pure” olfactory memory cannot be formed, namely, odorants cannot be remembered without being co-registered and associated with other cognitive properties (such as taste, texture, etc.) processed by other brain regions. The hippocampus, basolateral amygdala, and hypothalamus are heavily involved in memory and learning, including that related to olfactory stimuli.52 Olfactory memory might require certain attachment to other entities to be recalled. Using odor habituation as a measure, it was demonstrated53 that if an odor was paired with stimulation of locus ceruleus and noradrenaline release, the stimulation-paired odor could be recognized 24 h later even when pairing occurred under anesthesia. Without locus ceruleus stimulation, no behavioral recall was observed, consistent with our results. Thus, if anesthesia interferes with integration or synchronization of information processing among different brain regions or disrupts association between olfactory and other processing centers, behavioral recall may be impaired. (4) The registration of sensory information under general anesthesia might be fragmented and incoherent. The threshold might be lower to trigger changes in c-Fos immunoreactivity than to achieve a complete recall. This scenario is consistent with a recent chronic two-photon calcium imaging study,54 which showed that the olfactory sensory representations in the olfactory bulb are differently influenced by the wakeful and anesthetized states. Specifically, granule cells, a major class of GABAergic inhibitory interneurons, had significantly lower spontaneous and odor-evoked cell activities under urethane anesthesia. This is counter-intuitive because urethane is known to strongly potentiate GABAergic current.55 As a consequence of lower inhibitory input, mitral cell activities—both in cell number and in intensity—were found to be more pronounced in anesthetized state than in wakeful state. The increase in mitral cell density of odor representations, as the authors asserted, broadens “the odor tuning of mitral cells” and reduces the efficiency of odor coding.54 Thus, it is possible that anesthetized state is a state with too much information for the encoded information to be meaningful. In other words, the information consolidation under general anesthesia, as reflected in c-Fos immunoreactivity, might produce a “hidden” or “broadened” memory that cannot be detected by the current odor habituation test but nevertheless implicated by the latency in the forced-choice odor recognition test. More refined protocols for behavioral testing to ascertain memory consolidation under general anesthesia, or the lack thereof, will be needed. (5) It is tempting to speculate that if odorants are perceived under general anesthesia, its characteristics might be distorted. In other words, if anesthesia alters the percept of odorant X to that of odorant Y without changing the registration for odorant X, the second exposure to odorant X would be histologically registered as “familiar,” but behaviorally as novel. Conscious animals would perceive odorant X as X, which is not in line with the previously distorted percept.
Implications of state of mind under anesthesia
Our results have rather interesting implications for the study of consciousness. The relationship and interdependence between visual attention and awareness (or orienting) as a cortical-thalamic network model of consciousness have engendered much debate.56–58 Our study addresses a sensory system divorced from the cortical-thalamic network, yet we discovered potential implicit or “hidden” learning of novel stimuli that alter both the representation and sensitivity of subsequent c-Fos responses without the stimuli first entering consciousness. Although olfactory attention is difficult to measure and the current behavioral testing is not sensitive enough to detect olfactory awareness of cellular “memory,” our data nevertheless indicate an interesting scenario where the physical entity (c-Fos registration) of stimuli and the “qualia” of the same stimuli can be separated and are unlikely to be one and same process.
Consideration of anesthetic choice
Our choice of ketamine-xylazine anesthesia requires comment. Several studies showed that mitral cell firing was not different under ketamine-xylazine anesthesia and pentobarbital or urethane anesthesia.38,54,59,60 At low concentrations, ketamine is a so-called dissociative anesthetic, and its action is mediated by antagonizing the N-methyl-D-aspartate receptor. At high concentrations, the ketamine (100 mg/kg) and xylazine (5 mg/kg) combination was found to suppress α, β, and γ waves and increase δ oscillations, resembling the electroencephalography pattern in slow-wave sleep or unconsciousness under γ-aminobutyric acid type A receptor-mediated anesthesia.61 Moreover, xylazine is a centrally acting sedative and ketamine-xylazine anesthesia reduces energy consumption in the frontal and cingulate cortices, basal forebrain, and hippocampus,61 indicating LOC in a classical sense. Although recent studies, including high-resolution structural studies in our lab,62 have elucidated the possible ketamine action on pentameric ligand-gated ion channels including γ-aminobutyric acid type A receptors63 and neuronal acetylcholine receptors,64,65 it is important to emphasize that our results should not be extrapolated to implying that other general anesthetics with putative actions on γ-aminobutyric acid type A receptors or acetylcholine receptors would produce the same effects as discovered in this study. Further in-depth experimental investigations with other general anesthetics are still required before our conclusion can be generalized.
Clinical relevance and conclusion
In this study, all endpoints essential for clinical anesthesia and behavioral LOC were reached and the classical definition of unconsciousness was met, yet we discovered that higher-order sensory processes continue to occur in behaviorally unconscious brains. This discovery, however, should not be misconstrued as indicating awareness under general anesthesia. There are still many unknowns about the state of the mind under anesthesia that require further investigations in humans. For example, what exactly do the surrogate measures in the clinical monitors of anesthesia depth truly indicate if high-order sensory processing is partially maintained? Will the cellular registration of a novel sensory input under general anesthesia make the learning of the same sensory input more difficult later in an awake state given that some level of cellular registration has occurred? With the ability to investigate histology and behavior in the same animals, our study revealed a fundamental disconnection between neural registration and behavioral reporting of the same sensory information under general anesthesia that requires future clinical validation.
Final Boxed Summary Statement.
What we already know about this topic:
Whether and to what extent the brain processes sensory inputs during general anesthesia remains unclear.
What this article tells us that is new:
The authors demonstrated that in ketamine-xylazine anesthetized rats, odor presentation resulted in histologically distinguishable registration of sensory process, suggesting that general anesthesia causes disintegration of information processing but may preserve cellular communication.
Acknowledgments
Funding Information: This work was supported by a MERIT Award from the National Institute of General Medical Sciences, Bethesda, MD (R37GM049202 to YX and PT).
The authors thank Mr. James D. Bowman, B.S., Student Research Assistant, Department of Anesthesiology, for technical assistance in animal handling and histology, Dr. Eloise Peet and Professor Edda Thiels, Ph.D., Department of Neurobiology, for help with animal behavior testing, and Ms. Sandra Hirsch, M.B.A, Research Coordinator, Department of Anesthesiology, for editorial assistance. These individuals were affiliated with the University of Pittsburgh School of Medicine, Pittsburgh, PA 15213, USA.
Footnotes
Conflict of Interest: The authors declare no competing interests.
References
- 1.Kroeger D, Amzica F. Hypersensitivity of the anesthesia-induced comatose brain. J Neurosci. 2007;27:10597–607. doi: 10.1523/JNEUROSCI.3440-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cimenser A, Purdon PL, Pierce ET, Walsh JL, Salazar-Gomez AF, Harrell PG, Tavares-Stoeckel C, Habeeb K, Brown EN. Tracking brain states under general anesthesia by using global coherence analysis. Proc Natl Acad Sci U S A. 2011;108:8832–7. doi: 10.1073/pnas.1017041108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Campagna JA, Miller KW, Forman SA. Mechanisms of actions of inhaled anesthetics. N Engl J Med. 2003;348:2110–24. doi: 10.1056/NEJMra021261. [DOI] [PubMed] [Google Scholar]
- 4.Angel A. The G. L. Brown lecture. Adventures in anaesthesia. Exp Physiol. 1991;76:1–38. doi: 10.1113/expphysiol.1991.sp003471. [DOI] [PubMed] [Google Scholar]
- 5.Heinke W, Koelsch S. The effects of anesthetics on brain activity and cognitive function. Curr Opin Anaesthesiol. 2005;18:625–31. doi: 10.1097/01.aco.0000189879.67092.12. [DOI] [PubMed] [Google Scholar]
- 6.Hudetz AG. Suppressing consciousness: Mechanisms of general anesthesia. Seminars in Anesthesia, Perioperative Medicine and Pain. 2006;25:196–204. [Google Scholar]
- 7.Franks KM, Russo MJ, Sosulski DL, Mulligan AA, Siegelbaum SA, Axel R. Recurrent circuitry dynamically shapes the activation of piriform cortex. Neuron. 2011;72:49–56. doi: 10.1016/j.neuron.2011.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Onoda N. Odor-induced fos-like immunoreactivity in the rat olfactory bulb. Neurosci Lett. 1992;137:157–60. doi: 10.1016/0304-3940(92)90393-l. [DOI] [PubMed] [Google Scholar]
- 9.Woo CC, Oshita MH, Leon M. A learned odor decreases the number of Fos-immunopositive granule cells in the olfactory bulb of young rats. Brain Res. 1996;716:149–56. doi: 10.1016/0006-8993(96)00037-6. [DOI] [PubMed] [Google Scholar]
- 10.Sheng M, Greenberg ME. The regulation and function of c-fos and other immediate early genes in the nervous system. Neuron. 1990;4:477–85. doi: 10.1016/0896-6273(90)90106-p. [DOI] [PubMed] [Google Scholar]
- 11.Mandairon N, Didier A, Linster C. Odor enrichment increases interneurons responsiveness in spatially defined regions of the olfactory bulb correlated with perception. Neurobiol Learn Mem. 2008;90:178–84. doi: 10.1016/j.nlm.2008.02.008. [DOI] [PubMed] [Google Scholar]
- 12.Inaki K, Takahashi YK, Nagayama S, Mori K. Molecular-feature domains with posterodorsal-anteroventral polarity in the symmetrical sensory maps of the mouse olfactory bulb: Mapping of odourant-induced Zif268 expression. Eur J Neurosci. 2002;15:1563–74. doi: 10.1046/j.1460-9568.2002.01991.x. [DOI] [PubMed] [Google Scholar]
- 13.Paylor R, Johnson RS, Papaioannou V, Spiegelman BM, Wehner JM. Behavioral assessment of c-fos mutant mice. Brain Res. 1994;651:275–82. doi: 10.1016/0006-8993(94)90707-2. [DOI] [PubMed] [Google Scholar]
- 14.Grimm R, Schicknick H, Riede I, Gundelfinger ED, Herdegen T, Zuschratter W, Tischmeyer W. Suppression of c-fos induction in rat brain impairs retention of a brightness discrimination reaction. Learn Mem. 1997;3:402–13. doi: 10.1101/lm.3.5.402. [DOI] [PubMed] [Google Scholar]
- 15.Fleischmann A, Hvalby O, Jensen V, Strekalova T, Zacher C, Layer LE, Kvello A, Reschke M, Spanagel R, Sprengel R, Wagner EF, Gass P. Impaired long-term memory and NR2A-type NMDA receptor-dependent synaptic plasticity in mice lacking c-Fos in the CNS. J Neurosci. 2003;23:9116–22. doi: 10.1523/JNEUROSCI.23-27-09116.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Terleph TA, Tremere LA. In: The Use of Immediate Early Genes as Mapping Tools for Neuronal Activation: Concepts and Methods, Immediate early genes in sensory processing, cognitive performance and neurological disorders. Pinaud R, Tremere LA, editors. New York: Springer; 2006. pp. 1–10. [Google Scholar]
- 17.Morgan JI, Curran T. Stimulus-transcription coupling in the nervous system: Involvement of the inducible proto-oncogenes fos and jun. Annu Rev Neurosci. 1991;14:421–51. doi: 10.1146/annurev.ne.14.030191.002225. [DOI] [PubMed] [Google Scholar]
- 18.Bepari AK, Watanabe K, Yamaguchi M, Tamamaki N, Takebayashi H. Visualization of odor-induced neuronal activity by immediate early gene expression. BMC Neurosci. 2012;13:140. doi: 10.1186/1471-2202-13-140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Spinetta MJ, Woodlee MT, Feinberg LM, Stroud C, Schallert K, Cormack LK, Schallert T. Alcohol-induced retrograde memory impairment in rats: Prevention by caffeine. Psychopharmacology. 2008;201:361–71. doi: 10.1007/s00213-008-1294-5. [DOI] [PubMed] [Google Scholar]
- 20.O’Dell SJ, Feinberg LM, Marshall JF. A neurotoxic regimen of methamphetamine impairs novelty recognition as measured by a social odor-based task. Behav Brain Res. 2011;216:396–401. doi: 10.1016/j.bbr.2010.08.022. [DOI] [PubMed] [Google Scholar]
- 21.Canet J, Castillo J. Ketamine: A familiar drug we trust. Anesthesiology. 2012;116:6–8. doi: 10.1097/ALN.0b013e31823da398. [DOI] [PubMed] [Google Scholar]
- 22.Eikermann M, Grosse-Sundrup M, Zaremba S, Henry ME, Bittner EA, Hoffmann U, Chamberlin NL. Ketamine activates breathing and abolishes the coupling between loss of consciousness and upper airway dilator muscle dysfunction. Anesthesiology. 2012;116:35–46. doi: 10.1097/ALN.0b013e31823d010a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Stettler DD, Axel R. Representations of odor in the piriform cortex. Neuron. 2009;63:854–64. doi: 10.1016/j.neuron.2009.09.005. [DOI] [PubMed] [Google Scholar]
- 24.Poo C, Isaacson JS. Odor representations in olfactory cortex: “Sparse” coding, global inhibition, and oscillations. Neuron. 2009;62:850–61. doi: 10.1016/j.neuron.2009.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Miyamichi K, Amat F, Moussavi F, Wang C, Wickersham I, Wall NR, Taniguchi H, Tasic B, Huang ZJ, He Z, Callaway EM, Horowitz MA, Luo L. Cortical representations of olfactory input by trans-synaptic tracing. Nature. 2011;472:191–6. doi: 10.1038/nature09714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Buck LB. Olfactory receptors and odor coding in mammals. Nutr Rev. 2004;62:S184–8. doi: 10.1111/j.1753-4887.2004.tb00097.x. discussion S224–41. [DOI] [PubMed] [Google Scholar]
- 27.Kristiansen SL, Nyengaard JR. Digital stereology in neuropathology. APMIS. 2012;120:327–40. doi: 10.1111/j.1600-0463.2012.02889.x. [DOI] [PubMed] [Google Scholar]
- 28.Boyce RW, Dorph-Petersen KA, Lyck L, Gundersen HJ. Design-based stereology: Introduction to basic concepts and practical approaches for estimation of cell number. Toxicol Pathol. 2010;38:1011–25. doi: 10.1177/0192623310385140. [DOI] [PubMed] [Google Scholar]
- 29.Peterson DA. Quantitative histology using confocal microscopy: Implementation of unbiased stereology procedures. Methods. 1999;18:493–507. doi: 10.1006/meth.1999.0818. [DOI] [PubMed] [Google Scholar]
- 30.King MA, Scotty N, Klein RL, Meyer EM. Particle detection, number estimation, and feature measurement in gene transfer studies: Optical fractionator stereology integrated with digital image processing and analysis. Methods. 2002;28:293–9. doi: 10.1016/s1046-2023(02)00235-9. [DOI] [PubMed] [Google Scholar]
- 31.Swanson LW. Brain maps: Structure of the rat brain: A laboratory guide with printed and electronic templates for data, models, and schematics. 2. Chapter VII. Amsterdam ; New York: Elsevier; 1998. pp. 17–163. Rev Edition. [Google Scholar]
- 32.Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 4. San Diego: Academic Press; 1998. [Google Scholar]
- 33.Gjerstad J, Lien GF, Pedersen LM, Valen EC, Mollerup S. Changes in gene expression of Zif, c-fos and cyclooxygenase-2 associated with spinal long-term potentiation. Neuroreport. 2005;16:1477–81. doi: 10.1097/01.wnr.0000177004.19946.12. [DOI] [PubMed] [Google Scholar]
- 34.Abraham WC, Mason SE, Demmer J, Williams JM, Richardson CL, Tate WP, Lawlor PA, Dragunow M. Correlations between immediate early gene induction and the persistence of long-term potentiation. Neuroscience. 1993;56:717–27. doi: 10.1016/0306-4522(93)90369-q. [DOI] [PubMed] [Google Scholar]
- 35.Abraham WC, Dragunow M, Tate WP. The role of immediate early genes in the stabilization of long-term potentiation. Mol Neurobiol. 1991;5:297–314. doi: 10.1007/BF02935553. [DOI] [PubMed] [Google Scholar]
- 36.Wisden W, Errington ML, Williams S, Dunnett SB, Waters C, Hitchcock D, Evan G, Bliss TV, Hunt SP. Differential expression of immediate early genes in the hippocampus and spinal cord. Neuron. 1990;4:603–14. doi: 10.1016/0896-6273(90)90118-y. [DOI] [PubMed] [Google Scholar]
- 37.Wilson DA. Rapid, experience-induced enhancement in odorant discrimination by anterior piriform cortex neurons. J Neurophysiol. 2003;90:65–72. doi: 10.1152/jn.00133.2003. [DOI] [PubMed] [Google Scholar]
- 38.Rinberg D, Koulakov A, Gelperin A. Sparse odor coding in awake behaving mice. J Neurosci. 2006;26:8857–65. doi: 10.1523/JNEUROSCI.0884-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kadohisa M, Wilson DA. Separate encoding of identity and similarity of complex familiar odors in piriform cortex. Proc Natl Acad Sci U S A. 2006;103:15206–11. doi: 10.1073/pnas.0604313103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chapuis J, Wilson DA. Bidirectional plasticity of cortical pattern recognition and behavioral sensory acuity. Nat Neurosci. 2012;15:155–61. doi: 10.1038/nn.2966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sahraie A, Hibbard PB, Trevethan CT, Ritchie KL, Weiskrantz L. Consciousness of the first order in blindsight. Proc Natl Acad Sci U S A. 2010;107:21217–22. doi: 10.1073/pnas.1015652107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Franks NP. General anaesthesia: From molecular targets to neuronal pathways of sleep and arousal. Nat Rev Neurosci. 2008;9:370–86. doi: 10.1038/nrn2372. [DOI] [PubMed] [Google Scholar]
- 43.Alkire MT, Hudetz AG, Tononi G. Consciousness and anesthesia. Science. 2008;322:876–80. doi: 10.1126/science.1149213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Boly M, Moran R, Murphy M, Boveroux P, Bruno MA, Noirhomme Q, Ledoux D, Bonhomme V, Brichant JF, Tononi G, Laureys S, Friston K. Connectivity Changes Underlying Spectral EEG Changes during Propofol-Induced Loss of Consciousness. J Neurosci. 2012;32:7082–90. doi: 10.1523/JNEUROSCI.3769-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Guzowski JF, Setlow B, Wagner EK, McGaugh JL. Experience-dependent gene expression in the rat hippocampus after spatial learning: A comparison of the immediate-early genes Arc, c-fos, and zif268. J Neurosci. 2001;21:5089–98. doi: 10.1523/JNEUROSCI.21-14-05089.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Duncan GE, Moy SS, Knapp DJ, Mueller RA, Breese GR. Metabolic mapping of the rat brain after subanesthetic doses of ketamine: Potential relevance to schizophrenia. Brain Res. 1998;787:181–90. doi: 10.1016/s0006-8993(97)01390-5. [DOI] [PubMed] [Google Scholar]
- 47.Sallaz M, Jourdan F. Odour-induced c-fos expression in the rat olfactory bulb: Involvement of centrifugal afferents. Brain Res. 1996;721:66–75. doi: 10.1016/0006-8993(96)00150-3. [DOI] [PubMed] [Google Scholar]
- 48.Cirelli C, Pompeiano M, Tononi G. Neuronal gene expression in the waking state: A role for the locus coeruleus. Science. 1996;274:1211–5. doi: 10.1126/science.274.5290.1211. [DOI] [PubMed] [Google Scholar]
- 49.Jin BK, Franzen L, Baker H. Regulation of c-Fos mRNA and fos protein expression in olfactory bulbs from unilaterally odor-deprived adult mice. Int J Dev Neurosci. 1996;14:971–82. doi: 10.1016/s0736-5748(96)00044-5. [DOI] [PubMed] [Google Scholar]
- 50.Rose SP. How chicks make memories: The cellular cascade from c-fos to dendritic remodelling. Trends Neurosci. 1991;14:390–7. doi: 10.1016/0166-2236(91)90027-r. [DOI] [PubMed] [Google Scholar]
- 51.Kaczmarek L. Molecular biology of vertebrate learning: Is c-fos a new beginning? J Neurosci Res. 1993;34:377–81. doi: 10.1002/jnr.490340402. [DOI] [PubMed] [Google Scholar]
- 52.Lai WS, Chen A, Johnston RE. Patterns of neural activation associated with exposure to odors from a familiar winner in male golden hamsters. Horm Behav. 2004;46:319–29. doi: 10.1016/j.yhbeh.2004.06.007. [DOI] [PubMed] [Google Scholar]
- 53.Shea SD, Katz LC, Mooney R. Noradrenergic induction of odor-specific neural habituation and olfactory memories. J Neurosci. 2008;28:10711–9. doi: 10.1523/JNEUROSCI.3853-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kato HK, Chu MW, Isaacson JS, Komiyama T. Dynamic sensory representations in the olfactory bulb: Modulation by wakefulness and experience. Neuron. 2012;76:962–75. doi: 10.1016/j.neuron.2012.09.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hara K, Harris RA. The anesthetic mechanism of urethane: The effects on neurotransmitter-gated ion channels. Anesth Analg. 2002;94:313–8. doi: 10.1097/00000539-200202000-00015. [DOI] [PubMed] [Google Scholar]
- 56.Prinz J. A neurofunctional theory of visual consciousness. Conscious Cogn. 2000;9:243–59. doi: 10.1006/ccog.2000.0442. [DOI] [PubMed] [Google Scholar]
- 57.Kentridge RW, Heywood CA, Weiskrantz L. Attention without awareness in blindsight. Proc Biol Sci. 1999;266:1805–11. doi: 10.1098/rspb.1999.0850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Posner MI. Attention: The mechanisms of consciousness. Proc Natl Acad Sci U S A. 1994;91:7398–403. doi: 10.1073/pnas.91.16.7398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Nagayama S, Takahashi YK, Yoshihara Y, Mori K. Mitral and tufted cells differ in the decoding manner of odor maps in the rat olfactory bulb. J Neurophysiol. 2004;91:2532–40. doi: 10.1152/jn.01266.2003. [DOI] [PubMed] [Google Scholar]
- 60.Wellis DP, Scott JW. Intracellular responses of identified rat olfactory bulb interneurons to electrical and odor stimulation. J Neurophysiol. 1990;64:932–47. doi: 10.1152/jn.1990.64.3.932. [DOI] [PubMed] [Google Scholar]
- 61.Dworak M, McCarley RW, Kim T, Basheer R. Delta oscillations induced by ketamine increase energy levels in sleep-wake related brain regions. Neuroscience. 2011;197:72–9. doi: 10.1016/j.neuroscience.2011.09.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Pan J, Chen Q, Willenbring D, Mowrey D, Kong XP, Cohen A, Divito CB, Xu Y, Tang P. Structure of the pentameric ligand-gated ion channel GLIC bound with anesthetic ketamine. Structure. 2012;20:1463–9. doi: 10.1016/j.str.2012.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hevers W, Hadley SH, Luddens H, Amin J. Ketamine, but not phencyclidine, selectively modulates cerebellar GABA(A) receptors containing alpha6 and delta subunits. J Neurosci. 2008;28:5383–93. doi: 10.1523/JNEUROSCI.5443-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Yamakura T, Chavez-Noriega LE, Harris RA. Subunit-dependent inhibition of human neuronal nicotinic acetylcholine receptors and other ligand-gated ion channels by dissociative anesthetics ketamine and dizocilpine. Anesthesiology. 2000;92:1144–53. doi: 10.1097/00000542-200004000-00033. [DOI] [PubMed] [Google Scholar]
- 65.Coates KM, Flood P. Ketamine and its preservative, benzethonium chloride, both inhibit human recombinant alpha7 and alpha4beta2 neuronal nicotinic acetylcholine receptors in Xenopus oocytes. Br J Pharmacol. 2001;134:871–9. doi: 10.1038/sj.bjp.0704315. [DOI] [PMC free article] [PubMed] [Google Scholar]