

Abstract
The platelet-derived growth factor (PDGF) signaling pathway is essential for inducing a dedifferentiated state of vascular smooth muscle cells (VSMCs). Activation of PDGF inhibits smooth muscle cell (SMC)-specific gene expression and increases the rate of proliferation and migration, leading to dedifferentiation of VSMCs. Recently, microRNAs have been shown to play a critical role in the modulation of the VSMC phenotype in response to extracellular signals. However, little is known about microRNAs regulated by PDGF in VSMCs. Herein, we identify microRNA-15b (miR-15b) as a mediator of VSMC phenotype regulation upon PDGF signaling. We demonstrate that miR-15b is induced by PDGF in pulmonary artery smooth muscle cells and is critical for PDGF-mediated repression of SMC-specific genes. In addition, we show that miR-15b promotes cell proliferation. These results indicate that PDGF signaling regulates SMC-specific gene expression and cell proliferation by modulating the expression of miR-15b to induce a dedifferentiated state in the VSMCs. [BMB Reports 2013; 46(11): 550-554]
Keywords: Cell proliferation, Dedifferentiation, microRNA, PDGF, Vascular smooth muscle cell
INTRODUCTION
Vascular smooth muscle cells (VSMCs) are highly differentiated cells that form blood vessel walls and have the unique ability to switch between differentiated and dedifferentiated phenotypes in response to changes in the local environment (1). For example, in response to vascular injury, VSMCs switch from a differentiated, contractile state to a dedifferentiated, synthetic state by reducing SMC-specific genes such as the α-smooth muscle actin (ASMA), as well as promoting VSMC proliferation and migration (1,2). This phenotype switching is critical for the repair of vascular injuries. It has been demonstrated that contractile VSMCs have a high expression of SMC-specific genes and a very low rate of proliferation. In contrast, synthetic VSMCs have reduced expression of SMC-specific genes and an increased rate of proliferation and migration (3).
Growth factor signaling pathways are involved in the phenotype modulation of VSMCs (4,5). Growth factor signals, such as the transforming growth factor β (TGFβ) and PDGF, modulate cell growth and migration of VSMCs directly or indirectly by transcriptional regulation of the target gene expression. Recently, it has been proposed that growth factor signals regulate the expression of microRNA (miRNA) transcriptionally and/or post-transcriptionally, in which modulate VSMC differentiation and proliferation (3,6). It has been demonstrated that the TGFβ family of growth factors regulate generation of miRNA-21 on a post-transcriptional basis by enhancing the association of the Drosha Microprocessor complex with the primary miRNA-21 transcript via Smad proteins (7). Moreover, BMP signaling represses the expression of miRNA-302-367 gene cluster for the autoregulatory mechanism of the BMP signaling pathway (8). These changes in miRNA expression upon TGFβ and BMP signaling promote differentiation of VSMCs. On the other hand, PDGF induces transcription of miR-221, promoting VSMC proliferation (9).
miRNAs are ∼22 nucleotide small non-coding RNAs that are evolutionarily conserved in metazoans and play a critical role in development, cell differentiation, proliferation, metabolism and death (10,11). miRNAs are known to regulate expression of target genes by degradation of target mRNAs or repression of target mRNA translation (12). As aberrant miRNA expression is associated with developmental abnormalities and many human diseases, including cardiovascular disorders, study of their expression level in cells is important (3). In our previous study, we saw that downregulation of miR-15b increases expression of ASMA, which is a differentiation marker of VSMCs, suggesting the possibility of miR-15b function in the regulation of VSMC differentiation and proliferation (7). Moreover, miR-15b has been shown to be induced by cardiac injury, suggesting that miR-15b play a role in cellular dedifferentiation during the repair of cardiac injuries (14). However, it is unclear whether miR-15b expression is modulated by PDGF, which is the essential signal for VSMC dedifferentiation.
In this study, we demonstrate that PDGF induces the expression of miR-15b in human primary pulmonary artery smooth muscle cells (PASMCs). Modulation of miR-15b expression has no effect on cell migration, but inhibits SMC-specific genes and promotes cell proliferation. Therefore, miR-15b regulates the VSMC synthetic phenotype by controlling SMC-specific gene expression and cell proliferation in response to PDGF.
RESULTS AND DISCUSSION
miR-15b is essential for the PDGF-mediated repression of a SMC-specific gene
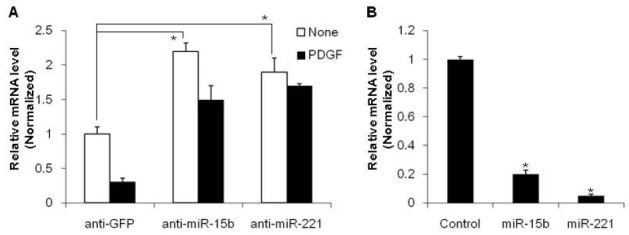
Previous study has indicated that miR-15b is involved in the regulation of SMC-specific gene expression to promote differentiation of VSMCs (7). Downregulation of miR-15b by 2’-O-methyl-modified RNA oligonucleotides complementary to the miR-15b sequence (anti-miR-15b) increased expression of ASMA in VSMCs from immnunostaining. To examine whether the effect of miR-15b on the VSMC phenotype is through transcriptional repression of the SMC-specific gene, we transfected PASMC with anti-miR-15b to inhibit miR-15b function. The level of SMC-specific gene, ASMA, was then examined by quantitative reverse transcriptase-PCR (qRT-PCR). RNA oligonucleotides against GFP (anti-GFP) were used as a control. When endogenous miR-15b is downregulated by anti-miR-15b, ASMA mRNA expression is elevated approximately 2-fold compared to the anti-GFP-transfected control (Fig. 1A). Conversely, exogenous miR-15b reduced the expression of ASMA to about 20% compared to miRNA control mimic-transfected cells (Fig. 1B). These results suggest that miR-15b represses SMC-specific gene expression. Since reduced expression of the SMC-specific gene is a typical characteristic of synthetic VSMC and this phenotype is induced by PDGF, we determined whether miR-15b is involved in the regulation of the PDGF-mediated synthetic phenotype of VSMCs. Transfection of anti-miR-15b inhibits the effects of PDGF (Fig. 1A), indicating that miR-15b plays an essential role in PDGF-mediated downregulation of the SMC specific gene, leading to the synthetic phenotype of VSMCs.
Fig. 1. miR-15b inhibits contractile gene expression. (A) PASMCs were transfected with antisense oligonucleotides to miR-15b, miR-221, or GFP as the control. Cells were then treated with PDGF-BB (20 mg/ml) for 24 h. The relative level of expression of the SMC marker, ASMA, normalized to GAPDH was determined by qRT-PCR and was presented as a mean plus SEM. *P < 0.01. (B) PASMCs were transfected with 5 nM miRNA control mimic, miR-15b mimic or miR-221 mimic for 24 h. The relative level of expression of ASMA normalized to GAPDH was examined by qRT-PCR and the results were presented as a mean plus SEM. *P < 0.001.
PDGF-BB induces miR-15b
The regulation of ASMA by miR-15b is consistent with that by miR-221 (Fig. 1), which is induced in response to PDGF (9). Induction of miR-221 by PDGF modulates VSMC proliferation and SMC gene expression. To determine whether miR-15b expression is activated by PDGF in a similar way to miR-221, a time-course expression of miR-15b and miR-221 was examined after PDGF-BB treatment in PASMCs (Fig. 2A). miR-15b is induced 2-fold within 4 h of treatment with PDGF and the induced level of miR-15b is maintained for 24 h after PDGF stimulation, suggesting that PDGF signaling modulates the expression level of miR-15b, as well as miR-221.
Fig. 2. miR-15b is regulated by the PDGF signaling pathway. (A) Time-course expression of miR-15b and miR-221 normalized to U6 small nuclear RNA was examined by qRT-PCR in PASMC treated with PDGF-BB (20 ng/ml) for the time indicated. (B) Time-course analysis of the relative expression of miR-15b or miR-21 normalized to U6 small nuclear RNA in PASMC stimulated with BMP4 for the time indicated. The average of 3 experiments, each performed in triplicate, is presented with S.E.
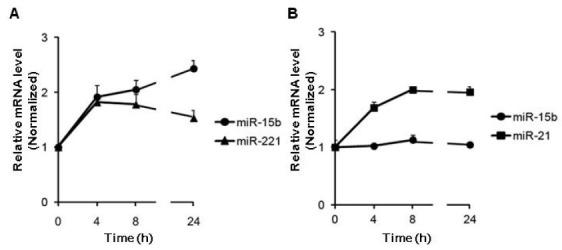
PDGF and BMP signaling play contrary roles as the switch to change VSMC phenotype. Indeed, miR-24 is induced by PDGF, in which abolishes BMP-mediated phenotypes of VSMC, such as induction of miR-21 and cell growth inhibition. Therefore, we examined whether BMP signal also modulates the level of miR-15b. After BMP4 was used to stimulate the PASMCs, we examined the time-course of miR-15b and miR-21 expression (Fig. 2B). The level of miR-15b is not altered by BMP4, while miR-21 expression is elevated by BMP4 as previously described (15), suggesting that PDGF signaling modulates the expression level of miR-15b, specifically.
miR-15b is essential for the cell proliferation
PDGF has been shown to promote the synthetic phenotype of VSMCs by regulating cell migration and growth, as well as reducing SMC-specific genes (9). To regulate cell migration and growth, PDGF modulates gene expression through miRNA, miR-221 (9). However, it is unclear whether the modulation of miR-15b generation by PDGF affects migration or growth of VSMCs. First, we examined whether endogenous miR-15b plays a role in cell migration regulation by an in vitro scratch wound assay. PASMCs were transfected with anti-miR-15b, anti-221, or anti-GFP as a control then scratch wounds were created 24 h after transfection. The wound closure was monitored in a time course (Fig. 3A). When miR-221 function was blocked by anti-miR-221, the rate of cell migration was inhibited, confirming that miR-221 plays a role in cell migration. Transfection of anti-miR-15b, however, did not alter the rate of cell migration as compared with control (anti-GFP). It is of note that endogenous miR-15b is not required to regulate cell migration unlike miR-221.
Fig. 3. miR-15b promotes VSMC proliferation. (A) PASMCs transfected with anti-GFP, anti-miR-15b, or anti-miR-221 were subjected to the scratch wound assay. Results show the mean plus S.E. of triplicate measurements from 3 independent experiments. (B) PASMCs were treated with PDGF-BB, or transfected with the control mimic (control) or miR-15b mimic for 72 hr. The number of cells was counted and presented as a mean plus SEM. *P < 0.01.
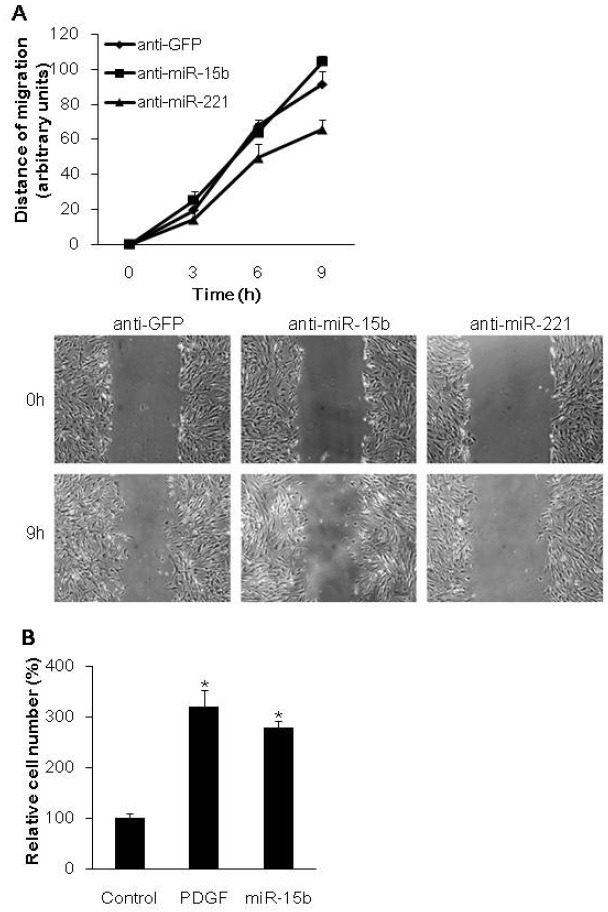
Since miR-221 induced by PDGF promotes cell growth as well as cell migration of PASMCs, we examined whether induction of miR-15b by PDGF affects cell growth. PASMCs were transfected with a control mimic or miR-15b, or treated with PDGF-BB, and the proliferation of PASMC was monitored by cell number counts after 48 h (Fig. 3B). PDGF elevated the rate of cell proliferation about 3-fold compared to the control. Similar to stimulation of PDGF signaling, exogenous miR-15b mimic enhances cell growth, suggesting that miR-15b is essential for cell proliferation. Together these results demonstrate that miR-15b facilitates PDGF-mediated prosynthetic activity by promoting VSMC proliferation.
Target prediction of miR-15b
Recent studies have revealed that miRNAs are important mediators for the modulation of the VSMC phenotype by repressing their targets, which function as molecular switches for VSMC differentiation (3,6). While it is known that a single miRNA affects the expression of a large number of protein coding genes (∼300) to regulate multiple cellular events simultaneously (16,17), only 5 targets have been identified and validated for miR-15b (Table 1) (18-22). Most studies of miR-15b and targets have been performed using cancer cells. In tongue squamous cell carcinoma (TSCC), miR-15b is essential for the cancer cell epithelial-mesenchymal transition through targeting BMI1 (18). In various cancer cells, miR-15b promotes malignant cell behavior by targeting a tumor suppressor, RECK (20). Furthermore, miR-15b regulates cell cycle progression in glioma cells by targeting cell cycle-related molecules (21) and also modulates apoptosis via targeting BCL2 in gastric cancer cells (22). All these studies indicate that miR-15b plays a critical role in the regulation of cell proliferation by targeting the regulatory molecules of cell growth or cell death. Indeed, major cell cycle regulators, such as cyclin D and cyclin E, are predicted for miR-15b targets by computational algorithms for miRNA target prediction, such as TargetScan and PicTar (Table 1).
Table 1.
Targets of miR-15b
| Gene | Accession # | Known | Reference |
|---|---|---|---|
|
| |||
| BCL2 | NM_000633 | Validated | Xia, et al. (21) |
| RECK | NM_021111 | Validated | Loayza-Puch, et al. (19) |
| CCNE1 | NM_057182 | Validated | Xia, et al. (20) |
| MAP2K4 | NM_003010.2 | Validated | Marasa, et al. (18) |
| BMI1 | NM_005180.8 | Validated | Sun, et al. (17) |
| Cyclin D1 | NM_053056 | Predicted | TargetScan, PicTar |
| Cyclin D2 | NM_001759 | Predicted | TargetScan, PicTar |
| Cyclin D3 | NM_001136017 | Predicted | TargetScan |
| Cyclin E1 | NM_001238 | Predicted | TargetScan, PicTar |
| CHK1 | NM_001114122 | Predicted | TargetScan, PicTar |
| CDK6 | NM_001145306 | Predicted | TargetScan |
| CDC14 | NM_001077181 | Predicted | TargetScan |
| CDC23 | NM_004661 | Predicted | TargetScan |
| CDC25A | NM_001789 | Predicted | TargetScan |
| CDC25B | NM_004358 | Predicted | TargetScan |
| CDC27 | NM_001114091 | Predicted | TargetScan |
| E2F3 | NM_001949 | Predicted | TargetScan, PicTar |
| Transcription factor Dp-2 | NM_001178138 | Predicted | TargetScan |
| WEE1 | NM_001143976 | Predicted | TargetScan, PicTar |
Genes involved in cell proliferation, growth, and death, were validated as miR-15b targets (shown at the top of the table). Additional key regulators of the cell cycle are predicted to be miR-15b targets by TargetScan and/or PicTar.
It has been shown that PDGF induces miR-221 levels and the repression of the miR-221 target, p27Kip1, is critical for PDGF-mediated induction of cell proliferation, leading to the dedifferentiated synthetic phenotype of VSMCs (9). Since miR-15b expression is induced by PDGF and miR-15b is sufficient for cell proliferation, it is likely that induction of miR-15b by PDGF is essential for PDGF-medicated cell proliferation of VSMC through targeting predicted molecules that regulate cell growth or cell death (Table 1). Further validation of predicted targets of miR-15b and the unraveling of the mechanisms responsible for regulation of miRNA-15b by PDGF will be important to elucidate the pathophysiological significance of miR-15b in VSMC phenotype regulation.
MATERIALS AND METHODS
Cell culture
Human primary pulmonary artery smooth muscle cells (PASMCs) were purchased from Lonza (CC-2581) and were maintained in Sm-GM2 medium (Lonza) containing 5% fetal bovine serum (FBS). Recombinant human platelet-derived growth factor (PDGF-BB) and BMP4 were purchased from R&D Systems. The cells were treated with PDGF-BB (20 ng/ml) or 3nM BMP4 under starvation conditions (0.2% FBS), as described previously (4).
Quantitative reverse transcriptase-PCR (qRT-PCR)
Quantitative analysis of the change in expression levels was calculated by a real-time PCR machine (iQ5; Bio-Rad), as described previously (23). mRNA levels were normalized to GAPDH. The sequences of the real-time PCR primers used for human smooth muscle α-actin (ASMA) were 5’-TTGCAGGAA CCTCTTCGGCC-3’ and 5’-GGTCGCTTCCTTATTCCTGC-3’. For detection of mature miRNAs, a TaqMan MicroRNA assay kit (Applied Biosystems) was used according to the manufacturer's instructions. Data analysis was performed using the comparative CT method in Bio-Rad software. The miRNA levels were normalized to the U6 small nuclear RNA. The average of 3 experiments, each performed in triplicate, is presented with standard errors.
miRNA mimic
Chemically modified double-stranded RNAs designed to mimic the endogenous mature miR-15b, miR-221, and negative control miRNA were purchased from Ambion. miRNA mimics were transfected using RNAi Max (Invitrogen) according to the manufacturer's directions at 5 nM.
Anti-miRNA oligonucleotides
2’-O-methyl-modified RNA oligonucleotides complementary to miRNA or green fluorescent protein (GFP; control) sequence were purchased from IDT and transfected at a concentration of 40 nM using RNAi Max (Invitrogen) according to the manufacturer's directions. Sequences are: anti-miR-221, 5’-G AAACCCAGCAGACAAUGUAGCU-3’; anti-miR-15b, 5’-UGU AAACCAUGAUGUGCUGCUA-3’; anti-GFP, 5’-AAGGCAAGCUGACCCUGAAGU-3’.
In vitro scratch wound assay
PASMCs transfected with anti-GFP, anti-miR-15b, or antimiR221 were plated in 6-well plates, and 3 scratch wounds were generated with a 200 μl pipette tip. Scratch wounds were photographed over 9 h with a Zeiss inverted microscope and their widths were quantitated with the Image J software.
Counting cell number
PASMCs were plated in 12-well plates 24 h after miRNA mimic transfection or PDGF-BB stimulation. After 48 h, cells were trypsinized and the total number of cells was counted using an hemocytometer.
Acknowledgments
This work was supported by the Incheon National University Research Grant in 2012 to H. K.
References
- 1.Owens G. K., Kumar M. S., Wamhoff B. R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. (2004);84:767–801. doi: 10.1152/physrev.00041.2003. [DOI] [PubMed] [Google Scholar]
- 2.Owens G. K. Regulation of differentiation of vascular smooth muscle cells. Physiol. Rev. (1995);75:487–517. doi: 10.1152/physrev.1995.75.3.487. [DOI] [PubMed] [Google Scholar]
- 3.Kang H., Hata A. MicroRNA regulation of smooth muscle gene expression and phenotype. Curr. Opin. Hematol. (2012);19:224–231. doi: 10.1097/MOH.0b013e3283523e57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lagna G., Ku M. M., Nguyen P. H., Neuman N. A., Davis B. N., Hata A. Control of phenotypic plasticity of smooth muscle cells by bone morphogenetic protein signaling through the myocardin-related transcription factors. J. Biol. Chem. (2007);282:37244–37255. doi: 10.1074/jbc.M708137200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.ten Dijke P., Arthur H. M. Extracellular control of TGFbeta signalling in vascular development and disease. Nat. Rev. Mol. Cell. Biol. (2007);8:857–869. doi: 10.1038/nrm2262. [DOI] [PubMed] [Google Scholar]
- 6.Hata A. Functions of microRNAs in cardiovascular biology and disease. Annu. Rev. Physiol. (2013);75:69–93. doi: 10.1146/annurev-physiol-030212-183737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Davis B. N., Hilyard A. C., Lagna G., Hata A. SMAD proteins control DROSHA-mediated microRNA maturation. Nature. (2008);454:56–61. doi: 10.1038/nature07086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kang H., Louie J., Weisman A., Sheu-Gruttadauria J., Davis-Dusenbery B. N., Lagna G., Hata A. Inhibition of MicroRNA-302 (miR-302) by bone morphogenetic protein 4 (BMP4) facilitates the BMP signaling pathway. J. Biol. Chem. (2012);287:38656–38664. doi: 10.1074/jbc.M112.390898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Davis B. N., Hilyard A. C., Nguyen P. H., Lagna G., Hata A. Induction of microRNA-221 by platelet-derived growth factor signaling is critical for modulation of vascular smooth muscle phenotype. J. Biol. Chem. (2009);284:3728–3738. doi: 10.1074/jbc.M808788200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bartel D. P. MicroRNAs: target recognition and regulatory functions. Cell. (2009);136:215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kim V. N., Han J., Siomi M. C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. (2009);10:126–139. doi: 10.1038/nrm2632. [DOI] [PubMed] [Google Scholar]
- 12.Jost D., Nowojewski A., Levine E. Small RNA biology is systems biology. BMB Rep. (2011);44:11–21. doi: 10.5483/BMBRep.2011.44.1.11. [DOI] [PubMed] [Google Scholar]
- 13.Davis-Dusenbery B. N., Hata A. Mechanisms of control of microRNA biogenesis. J. Biochem. (2010);148:381–392. doi: 10.1093/jb/mvq096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liu L. F., Liang Z., Lv Z. R., Liu X. H., Bai J., Chen J., Chen C., Wang Y. MicroRNA-15a/b are up-regulated in response to myocardial ischemia/reperfusion injury. J. Geriatr. Cardiol.: JGC. (2012);9:28–32. doi: 10.3724/SP.J.1263.2012.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kang H., Davis-Dusenbery B. N., Nguyen P. H., Lal A., Lieberman J., Van Aelst L., Lagna G., Hata A. Bone morphogenetic protein 4 promotes vascular smooth muscle contractility by activating microRNA-21 (miR-21), which down-regulates expression of family of dedicator of cytokinesis (DOCK) proteins. J. Biol. Chem. (2012);287:3976–3986. doi: 10.1074/jbc.M111.303156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Selbach M., Schwanhausser B., Thierfelder N., Fang Z., Khanin R., Rajewsky N. Widespread changes in protein synthesis induced by microRNAs. Nature. (2008);455:58–63. doi: 10.1038/nature07228. [DOI] [PubMed] [Google Scholar]
- 17.Baek D., Villen J., Shin C., Camargo F. D., Gygi S. P., Bartel D. P. The impact of microRNAs on protein output. Nature. (2008);455:64–71. doi: 10.1038/nature07242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sun L., Yao Y., Liu B., Lin Z., Lin L., Yang M., Zhang W., Chen W., Pan C., Liu Q., Song E., Li J. MiR-200b and miR-15b regulate chemotherapy-induced epithelial-mesenchymal transition in human tongue cancer cells by targeting BMI1. Oncogene. (2012);31:432–445. doi: 10.1038/onc.2011.263. [DOI] [PubMed] [Google Scholar]
- 19.Marasa B. S., Srikantan S., Masuda K., Abdelmohsen K., Kuwano Y., Yang X., Martindale J. L., Rinker-Schaeffer C. W., Gorospe M. Increased MKK4 abundance with replicative senescence is linked to the joint reduction of multiple microRNAs. Sci. Signal. (2009);2:ra69. doi: 10.1126/scisignal.2000442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Loayza-Puch F., Yoshida Y., Matsuzaki T., Takahashi C., Kitayama H., Noda M. Hypoxia and RAS-signaling pathways converge on, and cooperatively downregulate, the RECK tumor-suppressor protein through microRNAs. Oncogene. (2010);29:2638–2648. doi: 10.1038/onc.2010.23. [DOI] [PubMed] [Google Scholar]
- 21.Xia H., Qi Y., Ng S. S., Chen X., Chen S., Fang M., Li D., Zhao Y., Ge R., Li G., Chen Y., He M. L., Kung H. F., Lai L., Lin M. C. MicroRNA-15b regulates cell cycle progression by targeting cyclins in glioma cells. Biochem. Biophs. Res. Co. (2009);380:205–210. doi: 10.1016/j.bbrc.2008.12.169. [DOI] [PubMed] [Google Scholar]
- 22.Xia L., Zhang D., Du R., Pan Y., Zhao L., Sun S., Hong L., Liu J., Fan D. miR-15b and miR-16 modulate multidrug resistance by targeting BCL2 in human gastric cancer cells. International journal of cancer. J. Int. Canc. (2008);123:372–379. doi: 10.1002/ijc.23501. [DOI] [PubMed] [Google Scholar]
- 23.Chan M. C., Hilyard A. C., Wu C., Davis B. N., Hill N. S., Lal A., Lieberman J., Lagna G., Hata A. Molecular basis for antagonism between PDGF and the TGFbeta family of signalling pathways by control of miR-24 expression. EMBO J. (2010);29:559–573. doi: 10.1038/emboj.2009.370. [DOI] [PMC free article] [PubMed] [Google Scholar]