

Abstract
The purpose of this study was to investigate the relationship between 25-m sprint front crawl swimming performance and muscle fascicle length in young male swimmers. 23 swimmers were selected and divided into two groups according to their best records of 25-m sprint performance: 14.6-15.7 sec (S1, n = 11) and 15.8-17 sec (S2, n = 12). Muscle thickness and pennation angle of Biceps Brachii (BB; only muscle thickness), Triceps Brachii (TB), Vastus Lateralis (VL), Gastrocnemius Medialis (GM) and Lateralis (GL) muscles were measured by B-mode ultrasonography, and fascicle length was estimated. Although, there was no significant differences between groups in anthropometrical parameter as standing height, body mass, arm length, thigh length and leg length (p < 0.001), however, S1 significantly had a greater muscle thickness in VL, GL, and TB muscles (p < 0.05). Pennation angle only in TB was significantly smaller in S1 (p < 0.05). S1 in VL, GL, and TB muscles significantly had greater absolute fascicle length and in VL and TB muscles had relatively (relative to limb length) greater fascicle length (p < 0.05). Moreover, there was a significant relationship between sprint swimming time and absolute and relative fascicle length in VL (absolute: r = -0.49 and relative: r = -0.43, both p < 0.05) and GL (absolute: r = -0.47 and relative: r = -0.42, both p < 0.05). Potentially, it seems that fascicle geometry developed in muscles of faster young swimmers to help them to perform their high speed movement.
Key Points.
This study investigated the relationship between muscle fascicle length and sprint front crawl performance in young male swimmers.
It seems that young swimmers with faster front crawl sprint swimming performance trend to have smaller pennation angle and greater absolute and relative fascicle length (relative to limb length) in their locomotor muscles.
Potentially, fascicle geometry developed in faster swimmers to help them to perform higher speed movement via higher output power, however, the adaptive response of fiber length follownig training is not well underestood.
Key words: Muscle architecture, fascicle length, ultrasonography, sprint swimming, front crawl
Introduction
Performance in swimming has been related to different anthropometrical, physiological, and biomechanical parameters (Lätt et al., 2010; Vitor and Böhme, 2010). From physiological point of view one of the main factors that affect’s sprint performance capacity in swimming is power generation of locomotor muscles. In this sense, strength and speed are among major factors in a sprint swimmer’s training program normally (Girold et al., 2006). Considering these two factors as components of power generation during sprint swimming (Hawley et al., 1992); therefore, the active muscles should generate a high rate of powerful shortening to produce maximal mechanical power outputs (Kumagai et al., 2000; Stager and Tanner, 2005).
Among the factors that could affect the ability of muscle power generation, muscle architectural characteristic such as fascicle length is an important indicator (Blazevich, 2006). Although biochemical properties (myosin ATPase activity) are important in determining maximal shortening velocity of muscle (Barany, 1967), however, muscle architectural characteristics such as muscle fascicle length have been shown to play an important role in modulating biochemical effects (Blazevich, 2006; Sacks and Roy, 1982; Spector et al., 1980). Moreover, differences in maximal shortening velocity between muscles are more closely associated with differences in muscle fiber length rather than its biochemical differences, and muscle fascicle arrangement is probably a greater determinant of general muscle function than other aspects such as fiber type (Burkholder et al., 1994).
Recently, Nasirzade et al. (2013) demonstrated the relative contribution of different architectural parameters such as fascicle length and muscle thickness of vastus lateralis from different biomechanical, anthropometrical, and muscle architectural parameters in predicting 50-m sprint front crawl swimming performance in young male swimmers. Although there has not been another study to examine these parameters in swimmers, the results of similar studies on athletes that required high muscle power production such as sprinting demonstrated that sprinters compared to endurance runners and untrained individuals have greater fascicle in their locomotor muscles (Abe et al., 2000). Also, significant relationship between longer fascicle length and the best-personal sprint performance have been found (Abe et al., 2001; Kumagai et al., 2000). These findings demonstrated that the difference in sprint performance could be related to the difference in muscles fascicle length. Based on this information, we assumed that fascicle length might play an important role in sprint swimming performance. Hence, the purpose of present study was to investigate the relationship between fascicle length of different muscles and sprint performance of front crawl swimming in young male swimmers.
Methods
Subjects
Twenty-three young male country level swimmers (age: 13.86 ± 0.86 years) belonging to the same local swimming club, with background of 4 to 6 years of regular training experience (8.3 ± 1.1 h/week) participated in this study. All children and parents were thoroughly informed of the purposes and contents of the study and a written informed consent was obtained from the parents before participation. The study was approved by the local University Ethics Committee.
Study design
Each subject was measured on 2 separate days at approximately the same time of day on 3 different occasions. On the first day, selected anthropometrical parameters and swimming performance time were determined. Anthropometric measurements were taken before swimming test. On the second day, ultrasound images for muscle architectural parameters assessment were taken.
Swimming performance
A video camera with rate of 30 Hz filmed all trials with a profile view from above the pool. After taking the 25-m sprint front crawl swimming times, the records ranged between 14.62 and 16.94 sec. For comparison the sample was divided into two groups based on the mean (15.70 ± 0.74 s) of the 25-m swimming records: 14.62-15.67 sec (S1, n = 11) and 15.79-16.94 sec (S2, n = 12). All the swimmers performed 2 maximal front crawl 25-m trials with a 15 min active recovery period between the 2 trials. The faster trial was accepted as the swimmer performance. In-water starts were used and swimmers performed the maximal trial alone in each lane. The time spent to cover the 25-m between pushing off from the wall and touching the wall on the other side of the pool was recorded as the swimmer performance time.
Anthropometric data
Height and body mass were measured using standard laboratory equipment, and body-mass index (BMI) was calculated. Limbs length were measured from the right side of body in standing position by using anatomic landmarks: upper leg length, the distance between the lateral condyle of the femur and greater trochanter, lower leg length, the distance between the lateral malleolus of the fibula and the lateral condyle of the tibia, and upper arm length, the distance between the lateral epicondyle of the humerus and the acromial process of the scapula. All anthropometric parameters were measured to the nearest 0.1 cm by two expert evaluators in accordance with the ISAK guidelines (Stewart et al., 2011), using a sliding caliper (Sanny, American Medical from Brazil, BR). Mean value of both measurements was computed (ICC=0.95) (Weir, 2005).
Skeletal muscle architectural parameters
Five different muscles were investigated including Vastus Lateralis (VL; midway between the lateral condyle of the femur and greater trochanter), Gastrocnemius Lateralis (GL; 30% proximal between the lateral malleolus of the fibula and the lateral condyle of the tibia), and Gastrocnemius Medialis (GM; at the same level as GL) in lower extremity (Kumagai et al., 2000) and lateral head of Triceps Brachii (TB; midway of lateral epicondyle of the humerus and the acromial process of the scapula) and Biceps Brachii (BB; at the same level as TB) in upper extremity (Matta et al., 2011). These muscles are highly activated and recruited during front crawl swimming propulsion (Birrer, 1986; McLeod, 2010).
Muscle architectural characteristics were measured in vivo as described previously (Kumagai et al., 2000; Matta et al., 2011; Reeves and Narici, 2003). Briefly, after determining the measurement sites and coating them with water-soluble transmission gel to aid optimum acoustic coupling, the probe was placed perpendicularly to the specific muscle to observe a cross-sectional image to measure the muscle thickness. After imaging and marking the images for later analyzing, the probe was placed at the same point and parallel to the specific muscle resulting in a longitudinal image to measure the pennation angle. The muscle thickness was defined as the distance between the superficial and deep aponeurosis of the muscle in the cross-sectional image (Figure 1; left). The pennation angle was measured as the acute angle of insertion of muscle fiber fascicles into the deep aponeurosis in the longitudinal image (α in Figure 1; right). Finally, fascicle length was estimated by the following geometric formula (L in Figure 1; right):
Figure 1.
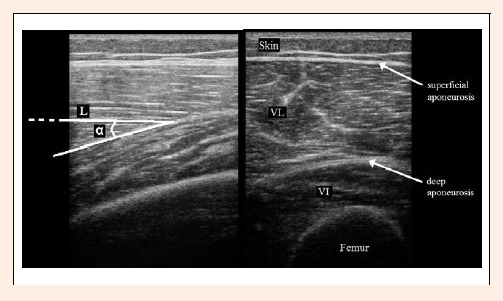
Ultrasound images of Vastus Lateralis (VL) and Intermedius (VI) muscles. Left: cross-sectional image. Right: longitudinal image representing pennation angle (α) and fascicle length (L) of VL.
| Fascicle length = Muscle thickness · (Sin (α))-1 | (1) |
where α is the pennation angle (Kumagai et al., 2000). Thus, fascicles were assumed straight and the model did not account for fascicle curvature. The error involved with this technique has been shown to be reasonably low (~2.3%) in contracted Tibialis Anterior (Reeves and Narici, 2003), where fascicle curvature is significant. Given that fascicles in the relaxed muscle are relatively straight (see Figure 1; right), we estimated that our error would be somewhat smaller.
All measurements were conducted from the right side of body in the morning before performing any physical activity, and with the same condition for all subjects. Imaging from BB and VL were taken in supine position and TB, GM, and GL recorded in prone position with arms and legs extended. Images were taken by B-mode ultrasonography (EUB-405, Hitachi, Japan) with a 7.5-MHz linear-array probe.
Two ultrasound images were taken from each trial, and the average of the two values for each variable was used for further analysis. All measurements were carried out by the same experienced sonographer. Images were analyzed with publicly available imaging software (ImageJ 1.47c, National Institutes of Health, Bethesda, MD, USA). The coefficient of variation for muscle thickness, pennation angle, and fascicle length from the two images were 2.4, 1.8, and 2.3%, respectively.
Statistical analysis
Results are expressed as means ± SD. Normality of distribution was checked with Kolmogorov-Smirnov test and normal distribution was found in all the subgroups. Independent-Samples T Test was used for comparison between the two performance groups and Pearson-product moment correlation was computed in SPSS (version 17) to verify the relationship between sprint front crawl swimming time and selected muscle architectural variables. In the entire study statistical significance set at p ≤ 0.05.
Results
Participant’s physical characteristics
Descriptive statistics for selected anthropometrical parameters and 25-m sprint swimming performance time are presented in Table 1. There were no differences in the anthropometrical parameters between groups, although S1 significantly had a faster swimming performance time than S2 (p > 0.05).
Table 1.
Anthropometrical parameters and 25-m sprint swimming performance time in S1 (n = 11) and S2 (n = 12).
| Variable | S1 | S2 | ||
|---|---|---|---|---|
| Mean (±SD) | Range | Mean (±SD) | Range | |
| Height (m) | 1.71 (.06) | 1.61 – 1.81 | 1.70 (.06) | 1.59 – 1.80 |
| Body mass (kg) | 57.8 (6.3) | 53.5 – 70.6 | 59.9 (6.1) | 56.1 – 71.2 |
| BMI (kg·m-2) | 19.6 (1.5) | 17.8 – 21.6 | 20.6 (1.2) | 18.2 – 22.1 |
| Thigh length (cm) | 40.1 (2.2) | 37.1 – 44.2 | 39.8 (2.0) | 36.3 – 43.1 |
| Lower leg length (cm) | 41.2 (2.3) | 38.5 – 45.2 | 40.6 (2.2) | 36.3 – 44.7 |
| Arm length (cm) | 37.5 (1.6) | 33.2 – 41.6 | 36.9 (2.0) | 33.4 – 40.8 |
| Swimming time (Sec) * | 15.07 (.37) | 14.62 – 15.67 | 16.31 (.40) | 15.79 – 16.94 |
* Significant differences between S1 and S2, p < 0.05.
Skeletal muscle architecture characteristics
S1 significantly had a greater VL, GL, and TB muscle thickness than S2 (Table 2). Only in the TB did the S1 have a significantly lower pennation angle than S2 (Table 2). S1 significantly had a greater fascicle length in VL, GL, and TB (Table 2). Relative fascicle length (relative to limb length) in VL and TB was significantly greater in S1 than in S2 (Table 2).
Muscle thickness had negative correlation with 25-m sprint front crawl swimming time in GM and GL (r = -0.41 and r = -0.45, both p >0.05), but not in BB, TB, and VL (respectively: r = -0.01; p = 0.68, r = -0.32; p = 0.13 and r = -0.30; p = 0.17). Pennation angle in GM was significantly correlated to swimming performance time (r = 0.42, p < 0.05), but there was no such correlation in VL, GL, and TB (respectively: r = 0.38; p = 0.07, r = 0.32; p = 0.13 and r = 0.18; p = 0.40). There were negative correlation between absolute fascicle length and swimming performance time in VL and GL (r = -0.49 and r = -0.47, both p > 0.05), but not in TB and GM (r = -0.23; p = 0.28 and r = -0.19; p = 0.38). Moreover, there were negative correlation between relative fascicle length (relative to limb length) and swimming performance time in VL and GL (Figure 2).
Figure 2.
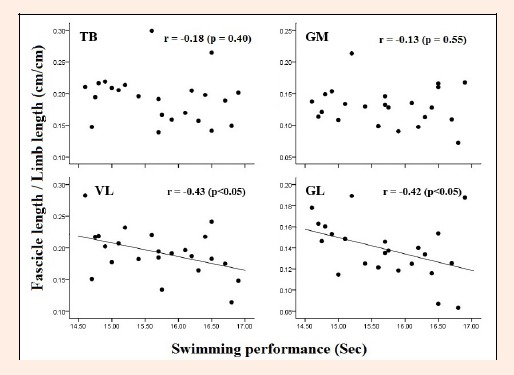
Relationships between 25-m front crawl swimming sprint performance time and relative fascicle length of Triceps Brachii (TB; left top), Vastus Lateralis (VL; right top), Gastrocnemius Medialis (GM; left bottom), and Gastrocnemius Lateralis (GL; right bottom) muscles.
Discussion
The purpose of the present study was to investigate the relationship between fascicle length of different muscles and sprint performance of front crawl swimming in young male swimmers. The results demonstrated that young swimmers with a faster front crawl swimming sprint performance significantly had a greater absolute fascicle length in VL, GL, and TB. Also, they had a significantly greater relative fascicle length (relative to limb length) in VL and GL. There was a significant relationship between muscle fascicle length (absolute and relative) and swimming performance time in VL and GL muscles.
It has been demonstrated that muscle fiber shortening velocity is determined by muscle fiber type composition and muscle fascicle length (Barany, 1967; Schluter and Fitts, 1994). Previous studies (Costill, 1978; Gerard et al., 1986) revealed that sprint swimmers have a high percentage of type II muscle fiber in their involved muscles. Nygaard and Nielsen (1978) did a cross-sectional study in which they found that swimmers had smaller Type I and IIa fibers in the deltoid muscle compared to controls, despite the fact that the overall size of the deltoid muscle was greater. However, muscle fiber type percentage could be varied among swimmers, even among those who have been trained for specific swimming distance (Sortwell, 2010). Also, evidence indicates that muscle fiber composition bears little relationship to performance capacities (Campbell et al., 1979). Despite the fact, it would seem likely that muscle fiber type is a more important muscle-based determinant of muscle endurance while fascicle geometry affects more the length-tension and force-velocity characteristics of a muscle (Blazevich, 2006).
Fascicle-shortening velocity is an important factor in determining muscle power output (Stager and Tanner, 2005). Potentially, there are two possibilities for longer fascicle length leading to a faster sprint performance. First, the longer fascicle length would result in greater maximal shortening velocity (Abe et al., 2001; Kumagai et al., 2000). Therefore, faster velocity would results in greater power, which leads to improvement of sprint performance. Thom et al. (2007) estimated that the decreasing of fascicle length due to losing of sarcomeres placed in one series results in 50% decrease in total maximum velocity of muscle shortening of GM in the old men compared with young men. Second, according to Hill’s equation (Hill, 1970), with increasing the velocity of muscle shortening, its force generating would decrease. However, the shortening speed of each sarcomere in a fiber or fascicle would be slower for a given speed of whole-fiber shortening when there are more sarcomeres in series. Also, according to the force-velocity relationship, sarcomere force would not decrease as rapidly as fiber-shortening speed increase (Blazevich, 2006; Kumagai et al., 2000). Therefore, at high shortening velocities, longer fascicles are capable of generating greater force (Blazevich, 2006). As a result, power would be greater which improve the sprint performance potentially.
Previously, the effects of muscle architectural parameters on swimmers performance had not been studied. However, similar studies in this field demonstrated that fascicle length (absolute and relative) is significantly greater in locomotor muscles such as VL and GL of elite sprinters with a faster sprint running performance time (Abe et al., 2001; Kumagai et al., 2000). In present study negative relationship has found between fascicle length (absolute and relative) of VL and GL muscles with sprint swimming performance. This is maybe due to the strong relationship (r = 0.76) between lower-body power output and sprint (50-m) front crawl swimming performance in young swimmers (Hawley et al., 1992). Moreover, the correlation of r=0.90, has been reported between 25-m sprint front crawl swimming performance and leg power in 8-12 year old children (Inbar and Bar-Or, 1977). Although in front crawl actual contribution of leg kick in the 10% gain in maximal velocity in front crawl swimming (Deschodt et al., 1999), however, leg action smooth the intracycle fluctuations of swimming velocity (Persyn et al., 1975), and also useful in keeping the body horizontal in the water, especially during sprints (Hollander et al., 1988), and therefore can be improved arm stroke efficiency.
In the musculoskeletal system, muscles generate force and transmit that force via tendons to bone. The greater the force generated by muscle, the greater the stress transmitted through the tendon. Increases in the power requirement of our muscles (e.g. with speed) require a stiffer tendon to produce optimal efficiency and also to produce the required power with the given muscle volume (Lichtwarh and Wilson, 2008). However, there is no evidence about front crawl flatter kick, it seems that in such activity like walking and running the GM fascicles contracted almost isometrically (Alexander, 2002). Indeed, the tendon stretched and recoiled as the ankle flexed and extended, but the muscle fascicles remained almost constant in length and hardly did any work. Therefore, tendon stiffness becomes an important factor in muscle power generation ability. However, as opposed to the well-documented changes in muscle with exercise, the adaptive significance of changes in tendon stiffness with exercise is unclear (Buchanan and Marsh, 2002). Although, to our best knowledge no investigation conducted on swimmers, Simonsen et al. (1995) found that tendons increased stiffness in rats in response to a swim-training program even though this type of exercise has low impact loads. In this sense, it would be interesting to determine the optimum fascicle length and tendon stiffness for swimming activities and compare them to data from specialists in this sport.
Considering the determining role of arm movement in generating propulsive force in front crawl swimming (Deschodt et al., 1999; Sharp et al., 1982), we investigated BB and TB muscles. Based on this consideration, muscle thickness of BB was similar between groups, but S1 had significantly greater muscle thickness in TB. Also, the group with faster sprint swimming performance (S1) had significantly smaller pennation angle and greater absolute and relative fascicle length (relative to arm length) in TB. So it seems that young swimmers with faster sprint performance benefit from greater fascicle length of TB in generating greater power.
Hawley et al. (1992) reported a positive correlation between mean power of active muscles on shoulder joint and sprint front crawl swimming performance in a group of trained young swimmers with same average age to the present study (r = 0.63, p < 0.01). Also, Inbar and Bar-Or (1977) found a correlation of r = -0.92 between mean power of the arms and 25-m freestyle time in a small group of young swimmers (n = 9). As a result, it would be expected that there should be a correlation between architectural characteristics especially fascicle length (absolute and relative) of TB and sprint front crawl swimming performance.
In the present study there was no correlation between architectural characteristics of TB and sprint front crawl swimming performance. These results can be due to several reasons that affect sprint performance of young swimmers: (i) Based on literature (Stager and Tanner, 2005; Van Praagh, 2000), genetic factors influenced anaerobic muscle function and performance, and genetic predispositions contribute around 50% of variance in short-term anaerobic performance. This fact supports the contention that sprinters might indeed be born, rather than developed (Stager and Tanner, 2005). (ii) Indeed, usage of potentially higher output muscle power due to the longer fascicle length to generate faster forward motion in swimming affected by swimmers technique. As shown by Lätt et al. (2010) from 28 different biomechanical, anthropometrical, and physiological parameters, stroke rate and stroke index alone explained 92.6% of 100-m front crawl swimming performance variability in adolescent male swimmers (15.2±1.9 years). Also, Vitor and Böhme (2010) reported that stroke index (r2 = 0.62) is one of the effective factors for predicting 100-m front crawl swimming performance in young male swimmers (13.0 ± 0.7 years). Thus, stroke technique parameters play an important role in front crawl swimming performance.
During these ages, long distance aerobic training is a major component of young swimmers training program as our subjects. These trainings could be affected and interrupted the relationship between sprint swimming performance and muscular architecture parameters. Therefore, to verify this relationship more clearly it is necessary to perform similar works on elite sprint swimmers. Moreover, it would be required to consider the impact of different muscular architecture parameters concomitant with major factors such as anthropometrical, physiological, and technical those affecting swimming performance. Such studies will help elucidate the most important factors that are responsible for differentiation between individuals performance.
Conclusion
Potentially, it seems that fascicle geometry developed in faster swimmers to help them to perform higher speed movement via higher output power; However, in present study the relationship between sprint swimming performance and fascicle length (absolute and relative) only confirmed for VL and GL muscles, and unexpectedly not in TB; therefore, to achieve more reliable data similar studies on high-class swimmers are required.
Table 2.
Architectural parameters of Vastus Lateralis (VL), Gastrocnemius Medialis (GM), Gastrocnemius Lateralis (GL), Triceps Brachii (TB), and Biceps Brachii (BB) muscles in S1 (n = 11) and S2 (n = 12).
| Variable | S1 | S2 | |||
|---|---|---|---|---|---|
| Mean (±SD) | Range | Mean (±SD) | Range | ||
| Muscle thickness (cm) | VL * | 2.25 (.12) | 2.01 - 2.45 | 2.14 (.11) | 1.98 - 2.31 |
| GM | 1.88 (.19) | 1.52 - 2.15 | 1.75 (.17) | 1.41 - 2.02 | |
| GL * | 1.70 (.21) | 1.18 - 2.03 | 1.51 (.20) | 1.08 - 2.01 | |
| TB * | 2.24 (.15) | 2.04 - 2.48 | 2.10 (.13) | 1.92 - 2.41 | |
| BB | 2.08 (.21) | 1.79 - 2.47 | 2.04 (.22) | 1.78 - 2.52 | |
| Pennation angle (°) | VL | 16.0 (2.6) | 12.3 - 22.3 | 18.3 (3.7) | 12.5 - 25.1 |
| GM | 20.2 (3.8) | 15.4 - 27.3 | 21.3 (5.1) | 13.3 - 29.0 | |
| GL | 16.1 (1.6) | 13.3 - 19.7 | 17.2 (2.1) | 14.1 - 21.2 | |
| TB * | 16.9 (2.3) | 12.6 - 21.6 | 19.1 (2.8) | 13.2 - 22.8 | |
| Absolute fascicle length (cm) | VL * | 8.30 (1.1) | 6.19 - 10.71 | 7.04 (1.2) | 4.91 - 9.16 |
| GM | 5.59 (1.0) | 4.23 - 8.11 | 5.04 (1.1) | 3.18 - 6.87 | |
| GL * | 6.13 (.7) | 5.13 - 7.19 | 5.23 (1.1) | 3.22 - 7.69 | |
| TB * | 7.86 (1.4) | 5.59 - 11.42 | 6.55 (1.1) | 5.08 - 8.99 | |
| Fascicle length/limb length (cm/cm) | VL * | .21 (.03) | .15 - .28 | .18 (.03) | .11 - .24 |
| GM | .14 (.03) | .10 - .21 | .12 (.03) | .07 - .17 | |
| GL | .15 (.02) | .11 - .19 | .13 (.03) | .08 - .19 | |
| TB * | .21 (.03) | .15 - .28 | .18 (.04) | .14 - .26 |
* Significant differences between S1 and S2, p < 0.05.
Acknowledgments
The authors would like to thank Mr. Seyed Abbas Musavi, head of Swimming Commission of Southern Khorasan for his cooperation. Special thanks to all the participating swimmers, their coaches, and their parents.
Biographies

Alireza NASIRZADE
Employment
PhD student in Sport Biomechanics at the Department of Physical Education and Sport Sciences of Kharazmi University, Tehran, Iran.
Degree
MSc
Research interest
Biomechanical research in swimming and gait analysis
E-mail: alireza.nasirzade@gmail.com

Alireza EHSANBAKHSH
Employment
Associate professor of radiology, Radiology Department of Birjand University of Medical Sciences, Birjand, Iran.
Degree
M.D, Associate professor
Research interest
Medical imaging
E-mail: a.r.ehsanbakhsh@gmail.com

Saeed ILBEIGI
Employment
Assistant Professor of Sport Biomechanics, Department of Sport Biomechanics, Faculty of Physical Education and Sport Sciences, University of Birjand, Birjand, Iran.
Degree
PhD
Research interest
Biomechanics of sprint and start
E-mail: silbeigi@birjand.ac.ir

Azadeh SOBHKHIZ
Employment
Instructor of Tehran University of Medical Sciences, Tehran, Iran.
Degree
MSc
Research interest
Improve performance, improve recovery time, and prevent injury
E-mail: Azadeh_sobh_khiz@yahoo.com

Hamed ARGHAVANI
Employment
PhD student, Department of Physical Education and Sport Sciences of Kharazmi University, Tehran, Iran.
Degree
MSc
Research interest
Biomechanics and skill acquisition in swimming
E-mail: hamed_arghavani1365@yahoo.com

Mahdi ALIAKBARI
Employment
MSc student in Sport and Exercise physiology at the Department of Physical Education and Sports Science University of Mazandaran, Baboulsar, Iran.
Degree
BSc
Research interest
Sport and exercise physiology
E-mail: m.aliakbari@stu.umz.ac.ir
References
- Abe T., Fukashiro S., Harada Y., Kawamoto K. (2001) Relationship between sprint performance and muscle fascicle length in female sprinters. Journal of Physiological Anthropology and Applied Human Science 20, 141-147 [DOI] [PubMed] [Google Scholar]
- Abe T., Kumagai K., Brechue W.F. (2000) Muscle fascicle length is greater in sprinters than long-distance runners. Medicine & Science in Sports & Exercise 32, 1125-1129 [DOI] [PubMed] [Google Scholar]
- Alexander R.M. (2002) Tendon elasticity and muscle function. Comparative Biochemistry and Physiology 133, 1001-1011 [DOI] [PubMed] [Google Scholar]
- Barany M. (1967) ATPase activity of myosin correlated with speed of muscle shortening. The Journal of General Physiology 50, 197-218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birrer P. (1986) The shoulder, EMG and the swimming stroke. Journal of Swimming Research 12, 20-23 [Google Scholar]
- Blazevich A.J. (2006) Effects of physical training and detraining, immobilization, growth and aging on human fascicle geometry. Sports Medicine 36, 1003-1017 [DOI] [PubMed] [Google Scholar]
- Buchanan C.I., Marsh R.L. (2002) Effects of exercise on the biomechanical, biochemical and structural properties of tendons. Comparative Biochemistry and Physiology 133, 1101-1107 [DOI] [PubMed] [Google Scholar]
- Burkholder T.J., Fingado B., Baron S., Lieber R.L. (1994) Relationship between muscle fiber types and sizes and muscle architecture properties in the mouse hindlimb. Journal of Morphology 221, 177–190 [DOI] [PubMed] [Google Scholar]
- Campbell C.J., Bonen A., Kirby R.L., Belcastro A.N. (1979) Muscle fiber composition and performance capacities of women. Medicine & Science in Sports & Exercise 11, 260-265 [PubMed] [Google Scholar]
- Costill D. (1978) Adaptations in skeletal muscle during training for sprint and endurance training. Swimming Medicine 194, 59-64 [Google Scholar]
- Deschodt V.J., Arsac L.M., Rouard A.H. (1999) Relative contribution of arms and legs in humans to propulsion in 25-m sprint front-crawl swimming. European Journal of Applied Physiology and Occupational Physiology 80, 192-199 [DOI] [PubMed] [Google Scholar]
- Gerard E.S., Caiozzo V.J., Rubin B.D., Prietto C.A., Davidson D.M. (1986) Skeletal muscle profiles among elite long, middle, and short distance swimmers. American Journal of Sports Medicine 14, 77-82 [DOI] [PubMed] [Google Scholar]
- Girold S., Calmels P., Maurin D., Mihau D., Chatard J.C. (2006) Assisted and resisted sprint training in swimming. The Journal of Strength & Conditioning Research 20, 547-554 [DOI] [PubMed] [Google Scholar]
- Hawley J.A., Williams M.M., Vickovic M.M., Handcock P.J. (1992) Muscle power predicts freestyle swimming performance. British Journal of Sports Medicine 26, 151-155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill A.V. (1970) First and last experiments in muscle mechanics. Cambridge University Press, London [Google Scholar]
- Hollander A., de Groot G., van lngen Schneau G., Kahman R., Toussaint H. (1988) Contributions of the legs to propulsion in front crawl swimming. : International Series on Sport Sciences. : Ungerechts B., Wilkie K., Reischle K., Swimming Science V. Champaign, IL: Human Kinetics; Vol 18 39-43, 69 [Google Scholar]
- Inbar O., Bar-Or O. (1977) Relationship of anaerobic and aerobic arm and leg capacities to swimming performance of 8-12 year old children. In: Frontiers of activity and child health. Eds: Shephard R.J., Lavellee H. 283-292 [Google Scholar]
- Kumagai K., Abe T., Brechue W.F., Ryushi T., Takano S., Mizuno M. (2000) Sprint performance is related to muscle fascicle length in male 100-m sprinters. Journal of Applied Physiology 88, 811-816 [DOI] [PubMed] [Google Scholar]
- Lätt E., Jürimäe J., Mäestu J., Purge P., Rämson R., Haljaste K., Keskinen K.L., Rodriguez F.A., Jürimäe T. (2010) Physiological, biomechanical and anthropometrical predictors of sprint swimming performance in adolescent swimmers. Journal of Sports Science and Medicine 9, 398-404 [PMC free article] [PubMed] [Google Scholar]
- Lichtwarh G.A., Wilson A.M. (2008) Optimal muscle fascicle length and tendon stiffness for maximizing gastrocnemius efficiency during human walking and running. Journal of Theoretical Biology 252, 662-673 [DOI] [PubMed] [Google Scholar]
- Matta T., Simão R., de Salles B.F., Spineti J., Oliveira L.F. (2011) Strength training's chronic effects on muscle architecture parameters of different arm sites. The Journal of Strength & Conditioning Research 25, 1711-1717 [DOI] [PubMed] [Google Scholar]
- McLeod I. (2010) Swimming anatomy. Human Kinetics, USA [Google Scholar]
- Nasirzade A., Ehsanbakhsh A., Argavani H., Sobhkhiz A., Aliakbari M. (2013) Selected anthropometrical, muscular architecture, and biomechanical variables as predictors of 50-m performance of front crawl swimming in young male swimmers. Science & Sport, In Press [Google Scholar]
- Nygaard E., Nielsen E. (1978) Skeletal muscle fiber capillarisation with extreme endurance training in man. : Swimming Medicine IV. : Eriksson B., Furberg B. University Park Press, Baltimore: Vol 6, 282-293 [Google Scholar]
- Persyn U., De Maeyer J., Vervaeke H. (1975) Investigation of hydrodynamic determinants of competitive swimming strokes. International Series on Sport Sciences 2, 214-222 [Google Scholar]
- Reeves N.D., Narici M.V. (2003) Behavior of human muscle fascicles during shortening and lengthening contractions in vivo. Journal of Applied Physiology 95, 1090-1096 [DOI] [PubMed] [Google Scholar]
- Sacks R.D., Roy R.R. (1982) Architecture of the hind limb of muscle of cats: functional significance. Journal of Morphology 173, 185-195 [DOI] [PubMed] [Google Scholar]
- Schluter J.M., Fitts R.H. (1994) Shortening velocity and ATPase activity of rat skeletal muscle fibers: effects of endurance exercise training. American Journal of Physiology 266, 1699-1713 [DOI] [PubMed] [Google Scholar]
- Sharp R.L., Troup J.P., Costill D.L. (1982) Relationship between power and sprint freestyle swimming. Medicine & Science in Sports & Exercise 14, 53-56 [DOI] [PubMed] [Google Scholar]
- Simonsen E.B., Klitgaard H., Bojsen-Moller F. (1995) The influence of strength training, swim training and aging on the Achilles tendon and m. soleus of the rat. Journal of Sports Science 13, 291–295 [DOI] [PubMed] [Google Scholar]
- Sortwell A. (2010) Strength and power training for 100m front crawl swimmers. Journal of the International Society of Swimming Coach 2, 4-29 [Google Scholar]
- Spector S.A., Gardiner P.F., Zernicke R.F., Roy R.R., Edgerton V.R. (1980) Muscle architecture and the force-velocity characteristics of cat soleus and medial gastrocnemius: implications for motor control. Journal of Neurophysiology 44, 951-960 [DOI] [PubMed] [Google Scholar]
- Stager J.M., Tanner D.A. (2005) Handbook of Sports Medicine and Science: Swimming. 2th edition Blackwell Publishing, United Kingdom [Google Scholar]
- Stewart A., Marfell-Jones M., Olds T., de Ridder H. (2011) International standards for anthropometric assessment. ISAK: Lower Hutt, New Zealand [Google Scholar]
- Thom J.M., Morse C.I., Birch K.M., Narici M.V. (2007) Influence of muscle architecture on the torque and power-velocity characteristics of young and elderly men. European Journal of Applied Physiology 100, 613-619 [DOI] [PubMed] [Google Scholar]
- Van Praagh E. (2000) Development of anaerobic function during childhood and adolescence. Pediatric Exercise Science 12, 150–173 [Google Scholar]
- Vitor F.M., Bohme M.T. (2010) Performance of young male swimmers in the 100-meters front crawl. Pediatric Exercise Science 22, 278-287 [DOI] [PubMed] [Google Scholar]
- Weir J.P. (2005) Quantifying test-retest reliability using the intraclass correlation coefficient and the SEM. The Journal of Strength & Conditioning Research 19, 231-240 [DOI] [PubMed] [Google Scholar]