

Abstract
Aggressive disorders are moderately heritable; therefore, identification of genetic influences is important. The X-linked MAOA gene, encoding the MAOA enzyme, has a functional 30bp repeat polymorphism in the promoter region (MAOA-LPR) that has been shown to influence aggression. Childhood trauma is a known risk factor for numerous psychopathologies in adulthood including aggressive behaviors. We investigated the interactive effect of MAOA-LPR genotype and a history of childhood trauma in predicting aggressive behaviors in a prisoner population. A total of 692 male prisoners were genotyped for MAOA-LPR with genotypes grouped into high and low transcriptional activity. Participant evaluations included measures of aggression (BGHA), hostility (Buss Durkee Hostility Inventory), impulsivity (Barratt Impulsiveness Scale), violence directed towards self and others, and childhood trauma (Childhood Trauma Questionnaire (CTQ)). MAOA-LPR interacted with CTQ physical neglect (PN), the most common (47%) form of childhood trauma in this sample, to predict BGHA aggression (P=0.002). Within the group not exposed to PN, carriers of the MAOA-LPR high activity variant were more aggressive: (t(R) =2.47, p<0.014). We observed a crossover effect in that the increase in aggression scores with PN was greater in low activity individuals (t(R) =5.55, p <0.0001) than in high activity individuals (t(R) =4.18, p <0.0001). These findings suggest that childhood trauma and the functional MAOA-LPR polymorphism may interact to specifically increase risk for over aggressive behavior but not impulsivity or hostility. The MAOA-LPR low activity variant may be protective against the development of aggressive behavior under low stress conditions, at least in this prisoner population.
Keywords: MAOA-LPR genotype, aggressive behavior, BGHA, childhood trauma, CTQ, physical neglect, prisoners, MAOA-LPR low activity variant, MAOA-LPR high activity variant, interactive effect
INTRODUCTION
A degree of aggressive behavior can be socially acceptable and considered part of the spectrum of normal behavior. However aggression directed towards self or others, for example, antisocial behavior, suicidal behavior or violent criminality (Carli et al., 2014, Dejong et al., 1992, Mann et al., 2009) is considered pathological. Genetic and environmental influences, such as childhood trauma, are likely to play a role in the development of aggression. Studies have shown that psychiatrically classified aggressive disorders such as antisocial personality disorder, borderline personality disorder and intermittent explosive disorder are moderately heritable (Coccaro et al., 1993, Kendler et al., 2008, Lyons et al., 1995, Torgersen et al., 2008). Childhood trauma is known to be a risk factor for numerous psychopathologies in adulthood (Enoch, 2011) including aggressive behaviors such as suicidal behavior (Carli et al., 2011, Roy et al., 2010) and delinquency and adult violent criminal behavior (Carli et al., 2014, Driessen et al., 2006, Sarchiapone et al., 2009, Widom, 1989). Childhood trauma, identified by the Childhood Trauma Questionnaire (CTQ), has been correlated with measures of lifetime aggression (Brown-Goodwin Lifetime History of Aggression (BGHA) interview (Brown et al., 1979) in the Italian prisoner population analyzed in this study (Sarchiapone et al., 2009) and in other studies (Garno et al., 2008).
One gene that has robustly been shown to influence aggression is the X-linked monoamine oxidase A (MAOA) gene that encodes the MAOA enzyme which plays an important role in the degradation of central nervous system serotonin and norepinephrine. A deficiency in MAOA has been associated with aggressive behavior in adult male knockout mice and men in a Dutch family (Brunner et al., 1993, Cases et al., 1995). In healthy men, MAOA enzyme activity in cortical regions correlates inversely with measures of aggression (Alia-Klein et al., 2008). MAOA binding in prefrontal cortex of healthy non-smoking men correlates negatively with angry-hostility (Soliman et al., 2011). The MAOA gene has a 30 base pair repeat in the promoter region (MAOA-LPR) that has been shown to affect transcriptional efficiency in vitro (Sabol et al., 1998). In healthy humans, MAOA-LPR genotype predicts limbic circuitry variation during emotional arousal together with diminished reactivity of regulatory prefrontal regions (Meyer-Lindenberg et al., 2006). The high and low activity MAOA-LPR variants also predict changes in different brain regions during tests for impulsivity (Passamonti et al., 2006).
Several (but not all) studies have shown that MAOA-LPR interacts with childhood trauma to predict aggressive or disinhibited behaviors (Byrd & Manuck, 2014, Caspi et al., 2002, Ducci et al., 2008, Enoch et al., 2010). For example, Byrd and Manuck (2014) in their recent meta-analysis of 20 male cohorts reported significant interactive effects of childhood trauma and the low activity MAOA-LPR variant on antisocial outcomes. However, this meta-analysis and other earlier studies have not included prisoners, who are more likely to have extreme aggressive phenotypes. As far as we are aware there has been only one study of MAOA-LPR in relation to aggression in prisoners which found converse results to general population studies in that the high activity variant was implicated in aggressive behavior (Tikkanen et al., 2010). The aim of the present study was to investigate the interactive effect of MAOA-LPR genotype and history of childhood trauma in predicting aggressive behaviors in a large group of male Italian prisoners who were evaluated for psychiatric disorders together with impulsive traits, lifetime aggressive behavior, and hostility and violent behavior during incarceration.
METHODS
Participants
Full details of the dataset are provided in Bevilacqua et al, 2012. The participants with analyzable MAOA-LPR genotypes included 692 male prisoners detained in the Penitentiary District of Abruzzo-Molise in central Italy. Prisoners had been convicted for violent crimes including homicide, aggression with weapons, violent robberies and terrorist activity, and non-violent crimes including drug use or sale, burglary and fraud. The mean (SD) age was 40.4 (11) years. All participants self-identified as Caucasians and ethnicity was also recorded by the interviewer. Informed consent was obtained from all participants after a detailed explanation of the study was provided by a psychiatrist. The Ethics Review Board of the University of Molise approved the study.
Psychiatric interviews were conducted by trained psychologists and psychiatrists. The Italian version of the structured Mini International Neuropsychiatric Interview (MINI) (Sheehan et al., 1998) was administered. Lifetime psychiatric disorders were determined according to DSM-IV criteria. Exclusion criteria included mental retardation, schizophrenia and the inability to read or speak Italian.
A total of 32.8% of the 692 prisoners had a lifetime DSM-IV diagnosis of substance dependence: opiates (4%), alcohol (8.4%), cannabis (11.9%), cocaine (30.8%), and multiple substances (44.9%). Since substance dependence is a predictor of aggression and violent behavior the term ‘any substance dependence’ (N = 227) was included in the analyses.
Measures of aggression, hostility, impulsivity and violence directed towards self and others
Brown-Goodwin Lifetime History of Aggression interview
The BGHA (Brown et al., 1979) is an 11 item interview that assesses lifetime aggressive behavior during adolescence and adulthood including temper tantrums and violence against self, property and others (including authority) in various social contexts, such as family, school and work environment. The number of times each item occurred is recorded. BGHA total possible scores range from 0 to 88. All psychometric properties are detailed in (Suris et al., 2004). A total of 678 participants completed the BGHA.
Buss-Durkee Hostility Inventory
The BDHI (Buss & Durkee, 1957) is a 75 item questionnaire developed to assess eight aspects of hostility: assault, indirect aggression, irritability, negativism, resentment, suspicion, verbal expression of negative affect and guilt. The BDHI total possible scores range from 0 to75. All psychometric properties are detailed in (Suris et al., 2004). A total of 672 participants completed the BDHI.
The Barratt Impulsiveness Scale
The BIS (Barratt, 1965, Patton et al., 1995) is a 30-item, 4 point Likert scale questionnaire that investigates personality/behavioral impulsiveness, including cognitive impulsiveness, motor impulsiveness and lack of planning. The BIS total possible scores range from 0 to 120. All psychometric properties are detailed in Suris et al, 2004. A total of 665 participants completed the BIS.
Violent behavior during incarceration
Prisoners were recorded as having exhibited violent behavior during their incarceration if there were disciplinary reports of physical aggression or assault against other inmates or prison officers while in prison. Verbal aggression and behaviors other than physical violence (for example drug dealing) were excluded from the definition of the variable. Examples for violent behavior in jail are fights between inmates or assault against a prison guard.
Lifetime history of suicide attempt and/or self-mutilation
A lifetime history of suicide attempt was assessed during the psychiatric interview. A suicide attempt was defined as a self-destructive act with some intent to end one’s life that was not self-mutilatory in nature
Eysenck Personality Questionnaire
Prisoners completed the EPQ (Eysenck Personality Questionnaire) that is a 69-item version specifically adapted for the Italian language. It is based on the original version of the Eysenck Personality Questionnaire (EPQ) (Eysenck, 1975a, Eysenck, 1975b), from which the psychoticism and neuroticism sores were obtained. Data was available for 680 prisoners. Since psychoticism and neuroticism are predictors of aggression (Eysenck H.J., 1985), these two measures of personality were included in the analyses as independent variables.
Measures of childhood trauma
Prisoners completed the CTQ-34 item version (Bernstein et al., 1997). The CTQ is an instrument for assessing the 5 subscales of physical abuse (PA), physical neglect (PN), emotional abuse (EA), emotional neglect (EN) and sexual abuse (SA). For each item there is a 5-point Likert scale to express the frequency of occurrence. The 34-item CTQ was converted into the 28-item version according to accepted criteria since this is the most recent and commonly used form of the questionnaire. CTQ subscale scores range from 5 to 25 and the total scores from 25 to 125. Reliability and validity of the Childhood Trauma Questionnaire have been previously demonstrated (Bernstein et al., 1997, Bernstein et al., 1994). Continuous CTQ scores were used in all analyses.
CTQ dichotomous clinical cutoff scores differentiate between the presence or absence of significant abuse and neglect (Walker et al., 1999). The cutoff points were 8 or higher for PA, PN and SA; 10 or higher for EA, and 15 or higher for EN. In this study we used CTQ clinical cutoff only for graphical illustration of G × E interaction.
Genotyping
DNA was extracted from whole blood using standard protocols. The MAOA gene promoter VNTR polymorphism (MAOA-LPR) genotyping was modified from the method of Ducci and colleagues (Ducci et al., 2006). MAOA-LPR was amplified from 10 ng genomic DNA using the primer sequences: forward 5'-(CCC AGG CTG CTC CAG AAA CAT G)-3' and reverse-5'(GTT CGG GAC CTG GGC AGT TGT G)-3'. Owing to the high GC content in the VNTR region, amplification was performed using Invitrogen's PlatinumTaq and PCR X Enhancer System kits, according to the manufacturer's protocol (Invitrogen, Carlsbad, CA, USA), with 5 µM of each primer and 25 mM dNTPs in a total reaction volume of 15µl. Amplifications were performed on a Perkin-Elmer 9700 thermocycler (Applied Biosystems, Foster City, CA, USA) with one cycle at 96°C for 10 min followed by 35 cycles of 94°C for 15 s, 55°C for 15 s, 72°C for 30 s, and a final 3 min extension at 72°C. The forward primer was labeled with the fluorescent dye 6-FAM. Amplicons were visualized with GeneScan-500 LIZ Size Standard (Applied Biosystems, Foster City, CA, USA) and analyzed on an ABI 3730 capillary sequencer. Allele sizes (allele 2, 183 bp; allele 3, 213 bp; allele 3.5, 232 bp; allele 4, 244 bp; allele 5, 272 bp) were determined using GeneMapper v4.0 (Applied Biosystems, Foster City, CA, USA). Genotyping accuracy was determined empirically by duplicate genotyping of 25% of the samples selected randomly. The overall error rate was <0.005, and the completion rate was >0.98.
The frequencies of the MAOA-LPR alleles were as follows: 3 repeat allele 0.322; 3.5 repeat allele 0.007; 4 repeat allele 0.658. Alleles with 5 and 2 repeats were detected in only eight and one individuals respectively of the original sample of 701 and were excluded from analysis due to the controversial effect on transcription (Deckert et al., 1999, Guo et al., 2008, Sabol et al., 1998).
As MAOA is an X linked gene and since our study included only males, for the remaining 692 individuals, genotypes were grouped by relative transcriptional activity of MAOA into two categories: high-activity (3.5 and 4 repeats, 0.67) versus low-activity (3 repeats, 0.33).
The distribution of the high activity (MAOA-H) and low activity (MAOA-L) alleles in the current study is similar to the distribution in other Caucasian male samples (Caspi et al., 2002, Manuck et al., 2000, Ni et al., 2007, Reif et al., 2007, Weder et al., 2009).
Statistical analysis
We set up a filtering process for the inclusion of dependent variables in analyses. The first criterion was that there was an effect of CTQ on the dependent variable. Correlation analyses were conducted between the continuous CTQ score and the continuous BGHA, BDHI, and BIS scores. In the event of a significant correlation (p < 0.05), the BGHA, BDHI and BIS scores would be included in separate linear regression analyses as the dependent variable. Likewise, if violent behavior in jail and self-directed violent behavior were associated with significantly (p < 0.05) higher CTQ scores, they would be included in separate logistic regression analyses as the dependent variables. The independent variables for the linear and logistic regression analyses included age, any substance dependence, EPQ neuroticism and psychoticism, continuous CTQ score, MAOA-LPR genotype (high and low activity variants) and the interaction term: MAOA-LPR genotype × CTQ score. If the CTQ did not show a main effect on the dependent variable within the full model, then that dependent variable was eliminated from further analysis.
Correction for multiple comparisons
Since three dependent variables (BGHA, violent behavior in jail, violent behavior towards self) survived the filtering process described above, P values were Bonferroni corrected for three independent tests: p < 0.017.
Primary analyses
A linear regression model was performed with BGHA scores as the dependent variable and age, any substance dependence, EPQ neuroticism and psychoticism scores, continuous CTQ score and MAOA-LPR genotype as the independent variables. All interaction terms were initially included: MAOA-LPR × CTQ score, MAOA-LPR × psychoticism score; MAOA-LPR × neuroticism score, MAOA-LPR × age, MAOA-LPR × any substance dependence. A backward stepwise regression was performed with variables being eliminated from the model in an iterative process. Interaction terms were included in the final model if p ≤ 0.1. The same process was performed for the logistic regression analyses for violent behavior in jail and violent behavior towards self.
Secondary analyses
If within each of the linear or logistic regression analyses, the MAOA-LPR × total CTQ score interaction term had a significance of p ≤ 0.1, secondary analyses were performed with the continuous scores of each of the 5 CTQ subscales. The 5 scores were entered into the whole model to determine whether any of the subscales had an independent effect on the dependent variable. If so, the relevant MAO-LPR × CTQ subscale interaction term was included.
Statistical analyses, including power analyses, were undertaken using JMP9 software.
RESULTS
Correlation between CTQ scores and measures of aggression, hostility, and impulsivity
The mean, SD, median and range of the total CTQ score and the 5 subscale scores are provide in supplementary Table S1. The full distribution statistics for the BGHA, BDHI and BIS are presented in supplementary Table S2. The mean (SD) scores were as follows: BGHA: 36 (12) (max possible score = 88); BDHI: 37 (11) (max possible score = 75); BIS: 48 (15) (max possible score = 120).
BGHA scores were correlated with CTQ scores: r2 = 0.18, p < 0.0001 and the CTQ had a significant main effect in the linear regression model (p < 0.001). BDHI scores were modestly correlated with CTQ scores: r2 = 0.05, p < 0.0001, however there was no main effect of CTQ in the linear regression model and therefore the BDHI was eliminated from further analysis.
BIS scores were not correlated with CTQ scores (p = 0.101) therefore the BIS was eliminated from further analysis.
Association between childhood trauma and violent behavior during incarceration
Violent behavior in jail was documented for 162 of the 692 prisoners. Violent behavior vs. none: CTQ (mean (SD)): 44.1(16.2) vs. 38.6(12.6); F (1,690) =20.9, p<0.0001. The CTQ had a significant main effect in the logistic regression model (p < 0.001).
Association between childhood trauma and lifetime history of suicide attempt and/or self-mutilation
A total of 146 of 692 prisoners had attempted self-harm: Suicide attempts /self mutilation vs. none: CTQ (mean (SD)): 47.4 (18.5) vs. 37.9(11.3); F (1,690) =61.2, p<0.0001. The CTQ had a significant main effect in the logistic regression model (p < 0.001).
Primary analyses
Table 1 shows the whole model analyses for the BGHA, violent behavior in prison and self-directed violent behavior. There were no main effects of MAOA-LPR genotype on any of these three measures of aggressive behavior. The MAOA-LPR × CTQ interaction term (p = 0.037) and the MAOA-LPR × psychoticism interaction term (p = 0.096) satisfied the criterion that p ≤ 0.1 only for the analysis for BGHA. The whole model accounted for 33% of the variance in BGHA score. A power analysis revealed that for MAOA-LPR × CTQ, there was 0.47 power to detect an effect (α = 0.05).
Table1.
Interaction of MAOA -LPR and CTQ scores on Brown-Goodwin Lifetime History of Aggression (BGHA) scores, violent behavior during incarceration and lifetime history of suicide attempts and/or self-mutilation.
| BGHA scores |
Violent behavior during incarceration |
Lifetime history of suicide attempts and/or self- mutilation |
||||
|---|---|---|---|---|---|---|
| Variable | F | P | L-R χ2 | P | L-R χ2 | P |
| Age | 24.83 | <0.0001 | 8.54 | 0.0035 | 0.07 | 0.8 |
| Psychoticism | 15.68 | <0.0001 | 2.73 | 0.098 | 6.98 | 0.0083 |
| Neuroticism | 39.8 | <0.0001 | 3.61 | 0.058 | 21.19 | <0.0001 |
| Any substance dependence | 10.10 | 0.0016 | 9.04 | 0.0026 | 9.68 | 0.0019 |
| CTQ total continuous score | 91.82 | <0.0001 | 8.61 | 0.0034 | 17.66 | <0.0001 |
| MAOA-LPR H vs L | 1.59 | 0.21 | 0.34 | 0.56 | 0.25 | 0.62 |
| G*E: MAOA-LPR × CTQ | 4.38 | 0.037 | 2.87 | 0.089 | 0.82 | 0.36 |
| G*E: MAOA-LPR × Psychoticism | 2.78 | 0.096 | -- | -- | -- | -- |
| Whole model | 40.13 | <0.0001 | 56.59 | <0.0001 | 101.92 | <0.0001 |
| Whole model variance, df | 0.33, 8 | 0.08, 7 | 0.15, 7 | |||
L-R χ2 = Likelihood Ratio chi square; df = degrees of freedom
MAOA-LPR H v L: high activity variant vs low activity variant
G*E: MAOA-LPR × CTQ contributes 0.013 of the total variance in BGHA aggression
Psychoticism and neuroticism scores derive from the Eysenck Personality Questionnaire
G*E interaction term MAOA-LPR × Psychoticism for violent behavior during incarceration, p = 0.85 and for suicide attempts / self-mutilation, p = 0.82. Therefore this interaction term was not included in these logistic regression analyses.
Secondary analyses for BGHA
Since the 5 subscales: PA, PN, EA, EN and SA are all highly correlated in this sample (Bevilacqua et al., 2012) we replaced the total CTQ continuous score by the continuous scores for the 5 subscales to determine which might have an independent effect on aggression as measured by the BGHA. PN, PA and SA had independent effects (p < 0.05) but not EN (p = 0.56) and EA (p = 0.90). Therefore the G × E interaction term for PN, PA and SA were included in the whole model analyses as shown in Table 2. The G × E interaction term was significant for PN (p = 0.002) but not for PA or SA. A power analysis revealed that for MAOA-LPR × PN there was sufficient power (0.81) to detect an effect (α = 0.05). However there was insufficient power for MAOA-LPR × SA (0.17) and MAOA-LPR × PA (0.11).
Table 2.
Interaction of MAOA-LPR and CTQ subscale scores for childhood physical neglect, physical abuse and sexual abuse on Brown-Goodwin Lifetime History of Aggression (BGHA) scores
| BGHA Scores |
||
|---|---|---|
| Variable | F | P |
| Age | 18.23 | <0.0001 |
| Psychoticism | 14.00 | 0.0002 |
| Neuroticism | 36.69 | <0.0001 |
| Any substance dependence | 11.60 | 0.0007 |
| MAOA-LPR H vs L | 1.12 | 0.29 |
| G*E: MAOA-LPR × Psychoticism | 2.61 | 0.11 |
| Physical neglect (PN) | 53.98 | <0.0001 |
| G*E: MAOA-LPR × PN | 10.37 | 0.001 |
| Physical abuse (PA) | 4.31 | 0.038 |
| G*E: MAOA-LPR × PA | 1.32 | 0.25 |
| Sexual abuse (SA) | 9.10 | 0.003 |
| G*E : MAOA-LPR × SA | 0.35 | 0.55 |
| Whole model | 30.35 | <0.0001 |
| Whole model - variance, df | 0.36, 12 | |
MAOA-LPR H v L: high activity variant vs low activity variant; df = degrees of freedom
G*E: MAOA-LPR × PN contributes 0.011 of the total variance in BGHA aggression
The continuous scores for PN, PA and SA were entered into this analysis.
Psychoticism and neuroticism scores derive from the Eysenck Personality Questionnaire
Clinical cut-off scores for the 5 CTQ subscales
As described in Methods, the CTQ provides clinical cutoffs that set thresholds for significant trauma within each of the 5 categories. In the total group of prisoners, 431 (63.2%) met the threshold for significant abuse and /or neglect in at least 1 of the 5 categories: 195 (45.2%) experienced only 1 type of childhood trauma, 101 (23.4%) reported 2 types, 59(13.7%) −3 types, 53 (12.3%) −4 types, and 23 (5.3%) reported all 5 categories of trauma. PN (325 (47%)) was the most common form of childhood trauma in this sample, followed by EN (164 (24%)), PA (158 (23%)), SA (143 (21%)) and EA (111 (16%)).
PN GxE interaction
Table 2 shows that the continuous PN score interacted with MAOA-LPR to predict aggression. In order to illustrate the G × E interaction we used the PN dichotomous clinical cutoff score to produce Figure 1. This shows that exposure to clinically significant PN was associated with increased BGHA scores (39.9±0.63 vs 32.4±0.56), t(R) =6.84, p<0.0001. However, MAOA-LPR genotype had no effect on scores of aggression within the group exposed to PN (p = 0.441). In contrast, within the group with no history of childhood PN, carriers of the MAOA-LPR high activity variant were more aggressive: *t(R) =2.47, p<0.014. Figure 1 shows that there is a crossover effect in that the increase in aggression scores with PN is greater in low activity individuals (t(R) =5.55, p <0.0001) than in high activity individuals (t(R) =4.18, p <0.0001).
Figure 1.
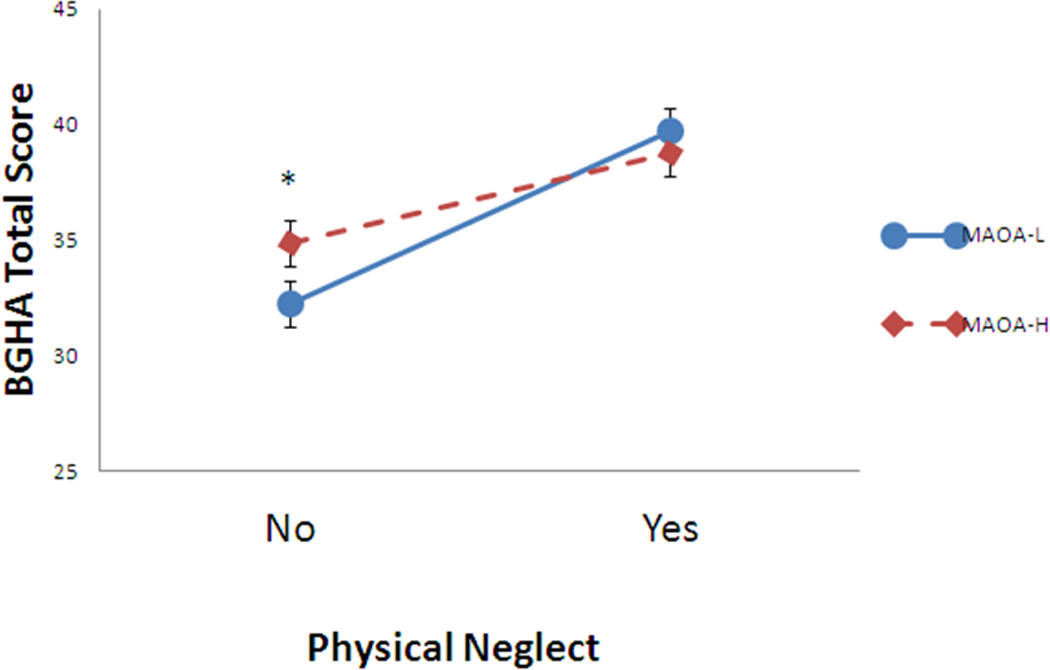
Interaction of MAOA-LPR low and high activity variants and presence or absence of childhood physical neglect on Brown-Goodwin Lifetime History of Aggression (BGHA) scores
MAOA-LPR high (MAOA-H) and low (MAOA-L) activity variants.
Childhood Trauma Questionnaire physical neglect subscale, clinical cut-off:
No physical neglect (n=361): MAOA-L, n=130; MAOA-H, n=213
Yes physical neglect (n=317): MAOA-L, n=90; MAOA-H, n=227
*p=0.014
Error bars are standard errors
BGHA scores range from 0 – 88
DISCUSSION
Numerous studies have shown that childhood trauma alters stress reactivity and is associated with psychopathology, including aggressive behaviors (Enoch, 2011, Frazzetto et al., 2007, Lovallo, 2013). In this sample of Caucasian Italian prisoners we have shown that CTQ childhood trauma influenced lifetime aggression, as measured by the BGHA, a lifetime history of suicidal behavior and/or self-mutilation and violent behavior during incarceration. However childhood trauma did not predict trait impulsivity, as measured by the BIS, or hostility as measured by the BDHI. When we investigated the MAOA-LPR – childhood trauma interaction we found that, within the group not exposed to PN, carriers of the MAOA-LPR high activity allele were more aggressive. However, as can be seen in Figure 1, carriers of the low activity allele were more vulnerable to the effects of PN on aggression: in the group exposed to PN, both high and low activity allele carriers had similar degrees of aggression. Our results can be considered to be robust in that the sizes for each of the four PN – MAOA-LPR genotype groups are large, ranging from N = 90 to N = 227.
Almost half (47%) of the Italian prisoners had experienced significant PN. In contrast, ‘only’ 16 – 24% had experienced EA, SA, PA or EN. We found that, as expected, exposure to childhood PN was associated with increased aggression however we did not find an effect of genotype on aggression in the PN exposed group. Indeed, results of studies on the interactive influence of childhood trauma and MAOA-LPR on human aggression are contradictory. A recent meta-analysis of 20 male cohorts provides support for the frequently reported interactive effect of childhood trauma and the low activity variant on antisocial outcomes (Byrd & Manuck, 2014). In contrast, a recent large study (not included in the meta-analysis) in 3356 white and 960 black men found no G × E effect for MAOA-LPR on adult antisocial behaviors (Haberstick et al., 2014). The aforementioned meta-analysis also looked at results from 11 female cohorts and found that in women it was the high activity variant that interacted with childhood maltreatment to predict antisocial behaviors (Byrd & Manuck, 2014). In the only other published MAOA-LPR study of prisoners, the high activity variant was implicated in aggressive behavior in a group of Finnish alcoholic violent offenders who were followed for eight years after release from prison. In that sample of prisoners, carriers of the MAOA-LPR high activity variant who had experienced childhood PA before age 13 were at increased risk of committing severe, impulsive, violent recidivist crimes, mediated through excessive drinking (Tikkanen et al., 2010). The results of our and the Tikkanen et al 2010 study, both in Caucasian men, differ from the results of studies derived from general population samples of males and it is possible that prisoners may be an atypical group for MAOA effects on aggression. For example, prisoners as a group have higher levels of anger, aggression, and self-injurious behaviors than general population samples (Jenkins et al., 2005, Ohlsson & Ireland, 2011, Rivlin et al., 2013, Sakelliadis et al., 2010). Another aspect might be resilience, i.e. the ability to thrive in the face of adversity. The prisoners in our study had completed the Connor-Davidson Resilience Scale (CD-RISC) that has validated mean scores in several populations including the US general population (80.7), primary care patients (71.8) and psychiatric outpatients (68.0) (Connor & Davidson, 2003). The prisoners in our study had very low mean (SD) resilience scores: 65.5 (13.6). Thus with stronger aggression phenotypes and increased vulnerability to stress, the use of prison populations might increase power to detect small G × E interactive effects. Therefore the results of our study may be generalizable to other prison samples or other antisocial groups rather than to general population samples.
It is tempting to speculate that our results are an indication that the MAOA-LPR low activity variant is protective against aggressive behavior under low stress conditions. However exposure to stressors during development might result in altered HPA axis stress reactivity. In fact we previously showed, in the same dataset, that there was an interaction between variation in FKBP5, a stress related gene influencing the HPA axis, and CTQ childhood trauma on BGHA aggression (Bevilacqua et al., 2012).
The five CTQ questions for PN are: “not enough to eat; got taken care of (reverse scored); parents were drunk or high; wore dirty clothes; got taken to doctor (reverse scored)”. Prisoners might be more likely to originate from impoverished backgrounds and therefore a high PN score could conceivably be the result of poverty rather than true parental neglect, hence in this sample of prisoners, PN might not be discriminating enough for the manifestation of MAOA-LPR genotype effects.
There are several strengths to this study. Firstly, we had access to a large selected extreme sample of individuals who have been incarcerated for committing an offence. Secondly, we used the CTQ which enabled us to examine subtypes of childhood trauma. Thirdly, we were able to analyze several aspects of this heterogeneous phenotype ‘aggression’: overt aggression and violence, indirect aggression, hostility and impulsive personality traits. As a result we were able to demonstrate that the MAOA-LPR × PN interaction had a specific effect on aggressive behavior and not on the other aspects of aggression / impulsivity listed above.
Limitations of this study include the fact that both the CTQ and BGHA questionnaires are self-report, and that the CTQ does not include an exhaustive list of the potentially traumatic events that could be experienced in childhood. Also, relative to PN, fewer prisoners had experienced significant PA and SA and, as shown by the power analysis, the negative G × E results for PA and SA may be due to a lack of power.
In conclusion this study in male Caucasian Italian prisoners has shown that CTQ childhood trauma influenced overt aggressive behavior but not impulsivity or hostility. MAOA-LPR interacted with PN, the most common form of childhood trauma in this sample, to increase the risk of aggressive behavior as measured by the BGHA. Within the group not exposed to PN, carriers of the MAOA-LPR high activity variant were more aggressive. However we observed a crossover effect in that the increase in aggression scores in PN exposed men was greater in low activity individuals than in high activity individuals. At least in this prisoner population, the MAOA-LPR low activity variant may have been protective during childhood development against the propensity to aggressive behaviors in low stress individuals i.e. those not exposed to physical neglect.
Supplementary Material
Acknowledgements
We wish to thank Dr. Bruna Brunetti for support in the organization of the study, the Societa’ Italiana di Medicina e Sanita’ Penitenziaria (SIMSPE), the Penitientiary Institutions and the Provveditorato dell’Amministrazione Penitenziaria dell’Abruzzo e Molise (PRAP-Abruzzo-Molise).
Financial Disclosure
This research was supported by the Intramural Research Program of the National Institute on Alcohol Abuse and Alcoholism, NIH and in part by grand R01 DA 10336-02 to AR from the National Institute of Drug Abuse, NIH. Dr. Sarchiapone has received financial support from Pfizer and Lundbeck.
Footnotes
The authors declare no conflict of interest.
References
- Alia-Klein N, Goldstein RZ, Kriplani A, Logan J, Tomasi D, Williams B, Telang F, Shumay E, Biegon A, Craig IW, Henn F, Wang GJ, Volkow ND, Fowler JS. Brain monoamine oxidase A activity predicts trait aggression. J Neurosci. 2008;28:5099–5104. doi: 10.1523/JNEUROSCI.0925-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barratt ES. Factor Analysis of Some Psychometric Measures of Impulsiveness and Anxiety. Psychol Rep. 1965;16:547–554. doi: 10.2466/pr0.1965.16.2.547. [DOI] [PubMed] [Google Scholar]
- Bernstein DP, Ahluvalia T, Pogge D, Handelsman L. Validity of the Childhood Trauma Questionnaire in an adolescent psychiatric population. J Am Acad Child Adolesc Psychiatry. 1997;36:340–348. doi: 10.1097/00004583-199703000-00012. [DOI] [PubMed] [Google Scholar]
- Bernstein DP, Fink L, Handelsman L, Foote J, Lovejoy M, Wenzel K, Sapareto E, Ruggiero J. Initial reliability and validity of a new retrospective measure of child abuse and neglect. Am J Psychiatry. 1994;151:1132–1136. doi: 10.1176/ajp.151.8.1132. [DOI] [PubMed] [Google Scholar]
- Bevilacqua L, Carli V, Sarchiapone M, George DK, Goldman D, Roy A, Enoch MA. Interaction between FKBP5 and childhood trauma and risk of aggressive behavior. Arch Gen Psychiatry. 2012;69:62–70. doi: 10.1001/archgenpsychiatry.2011.152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown GL, Goodwin FK, Ballenger JC, Goyer PF, Major LF. Aggression in humans correlates with cerebrospinal fluid amine metabolites. Psychiatry Res. 1979;1:131–139. doi: 10.1016/0165-1781(79)90053-2. [DOI] [PubMed] [Google Scholar]
- Brunner HG, Nelen M, Breakefield XO, Ropers HH, van Oost BA. Abnormal behavior associated with a point mutation in the structural gene for monoamine oxidase A. Science. 1993;262:578–580. doi: 10.1126/science.8211186. [DOI] [PubMed] [Google Scholar]
- Buss AH, Durkee A. An inventory for assessing different kinds of hostility. J Consult Psychol. 1957;21:343–349. doi: 10.1037/h0046900. [DOI] [PubMed] [Google Scholar]
- Byrd AL, Manuck SB. MAOA, childhood maltreatment, and antisocial behavior: meta-analysis of a gene-environment interaction. Biological psychiatry. 2014;75:9–17. doi: 10.1016/j.biopsych.2013.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carli V, Mandelli L, Zaninotto L, Alberti S, Roy A, Serretti A, Sarchiapone M. Trait-aggressiveness and impulsivity: Role of psychological resilience and childhood trauma in a sample of male prisoners. Nord J Psychiatry. 2014;68:8–17. doi: 10.3109/08039488.2012.756061. [DOI] [PubMed] [Google Scholar]
- Carli V, Roy A, Bevilacqua L, Maggi S, Cesaro C, Sarchiapone M. Insomnia and suicidal behaviour in prisoners. Psychiatry Res. 2011;185:141–144. doi: 10.1016/j.psychres.2009.10.001. [DOI] [PubMed] [Google Scholar]
- Cases O, Seif I, Grimsby J, Gaspar P, Chen K, Pournin S, Muller U, Aguet M, Babinet C, Shih JC. Aggressive behavior and altered amounts of brain serotonin and norepinephrine in mice lacking MAOA. Science. 1995;268:1763–1766. doi: 10.1126/science.7792602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caspi A, McClay J, Moffitt TE, Mill J, Martin J, Craig IW, Taylor A, Poulton R. Role of genotype in the cycle of violence in maltreated children. Science. 2002;297:851–854. doi: 10.1126/science.1072290. [DOI] [PubMed] [Google Scholar]
- Coccaro EF, Bergeman CS, McClearn GE. Heritability of irritable impulsiveness: a study of twins reared together and apart. Psychiatry Res. 1993;48:229–242. doi: 10.1016/0165-1781(93)90074-q. [DOI] [PubMed] [Google Scholar]
- Connor KM, Davidson JR. Development of a new resilience scale: the Connor-Davidson Resilience Scale (CD-RISC) Depression and anxiety. 2003;18:76–82. doi: 10.1002/da.10113. [DOI] [PubMed] [Google Scholar]
- Deckert J, Catalano M, Syagailo YV, Bosi M, Okladnova O, Di Bella D, Nothen MM, Maffei P, Franke P, Fritze J, Maier W, Propping P, Beckmann H, Bellodi L, Lesch KP. Excess of high activity monoamine oxidase A gene promoter alleles in female patients with panic disorder. Hum Mol Genet. 1999;8:621–624. doi: 10.1093/hmg/8.4.621. [DOI] [PubMed] [Google Scholar]
- DeJong J, Virkkunen M, Linnoila M. Factors associated with recidivism in a criminal population. J Nerv Ment Dis. 1992;180:543–550. doi: 10.1097/00005053-199209000-00001. [DOI] [PubMed] [Google Scholar]
- Driessen M, Schroeder T, Widmann B, von Schonfeld CE, Schneider F. Childhood trauma, psychiatric disorders, and criminal behavior in prisoners in Germany: a comparative study in incarcerated women and men. J Clin Psychiatry. 2006;67:1486–1492. doi: 10.4088/jcp.v67n1001. [DOI] [PubMed] [Google Scholar]
- Ducci F, Enoch MA, Hodgkinson C, Xu K, Catena M, Robin RW, Goldman D. Interaction between a functional MAOA locus and childhood sexual abuse predicts alcoholism and antisocial personality disorder in adult women. Mol Psychiatry. 2008;13:334–347. doi: 10.1038/sj.mp.4002034. [DOI] [PubMed] [Google Scholar]
- Ducci F, Newman TK, Funt S, Brown GL, Virkkunen M, Goldman D. A functional polymorphism in the MAOA gene promoter (MAOA-LPR) predicts central dopamine function and body mass index. Mol Psychiatry. 2006;11:858–866. doi: 10.1038/sj.mp.4001856. [DOI] [PubMed] [Google Scholar]
- Enoch MA. The role of early life stress as a predictor for alcohol and drug dependence. Psychopharmacology (Berl) 2011;214:17–31. doi: 10.1007/s00213-010-1916-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enoch MA, Steer CD, Newman TK, Gibson N, Goldman D. Early life stress, MAOA, and gene-environment interactions predict behavioral disinhibition in children. Genes Brain Behav. 2010;9:65–74. doi: 10.1111/j.1601-183X.2009.00535.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eysenck HJ, E MW. Personality and individual differences: A natural science approach. NY: Plenum: 1985. [Google Scholar]
- Eysenck MW. Arousal and speed of recall. Br J Soc Clin Psychol. 1975a;14:269–277. doi: 10.1111/j.2044-8260.1975.tb00179.x. [DOI] [PubMed] [Google Scholar]
- Eysenck MW. Extraversion, arousal, and speed of retrieval from secondary storage. J Pers. 1975b;43:390–401. doi: 10.1111/j.1467-6494.1975.tb00712.x. [DOI] [PubMed] [Google Scholar]
- Frazzetto G, Di Lorenzo G, Carola V, Proietti L, Sokolowska E, Siracusano A, Gross C, Troisi A. Early trauma and increased risk for physical aggression during adulthood: the moderating role of MAOA genotype. PloS one. 2007;2:e486. doi: 10.1371/journal.pone.0000486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garno JL, Gunawardane N, Goldberg JF. Predictors of trait aggression in bipolar disorder. Bipolar Disord. 2008;10:285–292. doi: 10.1111/j.1399-5618.2007.00489.x. [DOI] [PubMed] [Google Scholar]
- Guo G, Ou XM, Roettger M, Shih JC. The VNTR 2 repeat in MAOA and delinquent behavior in adolescence and young adulthood: associations and MAOA promoter activity. Eur J Hum Genet. 2008;16:626–634. doi: 10.1038/sj.ejhg.5201999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haberstick BC, Lessem JM, Hewitt JK, Smolen A, Hopfer CJ, Halpern CT, Killeya-Jones LA, Boardman JD, Tabor J, Siegler IC, Williams RB, Mullan Harris K. MAOA genotype, childhood maltreatment, and their interaction in the etiology of adult antisocial behaviors. Biological psychiatry. 2014;75:25–30. doi: 10.1016/j.biopsych.2013.03.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkins R, Bhugra D, Meltzer H, Singleton N, Bebbington P, Brugha T, Coid J, Farrell M, Lewis G, Paton J. Psychiatric and social aspects of suicidal behaviour in prisons. Psychological medicine. 2005;35:257–269. doi: 10.1017/s0033291704002958. [DOI] [PubMed] [Google Scholar]
- Kendler KS, Aggen SH, Czajkowski N, Roysamb E, Tambs K, Torgersen S, Neale MC, Reichborn-Kjennerud T. The structure of genetic and environmental risk factors for DSM-IV personality disorders: a multivariate twin study. Arch Gen Psychiatry. 2008;65:1438–1446. doi: 10.1001/archpsyc.65.12.1438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovallo WR. Early life adversity reduces stress reactivity and enhances impulsive behavior: implications for health behaviors. Int J Psychophysiol. 2013;90:8–16. doi: 10.1016/j.ijpsycho.2012.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyons MJ, True WR, Eisen SA, Goldberg J, Meyer JM, Faraone SV, Eaves LJ, Tsuang MT. Differential heritability of adult and juvenile antisocial traits. Arch Gen Psychiatry. 1995;52:906–915. doi: 10.1001/archpsyc.1995.03950230020005. [DOI] [PubMed] [Google Scholar]
- Mann JJ, Arango VA, Avenevoli S, Brent DA, Champagne FA, Clayton P, Currier D, Dougherty DM, Haghighi F, Hodge SE, Kleinman J, Lehner T, McMahon F, Moscicki EK, Oquendo MA, Pandey GN, Pearson J, Stanley B, Terwilliger J, Wenzel A. Candidate endophenotypes for genetic studies of suicidal behavior. Biological psychiatry. 2009;65:556–563. doi: 10.1016/j.biopsych.2008.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manuck SB, Flory JD, Ferrell RE, Mann JJ, Muldoon MF. A regulatory polymorphism of the monoamine oxidase-A gene may be associated with variability in aggression, impulsivity, and central nervous system serotonergic responsivity. Psychiatry Res. 2000;95:9–23. doi: 10.1016/s0165-1781(00)00162-1. [DOI] [PubMed] [Google Scholar]
- Meyer-Lindenberg A, Buckholtz JW, Kolachana B, A RH, Pezawas L, Blasi G, Wabnitz A, Honea R, Verchinski B, Callicott JH, Egan M, Mattay V, Weinberger DR. Neural mechanisms of genetic risk for impulsivity and violence in humans. Proc Natl Acad Sci U S A. 2006;103:6269–6274. doi: 10.1073/pnas.0511311103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ni X, Sicard T, Bulgin N, Bismil R, Chan K, McMain S, Kennedy JL. Monoamine oxidase a gene is associated with borderline personality disorder. Psychiatr Genet. 2007;17:153–157. doi: 10.1097/YPG.0b013e328016831c. [DOI] [PubMed] [Google Scholar]
- Ohlsson IM, Ireland JL. Aggression and offence motivation in prisoners: exploring the components of motivation in an adult male sample. Aggressive behavior. 2011;37:278–288. doi: 10.1002/ab.20386. [DOI] [PubMed] [Google Scholar]
- Passamonti L, Fera F, Magariello A, Cerasa A, Gioia MC, Muglia M, Nicoletti G, Gallo O, Provinciali L, Quattrone A. Monoamine oxidase-a genetic variations influence brain activity associated with inhibitory control: new insight into the neural correlates of impulsivity. Biological psychiatry. 2006;59:334–340. doi: 10.1016/j.biopsych.2005.07.027. [DOI] [PubMed] [Google Scholar]
- Patton JH, Stanford MS, Barratt ES. Factor structure of the Barratt impulsiveness scale. Journal of clinical psychology. 1995;51:768–774. doi: 10.1002/1097-4679(199511)51:6<768::aid-jclp2270510607>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- Reif A, Rosler M, Freitag CM, Schneider M, Eujen A, Kissling C, Wenzler D, Jacob CP, Retz-Junginger P, Thome J, Lesch KP, Retz W. Nature and nurture predispose to violent behavior: serotonergic genes and adverse childhood environment. Neuropsychopharmacology. 2007;32:2375–2383. doi: 10.1038/sj.npp.1301359. [DOI] [PubMed] [Google Scholar]
- Rivlin A, Hawton K, Marzano L, Fazel S. Psychosocial characteristics and social networks of suicidal prisoners: towards a model of suicidal behaviour in detention. PloS one. 2013;8:e68944. doi: 10.1371/journal.pone.0068944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy A, Gorodetsky E, Yuan Q, Goldman D, Enoch MA. Interaction of FKBP5, a stress-related gene, with childhood trauma increases the risk for attempting suicide. Neuropsychopharmacology. 2010;35:1674–1683. doi: 10.1038/npp.2009.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabol SZ, Hu S, Hamer D. A functional polymorphism in the monoamine oxidase A gene promoter. Hum Genet. 1998;103:273–279. doi: 10.1007/s004390050816. [DOI] [PubMed] [Google Scholar]
- Sakelliadis EI, Papadodima SA, Sergentanis TN, Giotakos O, Spiliopoulou CA. Self-injurious behavior among Greek male prisoners: prevalence and risk factors. European psychiatry : the journal of the Association of European Psychiatrists. 2010;25:151–158. doi: 10.1016/j.eurpsy.2009.07.014. [DOI] [PubMed] [Google Scholar]
- Sarchiapone M, Carli V, Cuomo C, Marchetti M, Roy A. Association between childhood trauma and aggression in male prisoners. Psychiatry Res. 2009;165:187–192. doi: 10.1016/j.psychres.2008.04.026. [DOI] [PubMed] [Google Scholar]
- Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, Hergueta T, Baker R, Dunbar GC. The Mini-International Neuropsychiatric Interview (M.I.N.I.): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry. 1998;59(Suppl 20):22–33. quiz 34–57. [PubMed] [Google Scholar]
- Soliman A, Bagby RM, Wilson AA, Miler L, Clark M, Rusjan P, Sacher J, Houle S, Meyer JH. Relationship of monoamine oxidase A binding to adaptive and maladaptive personality traits. Psychological medicine. 2011;41:1051–1060. doi: 10.1017/S0033291710001601. [DOI] [PubMed] [Google Scholar]
- Suris A, Lind L, Emmett G, Borman PD, Kashner M, Barratt ES. Measures of aggressive behavior: Overview of clinical and research instruments. Aggress Violent Beh. 2004;9:165–227. [Google Scholar]
- Tikkanen R, Ducci F, Goldman D, Holi M, Lindberg N, Tiihonen J, Virkkunen M. MAOA alters the effects of heavy drinking and childhood physical abuse on risk for severe impulsive acts of violence among alcoholic violent offenders. Alcohol Clin Exp Res. 2010;34:853–860. doi: 10.1111/j.1530-0277.2010.01157.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torgersen S, Czajkowski N, Jacobson K, Reichborn-Kjennerud T, Roysamb E, Neale MC, Kendler KS. Dimensional representations of DSM-IV cluster B personality disorders in a population-based sample of Norwegian twins: a multivariate study. Psychological medicine. 2008;38:1617–1625. doi: 10.1017/S0033291708002924. [DOI] [PubMed] [Google Scholar]
- Walker EA, Unutzer J, Rutter C, Gelfand A, Saunders K, VonKorff M, Koss MP, Katon W. Costs of health care use by women HMO members with a history of childhood abuse and neglect. Arch Gen Psychiatry. 1999;56:609–613. doi: 10.1001/archpsyc.56.7.609. [DOI] [PubMed] [Google Scholar]
- Weder N, Yang BZ, Douglas-Palumberi H, Massey J, Krystal JH, Gelernter J, Kaufman J. MAOA genotype, maltreatment, and aggressive behavior: the changing impact of genotype at varying levels of trauma. Biological psychiatry. 2009;65:417–424. doi: 10.1016/j.biopsych.2008.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widom CS. The cycle of violence. Science. 1989;244:160–166. doi: 10.1126/science.2704995. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.