

Abstract
Transcutaneous immunization refers to the topical application of antigens onto the epidermis. Transcutaneous immunization targeting the Langerhans cells of the skin has received much attention due to its safe, needle-free, and noninvasive antigen delivery. The skin has important immunological functions with unique roles for antigen-presenting cells such as epidermal Langerhans cells and dermal dendritic cells. In recent years, novel vaccine delivery strategies have continually been developed; however, transcutaneous immunization has not yet been fully exploited due to the penetration barrier represented by the stratum corneum, which inhibits the transport of antigens and adjuvants. Herein we review recent achievements in transcutaneous immunization, focusing on the various strategies for the enhancement of antigen delivery and vaccination efficacy. [BMB Reports 2013; 46(1): 17-24]
Keywords: Antigen delivery, Epidermis, Transcutaneous immunization, Vaccine
INTRODUCTION
Current vaccine delivery methods use the intradermal (ID), intramuscular (IM), subcutaneous (SC), intranasal (IN), and oral (OR) routes (1). Of these, IM, SC, and ID immunizations are classified as needle-based techniques. Needle-based methods are effective in achieving the desired immune responses; however, their use has numerous drawbacks, such as safety, cost and compliance. The development of safer and more effective vaccines is a global priority. One technology under development administers vaccines through the skin painlessly via a larger surface area than that used by a needle. Referred to as transcutaneous immunization (TCI), topical application of an antigen onto the skin targets immune cells resident therein, such as Langerhans cells (2). However, the skin poses a formidable barrier to hydrophilic macromolecules such as proteins, peptides, and vaccines due to the impermeability of the stratum corneum (3). To overcome the skin barrier to noninvasive delivery, numerous techniques for transcutaneous delivery are being developed. Significant advances in transcutaneous delivery of proteins, peptides, and large hydrophilic molecules have in recent years shown great promise, although commercialization of these products remains a future prospect. Clinical and pre-clinical trials in animals to test the safety and efficacy of vaccines using various TCI techniques and antigens are being conducted (4). This review will focus on the developments in TCI, with special emphasis on the concept of skin immunity and the recent achievements of transcutaneous antigen delivery systems.
SKIN IMMUNE SYSTEM
The skin is an immunologically active site composed of three layers: the stratum corneum, epidermis, and dermis (5). The barrier function of the skin resides in the stratum corneum, which is composed of dead corneocytes. Directly below the stratum corneum is the epidermis, underneath which is the dermis (6). The skin immune system includes keratinocytes and Langerhans cells (LCs) in the epidermis; fibroblasts, dendritic cells (DCs), and mast cells in the dermis; and T and B lymphocytes in the skin-draining lymph nodes (1). The number of resident T lymphocytes in the skin is approximately twice the total number of those circulating in the blood (7). Easy access to this skin immune system makes it an attractive site for vaccination.
LCs, specialized dendritic cells that contain large Birbeck granules, form a network around keratinocytes in the suprabasal layer of the epidermis. They represent a network of migratory immune cells that underlie approximately 25% of the total skin surface area, and can transport antigens from the skin into lymph nodes to elicit immune responses (8). The transfer of antigens to lymphoid organs is a critical step in the induction of immune responses. Immature LCs are typically located in the basal layers of the epidermis, where they sample and process antigens (1).
After encountering antigen in the epidermis, LCs become activated and undergo maturation, acquiring the ability to capture antigens and process them into immunogenic peptides, which are presented as complexes with major histocompatibility complex (MHC) proteins on their surfaces (1,6). These cells then migrate via afferent lymphatics to the skin-draining lymph nodes, where they present antigens to the resident lymphocytes. During their migration via the afferent lymphatics they mature and acquire the capacity to stimulate helper or cytotoxic activity in naïve resting T cells. Subsequent T cell activation results in the generation of both effector and memory cells, which play distinct roles in protection against various infections.
LCs are recognized mainly by virtue of their human leukocyte antigen (HLA)-DR, the adhesion marker E-cadherin, and unique cytoplasmic organelles named Birbeck granules (BGs) or Langerhans granules (1, 6). The C-type lectin Langerin (CD207) is responsible for BG formation and is a key marker of the LC lineage (9). LCs also strongly express CD1a (10). LC migration and maturation are associated with the expression of chemokine receptors on the surface of the cells. During LC maturation, the skin-homing chemokine receptors CCR1, CCR2, CCR5, and CCR6 are down-regulated, while receptors involved in homing to lymph nodes (CCR4, CXCR4, and CCR7) are up-regulated (11). Even in the absence of inflammatory signals, approximately 3% of LCs circulate naturally from the skin to the lymph nodes. The chemokine receptor CCR7 regulates the migration of LCs to the lymph nodes under the influence of secondary lymphoid tissue chemokines such as CCL20 (1, 12).
DCs are key regulators in the immune system. They constitute a link between the innate and adaptive immune systems, and determine whether immunity or tolerance is induced. In addition to classical LCs in the epidermis, several distinct subsets of Langerin-negative dermal DCs (dDCs) have been characterized, including CD14+ dDCs and CD1a+ dDCs in human skin (4, 12). CD14+ dDCs are most easily characterized by their expression of CD14, in addition to DC-SIGN (DC-specific intercellular adhesion molecule-3 (ICAM-3)-grabbing non-integrin, CD209), CD11c and CD11b (4, 13, 14). Dermal CD1a+ dDCs were shown to express CD1a, CD11c, and CD11b (15). In contrast, Langerin-positive CD103+ dDCs in mice showed low expression of CD11b and high expression of CD103 (16).
The distinct DC subsets in human skin differ with respect to their expression of cell markers and functions in the adaptive immune response. Human LCs are involved in the function of CD4+ and CD8+ T cells, and they preferentially induce the secretion of Th2-type cytokines (17). The CD1a+ dDCs also elicited some secretion of Th2 type cytokines, but less than that elicited by LCs. CD14+ dDCs were reported to promote the differentiation of naïve B cells into IgM-secreting plasma cells via the induction of CD4+ T cells and secretion of interleukin (IL)-6 and IL-12 (18,19), whereas LCs did not (17). Even though the precise role of skin DCs in the immune response has not been determined, soluble antigens can directly diffuse to the draining lymph nodes, and can be taken up by the large population of DCs therein (20). This topic is important for the design of novel vaccines targeting DC subsets.
TRANSCUTANEOUS IMMUNIZATION
TCI, the topical application of antigen onto the epidermis, is an emerging vaccine delivery system. Due to the easy access to the skin immune system by topical application of antigens, TCI is considered a promising technique for vaccine delivery. However, topical antigen application is limited by the impermeable barrier function of the stratum corneum, the outermost layer of the epidermis (21). For an antigen to be delivered passively via the stratum corneum, it must have suitable lipophilicity and a molecular mass of <500 Da (1,21,22).
The stratum corneum is comprised of dead corneocytes interspersed within a lipid-rich matrix. Its structure and lipophilic nature account for the barrier function of the skin. The stratum corneum exhibits selective permeability and allows only relatively lipophilic compounds to diffuse into the lower layers. Passive topical penetration of hydrophilic molecules and those of molecular mass >500 Da is difficult.
In recent years, advances in biotechnology have led to the development of techniques and instruments that can overcome the difficulties associated with the stratum corneum barrier. To overcome the skin barrier to noninvasive delivery, various sophisticated delivery systems, such as passive and active strategies to optimize percutaneous delivery, have emerged (23). The passive method needs strategies that increase permeability, such as optimization of formulation or vaccine carrier. Active methods rely on sophisticated delivery systems, such as complicated nanocarriers or mechanical devices, to enhance delivery. The delivery of antigens of different lipophilicities and molecular weights has been shown to be improved by active methods such as iontophoresis (24,25), electroporation (26), laser ablation (27), micro-/nano-needle techniques (28) and ultrasound (29). However, the challenges in creating such systems are the complicated processes involved and the potential negative impact on antigen stability. Moreover, the high manufacturing costs of these advanced systems often offset their advantages.
To circumvent these problems with the current methods, a novel strategy of creating “skin-permeable proteins” that possess a skin-penetrating ability and thereby eliminate the need for a transport vehicle was presented (3). Artificial skin-permeable proteins by simple conjugation of a protein to a cell-penetrating peptide, LMWP, which transported the attached protein into the skin by penetrating the stratum corneum barrier, were chemically constructed. This immunization strategy showed an advantage over conventional vaccination: it elicited specific immune responses. This methodology for constructing artificial skin-permeable antigens offered simple needle-free vaccination without the need for sophisticated vaccine carriers or expensive medical devices (3).
Bacterial and viral infectious diseases, neurodegenerative diseases, autoimmune diseases, cancers and biological weapons based on B. anthracis are promising targets of TCI vaccination (6,30).
Many complex events take place in the skin after vaccination to mount an effective immune response. Many parameters, including vaccine type, vaccination technique and the plasticity of skin antigen-presenting cells (APCs) targeted by vaccination will determine the outcome and effectiveness of the immune response. Studies with murine models have demonstrated that the vaccine dose used during TCI often determines the resultant immune response. It is now agreed that transcutaneous vaccination with low-dose antigen doses not distribute the antigen throughout the dermis, but targets solely epidermal LCs and stimulates Th1 and cytotoxic T lymphocyte (CTL) responses (1).
VACCINE CANDIDATES FOR TRANSCUTANEOUS IMMUNIZATION
Here, we will briefly discuss the wide variety of agents that have been studied as active vaccine candidates for TCI. DNA vaccines refer to plasmid DNA that encodes the antigenic protein (6). Upon introduction to the target cells, the protein antigen is expressed and induces an immune response against the antigen. Several DNA vaccines; for example those for influenza (31) and HBsAg (32), have been demonstrated to induce immune responses via TCI.
Immunodominant epitopes have been used widely as subunit vaccines, and are advantageous for TCI due to their short peptide sequences and ease of delivery across the skin (6,33). Specific peptide sequences from the antigen can improve immunogenicity for TCI via inducing a CD4+ T helper response (6,34) or CD8+ cytotoxic T cell response (6,35). Human recombinant proteins are probably the most widely used vaccine candidates to date. Notably, full-length protein vaccines can be delivered across the skin by topical application, resulting in induction of immune responses. Hydrogel patches (36), hyperthermia (37), and conjugation to cell-penetrating peptides (CPPs) stimulated delivery of the proteins across the stratum corneum (3,38). CPPs were notably able to transduce attached cargos, such as proteins or DNA, across many types of biological barrier, including the intestinal wall, nasal mucosa, and even the blood–brain barrier.
TCI with a carbohydrate-based vaccine using a synthetic hexasaccharide of the O-specific polysaccharide of V. cholerae conjugated to bovine serum albumin was immunogenic (6). Moreover, transcutaneous and subcutaneous boosting with a neoglycoconjugate following oral cholera vaccination boosted protective immune responses against V. cholerae (39). The capsule polysaccharide of H. influenzae b was conjugated to cross-reacting material, and induced a strong humoral immune response after TCI (40). Glycoproteins and glycolipids, expressed on the surface of cancer cells, were used to develop cancer immunotherapy (41,42).
TRANSCUTANEOUS ANTIGEN DELIVERY TECHNOLOGY
Numerous technologies using diverse methods to transport antigens across the skin have been developed (Fig. 1). Humoral, CTL, and mucosal immune responses are induced via TCI. Here, we will review key advances in transcutaneous antigen delivery technologies with diverse antigen models, focusing on the most-commonly studied vaccine carriers.
Fig. 1. Schematic illustration of several transcutaneous antigen delivery technologies used for transcutaneous vaccination. (A) Patch, (B) E. coli vectors/Viral vectors, (C) Vesicular systems, such as liposomes, niosomes, transfersomes, and vesosomes, (D) Ultrasound, (E) Nanoparticles or Microemulsions, (F) Iontophoresis or Electroporation (G) Micro-needles/Nano-needles, (H) Cell penetrating peptide-conjugated proteins (I) Tape-stripping.
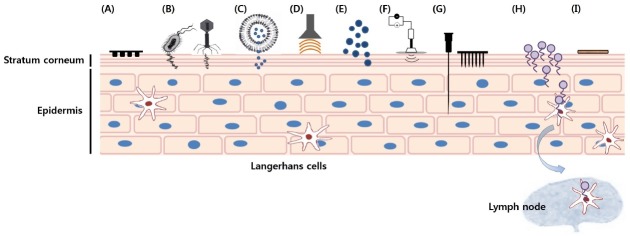
Combadiere and Liard categorized transcutaneous antigen delivery technologies into three generations, although some of the current TCI technologies can’t be properly classified (1). First-generation TCI technologies include those that use adsorption and passive distribution of very small lipophilic molecules across the stratum corneum. Most of the patches, gels, and sprays currently in clinical practice are first-generation TCI technologies. Patch-based TCI with whole inactivated viruses such as influenza (43) and herpes simplex virus (44) can induce humoral and cellular immune responses. Moreover, intact Escherichia coli particles can be used to activate APCs and T cells, thereby acting as natural adjuvants during TCI (45).
Second-generation TCI technologies comprise those that reformulate vaccine preparations, such as encapsulation in liposomes (46), transfersomes (47), and micro-emulsions (48). Exterior energetic methods of forcing antigens across the stratum corneum, including iontophoresis and ultrasound, also belong to this generation, although hydrophilic macromolecules are not always transported across the skin. Commonly employed vesicular systems for TCI, including niosomes (49) and vesosomes (50) have gained much attention recently as carriers of DNA and recombinant protein immunogens for TCI. Vesicular systems have been suggested to target epidermal LCs through hair follicles or pilosebaceous routes (6). In addition, vesicular systems have the following advantages: enhanced antigen stability, sustained antigen release, and synergistic function as an adjuvant. Nanoparticles targeting LCs in the hair follicle are a potentially useful TCI strategy, much like vesicular systems.
Third-generation TCI technologies include those that disrupt the stratum corneum barrier transiently and in a highly localized manner. Methods using micro-needles (including solid micro-needles, and coated, hollow, and dissolvable micro-needle arrays), lower-frequency ultrasound, and a pore-forming peptide named magainine (4,51) are under development with the goal of creating transient pores in the stratum corneum. Other techniques include thermal abrasion, using either heating patches or micro-lasers, and mechanical abrasion, such as shaving or friction, sandpapering, tape-stripping, and cyanoacrylate skin surface stripping (1).
ADJUVANTS FOR TRANSCUTANEOUS ANTIGEN DELIVERY
Adjuvants are the most important issue in TCI, because the immune response is adjuvant-dependent. A large number of potent adjuvants that enhance the immune response after TCI have emerged. Co-application of adjuvants with antigens is required to elicit successful immune responses by TCI (21,52). Successful adjuvants for vaccines include bacterial toxins, microbial products, cytokines, liposomes, oligodeoxynucleotides, and mineral salts (53). Of these, the most commonly used adjuvants, including alum, monophosphoryl lipid A, and an MF59 emulsion, are undesirable for TCI, likely due to their limited transport across the stratum corneum (4,54). Moreover, some adjuvants are too toxic for clinical use. To achieve a strong, directed immune response and reduce nonspecific adverse inflammatory responses, the optimal adjuvant must be identified. Moreover, newer vaccines based on recombinant proteins, synthetic peptides, and plasmid DNA tend to induce weaker immune responses, so proper adjuvants for selective activation of the receptors on skin DC subsets are vital. Here, we will discuss briefly the most promising TCI adjuvants.
ADP-ribosylating exotoxins
Bacterial ADP-ribosylating exotoxins, such as cholera toxin (CT) and E. coli heat-labile toxin (LT), and their derivatives, have received considerable attention owing to their marked adjuvanticity (6,55,56). They are used most often in pre-clinical studies of transcutaneous antigen delivery systems (4). CT and LT have ADP-ribosyl transferase activity, and bind to the GM1-ganglioside receptor (4,57). Application of CT to intact mouse skin resulted in not only anti-CT antibody production (4,39,56), but also provoked immune responses against antigens (58). Although the exact mechanisms are unclear, CT diffuses rapidly throughout the epidermis and results in a cAMP enhancement, which may stimulate the secretion of proinflammatory cytokines and tumor necrosis factor-α (6). These may in turn promote the activation LCs and their migration to regional lymph nodes. Novel TCI system induced an antigen-specific antibody response in the presence of sodium salicylate even without adjuvants, such as CT (53). Sodium salicylate had an adjuvant-like effect via activation of antigen presenting cells. Besides antibody production, CT was superior to other adjuvants in its ability to prime memory CD8 +T cells (58). The adjuvant effect of CT was independent of typical TLR and inflammasome signaling pathways, but CT adjuvanticity required Type-I IFN responses (59). Bacterial ADP-ribosylating exotoxins have been widely used and are popular adjuvants in TCI studies (60). However, application of CT to intact mouse skin can induce and enhance autoimmune disease (61). Specifically, dermal exposure to CT, with or without autoantigen, exacerbated the severity of autoimmune disease. Moreover, inclusion of adjuvants that exhibit high toxicity via the oral or intranasal routes always poses a safety concern, although they are relatively safe when used with TCI. So far, researchers have been unable to find a safer alternative; thus application of ADP-ribosylating exotoxin in a pharmaceutical product remains a challenge.
TLR ligands
Innate immunity is mediated by pathogen-recognition receptors (PRRs). Toll-like receptors (TLRs) are important PRRs involved in host defense (4). The distribution of TLRs in immune-active skin cells differs according to cell type. This heterogeneity may modulate the immune responses to TCI adjuvants. Therefore, use of TLR ligands as adjuvants for transcutaneous antigen vaccination is appropriate. One example is imiquimod, a synthetic immunomodulatory agonist of TLR7, which can activate dendritic cells and B cells to induce cytokines optimal for Th1 cell immunity, thus enhancing antibody production (62). A microneedle delivery of imiquimod with influenza subunit vaccine elicited strong immune responses compared to vaccine alone in skin-based influenza vaccines (63). Gardiquimod (64) and R848 (resiquimod) (65) are the ligands known to activate TLR7/TLR8 most strongly. As a vaccine adjuvant, resiquimod provoked prominent Th1 immune responses and promoted priming of CTLs to antigens.
CpG
Bacterial CpG motifs are unmethylated CpG dinucleotides within nucleic acid motifs that are recognized by vertebrate TLR9 (4,66-68). Under the influence of TLR9, CpG induced the secretion of the pro-inflammatory cytokines IL-12, TNF-α and IFN-γ, resulting in both Th1 and Th2 immune responses with a bias towards Th1 type (69). The adjuvanticity of CpG was dependent on the immunization route and carrier (70). CpG functioned as a TCI adjuvant when co-administered with DT, the model antigen ovalbumin, or DNA vaccine encoding influenza M protein, resulting in elevated antibody titers (4,69,71). Notably, topical application of an HIV peptide gp160 together with a mixture of CT and CpG induced a strong HIV-specific CTL response (4,72). CpG-ODN acted synergistically with tape-stripping (6,73) or CT in the lipid-based adjuvants Lipid C matrix (74). Therefore, co-administration of CpG-ODNs with other adjuvants could be a promising alternative to single high doses of adjuvants for transcutaneous vaccination.
Other approaches
Topically applied cytokines can act as potent adjuvants, because they are important mediators of cutaneous immune responses. Granulocyte macrophage colony-stimulating factor (GM-CSF), chemokine (C-C motif) ligand 20 (CCL-20), and fms-related tyrosine kinase 3 ligand (Flt3L) are known to be potent adjuvants in TCI (6,75). Surfactants, such as sodium lauryl sulfate, are known to enhance the antibody response to a topically applied antigen (76). Moreover, combinations of common chemicals (77) can enhance the delivery and immunogenicity of topically applied antigens.
Skin disruption is highly immunostimulatory, and can enhance antigen-specific immune responses through the secretion of proinflammatory cytokines and facilitate the migration of LCs. Several methods of skin barrier disruption, such as ultrasound, micro-needle techniques, electroporation, and gene gun delivery, have the potential to induce immune responses against topically applied vaccines (6). Hyperthermia activates the immune system, and can induce migration and maturation of LCs by increasing the expression of CD80 and CD86 (37). Mice transdermally immunized with diphtheria toxoid using local hyperthermia as an adjuvant without any adjuvant or penetration enhancing reagent resulted in an antibody response to the toxin. Heat-sensitive liposomes that quickly release their contents under hyperthermic conditions (40 to 41℃, the phase transition temperature of the liposomes) were also developed. The concept of using heat as a trigger to turn the immune system on or off, as well as to enhance gene expression, seems to be an attractive technique without risk of needle-stick injuries.
CONCLUDING REMARKS
Since the initial development of transcutaneous vaccination using bacterial exotoxin as an adjuvant by Glenn in 1998, numerous methods of efficient antigen delivery and vaccination have been developed. Although topical and transcutaneous administration of antigens has been limited, mainly due to the relatively low permeability of the stratum corneum, continued advances in the development of transcutaneous delivery systems and numerous technical innovations have led to the development of techniques and instruments that can overcome the difficulties associated with TCI. However, transcutaneous vaccination remains challenging and commercialization of transcutaneous vaccination products remains a future prospect. The main challenges are to ensure accurate antigen delivery across the epidermis, where LCs reside, and to enhance vaccine efficacy by effective activation of defined types of specific immune cells. The use of vaccines targeted to specific subsets of skin LCs is a promising way to improve vaccine efficacy by coupling the antigen to specific cell receptors on LCs. In addition, most TCI studies are currently performed in laboratory animal models. The physiological and structural differences of the skin and immune system between humans and laboratory animals should be taken into consideration to facilitate translation of the results of animal models to clinical studies. Finally, safer and more efficient adjuvants for TCI should be identified for the development of painless and self-administrable transcutaneous vaccination.
Acknowledgments
This work was supported in part by the sabbatical grant from Soonchunhyang University.
References
- 1.Combadiere B., Liard C. Transcutaneous and intradermal vaccination. Hum. Vaccine. (2011);7:811–827. doi: 10.4161/hv.7.8.16274. [DOI] [PubMed] [Google Scholar]
- 2.Glenn G. M., Scharton-Kersten T., Vassell R., Mallett C.P., Hale T. L., Alving C. R. Transcutaneous immunization with cholera toxin protects mice against lethal mucosal toxin challenge. J. Immunol. (1998);161:3211–3214. [PubMed] [Google Scholar]
- 3.Huang Y., Park Y. S., Moon C., David A. E., Chung H. S., Yang V. C. Synthetic skin-permeable proteins enabling needleless immunization. Angew. Chem. Int. Ed. Engl. (2010);49:2724–2727. doi: 10.1002/anie.200906153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bal S. M., Ding Z., van Riet E., Jiskoot W., Bouwstra J. A. Advances in transcutaneous vaccine delivery: do all ways lead to Rome? J. Control Release. (2010);148:266–282. doi: 10.1016/j.jconrel.2010.09.018. [DOI] [PubMed] [Google Scholar]
- 5.Kirschner N., Brandner J. M. Barriers and more: functions of tight junction proteins in the skin. Ann. N. Y. Acad. Sci. (2012);1257:158–166. doi: 10.1111/j.1749-6632.2012.06554.x. [DOI] [PubMed] [Google Scholar]
- 6.Karande P., Mitragotri S. Transcutaneous immunization: an overview of advantages, disease targets, vaccines, and delivery technologies. Annu. Rev. Chem. Biomol. Eng. (2010);1:175–201. doi: 10.1146/annurev-chembioeng-073009-100948. [DOI] [PubMed] [Google Scholar]
- 7.Clark R. A. Skin-resident T cells: the ups and downs of on site immunity. J. Invest. Dermatol. (2010);130:362–370. doi: 10.1038/jid.2009.247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Glenn G. M., Taylor D. N., Li X., Frankel S., Montemarano A., Alving C. R. Transcutaneous immunization: a human vaccine delivery strategy using a patch. Nat. Med. (2000);6:1403–1406. doi: 10.1038/82225. [DOI] [PubMed] [Google Scholar]
- 9.Valladeau J., Ravel O., Dezutter-Dambuyant C., Moore K., Kleijmeer M., Liu Y., Duvert-Frances V., Vincent C., Schmitt D., Davoust J., Caux C, Lebecque S., Saeland S. Langerin, a novel C-type lectin specific to Langerhans cells, is an endocytic receptor that induces the formation of Birbeck granules. Immunity. (2000);12:71–81. doi: 10.1016/S1074-7613(00)80160-0. [DOI] [PubMed] [Google Scholar]
- 10.Ochoa M. T., Loncaric A., Krutzik S. R., Becker T. C., Modlin R. L. "Dermal dendritic cells" comprise two distinct populations: CD1+ dendritic cells and CD209+ macrophages. J. Invest. Dermatol. (2008);128:2225–2231. doi: 10.1038/jid.2008.56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Toebak M. J., Gibbs S., Bruynzeel D. P., Scheper R. J., Rustemeyer T. Dendritic cells: biology of the skin. Contact Dermatitis. (2009);60:2–20. doi: 10.1111/j.1600-0536.2008.01443.x. [DOI] [PubMed] [Google Scholar]
- 12.Romani N., Clausen B. E., Stoitzner P. Langerhans cells and more: langerin-expressing dendritic cell subsets in the skin. Immunol. Rev. (2010);234:120–141. doi: 10.1111/j.0105-2896.2009.00886.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zaba L. C., Fuentes-Duculan J., Steinman R. M., Krueger J. G., Lowes M. A. Normal human dermis contains distinct populations of CD11c+BDCA-1+ dendritic cells and CD163+FXIIIA+ macrophages. J. Clin. Invest. (2007);117:2517–2525. doi: 10.1172/JCI32282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nestle F. O., Zheng X. G., Thompson C. B., Turka L. A., Nickoloff B. J. Characterization of dermal dendritic cells obtained from normal human skin reveals phenotypic and functionally distinctive subsets. J. Immunol. (1993);151:6535–6545. [PubMed] [Google Scholar]
- 15.Klechevsky E., Liu M., Morita R., Banchereau R., Thompson-Snipes L., Palucka A. K., Ueno H., Banchereau J. Understanding human myeloid dendritic cell subsets for the rational design of novel vaccines. Hum. Immunol. (2009);70:281–288. doi: 10.1016/j.humimm.2009.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bedoui S., Whitney P. G., Waithman J., Eidsmo L., Wakim L., Caminschi I., Allan R. S., Wojtasiak M., Shortman K., Carbone F. R., Brooks A. G., Heath W. R. Cross-presentation of viral and self antigens by skin-derived CD103+ dendritic cells. Nat. Immunol. (2009);10:488–495. doi: 10.1038/ni.1724. [DOI] [PubMed] [Google Scholar]
- 17.Klechevsky E., Morita R., Liu M., Cao Y., Coquery S., Thompson-Snipes L., Briere F., Chaussabel D., Zurawski G., Palucka A. K., Reiter Y., Banchereau J., Ueno H. Functional specializations of human epidermal Langerhans cells and CD14+ dermal dendritic cells. Immunity. (2008);29:497–510. doi: 10.1016/j.immuni.2008.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Caux C., Massacrier C., Vanbervliet B., Dubois B., Durand I., Cella M., Lanzavecchia A., Banchereau J. CD34+ hematopoietic progenitors from human cord blood differentiate along two independent dendritic cell pathways in response to granulocyte-macrophage colony-stimulating factor plus tumor necrosis factor alpha: II. Functional analysis. Blood. (1997);90:1458–1470. [PubMed] [Google Scholar]
- 19.Dubois B., Massacrier C., Vanbervliet B., Fayette J., Brière F., Banchereau J., Caux C. Critical role of IL-12 in dendritic cell-induced differentiation of naive B lymphocytes. J. Immunol. (1998);161:2223–2231. [PubMed] [Google Scholar]
- 20.Sixt M., Kanazawa N., Selg M., Samson T., Roos G., Reinhardt D. P., Pabst R., Lutz M. B., Sorokin L. The conduit system transports soluble antigens from the afferent lymph to resident dendritic cells in the T cell area of the lymph node. Immunity. (2005);22:19–29. doi: 10.1016/j.immuni.2004.11.013. [DOI] [PubMed] [Google Scholar]
- 21.Li N., Peng L. H., Chen X., Nakagawa S., Gao J. Q. Transcutaneous vaccines: novel advances in technology and delivery for overcoming the barriers. Vaccine. (2011);29:6179–6190. doi: 10.1016/j.vaccine.2011.06.086. [DOI] [PubMed] [Google Scholar]
- 22.Bos J. D., Meinardi M. M. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp. Dermatol. (2000);9:165–169. doi: 10.1034/j.1600-0625.2000.009003165.x. [DOI] [PubMed] [Google Scholar]
- 23.Brown M. B., Traynor M. J., Martin G. P., Akomeah F. K. Transdermal drug delivery systems: skin perturbation devices. Methods Mol. Biol. (2008);437:119–139. doi: 10.1007/978-1-59745-210-6_5. [DOI] [PubMed] [Google Scholar]
- 24.Sieg A., Wascotte V. Diagnostic and therapeutic applications of iontophoresis. J. Drug Target. (2009);17:690–700. doi: 10.3109/10611860903089750. [DOI] [PubMed] [Google Scholar]
- 25.Batheja P., Thakur R., Michniak B. Transdermal iontophoresis. Expert Opin. Drug Deliv. (2006);3:127–138. doi: 10.1517/17425247.3.1.127. [DOI] [PubMed] [Google Scholar]
- 26.Charoo N. A., Rahman Z., Repka M. A., Murthy S. N. Electroporation: an avenue for transdermal drug delivery. Curr. Drug Deliv. (2010);7:125–136. doi: 10.2174/156720110791011765. [DOI] [PubMed] [Google Scholar]
- 27.Chen X., Shah D., Kositratna G., Manstein D., Anderson R. R., Wu M. X. Facilitation of transcutaneous drug delivery and vaccine immunization by a safe laser technology. J. Control. Release. (2012);159:43–51. doi: 10.1016/j.jconrel.2012.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Vrdoljak A., McGrath M. G., Carey J. B., Draper S. J., Hill A. V., O'Mahony C., Crean A. M., Moore A. C. Coated microneedle arrays for transcutaneous delivery of live virus vaccines. J. Control. Release. (2012);159:34–42. doi: 10.1016/j.jconrel.2011.12.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Polat B. E., Hart D., Langer R., Blankschtein D. Ultrasound-mediated transdermal drug delivery: mechanisms, scope, and emerging trends. J. Control. Release. (2011);152:330–348. doi: 10.1016/j.jconrel.2011.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Peachman K. K., Rao M., Alving C. R., Burge R., Leppla S. H., Rao V. B., Matyas G. R. Correlation between lethal toxin-neutralizing antibody titers and protection from intranasal challenge with Bacillus anthracis Ames strain spores in mice after transcutaneous immunization with recombinant anthrax protective antigen. Infect. Immun. (2006);74:794–797. doi: 10.1128/IAI.74.1.794-797.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Watabe S., Xin K. Q., Ihata A., Liu L. J., Honsho A., Aoki I., Hamajima K., Wahren B., Okuda K. Protection against influenza virus challenge by topical application of influenza DNA vaccine. Vaccine. (2001);19:4434–4444. doi: 10.1016/S0264-410X(01)00194-3. [DOI] [PubMed] [Google Scholar]
- 32.Wang J., Hu J. H., Li F. Q., Liu G. Z., Zhu Q. G., Liu J. Y., Ma H. J., Peng C., Si F. G. Strong cellular and humoral immune responses induced by transcutaneous immunization with HBsAg DNA-cationic deformable liposome complex. Exp. Dermatol. (2007);16:724–729. doi: 10.1111/j.1600-0625.2007.00584.x. [DOI] [PubMed] [Google Scholar]
- 33.Partidos C. D., Beignon A. S., Brown F., Kramer E., Briand J. P., Muller S. Applying peptide antigens onto bare skin: induction of humoral and cellular immune responses and potential for vaccination. J. Control. Release. (2002);85:27–34. doi: 10.1016/S0168-3659(02)00268-7. [DOI] [PubMed] [Google Scholar]
- 34.Beignon A. S., Briand J. P., Muller S., Partidos C. D. Immunization onto bare skin with synthetic peptides:immunomodulation with a CpG-containing oligodeoxynucleotide and effective priming of influenza virus-specific CD4+ T cells. Immunology. (2002);105:204–212. doi: 10.1046/j.0019-2805.2001.01357.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Naohiro S., Yoshiki T., Takafumi N., Hideo H., Fukumi F., Masahiro T. Percutaneous peptide immunization via corneum barrier-disrupted murine skin for experimental tumor immunoprophylaxis. Proc. Natl. Acad. Sci. U.S.A. (2000);97:371–376. doi: 10.1073/pnas.97.1.371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hirobe S., Matsuo K., Quan Y. S., Kamiyama F., Morito H, Asada H., Takaya Y., Mukai Y., Okada N., Nakagawa S. Clinical study of transcutaneous vaccination using a hydrogel patch for tetanus and diphtheria. Vaccine. (2012);30:1847–1854. doi: 10.1016/j.vaccine.2011.12.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Joshi N., Duhan V., Lingwal N., Bhaskar S., Upadhyay P. Adjuvant properties of thermal component of hyperthermia enhanced transdermal immunization: effect on dendritic cells. PLoS One. (2012);7:e32067. doi: 10.1371/journal.pone.0032067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Brooks N. A., Pouniotis D. S., Tang C. K., Apostolopoulos V., Pietersz G. A. Cell-penetrating peptides: application in vaccine delivery. Biochim. Biophys. Acta. (2010);1805:25–34. doi: 10.1016/j.bbcan.2009.09.004. [DOI] [PubMed] [Google Scholar]
- 39.Tarique A. A., Kalsy A., Arifuzzaman M., Rollins S. M., Charles R. C., Leung D. T., Harris J. B., Larocque R. C., Sheikh A., Bhuiyan M. S., Saksena R., Clements J. D., Calderwood S. B., Qadri F., Kovác P., Ryan E. T. Transcutaneous immunization with a Vibrio cholerae O1 Ogawa synthetic hexasaccharide conjugate following oral whole-cell cholera vaccination boosts vibriocidal responses and induces protective immunity in mice. Clin. Vaccine Immunol. (2012);19:594–602. doi: 10.1128/CVI.05689-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mawas F., Peyre M., Beignon A. S., Frost L., Del Giudice G., Rappuoli R., Muller S., Sesardic D., Partidos C. D. Successful induction of protective antibody responses against Haemophilus influenzae type b and diphtheria after transcutaneous immunization with the glycoconjugate polyribosyl ribitol phosphate-cross-reacting material 197 vaccine. J. Infect. Dis. (2004);190:1177–1182. doi: 10.1086/423327. [DOI] [PubMed] [Google Scholar]
- 41.Slovin S. F., Keding S. J., Ragupathi G. Carbohydrate vaccines as immunotherapy for cancer. Immunol. Cell Biol. (2005);83:418–428. doi: 10.1111/j.1440-1711.2005.01350.x. [DOI] [PubMed] [Google Scholar]
- 42.Ouerfelli O., Warren J. D., Wilson R. M., Danishefsky S. J. Synthetic carbohydrate-based antitumor vaccines: challenges and opportunities. Expert Rev. Vaccines. (2005);4:677–685. doi: 10.1586/14760584.4.5.677. [DOI] [PubMed] [Google Scholar]
- 43.Skountzou I., Quan F. S., Jacob J., Compans R. W., Kang S. M. Transcutaneous immunization with inactivated influenza virus induces protective immune responses. Vaccine. (2006);24:6110–6119. doi: 10.1016/j.vaccine.2006.05.014. [DOI] [PubMed] [Google Scholar]
- 44.El-Ghorr A. A., Williams R. M., Heap C., Norval M. Transcutaneous immunisation with herpes simplex virus stimulates immunity in mice. FEMS Immunol. Med. Microbiol. (2000);29:255–261. doi: 10.1111/j.1574-695X.2000.tb01531.x. [DOI] [PubMed] [Google Scholar]
- 45.Zhang J., Shi Z., Kong F. K, Jex E., Huang Z., Watt J. M., Van Kampen K. R., Tang D. C. Topical application of Escherichia coli-vectored vaccine as a simple method for eliciting protective immunity. Infect. Immun. (2006);74:3607–3617. doi: 10.1128/IAI.01836-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mishra D., Mishra P. K., Dabadghao S., Dubey V., Nahar M., Jain N. K. Comparative evaluation of hepatitis B surface antigen-loaded elastic liposomes and ethosomes for human dendritic cell uptake and immune response. Nanomedicine. (2010);6:110–118. doi: 10.1016/j.nano.2009.04.003. [DOI] [PubMed] [Google Scholar]
- 47.Benson H. A. Transfersomes for transdermal drug delivery. Expert Opin. Drug Deliv. (2006);3:727–737. doi: 10.1517/17425247.3.6.727. [DOI] [PubMed] [Google Scholar]
- 48.Cui Z., Sloat B. R. Topical immunization onto mouse skin using a microemulsion incorporated with an anthrax protective antigen protein-encoding plasmid. Int. J. Pharm. (2006);317:187–191. doi: 10.1016/j.ijpharm.2006.04.013. [DOI] [PubMed] [Google Scholar]
- 49.Vyas S. P., Singh R. P., Jain S., Mishra V., Mahor S., Singh P., Gupta P. N., Rawat A., Dubey P. Non-ionic surfactant based vesicles (niosomes) for non-invasive topical genetic immunization against hepatitis B. Int. J. Pharm. (2005);296:80–86. doi: 10.1016/j.ijpharm.2005.02.016. [DOI] [PubMed] [Google Scholar]
- 50.Boyer C., Zasadzinski J. A. Multiple lipid compartments slow vesicle contents release in lipases and serum. ACS Nano. (2007);1:176–182. doi: 10.1021/nn7002025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kim Y. C., Ludovice P. J., Prausnitz M. R. Transdermal delivery enhanced by magainin pore-forming peptide. J. Control. Release. (2007);122:375–383. doi: 10.1016/j.jconrel.2007.05.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Himes R., Lee S., McMenigall K., Russell-Jones G. The influence of molecular adjuvants in the cutaneous response to antigen after topical vaccination. Vaccine. (2011);29:5393–5398. doi: 10.1016/j.vaccine.2011.05.085. [DOI] [PubMed] [Google Scholar]
- 53.Tomoka T-N., Erika T., Megumi K., Koichi T. Transcutaneous immunization system using a hydrotropic formulation induces a potent antigen-specific antibody response. PLoS One. (2012);7:e47980. doi: 10.1371/journal.pone.0047980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gupta R. K., Siber G. R. Comparison of adjuvant activities of aluminium phosphate, calcium phosphate and stearyl tyrosine for tetanus toxoid. Biologicals. (1994);22:53–63. doi: 10.1006/biol.1994.1008. [DOI] [PubMed] [Google Scholar]
- 55.Beignon A. S., Briand J. P., Muller S., Partidos C. D. Immunization onto bare skin with heat-labile enterotoxin of Escherichia coli enhances immune responses to coadministered protein and peptide antigens and protects mice against lethal toxin challenge. Immunology. (2001);102:344–351. doi: 10.1046/j.1365-2567.2001.01183.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Glenn G. M., Rao M., Matyas G. R., Alving C. R. Skin immunization made possible by cholera toxin. Nature. (1998);391:851. doi: 10.1038/36014. [DOI] [PubMed] [Google Scholar]
- 57.Connell T. D. Cholera toxin, LT-I, LT-IIa and LT-IIb: the critical role of ganglioside binding in immunomodulation by type I and type II heat-labile enterotoxins. Expert Rev. Vaccines. (2007);6:821–834. doi: 10.1586/14760584.6.5.821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Scharton-Kersten T., Yu Jm, Vassell R., O'Hagan D., Alving C. R., Glenn G. M. Transcutaneous immunization with bacterial ADP-ribosylating exotoxins, subunits, and unrelated adjuvants. Infect. Immun. (2000);68:5306–5313. doi: 10.1128/IAI.68.9.5306-5313.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Olvera-Gomez I., Hamilton S. E., Xiao Z., Guimaraes C. P., Ploegh H. L., Hogquist K. A, Wang L., Jameson S. C. Cholera toxin activates nonconventional adjuvant pathways that induce protective CD8 T-cell responses after epicutaneous vaccination. Proc. Natl. Acad. Sci. U.S.A. (2012);109:2072–2077. doi: 10.1073/pnas.1105771109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wilson-Welder J. H., Torres M. P., Kipper M. J., Mallapragada S. K., Wannemuehler M. J, Narasimhan B. Vaccine adjuvants: current challenges and future approaches. J. Pharm. Sci. (2009);98:1278–1316. doi: 10.1002/jps.21523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Riminton D. S., Kandasamy R., Dravec D., Basten A., Baxter A. G. Dermal enhancement: bacterial products on intact skin induce and augment organ-specific autoimmune disease. J. Immunol. (2004);1:302–309. doi: 10.4049/jimmunol.172.1.302. [DOI] [PubMed] [Google Scholar]
- 62.Rechtsteiner G., Warger T., Osterloh P., Schild H., Radsak M. P. Cutting edge: priming of CTL by transcutaneous peptide immunization with imiquimod. J. Immunol. (2005);174:2476–2480. doi: 10.4049/jimmunol.174.5.2476. [DOI] [PubMed] [Google Scholar]
- 63.Weldon W. C., Zarnitsyn V. G., Esser E. S., Taherbhai M. T., Koutsonanos D. G, Vassilieva E. V., Skountzou I., Prausnitz M. R., Compans R. W. Effect of adjuvants on responses to skin immunization by microneedles coated with influenza subunit vaccine. PLoS One. (2012);7:e41501. doi: 10.1371/journal.pone.0041501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Scott E. A., Stano A., Gillard M., Maio-Liu A. C., Swartz M. A., Hubbell J. A. Dendritic cell activation and T cell priming with adjuvant- and antigenloaded oxidation-sensitive polymersomes. Biomaterials. (2012);33:6211–6219. doi: 10.1016/j.biomaterials.2012.04.060. [DOI] [PubMed] [Google Scholar]
- 65.Chang B. A., Cross J. L., Najar H. M., Dutz J. P. Topical resiquimod promotes priming of CTL to parenteral antigens. Vaccine. (2009);27:5791–5799. doi: 10.1016/j.vaccine.2009.07.062. [DOI] [PubMed] [Google Scholar]
- 66.Verthelyi D., Zeuner R. A. Differential signaling by CpG DNA in DCs and B cells: not just TLR9. Trends Immunol. (2003);24:519–522. doi: 10.1016/S1471-4906(03)00243-6. [DOI] [PubMed] [Google Scholar]
- 67.Kim D., Kwon H. J., Lee Y. Activation of Toll-like receptor 9 and production of epitope specific antibody by liposome-encapsulated CpG-DNA. BMB Rep. (2011);44:607–612. doi: 10.5483/BMBRep.2011.44.9.607. [DOI] [PubMed] [Google Scholar]
- 68.Kim D., Kwon S., Ahn C. S., Lee Y., Choi S. Y., Park J., Kwon H. Y., Kwon H. J. Adjuvant effect of liposome-encapsulated natural phosphodiester CpG-DNA. BMB Rep. (2011);44:758–763. doi: 10.5483/BMBRep.2011.44.11.758. [DOI] [PubMed] [Google Scholar]
- 69.Diwan M., Tafaghodi M., Samuel J. Enhancement of immune responses by co-delivery of a CpG oligodeoxynucleotide and tetanus toxoid in biodegradable nanospheres. J. Control. Release. (2002);85:247–262. doi: 10.1016/S0168-3659(02)00275-4. [DOI] [PubMed] [Google Scholar]
- 70.Slütter B., Bal S. M., Ding Z., Jiskoot W., Bouwstra J. A. Adjuvant effect of cationic liposomes and CpG depends on administration route. J. Control. Release. (2011);154:123–130. doi: 10.1016/j.jconrel.2011.02.007. [DOI] [PubMed] [Google Scholar]
- 71.Ozaki T., Yauchi M., Xin K. Q., Hirahara F., Okuda K. Cross-reactive protection against influenza A virus by a topically applied DNA vaccine encoding M gene with adjuvant. Viral Immunol. (2005);18:373–380. doi: 10.1089/vim.2005.18.373. [DOI] [PubMed] [Google Scholar]
- 72.Belyakov I. M., Hammond S. A., Ahlers J. D., Glenn G.M., Berzofsky J. A. Transcutaneous immunization induces mucosal CTLs and protective immunity by migration of primed skin dendritic cells. J. Clin. Invest. (2004);113:998–1007. doi: 10.1172/JCI20261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Inoue J., Aramaki Y. Toll-like receptor-9 expression induced by tape-stripping triggers on effective immune response with CpG-oligodeoxynucleotides. Vaccine. (2007);25:1007–1013. doi: 10.1016/j.vaccine.2006.09.075. [DOI] [PubMed] [Google Scholar]
- 74.Hickey D. K., Aldwell F. E., Tan Z. Y., Bao S., Beagley K. W. Transcutaneous immunization with novel lipid-based adjuvants induces protection against gastric Helicobacter pylori infection. Vaccine. (2009);27:6983–6990. doi: 10.1016/j.vaccine.2009.09.078. [DOI] [PubMed] [Google Scholar]
- 75.Hickey D. K., Bao S., Ikeda L. T., Carey A. J., Beagley K. W. Induction of anti-chlamydial mucosal immunity by transcutaneous immunization is enhanced by topical application of GM-CSF. Curr. Mol. Med. (2005);5:559–605. doi: 10.2174/1566524054863906. [DOI] [PubMed] [Google Scholar]
- 76.Huang C. M., Wang C. C., Kawai M., Barnes S., Elmets C. A. Surfactant sodium lauryl sulfate enhances skin vaccination: molecular characterization via a novel technique using ultrafiltration capillaries and mass spectrometric proteomics. Mol. Cell. Proteomics. (2006);5:523–532. doi: 10.1074/mcp.M500259-MCP200. [DOI] [PubMed] [Google Scholar]
- 77.Karande P., Arora A., Pham T. K., Stevens D., Wojicki A., Mitragotri S. Transcutaneous immunization using common chemicals. J. Control. Release. (2009);138:134–140. doi: 10.1016/j.jconrel.2009.05.003. [DOI] [PubMed] [Google Scholar]