

Abstract
European white stork (Ciconia ciconia) populations have been object to several conservation measures such as reintroduction programs, habitat improvement or supplementary feeding in the last decades. Although recent white stork censuses revealed an upward trend of most of the western populations, evaluations of the relative importance of certain conservation measures are still scarce or even lacking. In our study we analyzed the effect of supplementary feeding on the reproductive success of white storks in conjunction with other factors such as weather or nest site characteristics. We present data of 569 breeding events at 80 different nest sites located in variable distances to an artificial feeding site at Affenberg Salem (south-western Germany) collected from 1990–2012. A multilevel Poisson regression revealed that in our study population (1) reproductive success was negatively affected by monthly precipitation in April, May and June, (2) pairs breeding on power poles had a lower reproductive success than pairs breeding on platforms or trees and (3) reproductive success was significantly higher in pairs breeding in close distance to the feeding site. The number of fledglings per nest decreased by 8% per kilometer distance to the feeding site. Our data suggest that supplementary feeding increases fledgling populations which may be a tool to attenuate population losses caused by factors such as habitat deterioration or unfavorable conditions in wintering habitats.
Introduction
White stork (Ciconia ciconia) populations in Western Europe decreased tremendously after 1945 leading to a severe threat of extinction or even extinction in most West European countries [1]. Breeding white storks disappeared in Belgium, Switzerland (1950) and in Sweden (1955) [2], [3]. A strong decline of the population was observed in the Netherlands (down to 5 pairs 1984), Denmark (down to 6 pairs in 1996) and France (down to 11 pairs 1974) [4]–[6]. Environmental changes in breeding and wintering habitats have been discussed as main reasons for the severe decline of West European populations [7]–[11]. As a consequence conservation actions on a national level such as reintroduction projects, supplementary feeding and habitat improvement have been applied in several West European countries to prevent migrating white storks from extinction [8], [12]. Reintroduction activities included continuous releases of white storks reared in captivity, installation of nest sites (poles) and supplementary feeding of free flying individuals [12].
White stork censuses in 1995 and 2004 revealed an upward trend of most western populations [9], [13]. Causes for the recovery of western white stork populations are discussed multidimensional. Reintroduction of white storks reared in captivity and less severe droughts in West African wintering habitats are regarded as key aspects for the recovery of certain populations [9], . Additionally, alternative food resources in wintering habitats such as rubbish dumps or the copious availability of invasive species (e.g., red swamp or Louisiana crayfish, Procambarus clarkii) are discussed to influence population recovery of white storks [9], [15], [16]. However, some of these management efforts have been stopped because conservation programs are often limited in space and time. Yet, analyses to what degree different conservation actions such as supplementary feeding, habitat improvement and reintroduction contributed to the recovery of West European white stork population are still scarce (but see [17], [18], [19]) and, to our knowledge, the effect of additional feeding on reproductive success of white storks has never been investigated in detail.
In most of the west European white stork populations reproductive success is low or highly variable [1], [20]–[22]. Multiple factors may affect the number of fledglings, including age, arrival date, breeding experience of pairs and physical fitness of parent storks [19], [23], [24], weather condition during the nestling period [22], [25], [26], nest site characteristics [27] and food availability [10], [11], [19], [26], [28], which depends largely on habitat quality. Here certain human farming practices can have a major negative impact [29]–[31]. Food availability is considered as a limiting factor for breeding success and hence for population trends [10], [11], [19], [26], [28]. Supplementary feeding of free flying white storks as a conservation measure should therefore compensate for low habitat quality and increase reproductive success of pairs [19], [32], [33].
Supplementary feeding is a common element of wildlife management that can aid in the establishment of a viable, self-sustaining population by increasing body condition, growth rates, survival, social interactions or reproductive success (e.g. [34]–[38], but for possible negative effects see also [31], [39]). However, the effects of supplementary feeding on reproduction rates and life history of white storks have never been evaluated in detail. The aim of this study was to explore effects of supplementary feeding in conjunction with a number of other factors proposed to affect reproductive success in white storks such as weather [22] and nest site characteristics [27]. We present data collected over a period of 23 years on reproductive rates of free flying pairs of white storks breeding in variable distances to a supplementary feeding site at Affenberg Salem (Baden Württemberg, Germany).
White storks, while breeding, forage on average in an area of not more than 1500 m from the nest site [40] and Massemin-Challet et al. 2006 [19] showed that pairs breeding close to a rubbish dump (<1000 m) fledged more young than pairs breeding further away. We therefore predicted that storks, nesting closer to the supplementary feeding site at the Affenberg had a higher reproductive success, measured as number of fledglings per nest and year, than storks nesting in greater distances.
Methods
Study Area
The study was conducted at Affenberg Salem (N 47°45′43″ E 09°14′44″) Baden-Württemberg, Germany (Fig. 1). Affenberg Salem harbours one of the largest colonies of free flying white storks belonging to the western population in the southern part of Germany. The colony was founded at Affenberg Salem with nine reintroduced individuals in 1978. In the last 34 years, the colony attracted wild breeding storks, reaching a current size (2012) of 28 breeding pairs located directly at Affenberg Salem and 36 pairs breeding in the surroundings. Nest sites are located at variable distances (min distance 21 m, max distance 7527 m; Fig. 1) to the feeding site at Affenberg Salem and all breeding sites are encircled by a mixture of intensively cultivated fields, plantations, meadows and ponds. Within the study period (1990–2012) the colony consisted exclusively of free flying pairs.
Figure 1. Geographic position of white stork nest sites (red dots) in the vicinity of the Affenberg, Salem.

The yellow star indicates the artificial feeding site and the white lines encircle the feeding site with a distance of 0.3 km and 3.0 km, respectively (background image derives from Landsat 4–5 TM scene, path/row 194/27, 08.07.2010; http://glovis.usgs.gov/).
Food provisioning at Affenberg Salem
At a defined feeding site at Affenberg Salem, supplementary food (small fish and one-day old chicken) was provided twice a day (11.00 h and 16.45 h) during the breeding season (including the nestling period) and once a day (14.00 h) outside the breeding season. The food was distributed randomly over an area of approximately 1000 m2. The number of storks visiting the feeding site was highly variable depending on season (Hilgartner, unpublished data) and most likely also on food availability in the surroundings. During the breeding season number of storks visiting the feeding site was highest but at times dropped to 0. Outside the breeding season number of storks at the feeding site was in general considerably lower. The majority of the population is migrating and therefore not present during winter feeding. We tried to adjust the quantity of food so that 30 min after the food had been distributed no more food was left at the feeding site. As an estimate for adjustment, we used the time storks needed to empty the feeding site the respective day before. As consequence feeding time per day and the amount of food provisioned were limited. The amount ranged between (0–35 kg of chicken and 0–10 kg of small fish per feeding session). Storks were able to ingest up to 12 chickens per feeding into their crops and transport them to their nests (pers. obs. D. Zinner).
Data Collection
Breeding pairs were determined following standardized census methodology [9]. Breeding activity of all pairs was monitored regularly. If possible pairs were identified via metal or PVC rings. We don't have identities of all breeding pairs because not all individuals had rings and in some years/cases it was not possible to decipher the rings with certainty. Therefore, data on individual fitness of breeding storks, their ages or pair experience are not available. Data on clutch size, hatching success and chick mortality were collected ad libitum and were only available for a few pairs. As a measure for reproductive success we used number of fledged chicks per breeding pair (nest) per year. As potential predictor variables we included (1) distance between nest site and feeding site (km, determined in ArcGIS 10, ESRI Inc.), assuming that storks nesting closer to the feeding site will obtain supplementary food more regularly and in larger quantities, (2) weather data for the breeding and nestling periods, i.e., monthly rainfall and average temperature for April, May and June 1990–2012, obtained from the closest meteorological station of the Deutscher Wetterdienst in Konstanz, which is about 10 km from the feeding site and in addition (3) type of nest base: (a) on artificial platforms, (b) wild nests on houses and in trees (not particularly supported nests) or (c) on power poles. Data are available from Table S1.
Statistical Analysis
We used a multilevel Poisson regression to examine potential associations between the number of fledglings and the distance to the feeding site while controlling for other potential predictors variables. As covariates we assessed monthly rainfall and average temperature in April, May and June of respective years and nest base type. We included nest ID as a random factor to model the dependency of breeding success due to the same nest (as a proxy for intrinsic nest-site quality, individual breeding stork quality, breeding pair tenure or nest-site fidelity of parent storks, which were in fact not known in most cases).
In a first step, we included all independent variables in the model and subsequently removed variables with a p>0.1. We then compared this final model with (1) a model with replacing weather data with ‘year’ as a categorical independent variable (‘good stork years’ vs. ‘bad stork years’) and (2) a model with distance replaced by a categorical variable with either two (<300 m or >300 m) or three levels (<300 m, 300 m–3 km, >3 km) (Fig. 2). The three models are not nested and cannot be assessed by statistical tests. Therefore we compared the models using AIC [41].
Figure 2. Dotplot of distances between white stork nest sites and feeding site (used to determine cut-off points).
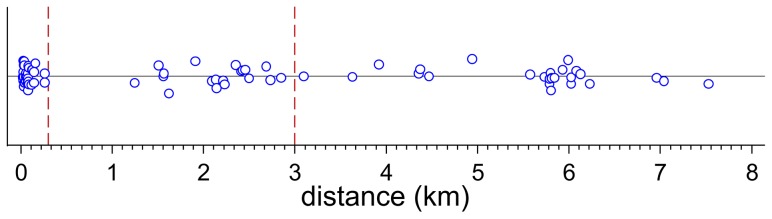
Each dot represents one nest site. Data points are slightly jittered (adding random noise to data) in order to prevent overplotting of data points. Vertical lines indicate cut-off distances.
A Poisson model assumes that the variance is equal to the mean. If there is more variation in the number of fledglings than predicted by the model (‘overdispersion’, e.g. due to more zero observations – in our case: no fledglings) standard errors are often underestimated and therefore p-values (and confidence intervals) become overoptimistic (type I error inflation). To account for possible overdispersion we used an observation–level random effect to model possible overdispersion assuming that overdispersion arises from errors taking on a log-normal mixing distribution [42], . This method also allows estimating and comparing the contribution of explained variance of the different variables [43]. We calculated the increase in the amount of variance in the response explained by adding a variable of interest, given that all other fixed and random variables are in the model (unique explained variance) using the approach described by Nakagawa and Schielzeth 2013 [44] for generalized linear mixed effects models. Incidence risk ratio (IRR) and 95% confidence intervals (95% CIs) of the final models are presented.
Ethical statement
Besides handling of white stork chicks for ring application, no other handling of storks was done. For all nests included in this study ring application was done by one of the authors (RH) and Walter Angst who hold a ringing permit issued by Regierungspräsidium Tübingen and Landratsamt Bodenseekreis (Germany). All handling of the storks was done in compliance with the laws of Baden-Württemberg (Germany). Number of fledglings was determined from a distance, if necessary with the help of binoculars. For these observational data collection no specific permission was required.
Results
Over a period of 23 consecutive years we collected data from 569 breeding events at 80 nest sites. The minimum number of nests per year was 10 in 1991. This number increased to 64 in 2012 (mean number of nests per year = 24.7, SD = 14.6). The number of fledglings per brood ranged from 0 to 5 (mean = 2.2, SD = 1.6, n = 569). ‘Bad years’ for stork reproduction were 1991 (0.9±1.3 fledglings per nest [mean ± SD, n = 10]) and 2007 (0.9±1.03 fledglings per nest [mean ± SD, n = 35]). In contrast, ‘good years’ were 2003 (3.0±1.6 fledglings per nest [mean ± SD, n = 22]) and 2011 (2.9±1.10 fledglings per nest [mean ± SD, n = 50]). On average 2.46 (SD = 1.53, N = 288) young storks fledged if breeding occurred in close proximity to the feeding site (<300 m); 1.99 (SD = 1.62, N = 148) chicks fledged in nests which were between 300 m and 3 km away form a feeding site and only 1.70 (SD = 1.45, N = 133) chicks fledged from nests if the distance to the feeding site was more than 3 km.
Our overall final model explained 28.7% of the variance. The fixed effects ‘nest base’, ‘distance to feeding site’ and ‘monthly precipitation in April, May and June’ explained 15.5% and the random factor ‘nest ID’ 13.2% (Table 1). Monthly mean temperatures did not predict significantly mean number of fledglings and were not part of the final model (all p>0.18) (Table 1). A model with year as a categorical variable instead of weather data resulted in a poor model (delta AIC compared to final model = −17.9). The final model suggests a negative relation between distance to feeding site and number of fledglings per nest and year (Fig. 3) with a reduction of the number of fledglings by 8% per kilometer distance from the feeding site (Incidence rate ratio = 0.92, unique r2 = 0.045).
Table 1. Results of multilevel Poisson regression with nest ID as a random factor.
| Variable | Incidence rate ratio (95% C.I.) | chi2 | z | p |
| nest base | 5.56 | 0.06 | ||
| power pole vs platform | 0.69 (0.51–0.94) | −2.35 | 0.019 | |
| wild vs. platform | 0.98 (0.75–1.29) | −0.15 | 0.88 | |
| power pole vs. wild | 0.708 (0.484–1.036) | 1.78 | 0.075 | |
| distance (km) | 0.920 (0.88–0.96) | −3.60 | <0.001 | |
| preci April (mm) | 0.997 (0.995–0.999) | −3.46 | <0.001 | |
| preci May (mm) | 0.996 (0.994–0.997) | −5.42 | <0.0001 | |
| preci June (mm) | 0.997 (0.995–0.999) | −3.19 | <0.001 |
Rate ratios (95% confidence intervals) are shown with chi2 or z test statistics and p values. An observation–level random effect was included to model possible overdispersion. Variance of i) random effects “nest ID”: 0.076 (95% C.I.: 0.035–0.17), p<0.05 and ii) observation-level random effect: 0 (exact 1.86e-23), n.s. The overall model explained 28.7% of the variance of which the random factor nest ID explained 13.2% and the fixed effects nest base, distance and precipitation in April, May and June 15.5%.
Figure 3. Number of fledglings per nest and year in relation to distance between the feeding place and nest site (km).

A Lowess (locally weighted scatterplot smoothing) curve is used to describe the trend of the data. Data points are slightly jittered (adding random noise to data) in order to prevent overplotting of data points.
In addition, monthly precipitation in April, May and June had an independent negative effect on the number of fledglings (combined unique r2 = 0.075). The type of nest-base tends to have an influence with nests on power poles having 31% less fledglings than storks breeding on platforms and 29% less fledglings than storks breeding in wild nests, whereas nests on platforms and wild nests on trees and buildings yielded similar numbers of fledglings (unique r2 for factor nest base = 0.02, Table 1).
Using distance as a categorical variable with two (<300 m or >300 m) and three levels (<300 m, 300 m–3 km, >3 km) resulted in a model with a slightly smaller AIC (2 levels: delta AIC = −0.77, 3 levels: delta AIC = −2.77) than the model with distance as a continuous variable. The 2-levels model suggests that storks breeding in a distance larger than 300 m from the feeding site have 29% less fledglings (IRR: 0.71 [95% C.I.: 0.585–0.86], z = −3.4, p = 0.001, unique r2 = 0.048) than storks breeding in a distance of less than 300 m from the feeding site. Using distance as a variable with three levels suggests that storks breeding in a distance of more than 3 km from the feeding site have 36% less fledglings (IRR: 0.64 [95% C.I. 0.49–0.82], z = −3.48, p<0.0001) than those breeding close to the feeding site (<300 m) while storks breeding in an intermediate distance (300 m–3000 m) have 20% less fledglings. But this difference is not significant compared to those living less than 300 m to the feeding site (IRR: 0.80 [95% C.I. 0.62–1.02], z = −1.79, p = 0.075). There is also no significant difference in the number of fledglings between those breeding in an intermediate and a long distance to the feeding site (IRR: 0.80 [95%C.I. 0.61–1.04], z = −1.64, p = 0.10). The distance factor accounted for 4.6% of the explained variance.
Discussion
Our study provides a first step to understand the effects of a particular conservation measure in a West European white stork population and to evaluate its consequences. We analyzed the effect of supplementary feeding on the reproductive success of white storks, which is a key variable in determining population trends [10], [11], [26], [28], [45], [46]. We included additional variables considered to influence reproductive success such as mean monthly temperature, monthly precipitation and type of nesting platforms and as a random factor ‘nest ID’ in our analyses.
The random factor ‘nest ID’ explains a large amount of the variance. Since we were not able to determine the identities of the breeding storks in all cases with certainty and the habitat quality surrounding the nests, we used this factor as a proxy for individual breeding stork quality and pair tenure, but also for quality of nest and the habitat around the nest, factors which are expected to influence reproductive success of breeding pairs [11], [21], [47]. It is therefore not surprising, that ‘nest ID’ has a significant impact on fledgling success in our study population.
Furthermore, environmental stressors such as extreme weather conditions affect survival rates in white storks [22] and indeed, breeding success was negatively affected by monthly precipitation in April, May and June but not by mean monthly temperatures. In our study population, most birds hatched in April and May with few outliers in March and June [Hilgartner & Angst unpublished data]. In the first three weeks after hatching, chicks do not have thermoregulatory abilities [48]. Even between three and five weeks after hatching, thermoregulation is still affected when chicks are too big to be protected by their parents but still have not fully developed their plumage [40], [49]. In accordance with our study Carrascal et al. [50] did not find an effect of mean spring temperatures on reproductive success, but number of rainy days was decisive for chick survival, with higher number of rainy days leading to lower chick survival. A number of other studies have also found a negative impact of rainy springs on the breeding success of white storks which seems to be general pattern across different populations [22], [26], [40].
The type of nest base tends to have an influence on reproductive success of white storks in our study population. White storks using nests of power poles had a lower number of fledglings than pairs breeding on platforms or wild nests. This effect is in contrast to the study of Tryjanowsky et al. [27] where no difference was found between the reproductive success of white storks nesting on power poles and other nesting sites such as chimneys, trees or roofs. A possible explanation for this difference maybe the yearly nest cleaning in autumn performed in our study colony. Nest cleaning involves the removal of the uppermost layer and the removal of rubbish e.g. strings, plastic etc. Nests on power poles, however, were excluded from cleaning as these nests are not easily accessible without presence of electricity suppliers and shutting off electricity. The cleaning may have an influence on survival of chicks as rubbish can have fatal effects on nestlings when they swallow plastic or get entangled in strings ([51]; Hilgartner pers. obs.). Moreover, nest cleaning may also lead to a better water permeability of the nest which could be advantageous for chick survival during long-lasting periods of rain.
Reproductive success of breeding white storks was significantly higher in nests in close distance to the artificial feeding site, as the number of fledglings decreased by 8% per kilometer distance from the feeding site. A similar observation was reported by Massemin-Challet et al. [19] who found that storks nesting near (<1000 m) a reliable artificial food source in Alsace produced significantly more fledglings than storks breeding further away. A plausible explanation for this negative correlation is that storks breeding closer to the feeding site made more frequently use of it and were able to supply their chicks with more food, which resulted in a higher reproduction. During breeding, white storks usually forage in close distance to their nest sites [20], [40], [47], [52], [53]. Moritzi et al. [40] found that 88% of all foraging records were within a radius of 1 km with a median distance of 380 m to the breeding colony. Hence, for white stork pairs not breeding in close distance to the feeding site greater flight expenditure might not compensate for supplementary food. Moreover, especially during the first weeks after hatching, at least one pair partner stays at the nest all the time for guarding the nestlings [54], [55]. Feeding times and food availability at the feeding site is limited. As a consequence pairs breeding in close distance to the feeding site are able to perform more successful provisioning flights than pairs of more distant nests. Hence, breeding storks in the population do not equally profit from supplementary feeding most probably leading to the observed distance related reproductive success in our stork colony.
A general impact of food availability on reproductive success in white storks has been found in a study by Tryanowski & Kuźniak [28]. They showed that reproductive success of white storks was higher in years with high food availability. A similar effect was found in pairs breeding in close distance to optimal feeding habitats such as meadows, pastures and wetlands [11], [52], [56]. Pairs breeding in close distance to such optimal feeding sites had a higher reproductive success than pairs breeding in suboptimal habitats. The intake of additional high quality food might increase the physical fitness of parent storks, in particular that of females and might therefore increase clutch size, egg size or investment in feeding rates of chicks [28], [53]. Unfortunately, we do not have data on these factors for our study population. However, it seems most likely that the main effect responsible for successful fledging is food availability for chicks during the nestling period [19], [21], [26], [57], [58].
Although supplementary feeding as a management tool has been controversially discussed [32], [33], [59] a raising number of studies underline its conservation relevance for several species (e.g., Florida shrub jay, Aphelocoma coerulescens, [38]; vultures, [60], [61]; Spanish imperial eagle, Aquila adalberti, [62], brown teal, Anas chloris, [63]; Iberian lynx, Lynx pardinus, [64]). Our study demonstrates that supplementary feeding enhances reproductive success, measured as number of fledglings, of white storks breeding in close proximity to the feeding site.
In West European White Storks overall reproductive success is still low [17], [21], [22], [40] and comparable to the low reproductive success during the dramatic population decline. Hence carrying capacity of habitats in many regions may not be sufficient to sustain a viable population. This could be also the case for our study population given the low reproductive success of 1.7 fledglings at the periphery compared to 2.5 fledglings in close distance to the feeding site (<300 m).
The increased fledgling population in close distance to the feeding site may have contributed to a positive population effect, at least in our study population. Storks fledged at Affenberg Salem recolonized several regions of Baden-Württemberg (south-western Germany, Fig 4) and ringing data indicate that about 34% of the 2012 breeding population in Salem fledged at the Affenberg colony (less than 300 m from the feeding site; Hilgartner unpublished data). However, to fully understand to which extent additional feeding, applied as a conservation tool in several parts of Baden-Württemberg and other regions of Western Europe has contributed to the population recovery of West European white storks, it is necessary to study the origin of the actual breeding population and the origin of the recruits globally. Furthermore, effects of additional feeding have to be evaluated in relation to other conservation measures, such as reintroduction programs or habitat restoration. Only if we assess the impact of various conservation measurements and their interactions we will be able to make sound conservation decisions. This is of special significance in a species that still has to cope with dramatic changes in its breeding and wintering habitats and that still suffers from low overall reproductive success.
Figure 4. Location of active white stork nests in south-western Germany in 2007 (dots and stars).

Nest locations with at least one breeding partner hatched in the Affenberg colony are indicated with stars. (Walther Feld; Storchenbeauftragter Baden-Württemberg unpublished data).
Supporting Information
(XLSX)
Acknowledgments
This study would have been impossible without data collection and management over decades by Walter Angst. We are very grateful to him. We thank all other persons involved in data collection. In particular we thank Sylvia Altheimer, Jens Bizenberger, Sebastian Häusler, Jan Leibeling, Elias Schardt, Marcel Uhlemannn. We thank Walther Feld for his support and help and for providing data on the status of the state wide stork population and Christian Schloegl for his valuable comments on an earlier draft of the paper. Last but not least we thank Gilbert de Turckheim, who initiated the establishment of a stork colony at Affenberg Salem, and Ellen Merz for their support in many ways.
Climate data
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
The authors have no support or funding to report.
References
- 1.Bairlein F (1991) Population studies of white storks (Ciconia ciconia) in Europe. In: Hirons GJM, editor. Bird Population Studies. Relevance to Conservation and Management. Oxford: Oxford University Press. pp.207–229. [Google Scholar]
- 2. Zink G (1967) Populationsdynamik des Weissen Storchs, Ciconia ciconia, in Mitteleuropa. Proc Int Orn Cong 14: 191–215. [Google Scholar]
- 3. Schütz E, Szijj J (1975) Bestandsveränderungen beim Weiβstorch, fünfte Übersicht: 1959-1972. Vogelwarte 28: 61–93. [Google Scholar]
- 4. Schierer A (1967) La cigogne blanche (Ciconia ciconia) en Alsace de 1948 a 1966. Lien Ornithol d'Alsace 12: 5–55. [Google Scholar]
- 5.Jonkers DA (1989) White storks in the Netherlands: a review. In: Rheinwald G, Ogden J, Schulz H, editors. Weiβstorch - White Stork. Walsrode, Germany: ICBP. pp.231–262.
- 6.Skov H (1999) White Stork (Ciconia ciconia) in Denmark. In: Schulz H, editor. Weiβstorch im Aufwind? - White Storks on the up? Proceedings of the International Symposium on the White Stork, Hamburg 1996. Bonn, Germany: Naturschutzbund Deutschland. pp.111–131.
- 7. Fiedler G, Wissner A (1980) Freileitungen als tödliche Gefahr für Störche Ciconia ciconia . Ökol Vögel, special issue 2: 59–109. [Google Scholar]
- 8.Schulz H (1988) Weiβstorchzug. Ökologie, Gefährdung und Schutz. Weikersheim, Germany: Margraf. 459 p. [Google Scholar]
- 9.Schulz H (1999) White stork on the up? Proceedings of the International Symposium on the White Stork, Hamburg, 1996. Bonn, Germany: Naturschutzbund Deutschland. 622 p. [Google Scholar]
- 10.Dallinga JH, Schoenmakers S (1989) Population changes of the white stork since the 1850s in relation to food resources. In: Rheinwald G, Ogden J, Schulz H, editors. Weiβstorch - White Stork. Walsrode, Germany: ICBP. pp.231–262.
- 11. Barbraud C, Barbraud JC, Barbraud M (1999) Population dynamics of the white stork Ciconia ciconia in western France. Ibis 141: 469–479. [Google Scholar]
- 12. Bloesch M (1980) Drei Jahrzehnte Schweizer Storchenansiedlungsversuch Ciconia ciconia in Altreu, 1948–1979. Ornithol Beob 77: 167–194. [Google Scholar]
- 13.Thomsen K, Hötker H (2006) The sixth International White Stork Census: 2004–2005. In: Boere GC, Galbraith CA, Stroud DA, editors. Waterbirds Around the World. Edinburgh, UK: The Stationery Office. pp.493–495.
- 14.Berthold P (2007) Vogelzug. Darmstadt, Germany: Wissenschaftliche Buchgesellschaft. 280 p. [Google Scholar]
- 15. Tortosa FS, Caballero JM, Reyes-Lopez J (2002) Effect of rubbish dumps on breeding success in the white stork in Southern Spain. Waterbirds 25: 39–43. [Google Scholar]
- 16. Kruszyk R, Ciach M (2010) White storks Ciconia ciconia forage on rubbish dumps in Poland – a novel behaviour in population. Eur J Wildl Res 56: 83–87. [Google Scholar]
- 17. Schaub M, Pradel R, Lebreton JD (2004) Is the reintroduced white stork (Ciconia ciconia) population in Switzerland self-sustainable? Biol Conserv 119: 105–114. [Google Scholar]
- 18. Doligez B, Thomson DL, van Noordwijk AJ (2004) Using large-scale data analysis to assess life history and behavioural traits: the case of the reintroduced white stork Ciconia ciconia population in the Netherlands. Anim Biodivers Conserv 27: 387–402. [Google Scholar]
- 19. Massemin-Challet S, Gendner JP, Samtmann S, Pichegru L, Wulgué A, et al. (2006) The effect of migration strategy and food availability on white stork Ciconia ciconia breeding success. Ibis 148: 503–508. [Google Scholar]
- 20.Lakeberg H (1993) Zur Nahrungsökologie des Weiβstorchs Ciconia ciconia in Oberschwaben: Raum-Zeit-Nutzungsmuster und Territorialverhalten. PhD Thesis. University of Tübingen, Tübingen, Germany.
- 21. Reinhard U (2007) Bestandsdynamik des Weiβstorchs (Ciconia ciconia) in Oberschwaben (Süddeutschland) - Eine kritische Bilanz der Auswilderung. Vogelwarte 45: 81–102. [Google Scholar]
- 22. Jovani R, Tella JL (2004) Age-related environmental sensitivity and weather mediated nestling mortality in white storks Ciconia ciconia . Ecography 27: 611–618. [Google Scholar]
- 23. Vergara P, Aguirre JI, Frenández-Cruz M (2007) Arrival date, age and breeding success in white stork Ciconia ciconia . Avian Biol 38: 573–579. [Google Scholar]
- 24. Nevoux M, Barbraud JC, Barbraud C (2008) Breeding experience and demographic response to environmental variability in the white stork. Condor 110: 55–62. [Google Scholar]
- 25. Lenz E, Zimmermann M (1990) Die Jugendsterblichkeit beim Weiβstorch. Die gefährlichen ersten 12 Lebenswochen: eine empirische Studie. Der bayrischen Akad Naturschutz und Landschaftspflege 4: 141–148. [Google Scholar]
- 26. Denac D (2006) Resource-dependent weather effect in the reproduction of the white stork Ciconia ciconia . Ardea 94: 233–240. [Google Scholar]
- 27. Tryjanowski P, Kosicki J, Kuźniak S, Sparks T (2009) Long-term changes and breeding success in relation to nesting structures used by the white stork, Ciconia ciconia . Ann Zool Fenn 46: 34–38. [Google Scholar]
- 28. Tryjanowski P, Kuźniak S (2002) Size and productivity of the white stork Ciconia ciconia population in relation to common vole Microtus arvalis density. Ardea 90: 213–217. [Google Scholar]
- 29. Pfeifer R, Brandl R (1991) Der Einfluβ des Wiesenmahdtermins auf die Vogelwelt. Ornithol Anz 30: 159–171. [Google Scholar]
- 30. Johst K, Brandl R, Pfeifer R (2001) Foraging in a patchy and dynamic landscape: human land use and the white stork. Ecol Appl 11: 60–69. [Google Scholar]
- 31. Nowakowski JJ (2003) Habitat structure and breeding parameters of the white stork Ciconia ciconia in the Kolno Upland (NE Poland). Acta Ornithol 38: 39–46. [Google Scholar]
- 32.Berthold P, Mohr G (2006) Vögel füttern aber richtig. Stuttgart, Germany: Kosmos Verlag. 112 p. [Google Scholar]
- 33. Robb GN, McDonald RA, Chamberlain DE, Bearhop S (2008) Food for thought: supplementary feeding as a driver of ecological change in avian populations. Front Ecol Environ 6: 476–484. [Google Scholar]
- 34. Hoodless AN, Draycott RAH, Ludiman MN, Robertson PA (1999) Effects of supplementary feeding on territoriality, breeding success and survival of pheasants. J Appl Ecol 36: 147–156. [Google Scholar]
- 35. Elliott GP, Merton DV, Jansen PW (2001) Intensive management of a critically endangered species: the kakapo. Biol Conserv 99: 121–133. [Google Scholar]
- 36. Jonsson P, Hartikainen T, Koskela E, Mappes T (2002) Determinants of reproductive success in voles: space use in relation to food and litter size manipulation. Evol Ecol 16: 455–467. [Google Scholar]
- 37. Heath SR, Kershner EL, Cooper DM, Lynn S, Turner JM, et al. (2008) Rodent control and food supplementation increase productivity of endangered San Clemente loggerhead shrikes (Lanius ludovicianus mearnsi). Biol Conserv 141: 2506–2515. [Google Scholar]
- 38. Schoech SJ, Bridge ES, Boughton RK, Reynolds SJ, Atwell JW, et al. (2008) Food supplementation: A tool to increase reproductive output? A case study in the threatened Florida scrub-jay. Biol Conserv 141: 162–173. [Google Scholar]
- 39. Carrete M, Donázar JA, Margalida A (2006) Density-dependent productivity depression in Pyrenean bearded vultures: implications for conservation. Ecol Appl 16: 1674–1682. [DOI] [PubMed] [Google Scholar]
- 40. Moritzi M, Maumary L, Schmid D, Steiner I, Vallotton L, et al. (2001) Time budget, habitat use and breeding success of White Storks Ciconia ciconia under variable foraging conditions during the breeding season in Switzerland. Ardea 89: 457–470. [Google Scholar]
- 41.Burnham KP, Anderson DR (2002) Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach. New York, NY: Springer. 488 p. [Google Scholar]
- 42.Hinde J (1982) Compound Poisson regression models. In: Gilchrist R, editor. GLIM 82: Proceedings of the International Conference on Generalized Linear Models. Berlin: Springer. pp.109–121.
- 43. Elston DA, Moss R, Boulinier T, Arrowsmith C, Lambin X (2001) Analysis of aggregation, a worked example: numbers of ticks on red grouse chicks. Parasitology 122: 563–569. [DOI] [PubMed] [Google Scholar]
- 44. Nakagawa S, Schielzeth H (2013) A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol Evol 4: 133–142. [Google Scholar]
- 45. Schoenmakers S (1985) Regional decrease in number of white storks (Ciconia ciconia) in relation to food resources. Colonial Waterbirds 10: 167–177. [Google Scholar]
- 46. Meijer T, Elmhagen B, Eide NE, Angerbjo A (2013) Life history traits in a cyclic ecosystem: a field experiment. Oecologia 173: 439–447. [DOI] [PubMed] [Google Scholar]
- 47. Alonso JC, Alonso JA, Carrascal LM (1991) Habitat selection by foraging white storks, Ciconia ciconia, during the breeding season. Canadian J Zool 69: 1957–1962. [Google Scholar]
- 48. Tortosa FS, Castro F (2003) Development of thermoregulatory ability during ontogeny in the White Stork Ciconia ciconia . Ardeola 50: 39–45. [Google Scholar]
- 49.O'Connor RJ (1984) The Growth and Development of Birds. London: Wiley. 326 p. [Google Scholar]
- 50. Carrascal LM, Bautista LM, Lázarom E (1993) Geographical variation in the density of the White Stork Ciconia ciconia in Spain: Influence of habitat structure and climate. Biol Conserv 65: 83–87. [Google Scholar]
- 51.Martinez Rodriguez E, Fernandez R (1995) Calidad de los nidos de Cigüeña Blanca. In: Biber O, Enggist P, Marti C, Slathé T, editors. Proceedings of the International Symposium of the White Stork (Western Population). Schweizerische Vogelwarte Sempach, Basel. pp. 155–158.
- 52. Dziewiaty K (2002) Zur Bedeutung des Deichvor- und -hinterlandes der Elbe als Nahrungshabitat für Weiβstörche (Ciconia ciconia). Vogelwarte 41: 221–230. [Google Scholar]
- 53. Tortosa FS, Pérez L, Hillström L (2003) Effect of food abundance on laying date and clutch size in the White Stork Ciconia ciconia.Bird Study. 50: 112–115. [Google Scholar]
- 54.Hancock JA, Kushlan JA, Kahl MP (1992) Storks, Ibises and Spoonbills of the World. Academc Press, London.
- 55. Böhning Gaese K (1992) Zur Nahrungsökologie des Weiβstorchs (Ciconia ciconia) in Oberschwaben: Beobachtung an zwei Paaren. J Ornithol 133: 61–71. [Google Scholar]
- 56. Tryjanowski P, Jerzak L, Radkiewicz J (2005) Effect of water level and livestock on the productivity and numbers of breeding White Storks. Waterbirds 28: 378–382. [Google Scholar]
- 57. Royle N (2000) Overproduction in the lesser black-backed gull - can marginal chicks overcome the initial handicap of hatching asynchrony? Avian Biol 31: 335–344. [Google Scholar]
- 58. Djerdali S, Tortosa FS, Doumandji S (2008) Do white stork (Ciconia ciconia) parents exert control over food distribution when feeding is indirect? Ethol Ecol Evol 20: 361–374. [Google Scholar]
- 59. Martínez-Abraín A, Oro D (2013) Preventing the development of dogmatic approaches in conservation biology: A review. Biol Conserv 159: 539–547. [Google Scholar]
- 60. Oro D, Margalida A, Carrete M, Heredia R, Donázar JA (2008) Testing the goodness of supplementary feeding to enhance population viability in an endangered vulture. PLoS ONE 3: e4084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Cortés-Avizanda A, Carrete M, Donázar JA (2010) Managing supplementary feeding for avian scavengers: Guidelines for optimal design using ecological criteria. Biol Conserv 143: 1707–1715. [Google Scholar]
- 62. González LM, Margalida A, Sánchez R, Oria J (2006) Supplementary feeding as an effective tool for improving breeding success in the Spanish imperial eagle (Aquila adalberti). Biol Conserv 129: 477–486. [Google Scholar]
- 63. Rickett J, Dey C, Stothart J, O'Connor CM, Quinn JS, et al. (2013) The influence of supplemental feeding on survival, dispersal and competition in translocated brown teal, or pateke (Anas chlorotis). Emu 113: 62–68. [Google Scholar]
- 64. López-Bao JV, Palomares F, Rodríguez A, Delibes M (2010) Effects of food supplementation on home-range size, reproductive success, productivity and recruitment in a small population of Iberian lynx. Anim Conserv 13: 35–42. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.