

Abstract
Remote sensing using airborne imaging spectroscopy (AIS) is known to retrieve fundamental optical properties of ecosystems. However, the value of these properties for predicting plant species distribution remains unclear. Here, we assess whether such data can add value to topographic variables for predicting plant distributions in French and Swiss alpine grasslands. We fitted statistical models with high spectral and spatial resolution reflectance data and tested four optical indices sensitive to leaf chlorophyll content, leaf water content and leaf area index. We found moderate added-value of AIS data for predicting alpine plant species distribution. Contrary to expectations, differences between species distribution models (SDMs) were not linked to their local abundance or phylogenetic/functional similarity. Moreover, spectral signatures of species were found to be partly site-specific. We discuss current limits of AIS-based SDMs, highlighting issues of scale and informational content of AIS data.
Keywords: species distribution, reflectance, hyperspectral data, alpine grasslands
1. Introduction
Spatial modelling of species distributions is commonly used to forecast environmental change effects, detect biodiversity hotspots or predict species' invasions [1]. As fine-grained environmental descriptors are difficult to obtain, coarse-grained (from hundreds of metres to kilometres) topo-climatic descriptors are usually used. Recent advances in airborne imaging spectroscopy (AIS) have allowed the acquisition of images with high spectral and sub-metre spatial resolution [2]. Spectral information provided by remotely sensed reflectance is influenced by phenology, variations in morphological, structural and biochemical properties of species [3], as well as by local environmental conditions (e.g. hydric stress, soil properties or productivity [4,5]) that determine species habitat suitability [6]. Nevertheless, previous attempts to predict species distributions with hyperspectral data have generated mixed results [7,8]. Sub-metre resolution allows the targeting of small plants and microhabitats where species find refuge, highlighting potential benefits of hyperspatial remote sensing for biodiversity monitoring [9]. However, despite increased spatial and spectral resolution of airborne data, little is known about its value in modelling species' distributions in species-rich ecosystems characterized by fine-scale heterogeneity.
Here, we explore the predictive power of AIS data for modelling plant species distributions in alpine grasslands in two distinct regions. Specifically, we aim to (i) identify key remotely sensed spectral information for predicting the distribution of grassland species and (ii) assess whether AIS data substantially improve model predictions. We also test for any phylogenetic or functional dependency of model characteristics among species.
2. Material and methods
(a). Study sites and species data
The study was conducted in the Western French (FR) and Western Swiss (CH) Alps (electronic supplementary material, S1). The French site included 103 vegetation plots of 2–5 m in radius, located between 2000 and 2830 metres above sea level (m.a.s.l.). The Swiss site included 68 quadrats (2 × 2 m) located between 1650 and 2150 m.a.s.l. Species cover was visually estimated using the Braun–Blanquet abundance scale. In total, 160 species were selected for species distribution analysis (119 species in FR, 78 in CH). Thirty-seven species were common to both sites (see electronic supplementary material, S1 for the details on selection criteria).
(b). Remote sensing data
AIS data were acquired with the dual Airborne Imaging Spectroradiometer for Applications (AISA; Specim Ltd., Finland). Raw AISA images contained 359 spectral bands between 400 and 2450 nm with spectral resolution ranging from 4.3 to 6.3 nm, and a pixel size of 0.8 m. After image processing, we extracted two types of AIS predictors: (i) reflectance in 75 spectral bands (avoiding bands with noisy radiometric response) and (ii) four vegetation indices. Vegetation indices characterized leaf chlorophyll (TCARI/OSAVI and ANCB) [10], leaf water content (SIWSI) [11] and leaf area index (MTVI2) [12] (for details see the electronic supplementary material, S1). Removal of poorly vegetated plots resulted in datasets with 70 FR and 53 CH plots.
(c). Topographic predictors
We computed five predictors derived from digital elevation models at 50 m resolution for FR and 25 m resolution for CH, representing mesoscale habitat conditions: (i) elevation (metre), (ii) slope (degree), (iii) aspect (degree), (iv) topographic position index (unitless) and (v) topographic wetness index (unitless; see the electronic supplementary material, S1).
(d). Species distribution modelling
Species distribution models (SDMs) were fitted with five different sets of variables: (i) topographic predictors only, (ii) reflectance predictors only, (iii) vegetation indices only, (iv) topographic and reflectance predictors combined and (v) topographic predictors and vegetation indices combined. We first used a conditional random forest algorithm to estimate the unbiased relative importance of predictors in the case of multicollinearity, then ran final models based on selection of the most important predictors [13] (see electronic supplementary material, S1). Their predictive accuracy was evaluated within each study site separately using a repeated split-sample procedure (100 iterations). Seventy per cent of the sample points were used for model calibration and 30% for model evaluation in each iteration.
(e). Model differences among species
The relative importance of AIS predictors and the predictive accuracy of SDMs was tested against (i) species' phylogenetic relatedness, (ii) species' functional similarity, including a set of morphological and physiological traits that are well correlated with the reflectance of canopy stands [14] (see electronic supplementary material, S2 and §5), and (iii) species' abundance patterns within plots. Phylogenetic and functional tests were computed as described in [15] (see the electronic supplementary material, S2 and §5).
3. Results
When fitting SDMs with reflectance data, the analysis of predictor importance indicated similarities in the selected spectral bands among sites (figure 1). The most important spectral bands were located between 500 and 900 nm for both sites, but site-specific differences in important spectral bands were also apparent (1500–1800 nm in FR, 1200–1500 nm and 2000–2500 nm in CH). These site differences existed for species present at only one or both sites (electronic supplementary material, S2 and figure S1). On average, all vegetation indices showed similar importance for SDM fitting (electronic supplementary material, S2 and figure S2).
Figure 1.
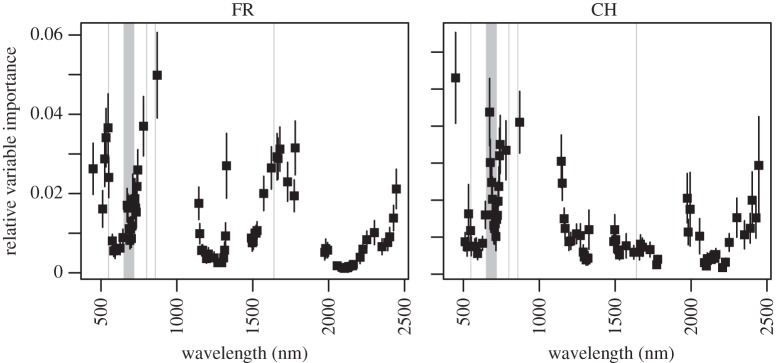
Relative importance of reflectance intensity in spectral bands for predicting species distributions at study sites in France (FR) and Switzerland (CH). Variable importance was assessed using conditional inference in random forest models. Grey areas represent bands used for the calculation of vegetation indices.
The prediction accuracy of SDMs based solely on topographic predictors, reflectance data or vegetation indices did not differ significantly. However, SDMs including both AIS and topographic predictors tended to be more accurate (figure 2 and the electronic supplementary material, S2 and table S1). The improvement was marginally significant for vegetation indices (Wilcoxon rank-sum test, p = 0.079) but non-significant for reflectance in FR. Conversely, CH showed significant improvement when using reflectance (Wilcoxon rank-sum test, p = 0.012), but non-significant effects when using vegetation indices. Improvements when including AIS predictors differed among species, with few species showing greater than or equal to 10% improved predictions and many showing reduced predictive accuracy (electronic supplementary material, S2 and figure S3). These variations were independent of species' abundance patterns and species' phylogenetic or functional similarity (electronic supplementary material, S2 and figures S4–S13).
Figure 2.
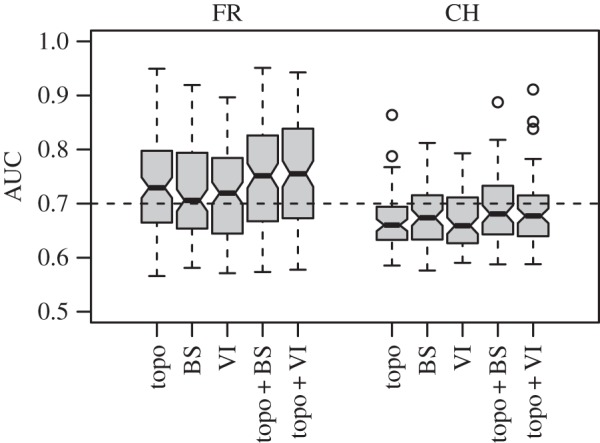
Prediction accuracy of species distribution models (based on the area under the curve of a receiver-operating characteristic plot: AUC) built with random forest models at study sites in France (FR) and Switzerland (CH). Topo indicates topographic predictors, BS indicates reflectance recorded in the spectral bands and VI indicates vegetation indices.
4. Discussion
Overall, topographic- and AIS-based SDMs revealed similar predictive accuracies in both sites. Model accuracy was on average higher in FR than in CH, whereas the topographical and spectral ranges observed in CH were much narrower than in FR (electronic supplementary material, S1 and figures S2, S4, S5). This agrees with previous studies where accuracy of SDMs derived from satellite images increased with steepness of ecological gradients [6]. Unlike vegetation indices, we found that importance of spectral bands differed between sites. Site-specific differences may partly reflect canopy differences owing to nutrient status or soil chemistry, because reflectance in these spectral regions is sensitive to light absorption by water [12], biochemical constituents [14] and scattering by plant architecture [11]. Additional field measurements of vegetation properties could probably improve ecological understanding of these spectral regions in SDMs.
The distribution models fit differed between species. Overall, models including both topographic and AIS predictors tended to be more accurate, even though significant improvements were confined to a limited number of species. This contrasts with results reported for invasive weeds [8], but agrees with results from meadows [7] where plant assemblages are inextricably mixed at the fine scale. Benefits of high spatial resolution of remote sensing data are a subject of debate [16]. Although our methodology considers the existence of geometric misalignment between AIS images and plot georeferencing, it still represents a source of uncertainty for matching reflectance of small pixels with local species occurrence. The significance of this uncertainty for SDM remains to be assessed.
We expected that differences between species models in terms of predictive accuracy and relative importance of AIS predictors would be linked to (i) abundance of species within-plots, because locally dominant species contribute more to canopy reflectance and (ii) phylogenetic or functional similarity, assuming that similar species show either comparable spectral signatures or similar habitat requirements as reflected by AIS data. These hypotheses were not supported. We suggest two possible explanations for such idiosyncrasy. First, accurate estimation of species' similarity may be limited by uncertainties in phylogenetic trait conservatism or availability of plant functional trait data. Phylogenies can often contribute to the integrated comparison of plant functional and life-history traits among species. However, the evolution of traits is characterized by both conservatism and diversification, and close links between functional similarity and phylogenetic relatedness are not always found [17]. In this study, we described species' functional similarity using morphological and ecophysiological traits that are recognized as key canopy reflectance drivers [14]. However, biochemical traits such as leaf nitrogen, chlorophyll or phosphorus content were not available for all species, and should be included wherever possible. Second, AIS-based SDMs may reflect both species' spectral signature and microhabitat suitability [18] (contrary to topography-based models which reflect solely habitat suitability at mesoscales). These two factors may differ in importance when fitting AIS variables across species and sites. This would explain why AIS-based models of both locally dominant (species detection scenario, e.g. Dryas octopetalla), and low-abundance species (habitat suitability scenario, e.g. Helictotrichon sedense) show equivalent accuracy despite very different species contributions to canopy characteristics and functional traits. Future research should focus on discriminating between species detection and habitat suitability for an array of species and ecosystem types (of varying degree of vegetation complexity), to better assess the ecological relevance of imaging spectroscopy for species' distribution modelling.
Supplementary Material
Supplementary Material
Acknowledgements
We are grateful to all persons supporting data collection. We also thank J.M.G. Bloor and four anonymous referees for helpful comments on previous version of the manuscript.
Data accessibility
Data available from the Dryad digital repository: doi:10.5061/dryad.n13hn.
Funding statement
This study was initiated and supported by the European ECOCHANGE project (GOCE-CT-2007-036866) and the Swiss National Science Foundation (BIOASSEMBLE, 31003A-125145). W.T. received funding from the ERC (EC FP7, 281422 (TEEMBIO)), M.S. from UZH URPP ‘Global Change and Biodiversity’. Computations were performed at the HPC Vital-IT Centre (Swiss Institute of Bioinformatics). Logistic support was provided by the ‘Station Alpine Joseph Fourier’ in France.
References
- 1.Guisan A, Thuiller W. 2005. Predicting species distribution: offering more than simple habitat models. Ecol. Lett. 8, 993–1009. ( 10.1111/j.1461-0248.2005.00792.x) [DOI] [PubMed] [Google Scholar]
- 2.Schaepman ME, Ustin SL, Plaza AJ, Painter TH, Verrelst J, Liang S. 2009. Earth system science related imaging spectroscopy: an assessment. Remote Sens. Environ. 113, S123–S137. ( 10.1016/j.rse.2009.03.001) [DOI] [Google Scholar]
- 3.Ustin SL, Gamon JA. 2010. Remote sensing of plant functional types. New Phytol. 186, 795–816. ( 10.1111/j.1469-8137.2010.03284.x) [DOI] [PubMed] [Google Scholar]
- 4.Schmidtlein S. 2005. Imaging spectroscopy as a tool for mapping Ellenberg indicator values. J. Appl. Ecol. 42, 966–974. ( 10.1111/j.1365-2664.2005.01064.x) [DOI] [Google Scholar]
- 5.Parviainen M, Luoto M, Heikkinen RK. 2010. NDVI-based productivity and heterogeneity as indicators of plant-species richness in boreal landscapes. Boreal Environ. Res. 15, 301–318. [Google Scholar]
- 6.Feilhauer H, He KS, Rocchini D. 2012. Modeling species distribution using niche-based proxies derived from composite bioclimatic variables and MODIS NDVI. Remote Sens. 4, 2057–2075. ( 10.3390/rs4072057) [DOI] [Google Scholar]
- 7.Schmidtlein S, Sassin J. 2004. Mapping of continuous floristic gradients in grasslands using hyperspectral imagery. Remote Sens. Environ. 92, 126–138. ( 10.1016/j.rse.2004.05.004) [DOI] [Google Scholar]
- 8.Lawrence RL, Wood SD, Sheley RL. 2006. Mapping invasive plants using hyperspectral imagery and Breiman Cutler classifications (randomForest). Remote Sens. Environ. 100, 356–362. ( 10.1016/j.rse.2005.10.014) [DOI] [Google Scholar]
- 9.Rocchini D. 2007. Effects of spatial and spectral resolution in estimating ecosystem α-diversity by satellite imagery. Remote Sens. Environ. 111, 423–434. ( 10.1016/j.rse.2007.03.018) [DOI] [Google Scholar]
- 10.Malenovský Z, Homolová L, Zurita-Milla R, Lukeš P, Kaplan V, Hanuš J, Gastellu-Etchegorry J-P, Schaepman ME. 2013. Retrieval of spruce leaf chlorophyll content from airborne image data using continuum removal and radiative transfer. Remote Sens. Environ. 131, 85–102. ( 10.1016/j.rse.2012.12.015) [DOI] [Google Scholar]
- 11.Haboudane D, Millera JR, Pattey E, Zarco-Tejada PJ, Strachan IB. 2004. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: modeling and validation in the context of precision agriculture. Remote Sens. Environ. 90, 337–352. ( 10.1016/j.rse.2003.12.013) [DOI] [Google Scholar]
- 12.Cheng Y-B, Zarco-Tejada PJ, Riaño D, Rueda CA, Ustin SL. 2006. Estimating vegetation water content with hyperspectral data for different canopy scenarios: relationships between AVIRIS and MODIS indexes. Remote Sens. Environ. 105, 354–366. ( 10.1016/j.rse.2006.07.005) [DOI] [Google Scholar]
- 13.Strobl C, Boulesteix A-L, Kneib T, Augustin T, Zeileis A. 2008. Conditional variable importance for random forests. BMC Bioinformatics 9, 307 ( 10.1186/1471-2105-9-307) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Homolová L, Malenovský Z, Clevers JGPW, García-Santos G, Schaepman ME. 2013. Review of optical-based remote sensing for plant trait mapping. Ecol. Complex. 15, 1–16. ( 10.1016/j.ecocom.2013.06.003) [DOI] [Google Scholar]
- 15.Hardy OJ, Pavoine S. 2012. Assessing phylogenetic signal with measurement error: a comparison of Mantel tests, Blomberg et al.'s K, and phylogenetic distograms. Evolution 66, 2614–2621. ( 10.1111/j.1558-5646.2012.01623.x) [DOI] [PubMed] [Google Scholar]
- 16.Nagendra H, Rocchini D. 2008. High resolution satellite imagery for tropical biodiversity studies: the devil is in the detail. Biodivers. Conserv. 17, 3431–3442. ( 10.1007/s10531-008-9479-0) [DOI] [Google Scholar]
- 17.Ackerly DD. 2009. Conservatism and diversification of plant functional traits: evolutionary rates versus phylogenetic signal. Proc. Natl Acad. Sci. USA 106, 19 699–19 706. ( 10.1073/pnas.0901635106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bradley BA, Olsson AD, Wang O, Dickson BG, Pelech L, Sesnie SE, Zachmann LJ. 2012. Species detection vs. habitat suitability: are we biasing habitat suitability models with remotely sensed data? Ecol. Model. 244, 57–64. ( 10.1016/j.ecolmodel.2012.06.019) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data available from the Dryad digital repository: doi:10.5061/dryad.n13hn.