

Abstract
Fluorescence in situ hybridization (FISH) is a name given to a variety of techniques commonly used for visualizing gene transcripts in eukaryotic cells and can be further modified to visualize other components in the cell such as infection with viruses and bacteria. Spatial localization and visualization of viruses and bacteria during the infection process is an essential step that complements expression profiling experiments such as microarrays and RNAseq in response to different stimuli. Understanding the spatiotemporal infections with these agents complements biological experiments aimed at understanding their interaction with cellular components. Several techniques for visualizing viruses and bacteria such as reporter gene systems or immunohistochemical methods are time-consuming, and some are limited to work with model organisms and involve complex methodologies. FISH that targets RNA or DNA species in the cell is a relatively easy and fast method for studying spatiotemporal localization of genes and for diagnostic purposes. This method can be robust and relatively easy to implement when the protocols employ short hybridizing, commercially-purchased probes, which are not expensive. This is particularly robust when sample preparation, fixation, hybridization, and microscopic visualization do not involve complex steps. Here we describe a protocol for localization of bacteria and viruses in insect and plant tissues. The method is based on simple preparation, fixation, and hybridization of insect whole mounts and dissected organs or hand-made plant sections, with 20 base pairs short DNA probes conjugated to fluorescent dyes on their 5' or 3' ends. This protocol has been successfully applied to a number of insect and plant tissues, and can be used to analyze expression of mRNAs or other RNA or DNA species in the cell.
Keywords: Infection, Issue 84, FISH, localization, insect, plant, virus, endosymbiont, transcript, fixation, confocal microscopy
Introduction
When studying the interactions between plant viruses and other pathogens with their infected plant hosts, it is important to visualize the pathogens and their respective nucleic acids in situ, regardless whether they cause negative effects on their hosts. This is most important when studying pathogen movement within and between plant cells. In situ localization of gene products of the pathogen is an essential step that complements other approaches for studying the pathogenicity process. Many plant pathogens, especially viruses, are transmitted by insects, having complex and intimate interactions with their vectors. Localization of these viruses in their vectors is important for studying the path for transmission, and the possible interaction sites inside the vector. The transmission of some insect-vectored plant viruses is aided by endosymbiotic bacteria that reside within these insects. To better study the transmission of these plant viruses by their vectors, it is also essential to visualize the endosymbiotic bacteria in general, and those involved in virus transmission, in particular. Colocalization of the virus and endosymbiotic bacteria is thus desired for investigating the possible relationships between these organisms within their insect host. Aside from virus transmission, endosymbiotic bacteria influence several aspects of the biology of insect vectors, thus spatial localization of these bacteria within insects is of high interest and importance.
Tomato yellow leaf curl virus (TYLCV) (Begomovirus, Geminiviridae) is the most important viral disease complex of cultivated tomato worldwide1-4. TYLCV is a phloem-limited virus and is exclusively vectored by the whitefly Bemisia tabaci5,6. A model describing the translocation of begomoviruses in their whitefly vectors has been proposed6-9. Two specific barriers are actively crossed during this circulative transmission: the midgut/hemolymph and the hemolymph/salivary gland barriers. This transmission process is hypothesized to be mediated by unknown receptors which recognize the virus capsid. TYLCV is thought to cross B. tabaci midgut6,10, and is absorbed through the primary salivary gland6 before it is injected into the plant. In the whitefly hemolymph, TYLCV interacts with a GroEL protein produced by the insect secondary endosymbiotic bacterium Hamiltonella. This interaction ensures safe transport of TYLCV in the hemolymph, and protects it from attack by the insect immune system11. Hamiltonella and Portiera, the primary endosymbiont of whiteflies, are housed in bacteriocytes, insect cells found in the hemolymph and house endosymbiotic bacteria11. B. tabaci harbors additional endosymbiotic bacteria including Rickettsia, Arsenophonus, Wolbachia, and Fritschea, which can be localized inside or outside the bacteriocytes, and have diverse effects on the insect’s biology12.
Several reports have attempted to study the localization of TYLCV in plants and whiteflies by using time-consuming and costly protocols such as Transmission Electron Microscopy (TEM), antibodies and RNA in situ on microscopic thick section10,13, a recent study described the localization of plant viruses inside their plant and vector hosts by using a simple protocol14. Here we describe a simple protocol for the localization of TYLCV in B. tabaci dissected midguts and salivary glands, and in sections prepared from TYLCV-infected plants. We further describe the localization of Portiera, the primary endosymbiont of B. tabaci, and its secondary endosymbionts Hamitonella, Rickettsia and Arsenophonus. This protocol is based on using short DNA probes, that are fluorescently labeled on their 5' end, and specifically hybridize to complementary sequences in viral or bacterial gene sequences. The sample processing is relatively easy and the signal obtained is highly specific. The described protocol can be used to localize viruses, bacteria and other pathogens in their plant, animal and insect hosts, and can further be used to localize mRNA in any given tissue.
Protocol
1. General Whitefly, Plant, and Virus Preparations for FISH Analysis
Rear B and Q biotype whiteflies on cotton seedlings (Gossypium hirsutum L. cv. Acala) and maintain inside insect-proof cages and growth rooms under standard conditions of 25±2 °C, 60% relative humidity, and a 14-hr light/10-hr dark photoperiod.
Perform PCR to test infection with endosymbionts using endosymbiont-specific primers as previously described15-19.
Purchase Tomato seedlings (Solanum esculentum cv. Beefsteak) from a commercial nursery or plant tomato seeds.
Maintain under the same rearing conditions described above until the plants reach the four true-leaf stage, the age most suitable for whitefly-mediated inoculation of TYLCV15 (see section 4.1 below).
2. Insect Handling, Fixation, Probe Design, Hybridization, and Visualization for Endosymbiont FISH
Collect 10 adult whiteflies by aspiration from cotton plants, or pick 10 3rd or 4th instar nymphs from cotton leaves.
Immediately immerse in Carnoy’s fixative (chloroform-ethanol-glacial acetic acid, 6:3:1, vol/vol) inside Eppendorf tubes. Fix for 2 hr to overnight (preferably overnight).
Completely remove the fixative and decolorize in 6% H2O2 in ethanol for 2 hr.
Preserve the samples in absolute ethanol for several days to several weeks at room temperature, or proceed to the next step.
Design probes as complementary reverse DNA primers based on the 16s ribosomal RNA genes of each bacterium. For designing the probes, take into the account the same considerations for designing PCR primers. Use the oligonucleotide probes BTP1 5'-Cy5-TGTCAGTGTCAGCCCAGAAG-3', Rb1 5'-Cy3-TCCACGTCGCCGTCTTGC-3', BTH 5'-Cy3-CCAGATTCCCAGACTTTACTCA-3' and Ars2 5'-cy3-TCATGACCACAACCTCCAAA-3' for specific targeting of Portiera, Rickettsia, Hamilonella, and Arsenophonus, respectively.
Hybridize samples overnight in the dark in hybridization buffer (20 mM Tris-HCl, pH 8.0, 0.9 M NaCl, 0.01% [wt/vol] sodium dodecyl sulfate, 30% [vol/vol] formamide) containing 10 pmol fluorescent probe per ml. Combine more than one probe with different fluorescent dyes if necessary. Use noninfected whiteflies with the targeted bacterium and no-probe samples as negative controls.
Mount the samples whole, on a microscope slide, in hybridization buffer contained with a liquid blocker, cover with a cover slip, seal with nail polish, and view under a fluorescence or confocal microscope.
3. Whitefly Organ Dissection, Fixation, Hybridization, and Visualization for TYLCV FISH
Collect adult whiteflies continuously reared on TYLCV-infected tomato plants by aspiration and anesthetize by exposure to paper towel impregnated with acetone. Expose whiteflies only for 1-2 min as too long exposure to acetone can fix whiteflies tissues. Use nonviruliferous whiteflies reared on noninfected tomato plants and no probe samples for controls to confirm specific hybridization.
Mount whiteflies on depression microscope slides for organ dissection using a stereo microscope.
Pull away the insect head from the prothorax and dissect salivary glands in 1x PBS supplemented with 1% toluidine blue stain for better visualization, due to their small size. Allow absorption of the stain for 2-3 min.
Pull the insect apart at the juncture between the thorax and the abdomen to observe and expel out the midgut. Expel the contents of the abdomen by pushing with forceps on the abdomen tip.
Gently remove the PBS and toluidine blue from successful dissections and gently add 300 µl of Carnoy’s fixative to fix the organs for 5 min.
Add the fixative from one side of the depressed well and not from the top to prevent uncontrolled movement of the dissected organs. Once touching the fixative, the organs will adhere to the glass, and throughout the process they will not move.
Remove fixative and add 500 µl of hybridization buffer supplemented with 10 pmol of the fluorescent DNA probe Cy5-5'-GGAACATCAGGGCTTCGATA-3', which is complementary to a sequence in TYLCV coat protein gene.
Hybridize overnight at room temperature in the dark by incubating the slide with the specimen inside a small humid chamber composed of small plastic box with wet towel paper at the bottom.
Following the hybridization, pick up the organs with a fine dissection tool and rapidly move them to a fresh microscope slide containing 30 µl of new hybridization buffer supplemented with DAPI (0.1 mg/ml in 1x PBS) and contained with liquid blocker.
Cover the specimens with a cover slip, seal with nail polish and view under a fluorescence or confocal microscope.
4. Plant Handling, Hand-sectioning, Fixation, and Hybridization for TYLCV FISH
Inoculate tomato seedlings with TYLCV using viruliferous whitefly-mediated inoculation. The virus can be detected using this FISH method in symptomless plants one week after inoculation. Typical disease symptoms are visible three weeks post inoculation. Use noninfected tomato plants and no probe samples for controls to confirm specific hybridization.
Use a sharp histological razor blade for preparing hand-cut 2-4 cm long longitudinal or cross sections of tomato stems and leaves.
In Eppendorf tubes, fix the sections in Carnoy’s fixative for 2 hr to overnight at room temperature.
Remove fixative and preserve the sections in absolute ethanol for several days to several weeks at room temperature, or proceed to the next step.
Wash the sections three times, 1 min each, in hybridization buffer.
Hybridize the sections with 500 µl of hybridization buffer supplemented with 10 pmol of the fluorescent oligonucleotide probe Cy3-5'-GGAACATCAGGGCTTCGATA-3' which is complementary to a sequence in TYLCV coat protein gene.
Wash the sections three times, 1 min each, in hybridization buffer.
Mount sections whole in hybridization buffer supplemented with DAPI (0.1 mg/ml in 1x PBS) and contained with liquid blocker, cover with a coverslip, seal with nail polish and view under a fluorescence or confocal microscope.
Representative Results
The system studied in this manuscript is shown in Figure 1 and includes an infected plant with TYLCV, an adult and a nymph of the whitefly B. tabaci, and the internal anatomy of the whitefly showing the path for TYLCV translocation in the insect. Figure 2 shows double FISH in an adult whitefly for the primary symbiont Portiera and the secondary symbiont Arsenophonus. Figure 3 shows double FISH for Portiera and Hamiltonella, and Figure 4 shows double FISH for Portiera and Rickettsia, both figures in B. tabaci 4th instar nymph. Figure 5 shows FISH for TYLCV and DAPI staining in a whitefly dissected midgut, and Figure 6 shows a FISH for TYLCV and DAPI staining in a dissected whitefly primary salivary glands. Figure 7 shows FISH for TYLCV and DAPI staining in hand-cut TYLCV-infected plant sections.
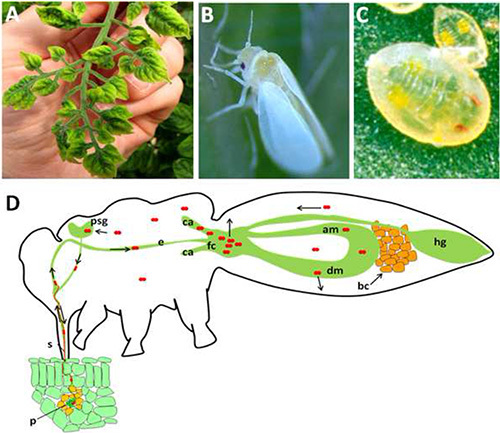 Figure 1. General overview of the system studied in this manuscript.
A) Typical disease symptoms caused by infection with Tomato yellow leaf curl virus (TYLCV). B) External view of an adult whitefly. C) External view of the 4th nymphal stage in the whitefly developmental life cycle. Other nymphal stages are similar in the general external view but smaller in size. D) Schematic view of the internal anatomy of an adult B. tabaci showing the path of TYLCV acquisition from the plant sieve elements, circulation and transmission (black arrows). Red particles refer to TYLCV virions. p: phloem; s: stylet; e: esophagus; fc: filter chamber; dm: descending midgut; am: ascending midgut; ca: caeca; hg: hindgut; bc: bacteriocytes; psg: primary salivary gland. Click here to view larger image.
Figure 1. General overview of the system studied in this manuscript.
A) Typical disease symptoms caused by infection with Tomato yellow leaf curl virus (TYLCV). B) External view of an adult whitefly. C) External view of the 4th nymphal stage in the whitefly developmental life cycle. Other nymphal stages are similar in the general external view but smaller in size. D) Schematic view of the internal anatomy of an adult B. tabaci showing the path of TYLCV acquisition from the plant sieve elements, circulation and transmission (black arrows). Red particles refer to TYLCV virions. p: phloem; s: stylet; e: esophagus; fc: filter chamber; dm: descending midgut; am: ascending midgut; ca: caeca; hg: hindgut; bc: bacteriocytes; psg: primary salivary gland. Click here to view larger image.
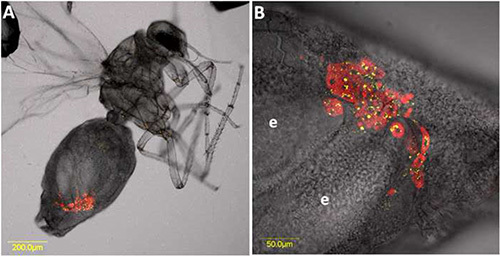 Figure 2. Double FISH of Portiera and Arsenphonus on B. tabaci Q biotype adult (A) and zoom on the labeled area (B) both under bright field channel and combined optical sections detecting the fluorescent signals emitted by each probe. Portiera-specific probe (red) conjugated to Cy5 and Arsenphonus-specific probe (yellow) conjugated to Cy3 were used. e: egg. Click here to view larger image.
Figure 2. Double FISH of Portiera and Arsenphonus on B. tabaci Q biotype adult (A) and zoom on the labeled area (B) both under bright field channel and combined optical sections detecting the fluorescent signals emitted by each probe. Portiera-specific probe (red) conjugated to Cy5 and Arsenphonus-specific probe (yellow) conjugated to Cy3 were used. e: egg. Click here to view larger image.
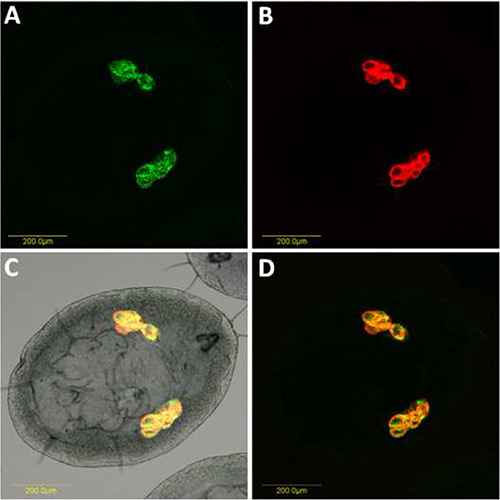 Figure 3. Double FISH of Portiera and Hamiltonella on B. tabaci nymph. Portiera-specific probe (red) conjugated to Cy5 and Hamiltonella-specific probe (green) conjugated to Cy3 were used. A) Hamiltonella hybridization signal obtained from combined optical sections and viewed under dark field. B) Portiera hybridization signal obtained from combined optical sections and viewed under dark field. C) Portiera and Hamiltonella merged signals under bright field. D) Portiera and Hamiltonella combined signals viewed under dark field. Click here to view larger image.
Figure 3. Double FISH of Portiera and Hamiltonella on B. tabaci nymph. Portiera-specific probe (red) conjugated to Cy5 and Hamiltonella-specific probe (green) conjugated to Cy3 were used. A) Hamiltonella hybridization signal obtained from combined optical sections and viewed under dark field. B) Portiera hybridization signal obtained from combined optical sections and viewed under dark field. C) Portiera and Hamiltonella merged signals under bright field. D) Portiera and Hamiltonella combined signals viewed under dark field. Click here to view larger image.
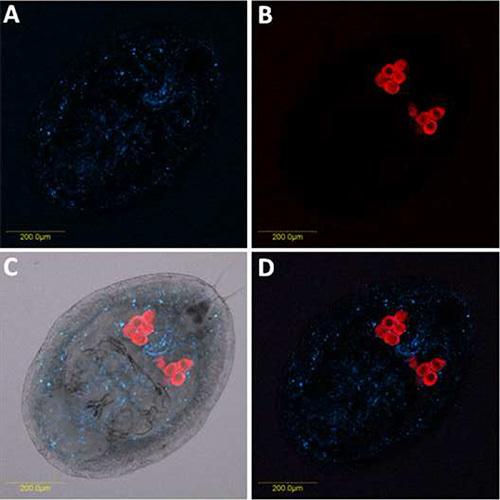 Figure 4. Double FISH of Portiera and Rickettsia on B. tabaci nymph. Portiera-specific probe (red) conjugated to Cy5 and Rickettsia-specific probe (blue) conjugated to Cy3 were used. A) Rickettsia hybridization signal obtained from combined optical sections and viewed under dark field. B) Portiera hybridization signal obtained from combined optical sections and viewed under dark field. C) Portiera and Rickettsia combined signals under bright field. D) Portiera and Rickettsia combined signals under dark field. Click here to view larger image.
Figure 4. Double FISH of Portiera and Rickettsia on B. tabaci nymph. Portiera-specific probe (red) conjugated to Cy5 and Rickettsia-specific probe (blue) conjugated to Cy3 were used. A) Rickettsia hybridization signal obtained from combined optical sections and viewed under dark field. B) Portiera hybridization signal obtained from combined optical sections and viewed under dark field. C) Portiera and Rickettsia combined signals under bright field. D) Portiera and Rickettsia combined signals under dark field. Click here to view larger image.
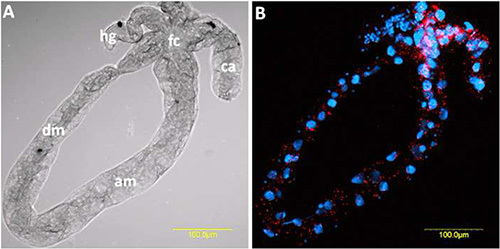 Figure 5. FISH of TYLCV on dissected midgut of an adult B. tabaci female that had acquired the virus for 48 hr from an infected tomato plant. TYLCV-specific probe (red) conjugated to Cy3 was used, and the midgut was stained with DAPI before mounting and visualization. A) The dissected midgut as seen from combined optical sections under bright field. B) TYLCV FISH signal (red) and DAPI stained nuclei (blue) as seen from combined optical sections under dark field. fc: filter chamber; dm: descending midgut; am: ascending midgut; ca: caeca; hg: hindgut. Click here to view larger image.
Figure 5. FISH of TYLCV on dissected midgut of an adult B. tabaci female that had acquired the virus for 48 hr from an infected tomato plant. TYLCV-specific probe (red) conjugated to Cy3 was used, and the midgut was stained with DAPI before mounting and visualization. A) The dissected midgut as seen from combined optical sections under bright field. B) TYLCV FISH signal (red) and DAPI stained nuclei (blue) as seen from combined optical sections under dark field. fc: filter chamber; dm: descending midgut; am: ascending midgut; ca: caeca; hg: hindgut. Click here to view larger image.
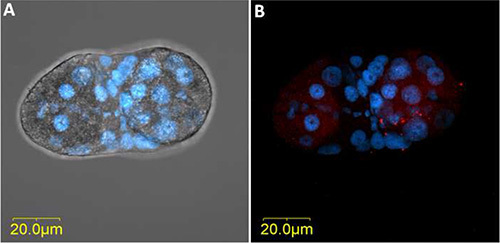 Figure 6. FISH of TYLCV on dissected primary salivary gland of an adult B. tabaci female that had acquired the virus for 48 hr from an infected tomato plant. TYLCV-specific probe (red) conjugated to Cy3 was used, and the salivary gland stained with DAPI before mounting and visualization. A) The dissected and DAPI stained primary salivary gland as seen from combined sections under bright field. B) TYLCV FISH signal (red) and DAPI stained nuclei (blue) as seen from combined sections under dark field. Click here to view larger image.
Figure 6. FISH of TYLCV on dissected primary salivary gland of an adult B. tabaci female that had acquired the virus for 48 hr from an infected tomato plant. TYLCV-specific probe (red) conjugated to Cy3 was used, and the salivary gland stained with DAPI before mounting and visualization. A) The dissected and DAPI stained primary salivary gland as seen from combined sections under bright field. B) TYLCV FISH signal (red) and DAPI stained nuclei (blue) as seen from combined sections under dark field. Click here to view larger image.
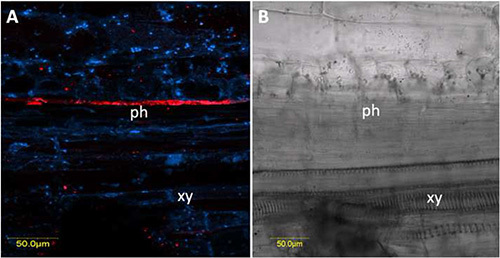 Figure 7. FISH of TYLCV on hand-cut longitudinal section through a stem of TYLCV-infected plant. TYLCV-specific probe (red) conjugated to Cy3 was used and combined with DAPI staining before mounting and visualization. A) One section shown with red TYLCV signal in a phloem sieve element, and DAPI stained nuclei seen from combined sections under dark field. B) The same combined sections shown in (A) under bright field for orientation. ph: phloem; xy: xylem. Click here to view larger image.
Figure 7. FISH of TYLCV on hand-cut longitudinal section through a stem of TYLCV-infected plant. TYLCV-specific probe (red) conjugated to Cy3 was used and combined with DAPI staining before mounting and visualization. A) One section shown with red TYLCV signal in a phloem sieve element, and DAPI stained nuclei seen from combined sections under dark field. B) The same combined sections shown in (A) under bright field for orientation. ph: phloem; xy: xylem. Click here to view larger image.
Discussion
The protocol described here for the localization of a plant virus in its plant host and insect vector, and endosymbiotic bacteria in their specific whitefly host, can be adapted for the localization of other viruses in plants and even in animal tissues. Furthermore, the protocol can be used to localize endosymbiotic and pathogenic bacteria and other microorganisms in plant and animal systems. The described methods rely on simple concept of hybridization between a short, fluorescently-labeled oligonucleotide DNA probe and the target DNA or RNA molecule in the cell. The outcome is a specific hybridization, with minimum background, and a signal detected using fluorescence or confocal microscopes. Several probes with different fluorescent dyes, that target more than one gene, can be used simultaneously. This enables visualizing more than one target in a single cell or tissue. The procedures described in these protocols are simple and the sample processing time is minimal. Problems associated with other protocols such as high background signal, long processing time for specimens and the use of numerous and costly materials for the analysis, are not constraints in the protocol described here.
The B and Q biotypes of B. tabaci whiteflies were used to describe the protocols in this manuscript; however, rearing conditions apply to any whitefly biotype, with their respective host plant. The whitefly completes its life cycle in three weeks under the indicated conditions. Each individual of the B biotype population used was infected with the primary endosymbiont Portiera, and the secondary endosymbiotic bacteria Hamiltonella and Rickettsia. The Q biotype individuals were infected with Portiera and Arsenophonus. Other whitefly populations around the world were shown to be infected with these or other endosymbiotic bacterial species, and with different spatial localization patterns in the body16-20.
TYLCV infected plants show typical disease symptoms three weeks following the inoculation. For TYLCV localization, symptomatic plants can be used for virus acquisition by whiteflies for 24 hr and the virus can be detected in infected plants and insects using the FISH protocol described above. While in insects the probes used can be conjugated with many fluorophore which can excised with different wavelengths, including Cy3 and Cy5, in plants Cy3 is the most suitable and does not fluoresce at similar wavelengths as chloroplasts, the most abundant organelles in the plant cell.
Successful implementation of the described protocols for the localization of bacteria and viruses in plants and insects depends on good fixation of the samples for better visualization and probe penetration, good probe design for specificity and clearing with H2O2 in the case of whole insects for better signal visualization. In the case of FISH in plants, hand-made sections must be as thin as possible for good probe penetration and better visualization of the signal. This can be improved by putting the leaf between two Styrofoam pieces to help with cutting and by using professional razor blades which can be purchased for this purpose from several vendors.
The protocol is not suitable for subcellular localization of targets, however it has the potential to be developed, using the probe format describe here, into a suitable method for subcellular localization. For example, the short probes described here can be conjugated to molecules such as biotin and not fluorescent molecules, which in turn can be targeted using gold particles conjugated to streptavidin that can be visualized under TEM. This later modification can be suitable for subcellular localization of several targets including gene transcripts and microorganisms. Finally, the described protocols are suitable for hybridization with nucleic acids such as DNA and RNA but not proteins.
For obtaining the best results in the FISH protocols described here, it is best to use fresh insect and plant material, as well as fresh fixative and hybridization buffers. The probes should always be kept in -20 ºC in small aliquots to avoid repeated freezing and thawing. The processing time of each described step can be calibrated depending on the organisms studied. As mentioned in the protocol, following fixation and decolorization, whitefly and plant samples can be preserved for long time in absolute ethanol at room temperature. This step is particularly useful when sending samples for processing from one location to another, or when performing long time course experiments. The quality of the hybridization signal was not affected following this long time preservation.
Disclosures
The authors have nothing to disclose.
Acknowledgments
Research in the Ghanim lab was supported by research grant no. 908-42.12/2006 from the German-Israeli Foundation (GIF), grant no. IS-4062-07 from the United States-Israel Binational Agricultural Research and Development Fund (BARD), and research grant no. 884/07 from Israel Science Foundation (ISF) to M.G.
References
- Czosnek H, Laterrot H. A worldwide survey of Tomato yellow leaf curl viruses. Arch. Virol. 1997;142:1391–1406. doi: 10.1007/s007050050168. [DOI] [PubMed] [Google Scholar]
- Ling KS, Simmons AM, Hassell RL, Keinath AP, Polston JE. First report of Tomato Yellow Leaf Curl Virus in South Carolina. Plant Dis. 2006;90 doi: 10.1094/PD-90-0379C. [DOI] [PubMed] [Google Scholar]
- Polston JE, McGovern RJ, Brown LG. Introduction of Tomato yellow leaf curl virus in Florida and implications for the spread of this and other geminiviruses of tomato. Plant Dis. 1999;83:984–988. doi: 10.1094/PDIS.1999.83.11.984. [DOI] [PubMed] [Google Scholar]
- Polston JE, Rosebrock TR, Sherwood T, Creswell T, Shoemaker PJ. Appearance of Tomato yellow leaf curl virus in North Carolina. Plant Dis. 2002;86 doi: 10.1094/PDIS.2002.86.1.73B. [DOI] [PubMed] [Google Scholar]
- Frohlich DR, Torres-Jerez I, Bedford ID, Markham PG, Brown JK. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol. Ecol. 2002;8:1683–1691. doi: 10.1046/j.1365-294x.1999.00754.x. [DOI] [PubMed] [Google Scholar]
- Ghanim M, Morin S, Czosnek H. Rate of Tomato yellow leaf curl virus translocation in the circulative transmission pathway of its vector, the whitefly Bemisia tabaci. Phytopathology. 2001;91:188–196. doi: 10.1094/PHYTO.2001.91.2.188. [DOI] [PubMed] [Google Scholar]
- Ghanim M, Rosell RC, Campbell LR, Czosnek H, Brown JK, Ullman DE. Digestive salivary and reproductive organs of Bemisia tabaci (Gennadius) Hemiptera: Aleyrodidae) B type. J. Morphol. 2001;248:22–40. doi: 10.1002/jmor.1018. [DOI] [PubMed] [Google Scholar]
- Hunter WB, Hiebert E, Webb SE, Tsai JK, Polston JE. Location of geminiviruses in the whitefly Bemisia tabaci (Homoptera: Aleyrodidae) Plant Dis. 1998;82:1147–1151. doi: 10.1094/PDIS.1998.82.10.1147. [DOI] [PubMed] [Google Scholar]
- Rosell RC, Torres-Jerez I, Brown JK. Temporal pathway of geminivirus in whitefly extracts, saliva, hemolymph and honeydew. Phytopathology. 1999;89:239–246. doi: 10.1094/PHYTO.1999.89.3.239. [DOI] [PubMed] [Google Scholar]
- Czosnek H, Ghanim M, Ghanim M. The circulative pathway of begomoviruses in the whitefly vector Bemisia tabaci—insights from studies with Tomato yellow leaf curl virus. Ann. Appl. Biol. 2002;140:215–231. [Google Scholar]
- Gottlieb Y, et al. The transmission efficiency of Tomato yellow leaf curl virusby the whitefly Bemisiatabaciis correlated with the presence of aspecificsymbiotic bacterium species. J. Virol. 2010;84:9310–9317. doi: 10.1128/JVI.00423-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brumin M, Levy M, Ghanim M. Transovarial transmission of Rickettsia spp. and organ-specific infection of the whitefly Bemisia tabaci. Appl. Environ. Microbiol. 2012;78:5565–5574. doi: 10.1128/AEM.01184-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghanim M, Medina V. In: Localization of Tomato yellow leaf curl virus in its whitefly vector Bemisia tabaci. In Tomato Yellow Leaf Curl Virus Disease, Management, Molecular Biology, Breeding for Resistance. Czosnek H, editor. New York: Springer; 2007. pp. 175–187. [Google Scholar]
- Ghanim M, Brumin M, Popovski S. A simple, rapid and inexpensive method for localization of Tomato yellow leaf curl virus and Potato leafroll virus in plant and insect vectors. J. Virol. Methods. 2009;159:311–314. doi: 10.1016/j.jviromet.2009.04.017. [DOI] [PubMed] [Google Scholar]
- Gotz M, et al. Implication of Bemisia tabaci heat shock protein 70 in Begomovirus-whitefly interactions. J. Virol. 2012;86:13241–13252. doi: 10.1128/JVI.00880-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nirgianaki A, et al. Wolbachia infections of the whitefly Bemisia tabaci. Curr. Microbiol. 2003;47:93–101. doi: 10.1007/s00284-002-3969-1. [DOI] [PubMed] [Google Scholar]
- Baumann P. Biology of bacteriocyte-associated endosymbionts of plant sap-sucking insects. Ann. Rev. Microbiol. 2005;59:155–189. doi: 10.1146/annurev.micro.59.030804.121041. [DOI] [PubMed] [Google Scholar]
- Gottlieb Y, et al. Identification and localization of a Rickettsia sp. in Bemisia tabaci (Homoptera) Aleyrodidae) App. Environ. Microbiol. 2006;72:3646–3652. doi: 10.1128/AEM.72.5.3646-3652.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li ZX, Lin HZ, Guo XP. Prevalence of Wolbachia infection in Bemisia tabaci. Curr. Microbiol. 2007;54:467–471. doi: 10.1007/s00284-007-0011-7. [DOI] [PubMed] [Google Scholar]
- Skaljac M, Zanic K, Hrncic S, Radonjic S, Perovic T, Ghanim M. Diversity and localization of bacterial symbionts in three whitefly species (Hemiptera: Aleyrodidae) from the east coast of the Adriatic. Bull. Entomol. Res. 2013;103:48–59. doi: 10.1017/S0007485312000399. [DOI] [PubMed] [Google Scholar]