

Abstract
Phenotypic analysis of gene-specific knockout (KO) mice has revolutionized our understanding of in vivo gene functions. As the use of mouse embryonic stem (ES) cells is inevitable for conventional gene targeting, the generation of knockout mice remains a very time-consuming and expensive process. To accelerate the large-scale production and phenotype analyses of KO mice, international efforts have organized global consortia such as the International Knockout Mouse Consortium (IKMC) and International Mouse Phenotype Consortium (IMPC), and they are persistently expanding the KO mouse catalogue that is publicly available for the researches studying specific genes of interests in vivo. However, new technologies, adopting zinc-finger nucleases (ZFNs) or Transcription Activator-Like Effector (TALE) Nucleases (TALENs) to edit the mouse genome, are now emerging as valuable and effective shortcuts alternative for the conventional gene targeting using ES cells. Here, we introduce the recent achievement of IKMC, and evaluate the significance of ZFN/TALEN technology in mouse genetics. [BMB Reports 2012; 45(12): 686-692]
Keywords: Gene targeting, Knockout, TALEN, ZFN
INTRODUCTION
Based on both genetic and physiological similarities to humans, the mouse has a long history of being used as an invaluable model organism in biological and clinical studies (1,2). Furthermore, various genetic tools modifying the mouse genome have enabled the researchers to study gene functions in vivo, and manipulation of the mouse genome through gene targeting in mouse embryonic stem (ES) cells has been central to mouse genetics over the past two decades. It is now generally accepted that the phenotype analysis of genetically-engineered mouse (GEM) models has accelerated our understanding of in vivo gene functions. Using GEM models, a plethora of human diseases (e.g., glioblastoma multiforme) has been reproduced and studied in mice (3). In appreciation of the valuable contribution, the Nobel Prize in Physiology or Medicine 2007 was awarded jointly to Mario R. Capecchi, Sir Martin J. Evans and Oliver Smithies for their discoveries of principles of gene targeting in mice using ES cells.
After completing the human genome project, GEM models have been recognized as essential materials for functional genomics, and the demands for developing novel GEM models have significantly increased. However, gene targeting using mouse ES cells is still a time-consuming, expensive, and laborious process. To initiate breakthroughs, global consortia have been organized for the high throughput production and phenotype analysis of GEM models. Furthermore, new technologies employing engineered nucleases are emerging and replacing the use of ES cells for genome editing in mice. Here, we introduce the recent achievement of the International Knockout Mouse Consortium (IKMC) and International Mouse Phenotype Consortium (IMPC), and evaluate the significance of engineered nucleases (e.g., ZFN and TALEN technologies) in mouse genetics.
INTERNATIONAL EFFORTS FOR THE LARGE-SCALE PRODUCTION AND PHENOTYPE ANALYSES OF KNOCKOUT MICE
The IKMC is an international consortium composed of Knockout Mouse Project (KOMP, USA), European Conditional Mouse Mutagenesis Program (EUCOMM, Europe), North American Conditional Mouse Mutagenesis Project (NorCOMM, Canada), and Texas A&M Institute for Genomic Medicine (TIGM, USA). KOMP includes CSD, a collaborative team at the Children's Hospital Oakland Research Institute (CHORI), the Wellcome Trust Sanger Institute (WTSI, UK), the University of California at Davis School of Veterinary Medicine, and Regeneron Pharmaceuticals, Inc. (USA).
The aim of IKMC is to mutate all protein-coding genes in the mouse by gene trapping and targeting using mouse embryonic stem (ES) cells. Historically, ES cell lines from 129 mouse strains are prevalently used to target genes in mice. However, as C57BL/6 (B6) mice are the most intensively studied mouse strain, there is plenty of data describing normative physiologies of B6 mice. Therefore, a laborious and time-consuming backcross had to be conducted to bring the targeted mutation into a pure B6 genetic background after KO mice were generated using ES cells derived from 129 strain. To this end, IKMC members established the utilization of ES cell lines with a pure B6 genetic background (4). Since the Agouti allele of B6 mice is recessive, tyrosinase-deficient B6 (thus albino) or Balb/c female mice are used to collect host embryos for chimera generation and to test germ-line transmission for coat color screening. Furthermore, they recently simplified this complex breeding scheme by restoring the dominant Agouti allele in B6 ES cells (5). This experimental setup provides us with gene targeted mice of a pure B6 genetic background that can be immediately utilized. The comprehensive phenotypes of GEM models are also being analyzed through the systematic platforms for mouse phenotyping under the International Mouse Phenotype Consortium (IMPC) (6). Furthermore, IKMC allows the public to freely access KO mouse resources and phenotyping data through the internet. Therefore, by generating and phenotyping GEM models, these international efforts will make it possible to annotate every single gene function in mice in the near future.
The gene targeting strategies with a conditional potential
The gene targeting strategies by the IKMC can be categorized into three types. First, the TIGM has been employing the gene-trap strategy. They use retroviral gene trap vectors containing 5' selectable marker β-geo, a functional fusion between the β-galactosidase and neomycin resistance genes, for identification of successful gene trap events. Later, it can be also used for the reporter gene in vivo.
VelociGene, another KOMP member generates definitive null alleles (7). In most cases, they delete all protein-coding genomic loci (i.e., both exons and introns ranging from start codon to stop codon). Since they use modified bacterial artificial chromosome (BAC) clones as a targeting construct, this strategy can be applied to all genes transcribed by RNA polymerase II, and does not need to be restricted by the size of a gene, intron-exon structure, RNA splicing pattern, or protein-coding capacity (7).
The other IKMC members are archiving mutant ES cells and mice containing a versatile allele called the knockout first allele (Fig. 1). The initial allele (tm1a) has a gene trapping cassette in an intron, and the critical exon(s) is flanked with LoxP sequences. After establishing a mutant mouse line, the gene trapping capacity can be immediately utilized. In addition, a lacZ gene contained in the promoter-less trapping cassette can be used as a reporter to monitor the gene expression in vivo. This gene trap cassette is flanked with Frt sequences, and thus can be removed by crossing mutant mice with transgenic mice expressing Flp recombinase in germ cells (tm1c) such as ACT:FLPe transgenic mice (8). Removal of the gene trap element restores normal gene activity. After eliminating the gene trap cassette, the floxed exon of the tm1c allele can be deleted using Cre recombinase (tm1d), which induces a frameshift mutation and subsequent nonsense mediated decay (NMD) of the mutant transcript (9). If both the trapping cassette and floxed exon of the tm1a allele are deleted, this mouse line can only be used as a lacZ-tagged reporter mouse line (tm1b).
Fig. 1. The basic IKMC knockout allele (tm1a). The lacZ cassette has a 5'-splicing acceptor. Flp removes both lacZ and neo cassettes (tm1c), and Cre deletes both the neo cassette and exon 2 (tm1b). Further action of Cre on tm1c allele removes the critical exon 2 (tm1d). Adopted from Skarnes et al. (9).
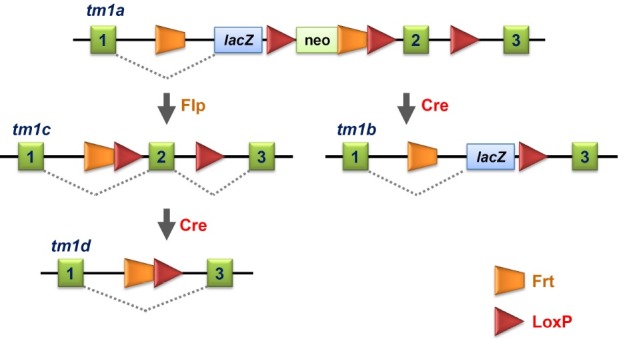
As of October, 2012, ES cell clones with conditional alleles for 8,897 genes, deletion alleles for 3,622 genes, and gene trap alleles for 9,583 genes are generated by the IKMC, and a small number of mutant mice are also produced using these ES cell clones (http://www.knockoutmouse.org/about/geneprogresssummary). The targeted ES cell lines and mutant mice can be easily searched at the IKMC web site (http://www.knockoutmouse.org/).
The practical use of Cre recombinase-expressing mouse lines
The conditional knockout mouse strategy is now prevailing as it has overwhelming advantages over classical knockout and gene trap strategies. For example, if a conditional allele is introduced to the mouse genome, germ-line mutation can be also generated using Protamin-Cre transgenic mice (10). In addition, germ-line mutations of essential genes frequently result in embryonic lethality, preventing the studies of gene functions in post-natal stages including adult mice. Thus spatio-temporal control of Cre recombinase can be employed to bypass the embryonic lethality. Furthermore, generating mouse models for human diseases that frequently occur sporadically in specific tissues is a more reasonable approach. Therefore, the generation of reliable Cre transgenic mouse lines may be central for the analysis of in vivo gene functions using conditional knockout mice. Diverse Cre transgenic mouse lines that were previously developed can be searched in the web site of Mouse Genome Informatics (MGI, http://www.informatics.jax.org/recombinase.shtml).
Cre transgenic mice have been being evolved further by adopting state-of-the-art technologies for the conditional modulation of Cre activity. ERT2, a modified ligand-binding domain of the estrogen receptor, has been successfully used to conditionally translocate a fusion protein into the nucleus upon the binding of 4-hydroxytamoxifen (4-OHT) (11). When 4-OHT binds to Cre-ERT2 fusion protein, this liganded complex translocates into the nucleus and deletes the floxed region. Combined with tissue-specific promoters, Cre-ERT2 transgenic mice are widely used to regulate gene knockout in a spatio-temporal manner.
Although Cre lines are the necessary reagents for generating their conditional mutations, their use is still limiting. Currently, a single satisfactory database that well integrates and updates all available Cre lines does not exist. Furthermore, similar Cre lines are often generated independently by disparate groups, but their reports do not always describe their detailed and critical characteristics including the efficiency of recombination, cell and tissue specificity, and genetic background effects. To improve the current insufficiency of and accessibility to appropriate Cre mouse lines, the CREATE (Coordination of resources for conditional expression of mutated mouse alleles) project (http://www.creline.org/) and EUCOMMTOOLS are planning to maximize the utility of the conditional IKMC resources by integrating and generating conditional Cre mouse lines (12).
The advanced version of Cre mouse lines
These days, the caveats of the previously generated Cre transgenic mouse lines including Cre expressions in off-target tissues, effect of the parent-of-origin, and inconsistent Cre recombination among littermates are being identified and addressed (13). For example, the expressions of Cre transgenes do not exactly coincide with those of the cognate genes. Emx1-Cre transgenic mice were generated using the 11-kb sequence located upstream of the Emx1 gene, but the Cre-mediated recombination did not match the endogenous expression pattern of Emx1 gene (14). That is caused by the use of a partial, but not the entire region of the promoter, thus discarding potentially important regulatory elements (e.g., enhancers) required for the intended Cre expression. Furthermore, the expression of Cre transgene may be affected by the genomic environment near the transgene integration site.
The knock-in strategy will make the Cre expression pattern much more similar to the cognate gene. However, it may be also problematic. For example, conventional Cre knock-in inevitably mutates one wild-type allele, and thus haplo-insufficient phenotypes are possibly induced. However, Chapman et al. described the novel strategy that can potentially solve the drawback (15). They integrated a “self-cleaving” E2A peptide sequence-fused Cre-ERT2 coding cassette into the stop codon of the endogenous surfactant C (SPC) gene. This strategy maintains endogenous SPC, and, at the same time, the same numbers of Cre-ERT2 protein are produced. It will be very helpful if these advantages are reflected for the future generation of all Cre mouse lines.
ENGINEERED NUCLEASES
Until recently, mouse ES cells were essential for custom-designed genetic modifications of the mouse genome. Now, engineered nucleases including zinc-finger nucleases (ZFNs) or Transcription Activator-Like Effector (TALE) Nucleases (TALENs) are rapidly evolving. Injection of ZFN or TALEN mRNA transcribed in vitro into one-cell stage mouse embryos allows rapid creation of genetically-engineered mouse (GEM) models without generating gene-targeted ES cell clones. Here, we discuss the basic characteristics and applications of engineered nucleases in regard to genetic engineering in vivo.
Sequence-specific recognition and restriction by ZFNs and TALENs
ZFNs and TALENs have been produced by connecting sequence-specific zinc finger arrays or TALEs for chosen DNA sequences to the cleavage domain of Fok I endonuclease through a linker (Fig. 2A, B). The modular structure of zinc fingers and TALEs support the generation of custom-designed DNA-binding domains for chosen DNA sequences with an exceptional specificity. After the pioneering work done by Bibikova et al. (16), zinc-finger domains have been intensively studied, and are being used widely for the genome modification (17-20). Individual zinc finger motifs are capable of binding triplets of DNA sequence, and thus zinc-finger proteins with adequate specificity can be produced through the modular assembly of individual zinc finger motifs (21,22); e.g., six zinc-finger motifs recognizing a specific 18-bp sequence is sufficient to pick up a single genomic locus in the mouse genome (23).
Fig. 2. The principles of ZFN- or TALEN-mediated gene targeting. (A) Zinc finger (ZF) motifs and (B) TALE modules are designed to detect specific sequences in an opposite direction. Previously developed, heterogeneous nuclease domains of FokI endonuclease can only form a heterodimer that is essential to exert endonuclease activity (32). (C) When connected to ZFs or TALEs through a linker, each pair of ZFNs or TALENs induces a double-strand break (DSB) at the spacer region between the binding sequences. DSBs are repaired through homologous recombination (HR) or non-homologous end joining (NHEJ). (D) If a donor DNA containing an additional sequence or modifications of the endogenous sequence, HR-mediated gene conversion can be achieved. (E) Without the template for HR-mediated DNA repair, DSB will be repaired through the error-prone NHEJ pathway, frequently resulting in deletion, insertion, and substitution of the original sequence.
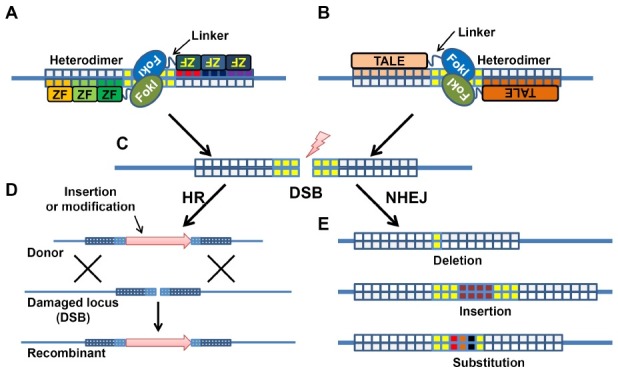
TALEs are transcription factors produced by bacterial plant pathogen, Xanthomonas, that enter into the nucleus and directly activate the expression of host genes (24). Boch et al. uncovered the cipher of TALEs responsible for their sequence-specific recognition of the target sequences (25). The DNA-binding domains of TALE are typically composed of repeats of 34 highly conserved amino-acid residues with the exception of the 12th and 13th amino acid. Each repeat recognizes one base pair in the target DNA, and these two variable amino-acid residues dictate the specificity of each repeat (e.g., HD, NG, NI, and NN for C, T, A, and G, respectively). Exploiting this modular interaction, Christian et al. created the first TALEN, and validated its activity on synthetic targets (26). Subsequent studies proved the versatile nuclease activity of TALENs against specific genomic loci in yeasts (27), and in cultured human cells (28). Further, the minimal DNA-binding domain of TALE proteins were determined, and their targeting specificity was validated (28,29).
The dimerization of FokI nuclease domains is required to cleave the target DNA (30). Normally, a pair of ZFNs or TALENs is designed to target a single locus, and the target sequence is placed between their binding sequences. Therefore, conventional ZFNs and TALENs act as heterodimers to induce double strand breaks (DSBs) at their target loci (Fig. 2C). This suggests that homodimer formation of engineered nucleases may induce DSBs at off-target sites, i.e., DNA sequences other than the intended target sequence (31). To minimize the homodimer formation, Miller et al. developed two heterodimeric FokI cleavage domain variants by engineering the interface mediating their interaction (32). This structural modification significantly decreases homodimer formation and the potential toxicity associated with off-target effects, and generally contributes to improving the specificity of ZFNs and TALENs.
Due to their relatively long history, ZFNs have been extensively studied not only to improve the targeting specificity, but also to minimize the cytotoxicity associated with off-target effects. However, there still seems to be debates on how to efficiently design ZFNs with sufficient specificities (33-35). As well as the simple cipher of TALE binding to DNA (25), TALENs are rapidly evolving through absorbing the systemic advantages achieved from studying ZFNs.
Genome editing by ZFNs and TALENs
The two subunits of a ZFN and TALEN pair bind to two adjacent sequences separated by a short spacer sequence in an inverted orientation (Fig. 2A, B) and the dimerized FokI nuclease domain introduces DSBs at the spacer region (Fig. 2C). These DSBs, inducing deleterious effects on the genomic integrity, are generally solved by two highly conserved DNA repair pathways (Fig. 2D, E): homologous recombination (HR) and non-homologous end-joining (NHEJ). These innate responses of the cells against DSBs are enabling us to edit the genomes of diverse species using ZFNs and TALENs. Especially, after the first successful application in Drosophila (16), a variety of mutant organisms, including mice, has been created using ZFNs (18).
DNA repair by NHEJ is a rapid and efficient process, but is error-prone (36). Previous studies indicate that ZFN-induced DSBs are prevalently repaired by NHEJ, and that null mutations induced by small insertions, deletions, and even substitutions frequently appear at the targeted locus (17). In contrast, HR is quite a rare event, but is essential for gene targeting in ES cells (37). However, when a DSB is introduced, the efficiency of HR can be significantly elevated in mammalian cells (38). Similarly, ZFN-induced DSBs effectively activates HR in human cells (39). Along this line, gene-targeted mice were generated by directly injecting ZFN mRNAs and the targeting construct into the pronuclear mouse embryos (40). Furthermore, instead of the plasmid-based gene-targeting vector that is laborious and time-consuming to construct, a single stranded synthetic oligonucleotide is also successfully used as a template for ZFN-induced homology-directed repair in mouse embryos of the pronuclear stage (41). These results indicate that ES cells are not an essential reagent for the targeted modification of the mouse genome any more.
Despite the short history, TALENs have rapidly developed as valuable tools for genome editing (42), and are being applied for generating genetically-engineered organisms of diverse species. In fact, TALENs were used to target genes in C. elegans, zebrafish, rats, cows, and pigs (43-47). In general, the efficiency of the genome modification by TALENs seems to approach the levels induced by ZFNs (28). In addition, much evidence points out the advantages of TALENs over ZNFs. For example, as compared to the corresponding ZFNs in human cells, even though TALENs were similar in gene disruption activities, they exhibited significantly reduced nuclease-associated cytotoxic activity, and revealed only minimal off-target activity at the highly homologous sequence (29). Consistently, compared to the efficiency obtained with ZFNs (48), TALEN showed less toxicity in generating TALEN-mediated knockout rats (47). These beneficial aspects, in conjunction with the simple decipher (25), may lead to the prevalent use of TALENs in directly generating GEM models.
Replacement of the use of ES cells
It is plausible to expect that engineered nucleases will considerably replace the use of ES cells in modifying the mouse genome. In addition to generating null alleles, engineered nucleases will be an effective tool for the knock-in strategies aiming to substitute amino acid(s) or to insert a short stretch of nucleotides. For example, to study endogenous transporter trafficking of the plasma membrane dopamine (DA) transporter (DAT), Rao et al. established a knock-in mouse line harboring a hemagglutinin (HA)-epitope tag at the second extracellular loop of mouse DAT via the homologous recombination in ES cells (49). As the use of engineered nucleases enables the sequence-specific modification of the mouse genome in embryos (41,50), it could be directly achieved by employing an efficient engineered nuclease and donor construct or oligonucleotide by avoiding the laborious and time-consuming steps of establishing targeted ES cells, generation of chimeras, and germ-line transmission. According to Cui et al., even an expression cassette can be inserted by the aid of engineered nucleases (50). It may provide a more expanded potential for generating diverse mouse models, and the use of engineered nucleases will make it possible to avoid the use of ES cells. Therefore, the mouse generation procedure should be simplified by the use of engineered nucleases.
The potential use of engineered nucleases for generating Cre mouse lines
The need for appropriate Cre mouse lines is enormously increasing, proportional to the expansion of publically available conditional KO mouse lines. As mentioned above, one of the most advanced versions of Cre mouse lines was produced by inserting a self-cleaving peptide-fused Cre-ERT2-encoding sequence into a stop codon of a gene (15). This strategy is compatible to the use of engineered nucleases in fertilized mouse eggs, and thus will be beneficial for the efficient and massive generation of Cre mouse lines.
CONCLUSIONS
The international efforts through the IKMC are expanding the mutant mouse resources, and are promoting public access to GEM models. In addition, the current development and versatility of engineered nucleases are pushing the generation of novel mouse models by circumventing the use of ES cells. Furthermore, these technologies will synergize to accelerate the establishment and use of GEM models as a more generalized tool for studying in vivo gene functions.
Acknowledgments
This study was supported by the Ministry of Education, Science, and Technology (20120006489, 20120000174, 2012009607) and by the Ministry of Health & Welfare (A085136), Republic of Korea.
References
- 1.Paigen K. One hundred years of mouse genetics: an intellectual history. II. The molecular revolution (1981-2002). Genetics. (2003);163:1227–1235. doi: 10.1093/genetics/163.4.1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Paigen K. One hundred years of mouse genetics: an intellectual history. I. The classical period (1902-1980). Genetics. (2003);163:1–7. doi: 10.1093/genetics/163.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lim S. K., Llaguno S. R., McKay R. M., Parada L. F. Glioblastoma multiforme: a perspective on recent findings in human cancer and mouse models. BMB Rep. (2011);44:158–164. doi: 10.5483/BMBRep.2011.44.3.158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.International Mouse Knockout, C. Collins F. S., Rossant J., Wurst W. A mouse for all reasons. Cell. (2007);128:9–13. doi: 10.1016/j.cell.2006.12.018. [DOI] [PubMed] [Google Scholar]
- 5.Pettitt S. J., Liang Q., Rairdan X. Y., Moran J. L., Prosser H. M., Beier D. R., Lloyd K. C., Bradley A., Skarnes W. C. Agouti C57BL/6N embryonic stem cells for mouse genetic resources. Nat. Methods. (2009);6:493–495. doi: 10.1038/nmeth.1342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kim I. Y., Shin J. H., Seong J. K. Mouse phenogenomics, toolbox for functional annotation of human genome. BMB Rep. (2010);43:79–90. doi: 10.5483/BMBRep.2010.43.2.079. [DOI] [PubMed] [Google Scholar]
- 7.Valenzuela D. M., Murphy A. J., Frendewey D., Gale N. W., Economides A. N., Auerbach W., Poueymirou W. T., Adams N. C., Rojas J., Yasenchak J., Chernomorsky R., Boucher M., Elsasser A. L., Esau L., Zheng J., Griffiths J. A., Wang X., Su H., Xue Y., Dominguez M. G., Noguera I., Torres R., Macdonald L. E., Stewart A. F., DeChiara T. M., Yancopoulos G. D. High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nat. Biotechnol. (2003);21:652–659. doi: 10.1038/nbt822. [DOI] [PubMed] [Google Scholar]
- 8.Rodriguez C. I., Buchholz F., Galloway J., Sequerra R., Kasper J., Ayala R., Stewart A. F., Dymecki S. M. High-efficiency deleter mice show that FLPe is an alternative to Cre-loxP. Nat. Genet. (2000);25:139–140. doi: 10.1038/75973. [DOI] [PubMed] [Google Scholar]
- 9.Skarnes W. C., Rosen B., West A. P., Koutsourakis M., Bushell W., Iyer V., Mujica A. O., Thomas M., Harrow J., Cox T., Jackson D., Severin J., Biggs P., Fu J., Nefedov M., de Jong P. J., Stewart A. F., Bradley A. A conditional knockout resource for the genome-wide study of mouse gene function. Nature. (2011);474:337–342. doi: 10.1038/nature10163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.O'Gorman S., Dagenais N. A., Qian M., Marchuk Y. Protamine-Cre recombinase transgenes efficiently recombine target sequences in the male germ line of mice, but not in embryonic stem cells. Proc. Natl. Acad. Sci. U.S.A. (1997);94:14602–14607. doi: 10.1073/pnas.94.26.14602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Indra A. K., Warot X., Brocard J., Bornert J. M., Xiao J. H., Chambon P., Metzger D. Temporally-controlled site-specific mutagenesis in the basal layer of the epidermis: comparison of the recombinase activity of the tamoxifen-inducible Cre-ER(T) and Cre-ER(T2) recombinases. Nucl. Acids Res. (1999);27:4324–4327. doi: 10.1093/nar/27.22.4324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Smedley D., Salimova E., Rosenthal N. Cre recombinase resources for conditional mouse mutagenesis. Methods. (2011);53:411–416. doi: 10.1016/j.ymeth.2010.12.027. [DOI] [PubMed] [Google Scholar]
- 13.Heffner C. S., Herbert Pratt C., Babiuk R. P., Sharma Y., Rockwood S. F., Donahue L. R., Eppig J. T., Murray S. A. Supporting conditional mouse mutagenesis with a comprehensive cre characterization resource. Nat. Commun. (2012);3:1218. doi: 10.1038/ncomms2186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jin X. L., Guo H., Mao C., Atkins N., Wang H., Avasthi P. P., Tu Y. T., Li Y. Emx1-specific expression of foreign genes using "knock-in" approach. Biochem. Biophys. Res. Commun. (2000);270:978–982. doi: 10.1006/bbrc.2000.2532. [DOI] [PubMed] [Google Scholar]
- 15.Chapman H. A., Li X., Alexander J. P., Brumwell A., Lorizio W., Tan K., Sonnenberg A., Wei Y., Vu T. H. Integrin alpha6beta4 identifies an adult distal lung epithelial population with regenerative potential in mice. J. Clin. Invest. (2011);121:2855–2862. doi: 10.1172/JCI57673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bibikova M., Beumer K., Trautman J. K., Carroll D. Enhancing gene targeting with designed zinc finger nucleases. Science. (2003);300:764. doi: 10.1126/science.1079512. [DOI] [PubMed] [Google Scholar]
- 17.Urnov F. D., Rebar E. J., Holmes M. C., Zhang H. S., Gregory P. D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genet. (2010);11:636–646. doi: 10.1038/nrg2842. [DOI] [PubMed] [Google Scholar]
- 18.Carroll D. Genome engineering with zinc-finger nucleases. Genetics. (2011);188:773–782. doi: 10.1534/genetics.111.131433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kim H. J., Lee H. J., Kim H., Cho S. W., Kim J. S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Res. (2009);19:1279–1288. doi: 10.1101/gr.089417.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kim S., Lee M. J., Kim H., Kang M., Kim J. S. Preassembled zinc-finger arrays for rapid construction of ZFNs. Nat. Methods. (2011);8:7. doi: 10.1038/nmeth0111-7a. [DOI] [PubMed] [Google Scholar]
- 21.Pabo C. O., Peisach E., Grant R. A. Design and selection of novel Cys2His2 zinc finger proteins. Annu. Rev. Biochem. (2001);70:313–340. doi: 10.1146/annurev.biochem.70.1.313. [DOI] [PubMed] [Google Scholar]
- 22.Bae K. H., Kwon Y. D., Shin H. C., Hwang M. S., Ryu E. H., Park K. S., Yang H. Y., Lee D. K., Lee Y., Park J., Kwon H. S., Kim H. W., Yeh B. I., Lee H. W., Sohn S. H., Yoon J., Seol W., Kim J. S. Human zinc fingers as building blocks in the construction of artificial transcription factors. Nat. Biotechnol. (2003);21:275–280. doi: 10.1038/nbt796. [DOI] [PubMed] [Google Scholar]
- 23.Kim J. S., Pabo C. O. Getting a handhold on DNA: design of poly-zinc finger proteins with femtomolar dissociation constants. Proc. Natl. Acad. Sci. U.S.A. (1998);95:2812–2817. doi: 10.1073/pnas.95.6.2812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kay S., Hahn S., Marois E., Hause G., Bonas U. A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science. (2007);318:648–651. doi: 10.1126/science.1144956. [DOI] [PubMed] [Google Scholar]
- 25.Boch J., Scholze H., Schornack S., Landgraf A., Hahn S., Kay S., Lahaye T., Nickstadt A., Bonas U. Breaking the code of DNA binding specificity of TAL-type III effectors. Science. (2009);326:1509–1512. doi: 10.1126/science.1178811. [DOI] [PubMed] [Google Scholar]
- 26.Christian M., Cermak T., Doyle E. L., Schmidt C., Zhang F., Hummel A., Bogdanove A. J., Voytas D. F. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics. (2010);186:757–761. doi: 10.1534/genetics.110.120717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li T., Huang S., Jiang W. Z., Wright D., Spalding M. H., Weeks D. P., Yang B. TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucl. Acids Res. (2011);39:359–372. doi: 10.1093/nar/gkq704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Miller J. C., Tan S., Qiao G., Barlow K. A., Wang J., Xia D. F., Meng X., Paschon D. E., Leung E., Hinkley S. J., Dulay G. P., Hua K. L., Ankoudinova I., Cost G. J., Urnov F. D., Zhang H. S., Holmes M. C., Zhang L., Gregory P. D., Rebar E. J. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. (2011);29:143–148. doi: 10.1038/nbt.1755. [DOI] [PubMed] [Google Scholar]
- 29.Mussolino C., Morbitzer R., Lutge F., Dannemann N., Lahaye T., Cathomen T. A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity. Nucl. Acids Res. (2011);39:9283–9293. doi: 10.1093/nar/gkr597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bitinaite J., Wah D. A., Aggarwal A. K., Schildkraut I. FokI dimerization is required for DNA cleavage. Proc. Natl. Acad. Sci. U.S.A. (1998);95:10570–10575. doi: 10.1073/pnas.95.18.10570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cornu T. I., Thibodeau-Beganny S., Guhl E., Alwin S., Eichtinger M., Joung J. K., Cathomen T. DNA-binding specificity is a major determinant of the activity and toxicity of zinc-finger nucleases. Mol. Ther. (2008);16:352–358. doi: 10.1038/sj.mt.6300357. [DOI] [PubMed] [Google Scholar]
- 32.Miller J. C., Holmes M. C., Wang J., Guschin D. Y., Lee Y. L., Rupniewski I., Beausejour C. M., Waite A. J., Wang N. S., Kim K. A., Gregory P. D., Pabo C. O., Rebar E. J. An improved zinc-finger nuclease architecture for highly specific genome editing. Nat. Biotechnol. (2007);25:778–785. doi: 10.1038/nbt1319. [DOI] [PubMed] [Google Scholar]
- 33.Ramirez C. L., Foley J. E., Wright D. A., Muller-Lerch F., Rahman S. H., Cornu T. I., Winfrey R. J., Sander J. D., Fu F., Townsend J. A., Cathomen T., Voytas D. F., Joung J. K. Unexpected failure rates for modular assembly of engineered zinc fingers. Nat. Methods. (2008);5:374–375. doi: 10.1038/nmeth0508-374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kim J. S., Lee H. J., Carroll D. Genome editing with modularly assembled zinc-finger nucleases. Nat. Methods. (2010);7:91. doi: 10.1038/nmeth0210-91a. author reply 91-92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Joung J. K., Voytas D. F., Cathomen T. Reply to “Genome editing with modularly assembled zinc-finger nucleases”. Nat. Methods. (2010);7:91–92. doi: 10.1038/nmeth0210-91b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lieber M. R. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. (2010);79:181–211. doi: 10.1146/annurev.biochem.052308.093131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Vasquez K. M., Marburger K., Intody Z., Wilson J. H. Manipulating the mammalian genome by homologous recombination. Proc. Natl. Acad. Sci. U.S.A. (2001);98:8403–8410. doi: 10.1073/pnas.111009698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rouet P., Smih F., Jasin M. Expression of a site-specific endonuclease stimulates homologous recombination in mammalian cells. Proc. Natl. Acad. Sci. U.S.A. (1994);91:6064–6068. doi: 10.1073/pnas.91.13.6064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Porteus M. H., Baltimore D. Chimeric nucleases stimulate gene targeting in human cells. Science. (2003);300:763. doi: 10.1126/science.1078395. [DOI] [PubMed] [Google Scholar]
- 40.Meyer M., de Angelis M. H., Wurst W., Kuhn R. Gene targeting by homologous recombination in mouse zygotes mediated by zinc-finger nucleases. Proc. Natl. Acad. Sci. U.S.A. (2010);107:15022–15026. doi: 10.1073/pnas.1009424107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Meyer M., Ortiz O., Hrabe de Angelis M., Wurst W., Kuhn R. Modeling disease mutations by gene targeting in one-cell mouse embryos. Proc. Natl. Acad. Sci. U.S.A. (2012);109:9354–9359. doi: 10.1073/pnas.1121203109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mussolino C., Cathomen T. TALE nucleases: tailored genome engineering made easy. Curr. Opin. Biotechnol. (2012);23:644–650. doi: 10.1016/j.copbio.2012.01.013. [DOI] [PubMed] [Google Scholar]
- 43.Carlson D. F., Tan W., Lillico S. G., Stverakova D., Proudfoot C., Christian M., Voytas D. F., Long C. R., Whitelaw C. B., Fahrenkrug S. C. Efficient TALEN-mediated gene knockout in livestock. Proc. Natl. Acad. Sci. U.S.A. (2012);109:17382–17387. doi: 10.1073/pnas.1211446109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wood A. J., Lo T. W., Zeitler B., Pickle C. S., Ralston E. J., Lee A. H., Amora R., Miller J. C., Leung E., Meng X., Zhang L., Rebar E. J., Gregory P. D., Urnov F. D., Meyer B. J. Targeted genome editing across species using ZFNs and TALENs. Science. (2011);333:307. doi: 10.1126/science.1207773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Huang P., Xiao A., Zhou M., Zhu Z., Lin S., Zhang B. Heritable gene targeting in zebrafish using customized TALENs. Nat. Biotechnol. (2011);29:699–700. doi: 10.1038/nbt.1939. [DOI] [PubMed] [Google Scholar]
- 46.Sander J. D., Cade L., Khayter C., Reyon D., Peterson R. T., Joung J. K., Yeh J. R. Targeted gene disruption in somatic zebrafish cells using engineered TALENs. Nat. Biotechnol. (2011);29:697–698. doi: 10.1038/nbt.1934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tesson L., Usal C., Menoret S., Leung E., Niles B. J., Remy S., Santiago Y., Vincent A. I., Meng X., Zhang L., Gregory P. D., Anegon I., Cost G. J. Knockout rats generated by embryo microinjection of TALENs. Nat. Biotechnol. (2011);29:695–696. doi: 10.1038/nbt.1940. [DOI] [PubMed] [Google Scholar]
- 48.Geurts A. M., Cost G. J., Freyvert Y., Zeitler B., Miller J. C., Choi V. M., Jenkins S. S., Wood A., Cui X., Meng X., Vincent A., Lam S., Michalkiewicz M., Schilling R., Foeckler J., Kalloway S., Weiler H., Menoret S., Anegon I., Davis G. D., Zhang L., Rebar E. J., Gregory P. D., Urnov F. D., Jacob H. J., Buelow R. Knockout rats via embryo microinjection of zinc-finger nucleases. Science. (2009);325:433. doi: 10.1126/science.1172447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rao A., Richards T. L., Simmons D., Zahniser N. R., Sorkin A. Epitope-tagged dopamine transporter knock-in mice reveal rapid endocytic trafficking and filopodia targeting of the transporter in dopaminergic axons. FASEB J. (2012);26:1921–1933. doi: 10.1096/fj.11-196113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cui X., Ji D., Fisher D. A., Wu Y., Briner D. M., Weinstein E. J. Targeted integration in rat and mouse embryos with zinc-finger nucleases. Nat. Biotechnol. (2011);29:64–67. doi: 10.1038/nbt.1731. [DOI] [PubMed] [Google Scholar]