

Abstract
The peptidoglycan precursor, Lipid II, produced in the model Gram-positive bacterium Bacillus subtilis differs from Lipid II found in Gram-negative bacteria such as Escherichia coli by a single amidation on the peptide side chain. How this difference affects the cross-linking activity of penicillin-binding proteins (PBPs) that assemble peptidoglycan in cells has not been investigated because B. subtilis Lipid II was not previously available. Here we report the synthesis of B. subtilis Lipid II and its use by purified B. subtilis PBP1 and E. coli PBP1A. While enzymes from both organisms assembled B. subtilis Lipid II into glycan strands, only the B. subtilis enzyme cross-linked the strands. Furthermore, B. subtilis PBP1 catalyzed the exchange of both d-amino acids and d-amino carboxamides into nascent peptidoglycan, but the E. coli enzyme only exchanged d-amino acids. We exploited these observations to design a fluorescent d-amino carboxamide probe to label B. subtilis PG in vivo and found that this probe labels the cell wall dramatically better than existing reagents.
Bacterial cells are surrounded by a cell wall composed of layers of peptidoglycan (PG). This mesh-like macromolecule stabilizes cell membranes against high internal osmotic pressures and is essential for survival. Peptidoglycan is assembled from the lipid-linked disaccharide pentapeptide precursor Lipid II (Figure 1a, 1).1 Peptidoglycan glycosyltransferases polymerize Lipid II into glycan strands and enzymes known as penicillin-binding proteins (PBPs) cross-link and process the peptide side chains to produce mature PG.2 Because PG is highly conserved and essential for cell survival, it is a target for antibiotics. Indeed, the most successful class of antibiotics in history, the β-lactams, inhibits the transpeptidases (TPs) that cross-link the carbohydrate strands of PG.3
Figure 1.
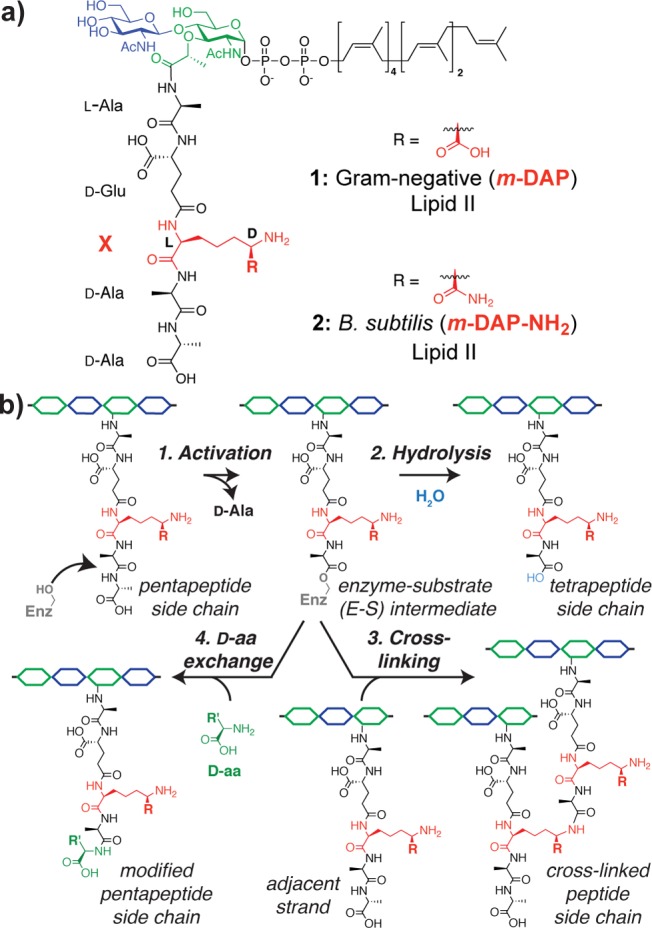
Reactions catalyzed by transpeptidases (TPs) that cross-link peptidoglycan. (a) Structures of Lipid II used by E. coli and B. subtilis.4 (b) Reactions catalyzed by TPs proceed via an enzyme–substrate intermediate that can be resolved by water to generate a tetrapeptide side chain, by a side chain on another glycan strand to yield a cross-link, or by a d-amino acid to give a pentapeptide with a new terminal amino acid.
TPs contain a conserved active-site serine that attacks the terminal d-Ala-d-Ala amide bond of the pentapeptide attached to the glycan polymer, forming a covalent enzyme–substrate (E-S) intermediate and releasing d-Ala (Figure 1b, activation step). An amino group on the peptide side chain of another glycan strand can then attack the E-S intermediate to form a cross-link and regenerate the enzyme (Figure 1b, cross-linking step). While the structure of Lipid II is highly conserved, the identity of the amino acid containing the reactive amino group varies by species.1a,5 As the Lipid II substrate is difficult to obtain,6,7 it is not well understood how these differences in the stem peptide affect TP cross-linking chemistry.8,9
The canonical Lipid II used by Gram-negative bacteria, including Escherichia coli, contains the pentapeptide l-Ala-γ-d-Glu-m-DAP-d-Ala-d-Ala (1).10 The third residue, m-DAP, which functions as the nucleophile in the cross-linking reaction, has a d-stereocenter in the side chain. Bacillus subtilis Lipid II is similar but contains a carboxamide instead of a carboxylic acid on the m-DAP side chain (2).11,12 Cell wall precursor 1 has been synthesized previously.8d We synthesized the B. subtilis Lipid II substrate 2 in 32 steps by adapting the synthesis of Gram-negative Lipid II8d,13 to introduce the carboxamide-containing m-DAP residue.14
Access to both 1 and 2 has allowed us to investigate the substrate preferences of TPs from E. coli and B. subtilis. E. coli PBP1A and B. subtilis PBP115 were purified as described in the Supporting Information, and each enzyme was then incubated with substrate 1 or substrate 2. These enzymes contain an N-terminal glycosyltransferase domain that polymerizes disaccharide subunits and a C-terminal TP domain that cross-links polymeric glycan strands. To assess cross-linking, we treated reaction mixtures with the glycosylhydrolase mutanolysin followed by NaBH4 and then used LC/MS to identify the products (Figure 2a).8d,16 As expected, each enzyme is able to polymerize and cross-link its native substrate (Figure 2b, traces i and ii). In addition, B. subtilis PBP1 is able to cross-link the canonical Gram-negative Lipid II (1, Figure 2b, trace iii), but E. coli PBP1A is not able to cross-link B. subtilis Lipid II (2, Figure 2b, trace iv), implying that the E. coli PBP discriminates against the carboxamide on the m-DAP.
Figure 2.
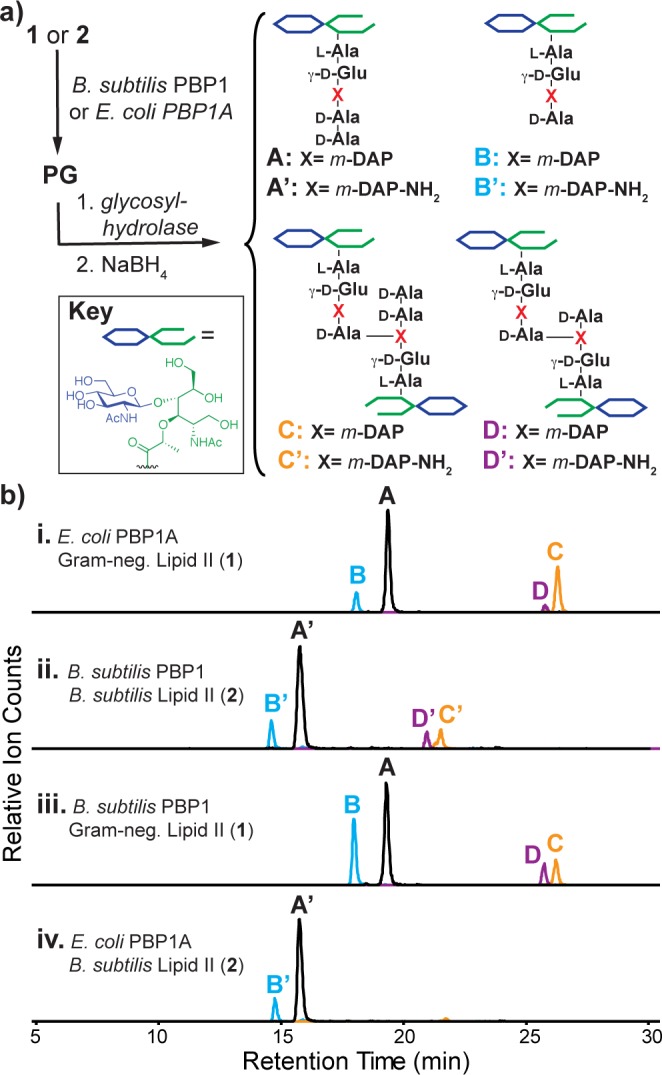
B. subtilis transpeptidases (TPs) can cross-link amidated peptidoglycan precursors in vitro. (a) Protocol for detecting cross-linking by E. coli and B. subtilis TPs in vitro. Fragments A/A′ are derived from unmodified polymer; the remaining products are derived from polymer that has been acted on by a TP. (b) LC/MS-extracted ion chromatograms showing unprocessed (A/A′), hydrolyzed (B/B′), and cross-linked products (C/C′, D/D′) formed in the presence of E. coli PBP1A (i,iv) or B. subtilis PBP1 (ii,iii) and substrate 1 (i,iii) or 2 (ii,iv).
We have previously shown that E. coli PBP1A can exchange the terminal d-Ala in cell wall precursors with other d-amino acids during in vitro PG synthesis.13a,16,17 To further assess the substrate scope of the E. coli and B. subtilis enzymes, we examined their ability to incorporate either d-Phe or d-Phe carboxamide (d-Phe-NH2) into synthetic PG (Figure 3a). E. coli PBP1A incorporated d-Phe but not d-Phe-NH2 (Figure 3b, traces i and ii), whereas B. subtilis PBP1 incorporated both (Figure 3b, traces iii and iv).18 Hence, the cross-linking and d-amino acid incorporation experiments are consistent in showing that the E. coli PBP1A TP domain discriminates against carboxamide substrates but the B. subtilis PBP1 TP domain does not.
Figure 3.
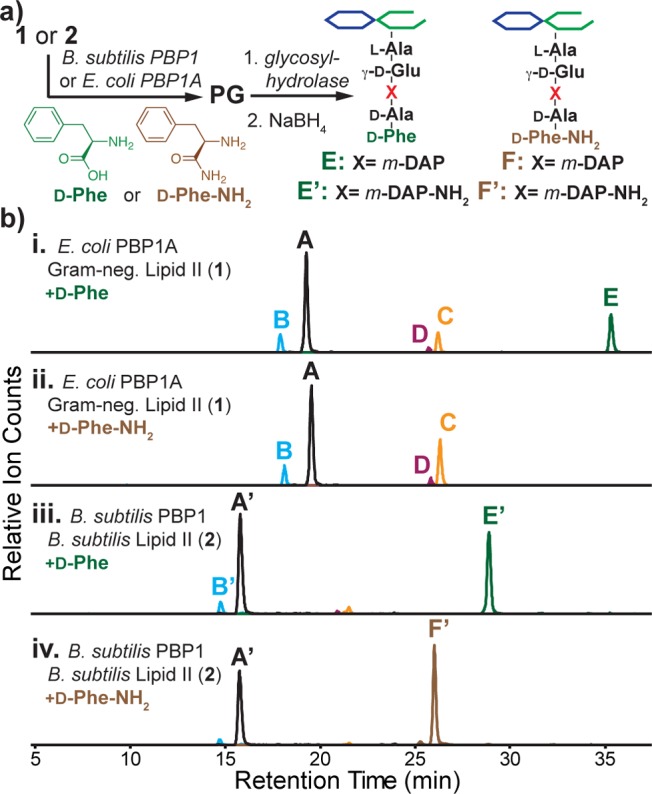
B. subtilis TPs can incorporate d-amino carboxamides into PG in vitro. (a) Protocol for detecting TP-mediated d-amino acid or carboxamide exchange in vitro. (b) LC/MS-extracted ion chromatograms of products generated by reaction of E. coli PBP1A (i,ii) or B. subtilis PBP1 (iii,iv) with Lipid II and 1 mM d-Phe (i,iii) or d-Phe-NH2 (ii,iv). Products E/E′ contain d-Phe; product F′ contains d-Phe-NH2.
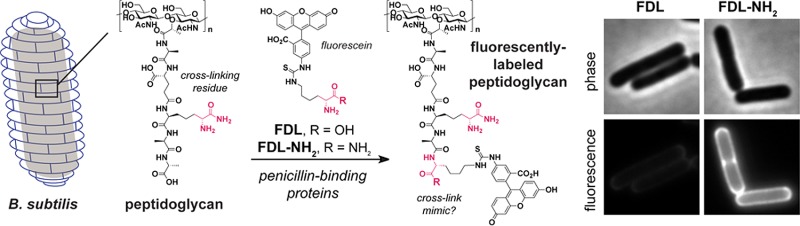
A variety of d-amino acid probes have recently been developed to fluorescently label PG in living cells,19 but labeling is poor in B. subtilis unless the cell wall hydrolase DacA, which removes terminal d-amino acids from PG, is absent.19a To determine if d-amino carboxamides can be stably incorporated into PG during B. subtilis growth, we grew cells to early log phase in medium supplemented with 500 μM d-Phe or d-Phe-NH2 and then analyzed the composition of the pentapeptide in PG fragments following enzymatic degradation. LC/MS analysis showed that d-Phe and d-Phe-NH2 were incorporated exclusively at the fifth position of the stem peptide, as observed previously for d-amino acids,20 but the levels of d-Phe-NH2 were much higher (Figure S1). These results led us to examine the utility of fluorescent d-amino carboxamide probes for imaging cell wall biosynthesis in B. subtilis.
We prepared previously reported fluorescent probe 3 (FDL)19a as well as new probe 4 (FDL-NH2) by appending fluorescein to d-Lys and d-Lys carboxamide, respectively (Figure 4a). Wild-type B. subtilis (dacA+) was grown in medium supplemented with 100 μM of either 3 or 4 for approximately four generations, and cells were then examined by fluorescence microscopy. FDL-NH24 efficiently labeled both new septa and the cylindrical walls of the cells. By contrast, FDL 3 inefficiently labeled the cells (Figure 4b). Even FDL-NH2 concentrations as low as 10 μM yielded strong labeling (Figure S2). Use of the fluorescent carboxamide probe FDL-NH2 may be preferable for imaging cell wall synthesis, as it does not require mutations (e.g., dacA deletion) that perturb peptidoglycan processing.
Figure 4.

Fluorescent d-amino carboxamide probes dramatically improve detection of PG synthesis. (a) Structures of fluorescent probes. (b) B. subtilis was grown with 100 μM probe to mid log phase. Images were adjusted to the same intensity scale to allow comparison. Scale bars: 1 μm.
To test whether d-amino carboxamides label PG in E. coli, we grew cells to early log phase in medium with 500 μM of either d-Phe or d-Phe-NH2, harvested the PG, and analyzed the PG fragments using LC/MS as before. As reported previously for d-amino acids,20b,21d-Phe incorporation was detected exclusively in the fourth position of the stem peptide (Figure S3b, trace ii). Strikingly, almost no d-Phe-NH2 incorporation was observed (Figure S3b, trace iii). It has previously been suggested that another subset of TPs called the l,d-transpeptidases are responsible for fourth-position incorporation,20b and these data suggest that, like the E. coli PBPs, E. colil,d-transpeptidases discriminate against carboxamides. Likewise, E. coli grown with 500 μM of either probe 3 or 4 for four generations incorporated 3 but not 4 (Figure S4), indicating that the ability to incorporate d-amino carboxamides may be specific to organisms like B. subtilis that cross-link substrates containing an amino group alpha to a carboxamide.
The ability of enzymes from B. subtilis to incorporate both d-amino acids and d-amino carboxamides suggests that there may be two trajectories for nucleophiles to interact with the B. subtilis transpeptidase enzyme–substrate intermediate: one for d-amino acids, which may constitute the reverse of the activation step in which a d-amino acid is the leaving group, and the other for forming cross-links to incoming m-DAP-NH2 side chains as well as d-amino carboxamides that resemble these side chains. For E. coli, the incoming side chain is essentially a d-amino acid, and other modifications are not well tolerated. The dramatically increased efficiency of incorporation of d-amino carboxamides in B. subtilis cells could be due to more efficient incorporation of this nucleophile, less efficient removal by endogenous PG hydrolases once installed, or both. d-amino carboxamide probes may also be useful for studying pathogenic bacteria that contain an amidated m-DAP residue in their peptidoglycan, such as Mycobacterium tuberculosis and Corynebacterium diphtheriae.19b,22 Using this general strategy, it may be possible to design probes that mimic cross-linking residues for other bacteria.
Acknowledgments
This research was supported by the NIH (R01 GM076710 to S.W.; R01 GM066174 to D.K.; R01 GM073831 to D.Z.R; R01 GM18568 to R.L.; CETR (U19 AI109764) to D.K., S.W., and D.Z.R.; F32 GM103056 to M.D.L.) and NSF (DGE-1144152 to J.M.M.). All high-resolution LC/MS data were acquired on an Agilent 6520 Q-TOF, spectrometer supported by the Taplin Funds for Discovery Program (P.I.: S.W.).
Supporting Information Available
Experimental procedures, synthesis of substrates and compound analysis, protein purification protocols, and LC/MS analysis of PG polymers. This material is available free of charge via the Internet at http://pubs.acs.org.
Author Contributions
⊥ M.D.L. and J.M.M. contributed equally.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
Supplementary Material
References
- a Vollmer W.; Blanot D.; de Pedro M. A. FEMS Microbiol. Rev. 2008, 32, 149–167. [DOI] [PubMed] [Google Scholar]; b Lovering A. L.; Safadi S. S.; Strynadka N. C. J. Annu. Rev. Biochem. 2012, 81, 451–478. [DOI] [PubMed] [Google Scholar]; c Typas A.; Banzhaf M.; Gross C. A.; Vollmer W. Nat. Rev. Microbiol. 2012, 10, 123–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sauvage E.; Kerff F.; Terrak M.; Ayala J. A.; Charlier P. FEMS Microbiol. Rev. 2008, 32, 234–258. [DOI] [PubMed] [Google Scholar]
- a Waxman D. J.; Strominger J. L. Annu. Rev. Biochem. 1983, 52, 825–869. [DOI] [PubMed] [Google Scholar]; b Walsh C.Antibiotics: Actions, Origins, Resistance; ASM Press: Washington, DC, 2003. [Google Scholar]
- A heptaprenyl lipid chain was used rather than the most commonly found natural lipid (undecaprenyl) because the former does not aggregate as extensively and thus is a better substrate for biochemical studies (ref (7d)).
- Bouhss A.; Trunkfield A. E.; Bugg T. D. H.; Mengin-Lecreulx D. FEMS Microbiol. Rev. 2008, 32, 208–233. [DOI] [PubMed] [Google Scholar]
- a Anderson J. S.; Matsuhashi M.; Haskin M. A.; Strominger J. L. Proc. Natl. Acad. Sci. U.S.A. 1965, 53, 881–889. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Breukink E.; van Heusden H. E.; Vollmerhaus P. J.; Swiezewska E.; Brunner L.; Walker S.; Heck A. J.; de Kruijff B. J. Biol. Chem. 2003, 278, 19898–19903. [DOI] [PubMed] [Google Scholar]; c El Ghachi M.; Bouhss A.; Barreteau H.; Touzé T.; Auger G.; Blanot D.; Mengin-Lecreulx D. J. Biol. Chem. 2006, 281, 22761–22772. [DOI] [PubMed] [Google Scholar]; d Lloyd A. J.; Gilbey A. M.; Blewett A. M.; De Pascale G.; El Zoeiby A.; Levesque R. C.; Catherwood A. C.; Tomasz A.; Bugg T. D. H.; Roper D. I.; Dowson C. G. J. Biol. Chem. 2008, 283, 6402–6417. [DOI] [PubMed] [Google Scholar]; e Patin D.; Barreteau H.; Auger G.; Magnet S.; Crouvoisier M.; Bouhss A.; Touzé T.; Arthur M.; Mengin-Lecreulx D.; Blanot D. Biochimie 2012, 94, 985–990. [DOI] [PubMed] [Google Scholar]
- a Men H.; Park P.; Ge M.; Walker S. J. Am. Chem. Soc. 1998, 120, 2484–2485. [Google Scholar]; b Ha S.; Chang E.; Lo M.-C.; Men H.; Park P.; Ge M.; Walker S. J. Am. Chem. Soc. 1999, 121, 8415–8426. [Google Scholar]; c Schwartz B.; Markwalder J. A.; Wang Y. J. Am. Chem. Soc. 2001, 123, 11638–11643. [DOI] [PubMed] [Google Scholar]; d Ye X.-Y.; Lo M.-C.; Brunner L.; Walker D.; Kahne D.; Walker S. J. Am. Chem. Soc. 2001, 123, 3155–3156. [DOI] [PubMed] [Google Scholar]; e Vannieuwenhze M. S.; Mauldin S. C.; Zia-Ebrahimi M.; Winger B. E.; Hornback W. J.; Saha S. L.; Aikins J. A.; Blaszczak L. C. J. Am. Chem. Soc. 2002, 124, 3656–3660. [DOI] [PubMed] [Google Scholar]; f Shih H.-W.; Chang Y.-F.; Li W.-J.; Meng F.-C.; Huang C.-Y.; Ma C.; Cheng T.-J. R.; Wong C.-H.; Cheng W.-C. Angew. Chem., Int. Ed. 2012, 51, 10123–10126. [DOI] [PubMed] [Google Scholar]
- Reconstitution of cross-linking has only been reported with enzymes from E. coli and S. pneumoniae, see:; a Bertsche U.; Breukink E.; Kast T.; Vollmer W. J. Biol. Chem. 2005, 280, 38096–38101. [DOI] [PubMed] [Google Scholar]; b Born P.; Breukink E.; Vollmer W. J. Biol. Chem. 2006, 281, 26985–26993. [DOI] [PubMed] [Google Scholar]; c Banzhaf M.; van den Berg van Saparoea B.; Terrak M.; Fraipont C.; Egan A.; Philippe J.; Zapun A.; Breukink E.; Nguyen-Distèche M.; den Blaauwen T.; Vollmer W. Mol. Microbiol. 2012, 85, 179–194. [DOI] [PubMed] [Google Scholar]; d Lebar M. D.; Lupoli T. J.; Tsukamoto H.; May J. M.; Walker S.; Kahne D. J. Am. Chem. Soc. 2013, 135, 4632–4635. [DOI] [PMC free article] [PubMed] [Google Scholar]; e Zapun A.; Philippe J.; Abrahams K. A.; Signor L.; Roper D. I.; Breukink E.; Vernet T. ACS Chem. Biol. 2013, 8, 2688–2696. [DOI] [PubMed] [Google Scholar]; f For computational modeling of cross-linking, see:Shi Q.; Meroueh S. O.; Fisher J. F.; Mobashery S. J. Am. Chem. Soc. 2011, 133, 5274–5283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- a While it has been difficult to characterize the transpeptidases, the substrate specificities of the low-molecular-mass (carboxypeptidase and endopeptidase) PBP classes have been studied. For a review, see:Pratt R. F. Cell. Mol. Life Sci. 2008, 65, 2138–2155. [DOI] [PMC free article] [PubMed] [Google Scholar]; b For a discussion of how amidated substrates are processed by a B. subtilis low-molecular-mass PBP, see:Nemmara V. V.; Adediran S. A.; Dave K.; Duez C.; Pratt R. F. Biochemistry 2013, 52, 2627–2637. [DOI] [PubMed] [Google Scholar]
- a Primosigh J.; Pelzer H.; Maass D.; Weidel W. Biochim. Biophys. Acta 1961, 46, 68–80. [DOI] [PubMed] [Google Scholar]; b Glauner B.; Höltje J. V.; Schwarz U. J. Biol. Chem. 1988, 263, 10088–10095. [PubMed] [Google Scholar]
- a Warth A. D.; Strominger J. L. Biochemistry 1971, 10, 4349–4358. [DOI] [PubMed] [Google Scholar]; b Atrih A.; Bacher G.; Allmaier G.; Williamson M. P.; Foster S. J. J. Bacteriol. 1999, 181, 3956–3966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- In B. subtilis spore PG, m-DAP is not amidated. See:Warth A. D.; Strominger J. L. Proc. Natl. Acad. Sci. U.S.A. 1969, 64, 528–535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- a Lupoli T. J.; Tsukamoto H.; Doud E. H.; Wang T.-S. A.; Walker S.; Kahne D. J. Am. Chem. Soc. 2011, 133, 10748–10751. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Tsukamoto H.; Kahne D. Bioorg. Med. Chem. Lett. 2011, 21, 5050–5053. [DOI] [PMC free article] [PubMed] [Google Scholar]; c Hesek D.; Lee M.; Zajíček J.; Fisher J. F.; Mobashery S. J. Am. Chem. Soc. 2012, 134, 13881–13888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Experimental procedures and characterization described in the SI.
- a Jackson G. E.; Strominger J. L. J. Biol. Chem. 1984, 259, 1483–1490. [PubMed] [Google Scholar]; b Popham D. L.; Setlow P. J. Bacteriol. 1995, 177, 326–335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lupoli T. J.; Lebar M. D.; Markovski M.; Bernhardt T.; Kahne D.; Walker S. J. Am. Chem. Soc. 2014, 136, 52–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PBPs also have been shown to transfer d-amino acids onto peptide substrates, see:; a Pollock J. J.; Ghuysen J. M.; Linder R.; Salton M. R.; Perkins H. R.; Nieto M.; Leyh-Bouille M.; Frère J. M.; Johnson K. Proc. Natl. Acad. Sci. U.S.A. 1972, 69, 662–666. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Adam M.; Damblon C.; Jamin M.; Zorzi W.; Dusart V.; Galleni M.; el Kharroubi A.; Piras G.; Spratt B. G.; Keck W. Biochem. J. 1991, 279, 601–604. [DOI] [PMC free article] [PubMed] [Google Scholar]; c Kumar I.; Pratt R. F. Biochemistry 2005, 44, 9961–9970. [DOI] [PubMed] [Google Scholar]
- This result provides direct evidence that PBPs are responsible for d-amino acid incorporation into B. subtilis PG in vivo (see ref (20b)).
- a Kuru E.; Hughes H. V.; Brown P. J.; Hall E.; Tekkam S.; Cava F.; de Pedro M. A.; Brun Y. V.; Vannieuwenhze M. S. Angew. Chem., Int. Ed. 2012, 51, 12519–12523. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Siegrist M. S.; Whiteside S.; Jewett J. C.; Aditham A.; Cava F.; Bertozzi C. R. ACS Chem. Biol. 2013, 8, 500–505. [DOI] [PMC free article] [PubMed] [Google Scholar]; c Liechti G. W.; Kuru E.; Hall E.; Kalinda A.; Brun Y. V.; VanNieuwenhze M.; Maurelli A. T. Nature 2014, 506, 507–510. [DOI] [PMC free article] [PubMed] [Google Scholar]; d Shieh P.; Siegrist M. S.; Cullen A. J.; Bertozzi C. R. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 5456–5461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- a Lam H.; Oh D.-C.; Cava F.; Takacs C. N.; Clardy J.; de Pedro M. A.; Waldor M. K. Science 2009, 325, 1552–1555. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Cava F.; de Pedro M. A.; Lam H.; Davis B. M.; Waldor M. K. EMBO J. 2011, 30, 3442–3453. [DOI] [PMC free article] [PubMed] [Google Scholar]; c Cava F.; Lam H.; de Pedro M. A.; Waldor M. K. Cell. Mol. Life Sci. 2011, 68, 817–831. [DOI] [PMC free article] [PubMed] [Google Scholar]; d Leiman S. A.; May J. M.; Lebar M. D.; Kahne D.; Kolter R.; Losick R. J. Bacteriol. 2013, 195, 5391–5395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- a Tsuruoka T.; Tamura A.; Miyata A.; Takei T.; Iwamatsu K.; Inouye S.; Matsuhashi M. J. Bacteriol. 1984, 160, 889–894. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Caparros M.; Pisabarro A. G.; de Pedro M. A. J. Bacteriol. 1992, 174, 5549–5559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- a Schleifer K. H.; Kandler O. Bacteriol. Rev. 1972, 36, 407–477. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Linnett P. E.; Strominger J. L. J. Biol. Chem. 1974, 249, 2489–2496. [PubMed] [Google Scholar]; c Lavollay M.; Arthur M.; Fourgeaud M.; Dubost L.; Marie A.; Veziris N.; Blanot D.; Gutmann L.; Mainardi J.-L. J. Bacteriol. 2008, 190, 4360–4366. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.