

Abstract
In previous studies examining the role of glucocorticoids and thyroid hormone on the maturation of the Na+/H+ antiporter (NHE3), we found attenuation in the maturational increase in proximal tubule apical Na+/H+ antiporter activity but no change in NHE3 mRNA abundance in either glucocorticoid-deficient or hypothyroid rats. In addition, prevention of the maturational increase in either hormone failed to totally prevent the maturational increase in Na+/H+ antiporter activity. We hypothesized that one hormone played a compensatory role when the other was deficient. The present study examined whether combined deficiency of thyroid and glucocorticoid hormones would completely prevent the maturation of the Na+/H+ antiporter. Adrenalectomy was performed in 9-day-old hypothyroid Sprague-Dawley rats, a time before the normal postnatal maturational increase in these hormones occurs. Nine- and 30-day-old adrenalectomized (ADX), hypothyroid rats had comparable NHE3 mRNA abundance, which was 5- to 10-fold less than 30-day-old ADX, hypothyroid rats that received corticosterone-thyroxine replacement and 30-day-old sham control rats (P < 0.05). Brush-border membrane NHE3 protein abundance was comparable in 9- and 30-day-old ADX, hypothyroid groups and ~20-fold lower than both the 30-day replacement and 30-day sham groups (P < 0.05). Similarly, the replacement and sham groups had higher sodium-dependent proton secretion than 9- and 30-day-old ADX, hypothyroid groups (P < 0.05). We conclude that combined deficiency of both hormones totally prevents the maturational increase in NHE3 mRNA and protein abundance and Na+/H+ antiporter activity.
Keywords: NHE3, adrenalectomy, renal development, microperfusion, cell pH
Neonates have a lower bicarbonate threshold than adults (21), which is due in large part to immaturity of neonatal proximal tubule acidification (10, 11, 35). There is a fourfold increase in rat apical membrane proximal convoluted tubule Na+/H+ antiporter activity during postnatal development (36). The factors that mediate the postnatal maturational increase in Na+/H+ antiporter activity have been elusive.
The levels of both glucocorticoids and thyroid hormone are lower in the first 1–2 wk of life compared with those of adults (24, 39). The maturational increase in these hormones parallels the maturational increase in NHE3 (36), the predominant Na+/H+ antiporter isoform on the apical membrane of the proximal tubule (15, 40, 41), and Na+/H+ antiporter activity (36). We examined previously the role of glucocorticoids and thyroid hormone in mediating the increase in Na+/H+ antiporter activity (8, 23). In these studies, we either prevented the maturational increase in glucocorticoids or thyroid hormone at a time well before the normal increase during postnatal development. We found that preventing the maturational increase in either thyroid hormone or glucocorticoids suppressed the maturational increase in Na+/H+ antiporter activity and brush-border membrane NHE3 protein abundance without any effect on the basal level of NHE3 mRNA abundance (8, 23). Furthermore, the rate of Na+/H+ antiporter activity in both adrenalectomized and hypothyroid animals was higher than in neonates. These results could be explained if thyroid hormone and glucocorticoids played a compensatory role in the face of deficiency of the other hormone.
The present study was designed to examine the maturation of Na+/H+ antiporter, brush-border membrane vesicle (BBMV) NHE3 protein and renal cortical mRNA abundance in the presence of a combined deficiency of glucocorticoid and thyroid hormones. This is the first study to use a unique animal model of adrenalectomized, hypothyroid neonatal rats. Our results using this model demonstrate that postnatal maturation of Na+/H+ antiporter activity, NHE3 protein, and RNA abundance is completely prevented using this novel model. Furthermore, this model will be useful in the study of other proteins mediated by both thyroid hormone and glucocorticoids.
METHODS
Animals
Pregnant Sprague-Dawley rats were received on day 13 of gestation; 0.01% n-propyl-thiouracil (PTU) drinking water was used to induce hypothyroidism starting from day 14 of gestation through the entire postnatal period (8). The number of pups in each litter was reduced to nine just after birth. The pups stayed with the mother until the day of death. Hypothyroid pups underwent bilateral adrenalectomy at 9 days of age, a time well before the postnatal increase in glucocorticoid levels (24). Control neonates from untreated Sprague-Dawley rats underwent sham operation at 9 days of age. A bilateral adrenalectomy was performed using two lumbodorsal incisions under light ether anesthesia as previously described (23). All hypothyroid pups destined for an adrenalectomy received 100 μg of the long-acting mineralocorticoid deoxycorticosterone pivalate (Percorten-V, Novartis Animal Health, Greensboro, NC) in a 0.9% saline suspension subcutaneously before surgery. Supplemental injections of 150 and 200 μg were administered on days 16 and 23, respectively (23). In preliminary experiments, we found that all hypothyroid neonates died shortly after an adrenalectomy. However, postoperative death was not seen if the pups received 0.5 ml of 0.9% saline subcutaneously just before surgery and immediately after surgery. Sham-operated rats received an equal volume of 0.9% saline injections. The adrenalectomized (ADX)-hypothyroid rats were divided into a corticosterone (cort) and thyroxine (T4) replacement group and vehicle-treated (ADX-hypothyroid) group. The replacement group received a daily subcutaneous injection of corticosterone (8 μg/g body wt) and thyroxine (0.05 μg/g body wt in 0.3 mM NaOH) from day 14 onward until the rats were killed at day 30 of age (12, 17, 19, 20). The ADX-hypothyroid group was injected with an equivalent volume of vehicle. All sham-operated rats received comparable vehicle injections. Animals were weighed on day 30 before death. Corticosterone and thyroxine levels were measured in serum to ensure complete adrenalectomy and hypothyroidism.
This study was approved by the IACRAC at the University of Texas Southwestern Medical Center and adhered to APS’s Guiding Principles in the Care and Use of Animals.
RNA isolation and analysis
Slices of renal cortex from decapsulated kidneys were homogenized in RNAzol [1:1, phenol-RNAzol stock (4 M guanidine thiocyanate, 25 mM disodium-citrate, pH 7.0), 0.5% sarcosyl] containing 3.6 μl/ml β-mercaptoethanol. RNA was extracted in the presence of 3 M NaOAc (pH 4.0) and chloroform, purified using isopropanol precipitation, and washed twice with 80% ethanol (16). RNA was quantitated with a LKB Ultra-spec III spectrophotometer at 260 nm, and 15 μg were fractionated using agarose-formaldehyde gel electrophoresis and transferred to a nylon filter (GeneScreen Plus, New England Nuclear, Boston, MA). The filter was prehybridized at 42°C for 4 h with 5× SSC, 5× Denhardt’s solution (Ficoll, BSA, and polyvinylpyrrolidone, each at 1 mg/ml), 0.5% SDS, and 0.5 mg/ml of sheared salmon sperm DNA and then hybridized to double-strand uniformly 32P-labeled cDNA probes (>106 cpm/ml) in the above hybridization solution at 42°C for ~16 h. The probes were synthesized by the random hexamer method using 50 to 100 ng of cDNA: NHE3 was the rat 1.2-kb PstI fragment (33) and β-actin was a 1.5-kb EcoRI fragment. The filter was then washed with 2× SSC twice and 0.1% SDS for 5 min at room temperature and then twice at 55°C for 40 min with 0.1× SSC and 1% SDS. NHE3 and β-actin mRNA abundance was quantitated with autoradiography and densitometry.
BBMV isolation
Kidneys were removed and placed in an ice-cold isolation buffer containing 300 mM mannitol, 16 mM HEPES, and 5 mM EGTA titrated to pH 7.4 with Tris. The isolation buffer contained aprotinin (2 μg/ml), leupeptin (2 μg/ml), and phenylmethylsulfonyl fluoride (100 μg/ml). The cortex was homogenized with 20 strokes of a Potter Eljevhem homogenizer at 4°C. BBMV were then isolated by differential centrifugation and magnesium precipitation as described previously (23, 36). The final BBMV fraction was resuspended in isolation buffer. Protein was assayed using the Lowry method with crystalline BSA as the standard (30). There was comparable enrichment of leucine amino peptidase in BBMV from neonatal 10.0 ± 1.6 compared with 13.4 ± 1.2 in 30-day-old rats [n = 7, P = not significant (NS)].
SDS-PAGE and immunoblotting
Brush-border membrane proteins (40 μg/lane) were denatured and then separated on a 7.5% polyacrylamide gel using SDS-PAGE as previously described (23, 36). The proteins were transferred overnight to a polyvinylidene diflouride membrane at 120–140 mA at 4°C. The blot was blocked with fresh Blotto (5% nonfat milk and 0.1% Tween 20 in PBS, pH 7.4) for 1 h followed by incubation with primary antibody to NHE3. NHE3 antibody, a gift from Dr. O. Moe, was a rabbit polyclonal antibody directed against a fusion protein of maltose-binding protein and rat NHE3 amino acids 405–831 (2). NHE3 antibody was added at 1:750 dilution and incubated for 16 h at 4°C. The blot was then washed extensively with Blotto. The secondary antibody, horseradish peroxidase-conjugated donkey anti-rabbit immunoglobulin, was added at 1:10,000 dilution and incubated in room temperature for 1 h. The blot was again washed with Blotto, and enhanced chemiluminescence was used to detect bound antibody (Amersham Life Science). The NHE3 protein abundance was quantitated using densitometry. Equal loading of the samples was confirmed using an antibody to β-actin at a 1:5,000 dilution (Sigma, St. Louis, MO).
In vitro microperfusion and measurement of intracellular pH
We recently demonstrated that 0.2- to 0.5-mm neonatal and young rat proximal convoluted tubules can be dissected free hand without collagenase and perfused in vitro (23, 36). Isolated segments of neonatal and adult rat proximal convoluted tubules were perfused using concentric glass pipettes using techniques previously described for rabbit proximal tubules (4, 5, 11). Briefly, rat proximal convoluted tubules were dissected from 9-day-old, 30-day-old sham, 30-day-old ADX, 30-day-old ADX-hypothyroid, and 30-day replacement group in Hanks’ balanced salt solution containing (in mM) 137 NaCl, 5 KCl, 0.8 MgSO4, 0.33 Na2HPO4, 0.44 KH2PO4, 1 MgCl2, 10 Tris hydrochloride, 0.25 CaCl2, 2 glutamine, 2 heptanoic acid, and 2 lactate at 4°C (pH 7.4). Tubules were transferred to a 0.2-ml chamber in which the bathing solution was preheated to 38°C.
The composition of the solutions used in these experiments is shown in Table 1. The fluorescent dye BCECF was used to determine intracellular pH (pHi) as described previously (1, 4, 5, 33, 37). pHi was measured with a Nikon inverted epifluorescent microscope attached to a PTI Ratiomaster at a rate of 30 measurements per second. A variable diaphragm was placed over the area to be measured. pHi was determined from the ratio of fluorescence (F500/F450) using a nigericin calibration curve as previously described (1, 4, 34).
Table 1.
Solutions used in pHi studies
| Bath | Luminal Na+ | Luminal 0Na+ | 0Na 0Cl− 0NH4+ | 0Na 0Cl +20 mM | |
|---|---|---|---|---|---|
| Solution A | Solution B | Solution C | Solution D | Solution E | |
| NaCl | 140 | 115 | |||
| NaHCO3 | 5 | 25 | |||
| TMA-OH | 115 | 95 | |||
| Gluconic acid lactone | 115 | 115 | |||
| NMDG-Cl | 115 | ||||
| Choline HCO3 | 25 | 25 | 25 | ||
| NH4OH | 20 | ||||
| KCl | 5 | ||||
| K2HPO4 | 2.5 | 2.5 | 2.5 | 2.5 | |
| MgCl2 | 1 | 1 | |||
| MgSO4 | 1 | ||||
| Na2HPO4 | 1 | ||||
| Glucose | 5 | 5 | 5 | ||
| L-Alanine | 5 | 5 | 5 | ||
| Urea | 5 | ||||
| Mg gluconate | 1 | 1 | |||
| CaCl2 | 1.8 | 1.8 | 1.8 | ||
| Ca gluconate | 12.5 | 12.5 | |||
| Heptanoic acid | 2 | ||||
| pH | 6.6 | 7.4 | 7.4 | 7.4 | 7.4 |
All constituents are in mM. All solutions were adjusted to an osmolality of 295 mosmol/kgH2O and were bubbled with 5% CO2-95% O2. pHi, intracellular pH; TMA, tetramethyl ammonium; NMDG, N-methyl-D-glucamine.
Measurement of proton flux rates
Proton flux rates (JH; in pmol · mm−1 · min−1) resulting from a luminal fluid change were calculated using the following formula JH = dpHi/dt · V/mm · β, where dpHi/dt was the rate of initial change in pHi after a luminal fluid change, V is the tubular volume in liters per millimeter, and β is the buffer capacity. Tubular volume was calculated from the measured inner and outer tubular diameters at ×400 magnification using an eyepiece reticle. Tubular volumes of proximal convoluted tubules from 9-day-old, 30-day-old ADX, 30-day-old ADX-hypothyroid, replacement and 30-day sham rats were 637 ± 42, 793 ± 52, 567 ± 51, 1,361 ± 114, and 1,281 ± 115 × 10−12 l/mm, respectively. The tubular volumes of the 9- and 30-day-old ADX and ADX-hypothyroid groups were smaller that the ADX-Cort and 30-day-old sham (P < 0.05). Apparent buffer capacity (β) was measured previously using (36). Solutions (D and E) were used for measurement of apparent buffer capacity. These solutions did not contain Na+ or Cl− so as to inhibit all acidification mechanisms due to Na+- and Cl−-dependent transporters. In the presence of 25 mM , the buffer capacity was 98.7 ± 16.4 mM/pH in neonatal PCT and 98.9 ± 17.6 mM/pH in adult proximal convoluted tubules (PCT; P = NS) (36) and a value of 99 as the buffer capacity for all groups in this study.
Tubules were incubated with an ultrafiltrate-like solution in the lumen and bath (solution B containing 5 mM glucose and 5 mM alanine) for at least 5 min before being loaded with 5 × 10−6 M BCECF, and tubules had a constant pHi for several minutes before the measurement of transporter activity. dpHi/dt was measured from the slope of change in pHi immediately after a luminal fluid change. Steady-state pHi values were reached within 1 min after a luminal fluid exchange, but pHi was measured for several minutes to ensure a steady-state pHi was achieved.
Na+/H+ antiporter activity was measured as previously described in adult rat PCT perfused in vivo and neonatal and adult rabbit and rat PCT perfused in vitro (1, 4, 11, 23, 36, 37). Neonatal and adult rat tubules were perfused with an ultrafiltrate-like solution without glucose and amino acids (solution B). Organic solutes were omitted from the luminal solution because sodium-coupled glucose and amino acid transport depolarize the basolateral membrane, which may affect bicarbonate exit, an electrogenic process (1). SITS (1 mM) was present in the bathing solution to inhibit the sodium bicarbonate cotransporter, a major regulator of pHi in proximal convoluted tubules (1, 4). The bathing solution had a bicarbonate concentration of 5 mM and a pH of 6.6 to compensate for the cell alkalinization caused by the addition of bath SITS (1, 4). The bathing solution was exchanged at a rate of at least 5 ml/min. Under these conditions, changes in pHi in response to a change in luminal sodium concentration are a measure of Na+/H+ antiporter activity (4, 15, 36, 37). In the experimental period, luminal sodium was removed (solution C).
Chemicals and RIA kits
Corticosterone, T4, and PTU were obtained from Sigma. Serum corticosterone levels were determined by using corticosterone 3H-RIA Kit (ICN Biomedicals, Costa Mesa, CA). Serum thyroxine was measured using 125I-labeled T4 RIA Kit (Diagnostic Systems Laboratories, Webster, TX).
Statistical analysis
Each experiment was performed at least four times. Data are expressed as means ± SE. Statistical significance was determined using analysis of variance, and statistical significance between the groups was performed using the Student-Newman-Keuls test.
RESULTS
Effect of adrenalectomy and hypothyroidism on serum concentration of corticosterone and thyroxine
As shown in Table 2, serum corticosterone levels were comparable in 9- and 30-day-old ADX-hypothyroid and significantly lower than in the replacement and sham-operated groups. The sham-operated group had a higher serum corticosterone level than any other group (P < 0.05); however, measurement of both corticosterone and thyroxine was performed in the replacement group 14 h after the last dose of replacement therapy. The serum thyroxine level was significantly less in the 30-day-old ADX-hypothyroid group than the 9-day-old rats (P < 0.05). The replacement and sham-operated groups had a higher level of thyroxine than both ADX-hypothyroid and 9-day-old rats (P < 0.05). Thus 0.01% PTU in drinking water and adrenalectomy prevented the maturational increase in serum thyroxine and corticosterone, respectively.
Table 2.
Weight of kidneys and of animals and serum corticosterone and thyroid hormone levels in rats
| 9-Day-Old Rats | 30-Day ADX-Hypothyroid Rats | 30-Day Replacement | 30-Day Sham | |
|---|---|---|---|---|
| Wt of right kidney, g | 0.08±0.01*† | 0.14±0.01† | 0.35±0.02 | 0.43±0.01‡ |
| n=9 | n=9 | n=10 | n=6 | |
| Wt of left kidney, g | 0.08±0.01*† | 0.13±0.01† | 0.36±0.02 | 0.44±0.01‡ |
| n=9 | n=9 | n=10 | n=6 | |
| Wt of rat, g | 23.06±0.75† | 25.75±0.69† | 58.02±2.06 | 89.71±1.70‡ |
| n=12 | n=13 | n=10 | n=11 | |
| Serum corticosterone, μg/dl | 0.36±0.14*† | 0.16±0.04† | 1.61±0.29 | 14.68±2.60‡ |
| n=11 | n=15 | n=13 | n=10 | |
| Serum T4, μg/dl | 2.38±0.16*† | 0.20±0.05† | 9.51±1.01 | 5.23±0.36‡ |
| n=11 | n=12 | n=12 | n=8 | |
| Serum bicarbonate, meq/l | 25.9±1.5 | 20.0±0.3§ | 26.8±2.3 | 27.8±0.7 |
| n=6 | n=5 | n=4 | n=6 |
Values are means ± SE; n = number of measurements.
Different from 30-day adrenalectomized (ADX) hypothyroid group at P < 0.05.
Different from replacement and sham at P < 0.05.
Different from replacement at P < 0.05.
Different from all groups at P < 0.01.
Effect of adrenalectomy and hypothyroidism on body and kidney weight and serum bicarbonate
As shown in Table 2, there was no significant difference in mean weights of 9- and 30-day-old ADX-hypothyroid groups, and those of both groups were less than 30-day-old replacement and sham control groups (P < 0.05). Replacement with corticosterone and T4 resulted in an increase in body weight, but the body weight in the replacement group was still less than that of the sham control group (P < 0.05). The mean weights of the right kidney and left kidney were less in the 9-day-old group than that of 30-day-old ADX-hypothyroid group, indicating that there was renal growth in the virtual absence of thyroid hormone and glucocorticoids (P < 0.05). Thus hypothyroid-ADX animals have a decreased rate of somatic and renal growth.
Serum bicarbonate was significantly less in the ADX-hypothyroid group than in the 9- and 30-day-old replacement and sham control groups. The bicarbonate level was likely similar in the 9- and 30-day-old replacement and sham control groups, because the 9-day-old rats were still nursing and rat milk contains significant quantities of bicarbonate (42). The serum bicarbonate level in the ADX-hypothyroid group was significantly less than in the 30-day-old replacement and sham control groups.
Effect of glucocorticoids and thyroid hormone on renal cortical NHE3 mRNA abundance
To determine the role of a combined deficiency of glucocorticoids and thyroid hormone on the maturation of NHE3 RNA, we measured renal cortical NHE3 RNA abundance in 9-day-old, 30-day-old ADX-hypothyroid, replacement, and sham control rats using Northern blot analysis. The results are shown in Fig. 1. The NHE3 mRNA-to-β-actin ratio in 9- day-old (0.62 ± 0.08) and in 30-day-old ADX-hypothyroid (1.44 ± 0.22) groups was comparable. Both of these groups had significantly lower NHE3 mRNA/β-actin ratios than that of the 30-day-old replacement (6.46 ± 1.14) group and the 30-day-old sham control (8.54 ± 1.26) groups (P < 0.05). There was no difference in the 30-day-old replacement group and sham-operated group. Thus prevention of a postnatal surge in glucocorticoid and thyroid hormones prevented the maturational increase in renal cortical NHE3 mRNA abundance.
Fig. 1.

Effect of glucocorticoids and thyroid hormones on NHE3 mRNA abundance. A: Northern blot with 15 μg of total RNA from renal cortex of rats of ages. B: β-actin mRNA was shown for comparison. NHE3 mRNA abundance was significantly less in the 9-day-old (9 d) and ADX-hypothyroid (adx-hypoT) groups than that of the other groups (P < 0.05). There were at least 4 measurements in each group. cort, Corticosterone; T4, thyroxine.
Effect of glucocorticoids and thyroid hormones on BBMV NHE3 protein abundance
Western blot analysis was used to quantify NHE3 protein abundance. As shown in Fig. 2, the NHE3 protein/β-actin ratio in the 9-day-old rats (0.03 ± 0.01) was not different from that of the 30-day-old ADX-hypothyroid group (0.06 ± 0.01). The 30-day-old replacement group (1.26 ± 0.16) and sham control group (0.90 ± 0.19) had comparable levels of NHE3 protein/β-actin ratios, and both had significantly higher levels than that of the 9- and 30-day-old ADX-hypothyroid groups (P < 0.05).
Fig. 2.
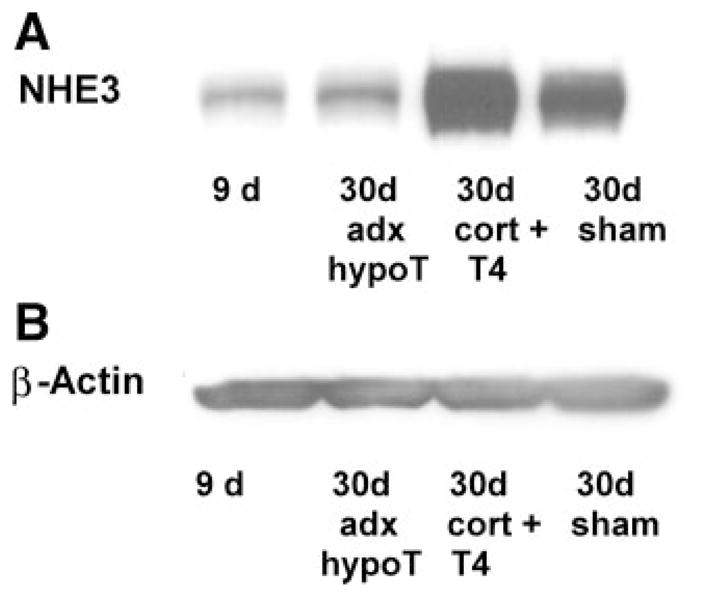
Effect of glucocorticoids and thyroid hormones on NHE3 protein abundance from renal brush-border membrane vesicles compared with β-actin. NHE3 protein abundance was comparable in the 9-day-old and ADX-hypothyroid groups and less than that in the 30-day-old replacement and the 30-day-old sham-operated group (P < 0.05). There were at least 4 measurements in each group.
Effect of glucocorticoids and thyroid hormone on Na+/H+ exchanger in rat PCT
The rates of sodium-dependent proton secretion (in pmol · mm−1 · min−1) in PCT are shown in Fig. 3. The steady-state pHi in the presence and absence of luminal sodium is shown in Table 3. Sodium-dependent proton secretion was significantly slower in 9-day-old (204 ± 21 pmol · mm−1 · min−1), 30-day-old ADX (339 ± 39 pmol · mm−1 · min−1), and 30-day-old ADX-hypothyroid (110 ± 24 pmol · mm−1 · min−1) groups than in 30-day-old replacement (718 ± 134 pmol · mm−1 · min−1) and sham-operated groups (607 ± 114 pmol · mm−1 · min−1; P < 0.05). The 30-day-old ADX group had a higher rate of sodium-dependent proton secretion than that of the 30-day-old ADX-hypothyroid group (P < 0.05). We previously found that adrenalectomy did not completely prevent the maturational increase in Na+/H+ antiporter activity and confirmed this here if one compares the 30-day-old ADX group vs. 9-day-old neonates using an unpaired Student’s t-test. However, maturation of the Na+/H+ antiporter was completely prevented in hypothyroid ADX animals.
Fig. 3.

Na+/H+ antiporter activity in rat proximal convoluted tubules perfused in vitro. Proton flux rate (JH) was measured after removal of luminal sodium. Na+/H+ antiporter activity in 30-day-old ADX-hypothyroid group was less than that of the 30-day-old ADX group (P < 0.05). Nine-day, 30-day ADX, and 30-day ADX-hypothyroid groups had less Na+/H+ antiporter activity than that of the 30-day-old replacement and the 30-day-old sham-operated group (P < 0.05). There were at least 5 measurements in each group.
Table 3.
Effect of luminal Na+ removal on pHi in rat PCT
| n | Na+ | 0 Na+ | Na+ | |
|---|---|---|---|---|
| 9-Day PCT | 7 | 7.35±0.04 | 7.12±0.06* | 7.39±0.04 |
| 30-Day ADX PCT | 6 | 7.20±0.07 | 6.89±0.06* | 7.25±0.08 |
| 30-Day ADX + Hypothyroid PCT | 10 | 7.24±0.05 | 6.97±0.07* | 7.27±0.05 |
| 30-Day ADX + Hypothyroid + Hormone Replacement PCT | 6 | 7.39±0.04 | 6.96±0.07* | 7.39±0.05 |
| 30-Day Sham PCT | 5 | 7.28±0.06 | 6.94±0.08* | 7.29±0.08 |
Values are means ± SE. PCT, proximal convoluted tubules.
P < 0.01 compared with sodium-containing solutions.
DISCUSSION
In the present study, we examined the effect of combined glucocorticoid and thyroid hormone deficiency as a novel model to examine the maturation of the proximal tubule Na+/H+ exchanger (NHE3). In this model, rats were made glucocorticoid and thyroid hormone deficient before the normal postnatal surge in serum glucocorticoid and thyroid hormone levels. We found that ADX-hypothyroid rats at 30 days of age had comparable renal cortical NHE3 mRNA abundance, brush-border membrane NHE3 protein abundance, and proximal tubule Na+/H+ antiporter activity as that of the 9-day-old neonatal rats. Replacement with physiological doses of corticosterone and thyroxine (replacement group) resulted in a comparable level of NHE3 mRNA and protein abundance and Na+/H+ antiporter activity as that of the sham control group.
The factors that induce the postnatal increase in proximal tubule acidification have been elusive. Prime candidates have been glucocorticoids and thyroid hormone, which increase ~25- and ~3-fold, respectively, during postnatal development (24, 39) and which are known to affect the Na+/H+ antiporter (NHE3) (8, 23). The promoter of rat NHE3 has been sequenced and characterized (13, 25). Analysis of 1.4 kb of the 5′-flanking promoter region showed multiple DNA sequence elements that are recognized by the glucocorticoid receptors, thyroid hormone receptors, SP1, AP-1, AP-2, and several other transcription factors that may participate in the regulation of the NHE3 gene (13, 25). OKP cells, a cell line with characteristics of proximal tubule, expresses NHE-3. We previously demonstrated that both dexamethasone and thyroid hormone increase NHE3 transcription using nuclear run-on assays (6, 14). Neither thyroid hormone nor dexamethasone affected NHE3 mRNA stability (6, 14). In addition, triiodothyronine did not affect rat NHE3 protein stability (14).
There is an interaction between serum thyroid hormone and glucocorticoid levels. Adrenalectomized rats have an increase in thyroid hormone levels compared with controls (32, 38). Plasma thyrotropin-releasing hormone, TSH, T4, and T3 were elevated a week following adrenalectomy (32) in one study; however, another found elevated T3 levels without any significant change in T4 level, 6 wk following adrenalectomy (38). In contrast, studies in neonatal hypothyroid rats have shown either normal (31) or subnormal basal levels of corticosterone (18), whereas hypothyroid adult rats have normal basal corticosterone levels (38) and they have a subnormal corticosterone response to stress (31).
We previously examined whether administration of dexamethasone would accelerate the maturation of proximal tubule acidification (10, 11). We found that the rate of juxtamedullary proximal convoluted tubule bicarbonate absorption and Na+/H+ antiporter activity in the first 2 days of life in rabbits were one-third that of the adult tubule (4, 10). Administration of 60 μg/kg of dexamethasone to a pregnant doe for 3 days before delivery resulted in an increase in the rates of bicarbonate absorption and Na+/H+ antiporter activity comparable to that in the adult proximal convoluted tubule (11). We also found that there is a fourfold maturational increase in rabbit NHE-3 mRNA and protein abundance during postnatal development (7). Administration of either 6 μg/100 g of dexamethasone to does starting at day 27 of gestation or 10 μg/100 g of dexamethasone to neonates resulted in levels of NHE3 mRNA abundance in neonatal rabbits that were comparable to that of adults (7). In the proximal tubule, NHE1 is located on the basolateral membrane. There was no maturational change in NHE1 mRNA or protein abundance, and neither administration of prenatal nor postnatal dexamethasone affected NHE1 mRNA or protein abundance (7).
We recently examined whether prevention of the postnatal increase in glucocorticoids would affect the maturation of PCT Na+/H+ antiporter activity, BBMV NHE3 protein abundance, and renal cortical NHE3 mRNA abundance (23). In that study, adrenalectomy at 9 days of age, a time before the postnatal increase in glucocorticoids, resulted in a significant attenuation of Na+/H+ exchanger activity at 30 days of age. Nonetheless, Na+/H+ antiporter activity was higher than that of the 9-day-old rats. Corticosterone replacement from day 14 of age in the ADX group restored PCT Na+/H+ antiporter activity. NHE3 mRNA abundance was fivefold lower in 9-day-old than that of 30-day-old control rats. Surprisingly, neonatal adrenalectomy did not prevent the maturational increase in NHE3 mRNA abundance. The 30-day-old ADX mRNA abundance was the same as that of ADX with corticosterone replacement and the sham control groups (23).
Similarly, we studied the role of thyroid hormone in the maturation of proximal tubule Na+/H+ antiporter activity (8). In this study, neonatal rats were made hypothyroid by administrating 0.01% PTU in drinking water from day 14 of gestation until the day of study at 21 days of age. Hyperthyroidism was induced by intraperitoneal injection of triiodothyronine on days 17 to 20 of postnatal life. Although there was a reduction in BBMV Na+/H+ antiporter activity in hypothyroid animals and an increase in BBMV Na+/H+ antiporter activity in hyperthyroid rats compared with euthyroid controls, the effect of thyroid hormone status on Na+/H+ antiporter activity was trivial. Although hyperthyroid rats showed a twofold increase in NHE3 mRNA abundance vs. euthyroid rats, hypothyroid rats had comparable NHE3 mRNA abundance as that of euthyroid rats (8). These studies taken together demonstrate that prevention of the maturational increase in either thyroid hormone or glucocorticoids alone does not affect the postnatal maturational increase in NHE3 mRNA. However, as shown in the present study, prevention of the maturational increase in glucocorticoids and thyroid hormone together prevents the maturational increase in NHE3 mRNA.
Studies have characterized the effect of both glucocorticoid excess and deficiency in adult rats (22, 26, 27, 29). The administration of dexamethasone in both adult and adrenalectomized adult rats stimulated the rate of Na+/H+ antiporter activity in BBMV without altering the affinity of the antiporter for Na+ or H+ (22, 27). Similarly, the increase in NHE3 protein abundance following dexamethasone administration was independent of whether adult rats were adrenalectomized 2 days before the study (29). Adrenalectomized adult rats had comparable Na+/H+ antiporter activity in BBMV as that of the sham control group (22, 26, 27). NHE3 protein abundance was also not found to be different in adrenalectomized and sham control groups (29). In agreement with these studies, use of aminoglutethimide in adult rabbits produced glucocorticoid deficiency but resulted in no change in the basal level of renal cortical NHE3 mRNA abundance (9). At variance with these studies, we found a significant attenuation in Na+/H+ antiporter activity as well as NHE3 protein abundance in BBMV in adrenalectomized neonatal rats. This discrepancy may be explained by the fact that adrenalectomy was performed at 9 days of age in our study, a time before the postnatal increase in glucocorticoid occurs in rats. The duration of glucocorticoid deficiency was also different in the two studies. We made measurements 21 days after adrenalectomy compared with 2–3 days after adrenalectomy in studies using adult rats described above. Both PCT Na+/H+ antiporter activity and NHE3 protein abundance in adrenalectomized neonatal rats were restored with administration of physiological doses of corticosterone.
Similar to our previous study of the effect of thyroid hormone on neonatal rats (8), the role of thyroid hormone status on Na+/H+ antiporter activity and NHE3 abundance has been studied in adult rats (3, 28). The thyroid hormone status was found to have a greater impact on Na+/H+ antiporter activity in adult rats than we found in our neonatal rats (8, 28). In agreement with our study, NHE3 mRNA abundance was comparable in both hypothyroid and euthyroid adult rats and was higher in hyperthyroid rats (3). However, this study in adult rats failed to show any difference in renal cortical NHE3 protein abundance between hypothyroid and hyperthyroid adult rats (3), in contrast to our finding of a low level of NHE3 protein in hypothyroid vs. hyperthyroid neonatal rats (8). The reason for this discrepancy is not clear but is likely due to the age of the rats.
Our previous study in neonatal adrenalectomized rats showed significant blunting (more than 60%) in postnatal maturation of Na+/H+ antiporter activity in a 30-day-old adrenalectomized group vs. a 30-day-old sham-operated group (23). Once again, we confirmed our previous findings in this experiment that deficiency of glucocorticoids did not totally prevent the maturation of Na+/H+ exchanger activity (23). Hypothyroid status showed a trivial (only 10%) reduction in Na+/H+ antiporter activity in BBMV (in hypothyroid rats) in contrast to euthyroid rats (8). These findings demonstrate that glucocorticoids have a more profound effect on maturation of Na+/H+ antiporter activity than the thyroid hormone. In the present study, Na+/H+ antiporter activity was comparable in the 30-day-old ADX-hypothyroid group and 9-day-old rats. However, the 30-day-old ADX group had a significantly higher level of Na+/H+ antiporter activity than that of the 30-day ADX-hypothyroid group. All these data suggest that the thyroid hormone contributes a small fraction to the maturation of Na+/H+ antiporter activity that persists after complete adrenalectomy.
In summary, the present study shows that maturation of Na+/H+ antiporter activity, BBM NHE3 protein, and renal cortical NHE3 mRNA abundance can be completely prevented by preventing ontogenic rise in glucocorticoid and thyroid hormone levels. Thus both glucocorticoid and thyroid hormones play a combined and interrelated role in maturation of proximal tubule acidification.
Acknowledgments
We thank Dr. R. Quigley for reviewing this manuscript.
GRANTS
This work was supported by National Institutes of Health Grant DK-41612.
References
- 1.Alpern RJ, Chambers M. Cell pH in the rat proximal convoluted tubule: regulation by luminal and peritubular pH and sodium concentration. J Clin Invest. 1986;78:502–510. doi: 10.1172/JCI112602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Amemiya M, Loffing J, Lötscher M, Kaissling B, Alpern RJ, Moe OW. Expression of NHE-3 in the apical membrane of rat renal proximal tubule and thick ascending limb. Kidney Int. 1995;48:1206–1215. doi: 10.1038/ki.1995.404. [DOI] [PubMed] [Google Scholar]
- 3.Azuma KK, Balkovetz DF, Magyar DE, Lescale-Matys L, Zhang Y, Chambrey R, Warnock DG, McDonough AA. Renal Na+/H+ exchanger isoforms and their regulation by thyroid hormone. Am J Physiol Cell Physiol. 1996;270:C585–C592. doi: 10.1152/ajpcell.1996.270.2.C585. [DOI] [PubMed] [Google Scholar]
- 4.Baum M. Neonatal rabbit juxtamedullary proximal convoluted tubule acidification. J Clin Invest. 1990;85:499–506. doi: 10.1172/JCI114465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Baum M. Developmental changes in rabbit juxtamedullary proximal convoluted tubule acidification. Pediatr Res. 1992;31:411–414. doi: 10.1203/00006450-199204000-00021. [DOI] [PubMed] [Google Scholar]
- 6.Baum M, Amemiya M, Dwarakanath V, Alpern RJ, Moe OW. Glucocorticoids regulate NHE-3 transcription in OKP cells. Am J Physiol Renal Fluid Electrolyte Physiol. 1996;270:F164–F169. doi: 10.1152/ajprenal.1996.270.1.F164. [DOI] [PubMed] [Google Scholar]
- 7.Baum M, Biemesderfer D, Gentry D, Aronson PS. Ontogeny of rabbit renal cortical NHE3 and NHE1: effect of glucocorticoids. Am J Physiol Renal Fluid Electrolyte Physiol. 1995;268:F815–F820. doi: 10.1152/ajprenal.1995.268.5.F815. [DOI] [PubMed] [Google Scholar]
- 8.Baum M, Dwarakanath V, Alpern RJ, Moe OW. Effects of thyroid hormone on the neonatal renal cortical Na+/H+ antiporter. Kidney Int. 1998;53:1254–1258. doi: 10.1046/j.1523-1755.1998.00879.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Baum M, Moe OW, Gentry DL, Alpern RJ. Effect of glucocorticoids on renal cortical NHE-3 and NHE-1 mRNA. Am J Physiol Renal Fluid Electrolyte Physiol. 1994;267:F437–F442. doi: 10.1152/ajprenal.1994.267.3.F437. [DOI] [PubMed] [Google Scholar]
- 10.Baum M, Quigley R. Ontogeny of proximal tubule acidification. Kidney Int. 1995;48:1697–1704. doi: 10.1038/ki.1995.467. [DOI] [PubMed] [Google Scholar]
- 11.Baum M, Quigley R. Prenatal glucocorticoids stimulate neonatal juxtamedullary proximal convoluted tubule acidification. Am J Physiol Renal Fluid Electrolyte Physiol. 1991;261:F746–F752. doi: 10.1152/ajprenal.1991.261.5.F746. [DOI] [PubMed] [Google Scholar]
- 12.Blake HH, Henning SJ. Effect of propylthiouracil dose on serum thyroxine, growth, and weaning in young rats. Am J Physiol Regul Integr Comp Physiol. 1985;248:R524–R530. doi: 10.1152/ajpregu.1985.248.5.R524. [DOI] [PubMed] [Google Scholar]
- 13.Cano A. Characterization of the rat NHE3 promoter. Am J Physiol Renal Fluid Electrolyte Physiol. 1996;271:F629–F636. doi: 10.1152/ajprenal.1996.271.3.F629. [DOI] [PubMed] [Google Scholar]
- 14.Cano A, Baum M, Moe OW. Thyroid hormone stimulates the renal Na/H exchanger NHE3 by transcriptional activation. Am J Physiol Cell Physiol. 1999;276:C102–C108. doi: 10.1152/ajpcell.1999.276.1.C102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Choi JY, Shah M, Lee MG, Schultheis PJ, Shull GE, Muallem S, Baum M. Novel amiloride-sensitive sodium-dependent proton secretion in the mouse proximal convoluted tubule. J Clin Invest. 2000;105:1141–1146. doi: 10.1172/JCI9260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chomczynmski P, Sacchi N. Single-step method of RNA isolation by acid guanidine thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 17.Clos J, Crepel F, Legrand C, Legrand J, Rabie A, Vigouroux E. Thyroid physiology during the postnatal period in the rat: a study of the development of thyroid function and of the morphogenetic effects of thyroxine with special reference to cerebellar maturation. Gen Comp Endocrinol. 1974;23:178–192. doi: 10.1016/0016-6480(74)90127-0. [DOI] [PubMed] [Google Scholar]
- 18.D’agostino J, Henning SJ. Role of thyroxine in coordinate control of corticosterone and CBG in postnatal development. Am J Physiol Endocrinol Metab. 1982;242:E33–E39. doi: 10.1152/ajpendo.1982.242.1.E33. [DOI] [PubMed] [Google Scholar]
- 19.Devenport LD, Devenport JA. The effects of adrenal hormones on brain and body size. Physiol Psychol. 1982;10:399–405. [Google Scholar]
- 20.Devenport LD, Devenport JA. Adrenocortical hormones and brain growth: reversibility and differential sensitivity during development. Exp Neurol. 1985;90:44–52. doi: 10.1016/0014-4886(85)90039-1. [DOI] [PubMed] [Google Scholar]
- 21.Edelman CM, Jr, Soriano JR, Boichis H, Gruskin AB, Acosta MI. Renal bicarbonate reabsorption and hydrogen ion excretion in normal infants. J Clin Invest. 1967;46:1309–1317. doi: 10.1172/JCI105623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Freiberg JM, Kinsella J, Sacktor B. Glucocorticoids increase the Na+/H+ exchange and decrease the Na+ gradient-dependent phosphate-uptake systems in renal brush-border membrane vesicles. Proc Natl Acad Sci USA. 1982;79:4932–4936. doi: 10.1073/pnas.79.16.4932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gupta N, Tarif SR, Seikaly M, Baum M. Role of glucocorticoids in the maturation of the rat renal Na+/H+ antiporter (NHE3) Kidney Int. 2001;60:173–181. doi: 10.1046/j.1523-1755.2001.00784.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Henning SJ. Plasma concentrations of total and free corticosterone during development in the rat. Am J Physiol Endocrinol Metab Gastrointest Physiol. 1978;235:E451–E456. doi: 10.1152/ajpendo.1978.235.5.E451. [DOI] [PubMed] [Google Scholar]
- 25.Kandasamy RA, Orlowski J. Genomic organization and glucocorticoid transcriptional activation of the rat Na+/H+ exchanger Nhe3 Gene. J Biol Chem. 1996;271:10551–10559. doi: 10.1074/jbc.271.18.10551. [DOI] [PubMed] [Google Scholar]
- 26.Kinsella J, Cujdik T, Sacktor B. Na+/H+ exchange activity in renal brush border membrane vesicles in response to metabolic acidosis: the role of glucocorticoids. Proc Natl Acad Sci USA. 1984;81:630–634. doi: 10.1073/pnas.81.2.630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kinsella J, Freiberg JM, Sacktor B. Glucocorticoid activation of Na+/H+ exchange in renal brush border membrane vesicles: kinetic effects. Am J Physiol Renal Fluid Electrolyte Physiol. 1985;248:F233–F239. doi: 10.1152/ajprenal.1985.248.2.F233. [DOI] [PubMed] [Google Scholar]
- 28.Kinsella J, Sacktor B. Thyroid hormones increase Na+/H+ exchange activity in renal brush border membranes. Proc Natl Acad Sci USA. 1985;82:3606–3610. doi: 10.1073/pnas.82.11.3606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Loffing J, Lotscher M, Kaissling B, Biber J, Murer H, Seikaly M, Alpern RJ, Levi M, Baum M, Moe OW. Renal Na/H exchanger NHE-3 and Na-PO4 cotransporter NaPi-2 protein expression in glucocorticoid excess and deficient states. J Am Soc Nephrol. 1998;9:1560–1567. doi: 10.1681/asn.v991560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurements with the Folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 31.Meserve LA, Juarez de Ku LM. Effect of thiouracil-induced hypothyroidism on time course of adrenal response in 15 day old rats. Growth Dev Aging. 1993;57:25–30. [PubMed] [Google Scholar]
- 32.Mitsuma T, Nogimori T. Effects of adrenalectomy on the hypothalamic-pituitary-thyroid axis in rats. Horm Metab Res. 1982;14:317–319. doi: 10.1055/s-2007-1019004. [DOI] [PubMed] [Google Scholar]
- 33.Orlowski J, Kandasamy RA, Shull GE. Molecular cloning of putative members of the Na/H exchanger gene family. cDNA cloning, deduced amino acid sequence, and mRNA tissue expression of the rat Na/H exchanger NHE-1 and two structurally related proteins. J Biol Chem. 1992;267:9331–9339. [PubMed] [Google Scholar]
- 34.Roos A, Boron WF. Intracellular pH. Physiol Rev. 1981;61:297–434. doi: 10.1152/physrev.1981.61.2.296. [DOI] [PubMed] [Google Scholar]
- 35.Schwartz GJ, Evan AP. Development of solute transport in rabbit proximal tubule. I. and glucose absorption. Am J Physiol Renal Fluid Electrolyte Physiol. 1983;245:F382–F390. doi: 10.1152/ajprenal.1983.245.3.F382. [DOI] [PubMed] [Google Scholar]
- 36.Shah M, Gupta N, Dwarakanath V, Moe WO, Baum M. Ontogeny of Na+/H+ antiporter activity in rat proximal convoluted tubules. Pediatr Res. 2000;48:206–210. doi: 10.1203/00006450-200008000-00014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Shah M, Quigley R, Baum M. Maturation of rabbit proximal straight tubule chloride-base exchange. Am J Physiol Renal Physiol. 1998;274:F883–F888. doi: 10.1152/ajprenal.1998.274.5.F883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Stith RD, Reddy YS. Myocardial contractile protein ATPase activities in adrenalectomized and thyroidectomized rats. Basic Res Cardiol. 1992;87:519–526. doi: 10.1007/BF00788662. [DOI] [PubMed] [Google Scholar]
- 39.Walker P, Dubois JD, Dussault JH. Free thyroid hormone concentrations during postnatal development in the rat. Pediatr Res. 1980;14:247–249. doi: 10.1203/00006450-198003000-00014. [DOI] [PubMed] [Google Scholar]
- 40.Wang T, Yang CL, Abbiati T, Schultheis PJ, Shull GE, Giebisch G, Aronson PS. Mechanisim of proximal tubule bicarbonate absorption in NHE3 null mice. Am J Physiol Renal Physiol. 1999;277:F298–F302. doi: 10.1152/ajprenal.1999.277.2.F298. [DOI] [PubMed] [Google Scholar]
- 41.Wu MS, Biemesderfer D, Giebisch G, Aronson PS. Role of NHE3 in mediating renal brush border Na+/H+ exchange. J Biol Chem. 1996;271:32749–32752. doi: 10.1074/jbc.271.51.32749. [DOI] [PubMed] [Google Scholar]
- 42.Yagil R, Lerner Z, Etzion Z, Berlyne GM. Acid-base changes in milk and blood of rats in acidosis and alkalosis. Am J Physiol. 1976;231:132–135. doi: 10.1152/ajplegacy.1976.231.1.132. [DOI] [PubMed] [Google Scholar]