

Abstract
A close association between the obesity hormone leptin and breast cancer progression has been suggested. The present study investigated the molecular mechanism for enhanced leptin expression in breast cancer cells and its functional significance in breast cancer aggressiveness. We examined whether leptin expression level is affected by the oncoprotein human epidermal growth factor receptor2 (HER2), which is overexpressed in ∼30% of breast tumors. Here, we report, for the first time, that HER2 induces transcriptional activation of leptin in MCF10A human breast epithelial cells. We also showed that p38 mitogen-activated protein kinase signaling was involved in leptin expression induced by HER2. We showed a crucial role of leptin in the invasiveness of HER2-MCF10A cells using an siRNA molecule targeting leptin. Taken together, the results indicate a molecular link between HER2 and leptin, providing supporting evidence that leptin represents a target for breast cancer therapy. [BMB Reports 2012; 45(12): 719-723]
Keywords: Breast cancer, ErBb2, HER2, Leptin
INTRODUCTION
Breast cancer is one of the most frequent types of cancer among women (1), and metastasis is the major cause of mortality in patients with breast cancer. Metastasis and invasion require matrix-degrading activities mainly exerted by matrix metalloproteinases (MMPs) (2). Our previous studies revealed key roles of MMP-2 and MMP-9 in the induction of breast cell invasion (3-5). Mounting evidence suggests a correlation between obesity and breast cancer risk. Obesity has been related to high breast cancer risk in postmenopausal women (6), and obese patients with breast cancer show larger, more advanced tumors, and aggressive cancer pathological factors, including lymph node metastases, advanced tumor stage, and high grade compared to those in non-obese patients (7).
Leptin is a 16 kDa circulating peptide hormone secreted mainly from adipose tissue (8). Although the main function of leptin is to control energy balance and food intake via hypothalamic-mediated effects (9,10), accumulating evidence suggests that leptin is involved in carcinogenesis, particularly in breast cancer (6,11,12). Leptin and the ObR leptin receptor, which are overexpressed in breast tumors, correlate with worse prognosis and poor survival of breast cancer (6,13). A recent study on human placental cells revealed that 17β-estradiol increases leptin expression in which membrane-associate estrogen receptor (ER)-α is involved (14). Leptin expression is enhanced under hyperinsulinemia and hypoxic conditions in MCF-7 breast cancer cells (15).
Human epidermal growth factor receptor 2 (HER2), also called Neu or ErbB2, is overexpressed in about 30% of breast tumors (16). HER2 overexpression correlates with poor prognosis because it enhances invasive and metastatic phenotypes (16,17). We have previously shown that HER2 induces an invasive phenotype in human breast epithelial cells (18). Leptin induces proliferation of breast cancer cell lines in relation to ER status as well as to the presence or absence of HER2 (19).
Despite accumulating data supporting a close relationship between leptin and breast cancer progression, limited information is available on the molecular mechanism for enhanced leptin expression in breast cancer cells and its functional significance in breast cancer aggressiveness. In the present study, we found that leptin gene expression was increased markedly by HER2 in MCF10A human breast epithelial cells. We further elucidated the functional significance of leptin in the HER2-induced invasive phenotype of breast cells.
RESULTS AND DISCUSSION
HER2 induces expression of leptin by transcriptional activation
Immunoblot analyses to detect leptin were performed on HER2-overexpressing MCF10A cells to investigate the effect of HER2 on leptin expression (18). HER2 overexpression was confirmed by immunoblot analysis (Fig. 1A). Leptin protein level increased markedly in HER2-MCF10A cells compared to that in parental MCF10A cells (Fig. 1B), demonstrating that HER2 induces leptin expression in MCF10A cells. A reverse transcription-polymerase chain reaction (RT-PCR) analysis showed that leptin mRNA level was increased significantly by HER2 (Fig. 1C), indicating that HER2 upregulated leptin at the transcriptional level.
Fig. 1. HER2 induces leptin expression by transcriptional activation. (A) HER2 expression was detected in cell lysates by immunoblot analysis. β-actin was used as the loading control. (B) Leptin expression was examined by immunoblot analysis. (C) Leptin mRNA level was detected by RT-PCR analysis. β-actin was used as the loading control. Band intensities were quantified and plotted. The results are means + S.E. of triplicates. *Statistically different at P < 0.05.
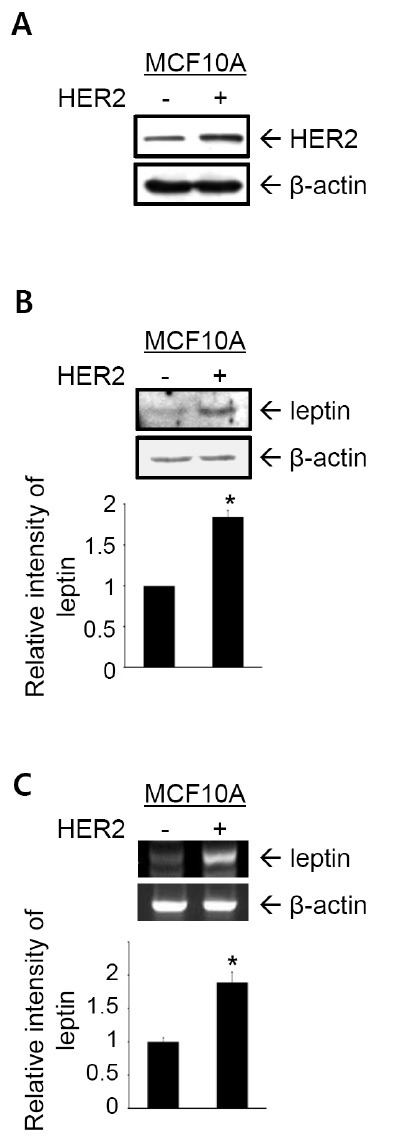
Involvement of ER in the regulatory mechanism for leptin expression has been elucidated in human placental cell line (14), and in the MCF-7 ER-positive breast cancer cell line (15). Our findings demonstrate that leptin expression is induced by HER2 in MCF10A ER-negative cell line, suggesting an ER-independent regulation of leptin expression.
p38 mitogen protein activated protein kinase (MAPK) is required for leptin expression induced by HER2
We next investigated the signaling pathway involved in HER2-induced leptin expression. Activation of HER2 by macrophage inhibitory cytokine-1 (MIC-1) was suggested to promote the ability of tumor cells to activate Akt and MAPKs signaling (20). Our previous study showed that Rac1, p38 MAPK, and Akt are activated by HER2 in MCF10A cells (18). In the present study, we assessed the role of p38 MAPK signaling in leptin expression. HER2-MCF10A cells were treated with SB203580, a pharmacological inhibitor of p38 MAPK (21). As shown in Fig. 2, leptin expression level in HER2-MCF10A cells decreased significantly by inhibiting p38 MAPK. These results demonstrate that the p38 MAPK signaling pathway is involved in HER2-induced leptin expression.
Fig. 2. p38 MAPK is required for leptin expression induced by HER2. HER2-MCF10A cells were pretreated with 50 μM SB203580 for 24 hr. Expression of leptin, phosphorylated p38 (pp38) MAPK, and total p38 MAPK were detected in cell lysates by immunoblot analysis. Band intensities were quantified and plotted. The results are means + S.E. triplicates. *Statistically different at P < 0.05.
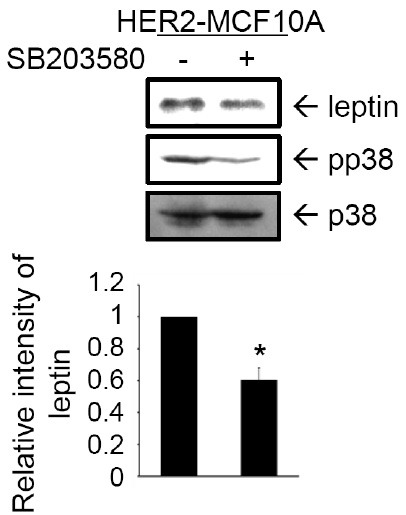
Recent studies have revealed the signaling pathways that mediate leptin expression. Extracellular regulated kinases (ERKs) and PI3K signal transduction pathways mediate leptin expression through 17β-estradiol in human placental cells (22). The role of hypoxia-inducible factor-1α signaling has been demonstrated in leptin expression in MCF-7 cells treated with insulin or cultured under hypoxic conditions (15). Further studies are required to elucidate additional signaling transduction pathways responsible for the induction of leptin expression in HER2-MCF10A cells.
The invasive phenotype induced by HER2 requires leptin expression
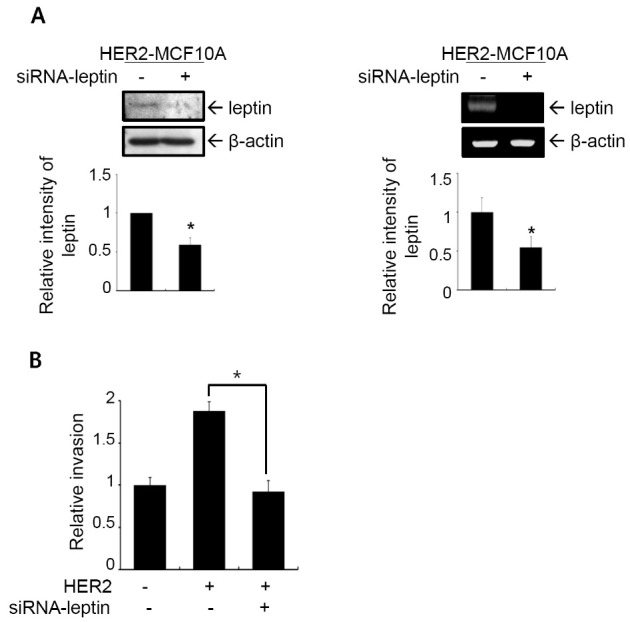
Leptin promotes growth, migration, invasion, and angiogenesis of breast cancer cells (6,23), and leptin enhances invasion and migration of breast cancer cells through crosstalk with insulin-like growth factor-1 (24). We performed an in vitro invasion assay in HER2-MCF10A cells in which leptin expression was knocked-down by an siRNA targeting leptin to investigate the functional significance of leptin in the HER2-induced invasive phenotype of MCF10A cells. Leptin knockdown was confirmed by immunoblot analysis (Fig. 3A, left) and RT-PCR analysis (Fig. 3A, right). Invasiveness was inhibited by 49.3% in HER2-MCF10A cells in which leptin was knocked-down by siRNA (Fig. 3B). These results demonstrate that leptin plays a crucial role in the induction of breast cell invasion by HER2.
Fig. 3. Invasive phenotype induced by HER2 requires leptin expression. (A) HER2-MCF10A cells were transfected with control siRNA or siRNA targeting leptin. Leptin knock-down was confirmed by immunoblot analysis (left) and RT-PCR analysis (right). Band intensities were quantified and plotted. The results are means + S.E. of triplicates. *Statistically different at P < 0.05. (B) Cells were subjected to the in vitro invasion assay. The number of invaded cells per field was counted (×400) in 13 arbitrary visual fields. The results represent means + S.E. of triplicates. *Statistically different from control at P < 0.05.
Leptin plays a crucial role in MMP-2 and MMP-9 expressions in HER2-MCF10A cells
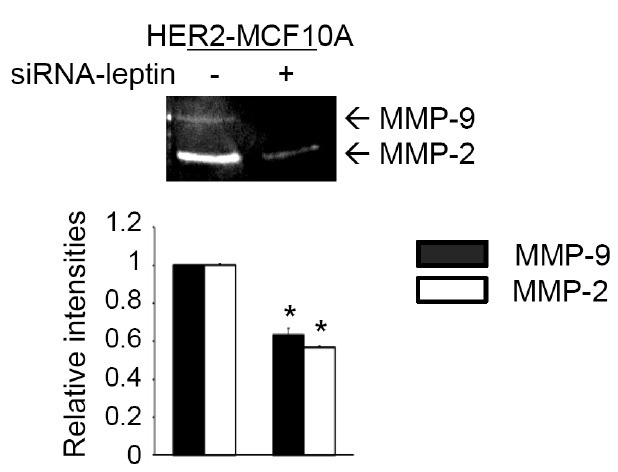
Among MMPs, MMP-2 is regulated by leptin in adult cardiac fibroblasts (25) and in rat glomerular mesangial cells (26). Our previous study showed that HER2-induced invasion of MCF10A cells involves MMP-9 (18). We next investigated the role of leptin in MMP-2 and/or MMP-9 expression. A gelatin zymogram assay was performed to detect MMP-2 and MMP-9 in HER2-MCF10A cells transfected with an siRNA targeting leptin. As shown in Fig. 4, levels of both MMP-9 and MMP-2 decreased significantly following leptin knockdown. These data demonstrate an essential role for leptin in MMP-2 and MMP-9 expression in HER2-MCF10A cells.
Fig. 4. Leptin plays a crucial role in the expressions of MMP-2 and MMP-9 in HER2-MCF10A cells. HER2-MCF10A cells were transfected with control siRNA or siRNA targeting leptin. Conditioned media were collected, and gelatinolytic activities of secreted MMP-2 and MMP-9 were determined by gelatin zymogram assay. Band intensities were quantified and plotted. The results are means + S.E. of triplicates. *Statistically different at P < 0.05.
MATERIALS AND METHODS
Cell lines
MCF10A cells were purchased and cultured as described previously (27). The cells were cultured in DMEM/F12 supplemented with 5% horse serum, 0.5 μg/ml hydrocortisone, 10 μg/ml insulin, 20 ng/ml epidermal growth factor, 0.1 μg/ml cholera enterotoxin, 100 units/ml penicillin-streptomycin, 2 mM L-glutamine, and 0.5 μg/ml amphotericin B. HER2-MCF10A cells were established as described previously (18).
Immunoblot analysis
Immunoblot analysis was performed as described previously (28). Anti-HER2 and anti-leptin antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Anti-β-actin antibody was purchased from Sigma-Aldrich (St. Louis, MO, USA). Anti-p38 MAPK and anti-pp38 MAPK antibodies were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). The enhanced chemiluminescence system (Amersham-Pharmacia, Buckinghamshire, UK) was used for detection. Relative band intensities were determined by quantifying each band using an Image Analyzer (Vilber Lourmet, France).
RT-PCR
The sequences of the leptin sense and antisense primers were 5'-CTGTGCCCATCCAAAAAGTCC-3' and 5'-CCCCCAGGCTGTCCAAGGTC-3', respectively. The sequences of the β-actin sense and antisense primers were 5'-ACTCTTCCAGCCTTCCTT-3' and 5'-TCTCCTTCTGCATCCTGTC-3', respectively. We used the experimental conditions for the reverse transcription and thermocycler as described previously (6).
In vitro invasion assay
The in vitro invasion assay was performed using a 24-well Transwell unit with polycarbonate filters (Corning Costar, Cambridge, MA, USA), as described previously (29). The lower side of the filter was coated with type I collagen, and the upper side was coated with Matrigel (Collaborative Research, Lexington, KY, USA). The lower compartment was filled with serum-free medium containing 0.1% BSA. Cells were placed in the upper part of the Transwell plate, incubated for 17 hr, fixed with methanol, and stained with hematoxylin for 10 min followed briefly by staining with eosin. The invasive phenotypes were determined by counting the cells that migrated to the lower side of the filter by microscopy at 400×. Thirteen fields were counted for each filter, and each sample was assayed in triplicate.
Leptin knockdown by small-interfering (si) RNA
Leptin knockdown was performed with an siRNA molecule targeting leptin (Santa Cruz Biotechnology). Cells were plated in 6-well plates at 1.5 × 105 cells/well, grown for 24 hours, and then transfected with 100 pmol siRNA for 6 hr using Lipofectamine 2000 reagent and OPTI-MEMI (Invitrogen, Carlsbad, CA, USA). Control cells were treated with negative control siRNA (Santa Cruz Biotechnology).
Gelatin zymogram assay
Cells were cultured in serum-free DMEM/F12 medium for 24 hr. Gelatinolytic activity of the conditioned medium was determined by the gelatin zymogram assay, as described previously (27).
Acknowledgments
This study was supported by the Duksung Women’s University Research Grant 2010.
References
- 1.Jemal A., Siegel R., Ward E., Murray T., Xu J., Thun M. J. Cancer statistics, 2007. CA Cancer J. Clin. (2007);57:43–66. doi: 10.3322/canjclin.57.1.43. [DOI] [PubMed] [Google Scholar]
- 2.Stetler-Stevenson W. G. Matrix metalloproteinases in angiogenesis: a movingtarget for therapeutic intervention, J. Clin. Invest. (1999);103:1237–1241. doi: 10.1172/JCI6870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kim M. S., Lee E. J., Choi Kim H. R., Moon A. p38 kinase is a key signaling molecule for H-ras-induced cell motility and invasive phenotype in human breast epithelial cell. Cancer Res. (2003);63:5454–5461. [PubMed] [Google Scholar]
- 4.Song H., Ki S. H., Kim S. G., Moon A. Activating transcription factor 2 mediates matrix metalloproteinase-2 transcriptional activation induced by p38 in breast epithelial cells. Cancer Res. (2006);66:10487–10496. doi: 10.1158/0008-5472.CAN-06-1461. [DOI] [PubMed] [Google Scholar]
- 5.Kim E. S., Kim J. S., Kim S. G., Hwang S., Lee C. H., Moon A. Sphingosine 1-phosphate regulates matrix metalloproteinase-9 expression and breast cell invasion through S1P3-Gαq coupling. J. Cell Sci. (2011);124:2220–2230. doi: 10.1242/jcs.076794. [DOI] [PubMed] [Google Scholar]
- 6.Garofalo C., Koda M., Cascio S., Sulkowska M., Kanczuga-Koda L., Golaszewska J., Russo A., Sulkowski S., Surmacz E. Increased expression of leptin and the leptin receptor as a marker of breast cancer progression: possible role of obesity-related stimuli. Clin. Cancer Res. (2006);12:1447–1453. doi: 10.1158/1078-0432.CCR-05-1913. [DOI] [PubMed] [Google Scholar]
- 7.Porter G. A., Inglis K. M., Wood L. A., Veugelers P. J. Effect of obesity on presentation of breast cancer. Ann. Surg. Oncol. (2006);13:327–332. doi: 10.1245/ASO.2006.03.049. [DOI] [PubMed] [Google Scholar]
- 8.Garofalo C., Surmacz E. Leptin and cancer. J. Cell Physiol. (2006);207:12–22. doi: 10.1002/jcp.20472. [DOI] [PubMed] [Google Scholar]
- 9.Zhang Y., Proenca R., Maffei M., Barone M., Leopold L., Friedman J. M. Positional cloning of the mouse obese gene and its human homologue. Nature. (1994);372:425–432. doi: 10.1038/372425a0. [DOI] [PubMed] [Google Scholar]
- 10.Schwartz M. W., Baskin D. G., Kaiyala K. J., Woods S. C. Model for the regulation of energy balance and adiposity by the central nervous system 1-3. Am. J. Clin. Nut. (1999);69:584–596. doi: 10.1093/ajcn/69.4.584. [DOI] [PubMed] [Google Scholar]
- 11.Vona-Davis L., Howard-McNatt M., Rose D. P. Adiposity, type 2 diabetes and the metabolic syndrome in breast cancer. Obes. Rev. (2007);8:395–408. doi: 10.1111/j.1467-789X.2007.00396.x. [DOI] [PubMed] [Google Scholar]
- 12.Schaffler A., Scholmerich J., Buechler C. Mechanisms of disease: adipokines and breast cancer - endocrine and paracrine mechanisms that connect adiposity and breast cancer. Nat. Clin. Pract. Endocrinol. Metab. (2007);3:345–354. doi: 10.1038/ncpendmet0456. [DOI] [PubMed] [Google Scholar]
- 13.Ishikawa M., Kitayama J., Nagawa H. Enhanced expression of leptin and leptin receptor (OB-R) in human breast cancer. Clin. Cancer Res. (2004);10:4325–4331. doi: 10.1158/1078-0432.CCR-03-0749. [DOI] [PubMed] [Google Scholar]
- 14.Gambino Y. P., Pérez Pérez A., Dueñas J. L., Calvo J. C., Sánchez-Margalet V., Varone C. L. Regulation of leptin expression by 17beta-estradiol in human placental cells involves membrane associated estrogen receptor alpha. Biochim. Biophys. Acta. (2012);1823:900–910. doi: 10.1016/j.bbamcr.2012.01.015. [DOI] [PubMed] [Google Scholar]
- 15.Cascio S., Bartella V., Auriemma A., Johannes G. J., Russo A., Giordano A., Surmacz E. Mechanism of leptin expression in breast cancer cells: role of hypoxia- inducible factor-1alpha. Oncogene. (2008);27:540–547. doi: 10.1038/sj.onc.1210660. [DOI] [PubMed] [Google Scholar]
- 16.Slamon D. J., Clark G. M., Wong S. G., Levin W. J., Ullrich A., McGuire W. L. Human breast cancer: correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science. (1987);235:177–182. doi: 10.1126/science.3798106. [DOI] [PubMed] [Google Scholar]
- 17.Yarden Y. Biology of HER2 and its importance in breast cancer. Oncology. (2001);61:1–13. doi: 10.1159/000055396. [DOI] [PubMed] [Google Scholar]
- 18.Kim I. Y., Yong H. Y., Kang K. W., Moon A. Overexpression of ErbB2 induces invasion of MCF10A human breast epithelial cells via MMP-9. Cancer Letters. (2009);275:227–233. doi: 10.1016/j.canlet.2008.10.013. [DOI] [PubMed] [Google Scholar]
- 19.Ray A., Nkhata K. J., Cleary M. P. Effects of leptin on human breast cancer cell lines in relationship to estrogen receptor and HER2 status. Int. J. Oncol. (2007);30:1499–1509. [PubMed] [Google Scholar]
- 20.Park Y. J., Lee H., Lee J. H. Macrophage inhibitory cytokine-1 transactivates ErbB family receptors via the activation of Src in SK-BR-3 human breast cancer cells. BMB Rep. (2010);43:91–96. doi: 10.5483/BMBRep.2010.43.2.091. [DOI] [PubMed] [Google Scholar]
- 21.Cuenda A., Rouse J., Doza Y. N., Meier R., Cohen P., Gallagher T. F., Young P. R., Lee J. C. SB 203580 is a specific inhibitor of a MAP kinase homologue which is stimulated by cellular stresses and interleukin-1. FEBS Lett. (1995);364:229–233. doi: 10.1016/0014-5793(95)00357-F. [DOI] [PubMed] [Google Scholar]
- 22.Gambino Y. P., Maymó J. L., Pérez-Pérez A., Dueñas J. L., Sánchez-Margalet V., Calvo J. C., Varone C. L. 17Beta-estradiol enhances leptin expression in human placental cells through genomic and nongenomic actions. Biol Reprod. (2010);83:42–51. doi: 10.1095/biolreprod.110.083535. [DOI] [PubMed] [Google Scholar]
- 23.Surmacz E. Obesity hormone leptin: a new target in breast cancer? Breast Cancer Res. (2007);9:301. doi: 10.1186/bcr1638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Saxena N. K., Taliaferro-Smith L., Knight B. B., Merlin D., Anania F. A., O R. M., Sharma D. Bidirectional crosstalk between leptin and insulin-like growth factor-I signaling promotes invasion and migration of breast cancer cells via transactivation of epidermal growth factor receptor. Cancer Res. (2008);68:9712–9722. doi: 10.1158/0008-5472.CAN-08-1952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Schram K., Ganguly R., No E. K., Fang X., Thong F. S., Sweeney G. Regulation of MT1-MMP and MMP-2 by leptin in cardiac fibroblasts involves Rho/ ㅌROCK-dependent actin cytoskeletal reorganization and leads to enhanced cell migration. Endocrinology. (2011);152:2037–2047. doi: 10.1210/en.2010-1166. [DOI] [PubMed] [Google Scholar]
- 26.Lee M. P., Madani S., Sekula D., Sweeney G. Leptin increases expression and activity of matrix metalloproteinase-2 and does not alter collagen production in rat glomerular mesangial cells. Endocr. Res. (2005);31:27–37. doi: 10.1080/07435800500320653. [DOI] [PubMed] [Google Scholar]
- 27.Moon A., Kim M. S., Kim T. G., Kim S. H., Kim H. E., Chen Y. Q., Choi Kim H. R. H-ras but not N-ras induces an invasive phenotype in human breast epithelial cells: a role for MMP-2 in the H-ras-induced invasive phenotype. Int. J. Cancer. (2000);85:176–181. doi: 10.1002/(sici)1097-0215(20000115)85:2<176::aid-ijc5>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- 28.Kim M. S., Lee E. J., Choi Kim H. R., Moon A. p38 kinase is a key signaling molecule for H-ras-induced cell motility and invasive phenotype in human breast epithelial cell. Cancer Res. (2003);63:5454–5461. [PubMed] [Google Scholar]
- 29.Zhang C., Li Y., Shi X., Kim S. K. Inhibition of the expression on MMP-2, 9 and morphological changes via human fibrosarcoma cell line by 6,6'-bieckol from marine alga Ecklonia cava. BMB Rep. (2010);43:62–68. doi: 10.5483/BMBRep.2010.43.1.062. [DOI] [PubMed] [Google Scholar]