

Abstract
Dopamine (DA) regulates emotional and motivational behavior through the mesolimbic dopaminergic pathway. Changes in DA signaling in mesolimbic neurotransmission are widely believed to modify reward-related behaviors and are therefore closely associated with drug addiction. Recent evidence now suggests that as with drug addiction, obesity with compulsive eating behaviors involves reward circuitry of the brain, particularly the circuitry involving dopaminergic neural substrates. Increasing amounts of data from human imaging studies, together with genetic analysis, have demonstrated that obese people and drug addicts tend to show altered expression of DA D2 receptors in specific brain areas, and that similar brain areas are activated by food-related and drug-related cues. This review focuses on the functions of the DA system, with specific focus on the physiological interpretation and the role of DA D2 receptor signaling in food addiction. [BMB Reports 2013; 46(11): 519-526]
Keywords: Addiction, Dopamine, Dopamine receptor, Food reward, Reward circuit
INTRODUCTION
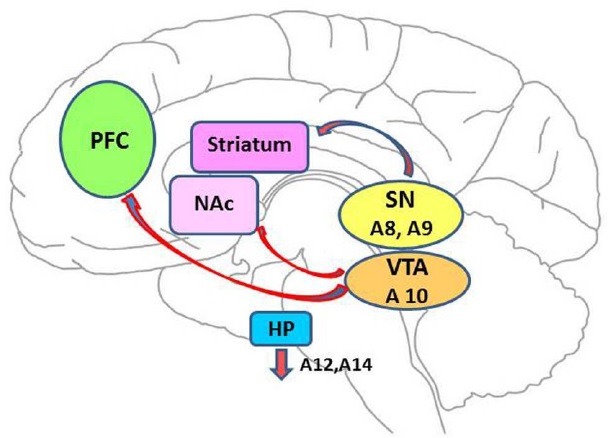
Catecholamines have often been linked to the behavioral pathology of a number of neurological and psychiatric disorders such as Parkinson's disease, Huntington's disease, drug addiction, depression, and schizophrenia. Dopamine (DA) is the predominant catecholamine in the brain and is synthesized by mesencephalic neurons in the substantia nigra (SN) and the ventral tegmental area (VTA). DA neurons project from the SN and VTA to many different areas of the brain. These dopaminergic cell groups are designated as group ‘A’ cells, indicating aminergic DA-containing cells, and are subdivided into cell groups A8 through A14. DA cells within the pars compacta (A8) and neighboring areas (groupA9) of the SN project to the basal ganglia (striatum, globus pallidus, and subthalamic nucleus). This projection constitutes the nigrostriatal pathway, which is involved primarily in the control of voluntary movement but also in goal-directed behaviors (Fig. 1). From the VTA, A10 cell group projects to the nucleus accumbens (NAc), prefrontal cortex, and other limbic areas. Thus, this group of cells is termed the mesolimbic and mesocortical pathways (Fig. 1). These neurons play a crucial role in reward-related behaviors and motivation. Another distinct group of cells constitutes the tubero-infundibular pathway. These cells arise from the arcuate nucleus (cellgroupA12) and periventricular nucleus (cellgroupA14) of the hypothalamus and project to the pituitary. This pathway is known to control the release and synthesis of pituitary hormone, primarily prolactin (1-4).
Fig. 1. DAergic pathways in brain. Major three dopaminergic pathways are presented: First, nigrostriatal pathway where DA cells within pars compacta (A8) and neighbouring area (group A9) from SN project to striatum, this projection is involved in mostly the control of vouluntary movement. Secondly, mesolimbic mesocortical pathway, which is the projection from VTA, cell group A10, to the nucleus accumbens (NAc), the preforntal cortex (PFC) and other limbic areas. These neurons play a crucial role in reward-related behaviors. Thirdly, the tubero-infundibular pathways, which are the cells fromarcuate nucleus (cell group A12) and periventricular nucleus (cell group A14) of the hypothalamus, projecting to the pituituary. This pathaway is known to control the release and synthesis of pituituary hormone, mostly prolactin. VTA, ventral tegmental area; SN, substanita nigra; NAc, nucleus accumbens; PFC, Prefrontal cortex; HP, hypothalamus.
Regulation of the DA system for reward-related behaviors is mediated by the mesolimbic and mesocortical pathways. The role of DA in reward-related behaviorshas received much attention because of severe consequences of dysfunction within the mesolimbic and mesocortical circuits, which include drug addiction and depression. It has recently become accepted that DA-mediated food reward is linked to obesity, a major public health problem.
It is well known that a homeostatic regulation center for feeding behaviors exists in the brain, in particular the hypothalamus, and serves to integrate different hormonal and neuronal signals that control appetite and energy homeostasis in controlling body weight. This homeostatic regulation of body weight monitors the level of body adiposity by employing different regulators such as leptin, insulin, and ghrelin (5). However, the motivation for food is strongly associated with reward, and responding to the hedonic properties of food such as its sight, smell, and taste may be associated with conditioning cues. These hedonic qualities can override the homeostatic system (6). Therefore, delineating how this food reward circuit in the brain can control appetite and eating behaviors in connection with the brain’s homeostatic system of energy balance is difficult.
Considerable evidence suggests that synaptic modifications of the mesolimbic DA system are critically associated with the rewarding effects of drugs of abuse as well as with food reward (7-9). However, DA reward signaling is far more complex than it appears, and it is also implicated in learning and conditioning processes, as evidenced by studies revealing that dopaminergic reward signals are involved in coding for reward prediction error in behavioral learning (10-13). In drug addiction, it is well known that the rewarding effects of drugs are primarily induced by increased DA release upon targeting of a specific substrate, such as the DA transporter in the case of cocaine. In food addiction, however, it remains to be elucidated how food reward can activate the DA reward signal in a manner similar to that evoked by drug addiction. It is important to understand the mechanisms by which these reward components induce adaptive changes in DA circuitry responsible for these addictive behaviors (7-9).
In this review, I will provide a short summary of dopaminergic signaling in food reward-related behaviors, with a focus on recent studies on the role of DA receptor subtypes, in particular D2 receptors, in this process.
DA D2 RECEPTORS
DA interacts with membrane receptors belonging to a family of seven transmembrane domain G-protein-coupled receptors. This leads to the formation of second messengers and the activation or repression of specific signaling pathways. To date, five different subtypes of DA receptor have been cloned from different species. A general subdivision into two groups has been made based on their structural and G-protein coupling properties: the D1-like receptors, which stimulate intracellular cAMP levels and comprising D1 (14,15) and D5 (16,17) receptors, and the D2-like receptors, which inhibit intracellular cAMP levels and comprise the D2 (18,19), D3 (20), and D4 (21) receptors.
D1 and D2 receptors are the most abundant DA receptors in the brain. The expression of D3, D4, and D5 receptors in the brain is considerably more restricted and weaker than that of D1 and D2 receptors. The D2 receptor is represented by two isoforms generated by alternative splicing of the same gene (18,22). These isoforms, namely D2L and D2S, are identical except for an insert of 29 amino acids present in the putative third intracellular loop of D2L, which is in fact encoded by exon 6 of the D2 receptor gene, an intracellular domain thought to have a role in coupling this class of receptor to particular second messengers. The large isoform appears to be the predominant form present in all brain regions, although the exact ratio of the two isoforms can vary (22). In fact, the phenotype of D2 receptor total knockout mice was revealed as being quite different from the D2L knockout mice (23-25), indicating that these two isoforms of D2 receptor might have different functions in vivo. Recent results from Moyer and coworkers support a differential in vivo function of the two D2 receptor isoforms in the human brain. They demonstratedthat the two variants of the D2 receptor gene (Drd2), caused by D2 receptor alternative splicing, possessed intronic single-nucleotide polymorphisms (SNPs) that were differentially associated with cocaine abuse in Caucasians (26,27). D2S and D2L mRNA levels were measured in tissues from human brain autopsies (prefrontal cortex and putamen) obtained from cocaine abusers and controls, and the relationship between the D2 receptor gene genotype, D2S/L splicing, and cocaine abuse was examined. The results supported a robust effect of difference of specific SNPs in decreasing the relative expression of D2S in humans, representing strong risk factors in cases of cocaine overdose (26). Given that these two isoforms are generated by alternative splicing of a single gene, it would also be interesting to see whether the ratio of the two isoforms could be a factor contributing to such disease.
D2 receptors are also localized presynaptically, as indicated by experiments examining receptor expression and binding sites in DA neurons throughout the midbrain (28). These D2 autoreceptors may either be somatodendritic autoreceptors, which are known to decrease neuronal excitability (29,30), or terminal autoreceptors, which mostly reduce DA synthesis and packaging (31,32) and inhibit DA release (33-35). It has been suggested that in the embryonic stage, the D2 autoreceptor may play a role in DA neuronal development (36-38).
Bello and coworkers recently generated mice conditionally deficient for the D2 receptor in midbrain DA neurons (referred to as autoDrd2KO mice). These autoDrd2 KO mice lacked DA-mediated somatodendritic synaptic responses and inhibition of DA release (39) and displayed elevated DA synthesis and release, hyperlocomotion, and supersensitivity to the psychomotor effects of cocaine. The mice also exhibited increased place preference for cocaine and enhanced motivation for food reward, indicating the importance of D2 autoreceptors in the regulation of DA neurotransmission and demonstrating that D2 autoreceptors are important for normal motor function, food-seeking behavior, and sensitivity to the locomotor and reward properties of cocaine (39). Therefore, the principal role of these autoreceptors appears to be the inhibition and modulation of DA neurotransmission. As demonstrated with D2 autoreceptor deficient mice, one can therefore hypothesize that modulating the level of sensitivity to the reward response via the presynaptic D2 receptor might be crucial in motivational behavioral responses to addictive drugs as well as food rewards, although the cellular and molecular role of these presynaptic D2 receptors remains to be further investigated.
DOPAMINE SIGNALING IN FOOD REWARD
As mentioned above, drugs of abuse can alter our brain reward systems, in particular the dopaminergic mesolimbic system. In addition, it has been demonstrated that palatable food with high fat and sugar content can significantly activate DA reward circuitry. These findings suggest common neural substrates exist for both food and drug addictions, and that both depend on dopaminergic circuits. Furthermore, human brain imaging studies strongly support a role for dopaminergic circuits in the control of food intake (40-43).
Drugs of abuse trigger large increases in synaptic DA concentrations in the mesolimbic system (44). Likewise, it has been reported that rewarding food stimulates dopaminergic transmission in the NAc (45-47). When DA was measured by microdialysis in the nucleus accumbens of freely moving rats in the presence of food rewards, it was observed that amphetamine and cocaine injection increased DA levels in the NAc, which is normally activated by eating; thus, suggesting that the release of DA by eating could be a factor in food addiction (46). In addition, using fast-scan cyclic voltammetry at carbon-fiber microelectrodes in the NAc of rats trained to press a lever for sucrose, Rotiman and coworkers have shown that cues signaling the opportunity to respond for sucrose reward, or the unexpected delivery of sucrose, evoked DA release in the NAc (47); thus, strongly implicating DA signaling in the NAc as a real-time modulator of food-seeking behavior. However, some other studies have revealed the importance of the dorsal striatum, rather than the NAc, in the control of food reward. For example, injection of the DA antagonist cis-flupenthixol into the dorsal striatum but not the NAc, amygdala, or frontal cortex of rats produces a decline in food reward-associated lever pressing (48). Additionally, DA-deficient mice are hypophagic, and virally mediated restoration of DA production in DA-deficient mice reverses aphagia only when DA signaling in the caudate-putamen and dorsal striatumhas been restored. In contrast, restoration of dopaminergic signaling to the NAc did not reverse aphagia, although the locomotor response to a novel environment or amphetamine was restored by viral delivery to the NAc (49,50).
In humans, mostly the dorsal striatum has been observed to correlate with feeding behaviors. For example, Small and coworkers used positron emission tomography (PET) on human subjectsto show that regional cerebral blood flow measured while eating chocolate correlated with pleasantness ratings in the dorsal caudate and putamen, but not in the NAc (41). In a PET imaging study of healthy human subjects, a correlation was observed between the reduction in DA ligand binding in the dorsal striatum and feeding (42). Consistent with this finding, striatal D2 receptor expression was decreased in obese individuals in proportion to their body mass index (40); this issue will be discussed further in the following section.
D2 receptors in food reward
Although feeding increases extracellular DA concentration in the nucleus accumbens in rats, (45,46), as do drugs of abuse, DA depletion in the NAc in rats following bilateral injections of the neurotoxic agent 6-hydroxydopamine (6-OHDA) into the nucleus accumbens alone does not alter feeding (51). Pharmacological blockade of D1 and D2 receptors in the NAc affects motor behavior and the frequency and duration of feeding, but does not reduce the amount of food consumed (52). Another study reported that when exposed to the same high-fat diet, mice with lower D2 receptor density in the putamen gain more weight than mice with higher D2 receptor density (53), showing that the dopaminergic system responds to palatable food. Davis and coworkers assessed the hypothesis that diet-induced obesity reduces mesolimbic DA function (54). They compared DA turnover in the mesolimbic DA system between rats fed a high-fat diet and those consuming a standard low-fat diet (54). The results demonstrated that animals consuming a high-fat diet, independent of the development of obesity, exhibited decreased DA turnover in the NAc, reduced preference for an amphetamine cue, and attenuated operant responses for sucrose. The authors also observed that obesity induced due to a high-fat diet attenuated mesolimbic DA turnover in the nucleus accumbens, while there were no differences in DA concentration or turnover in the orbitofrontal cortex, suggesting a specific effect of a high-fat diet restricted to the NAc (54).
Recently, Halpern and coworkers examined the effect of deep brain stimulation (DBS) of the NAc shell (55). Since this procedure is currently under investigation in humans for the treatment of major depression, obsessive–compulsive disorder, and addiction, they hypothesized that it may also be effective in limiting binge eating. Interestingly, DBS of the NAc shell was found to reduce binge eating and increased c-Fos levels in this region. Raclopride, a DA D2 receptor antagonist, attenuated the effects of DBS, whereas the D1 receptor antagonist SCH-23390 was ineffective, suggesting that DA signaling involving D2 receptors is required for the effect of DBS in the NAc shell (55). When they examined the effect of chronic NAc shell DBS in diet-induced obese mice, it was found to acutely reduce caloric intake and induce weight loss and, thus, supporting the involvement of D2 receptor-containing DA pathways in the food reward contributing to obesity, as well as the efficacy of NAc shell DBS in modulating this system (55).
A recent study conducted by Johnson and Kenny suggested a strong correlation between D2 receptor expression and compulsive eating behaviors (56). In this study, it was observed that in animals given a ‘cafeteria diet’, consisting of a selection of highly palatable, energy-dense food that is available at cafeterias for human consumption, these animals gained weight and demonstrated compulsive eating behavior (56). In addition to their excessive adiposity and compulsive eating, rats under cafeteria diet had decreased D2 receptor expression in the striatum. In another recent study, the selective deletion of insulin receptors in midbrain dopaminergic neurons in mice demonstrated that this manipulation results in increased body weight, increased fat mass, and hyperphagia (57). Interestingly, in these mice, DA D2 receptor expression in the VTA was decreased as compared to that in the control mice, suggesting a possible disinhibition of dopaminergic VTA/SN cells in a D2 receptor-dependent mechanism (57). However, in our laboratory, we observed that compared to wild-type (WT) mice, D2 receptor KO mice have a lean phenotype and exhibit reduced food intake and body weight with enhanced hypothalamic leptin signaling (58). Based on these findings, we cannot rule out that D2 receptor has a role in the homeostatic regulation of metabolism in association with homeostatic regulators of energy balance, such as leptin, in addition to its role in food motivation behavior. Therefore, it appears that the expression of D2 receptor is tightly associated with food reward and eating behaviors, and that depending on the localization of D2 receptors in the brain, this could lead to different outcomes in the relevant circuits.
DA D2 receptors in human obesity
Many human studies have indicated the importance of the DA D2 receptor in regulating food rewardin the context of obesity, particularly showing a change in striatal D2 receptor function and expression (59,60). Obese people and drug addicts tend to show reduced expression of DA D2 receptors in striatal areas, and imaging studies have demonstrated that similar brain areas are activated by food-related and drug-related cues (61,62). PET studies suggest that the availability of DA D2 receptors is decreased in obese individuals in proportion to their body mass index (40); thus, suggesting that DA deficiency in obese individuals may perpetuate pathological eating as a means to compensate for decreased activation of dopaminergic reward circuits. An alternative explanation is that individuals with low numbers of D2 receptors may be more vulnerable to addictive behaviors, including compulsive food intake, and, thus, providing direct evidence of a deficit in DA D2 receptors in obese individuals (40).
Based on the reduced D2 receptor availability in the striatal region of obese individuals, which suggests a possible role for D2 receptors in the inhibitory control of compulsive eating behaviors, Volkow and coworkers investigated whether D2 receptor availability in obese subjects would be associated with metabolism in prefrontal regions such as the cingulate gyrus (CG), dorsolateral prefrontal cortex (DLPFC), and orbitofrontal cortex, which are brain regions that have been implicated in various components of inhibitory control (63). Their study revealed a significant association between D2 receptor levels in the striatum and the activity in the DLPFC, medial OFC, and CG in obese subjects. Since these brain regions are implicated in inhibitory control, salience attribution, and emotional reactivity, this finding suggests that disruption of these areas can cause impulsive and compulsive behaviors, and that this may be one of the mechanisms by which the low D2 receptor levels in obesity contribute to over-eating and obesity (63).
The association between D2 receptor genotype and obesity in humans has been investigated, and it has been suggested that allelic variants of the Taq1A polymorphism in the D2 receptor gene affect D2 receptor expression (64,65). This polymorphism lies 10 kb downstream of the coding region of the gene and falls within the protein-encoding region of a neighboring gene ankyrin repeat and kinase domain containing 1 (ANKK1). The Taq1A polymorphism has three allelic variants: A1/A1, A1/A2, and A2/A2. Postmortem and PET studies suggest that individuals with one or two copies of the A1 allele have 30-40% fewer D2 receptors when compared to those without an A1 allele (64) and an association of the A1 allele with alcoholism has been suggested (64,66). Interestingly, it has been reported that food reinforcement has a significant effect on energy intake, and this effect is moderated by the A1 allele (67,68). Epstein and coworkers examined food reinforcement, polymorphisms in the dopamine D2 receptor and DA transporter genes, and laboratory energy intake in obese and non-obese humans. Food reinforcement was greater in obese than in non-obese individuals, especially in obese individuals with the TaqI A1 allele. Energy intake was greater for individuals with high levels of food reinforcement and greatest in those having high levels of food reinforcement as well as the TaqI A1 allele (68). However, no effect of the DA transporter genewas observed in this study, indicating an association between D2 receptor gene polymorphism and food reinforcement.
In accordance with this study, Stice and coworkers used functional magnetic resonance imaging (fMRI) to show that in individuals with the A1 allele of the TaqIA polymorphism in the D2 receptor gene, weaker striatal activation in response to food intake was significantly more strongly related to current body mass and future weight gain over a 1-year follow-up, compared to those lacking the A1 allele (59,69,70). Using a different fMRI experimental paradigm, Stice and coworkers demonstrated that weaker activation of the frontal operculum, lateral orbitofrontal cortex, and striatum in response to imagined eating of appetizing foods, as opposed to imagined eating of less palatable foods or drinking water, predicted elevated weight gain for those with the A1 allele (71). Weaker activation of the frontal operculum, lateral orbitofrontal cortex, and striatum in response to imagined intake of palatable foods also predicted future increases in body mass for those with the TaqIA A1 allele of the D2 receptor gene (71), suggesting that for those lacking this allele, greater responsivity of these food reward regions predicted future increases in body mass.
Interestingly, a recent report by Davis and coworkers demonstrated another aspect of the link between D2 receptor signals and compulsive eating behaviors (72). They showed that obese adults with binge eating disorder differ biologically from their counterparts who do not binge eat. In fact, obese adults with binge eating disorder were characterized by a stronger DA signal when compared to their obese but non-binging counterparts, a difference that was associated with a distinct genetic polymorphism of TaqIA of the D2 receptor gene (72).
In addition, while D2 receptor signaling in the dorsal striatum seems to be implicated in the inhibitory control of compulsive eating behaviors, Caravaggio and coworkers recently reported a positive correlation between body mass and D2/D3 receptor agonist binding in the ventral striatum (NAc) of non-obese humans, but found no relationship with antagonist binding. These data suggest that in non-obese individuals, higher body mass may be associated with increased D2 receptor affinity in the NAc, and that this increased affinity may potentiate the incentive salience of food cues and may increase motivation to consume palatable foods (73).
Therefore, even though considerable evidence indicates that low D2 receptor levels are associated with increases in food intake, weight gain, and risk for food addiction, as observed in humans with substance abuse problems (74), it would be valuable to determine how D2 receptor expression and its downstream signaling may control this association.
CONCLUSIONS AND FUTURE DIRECTIONS
Increasing evidence has been made to delineate the brain circuit controlling the homeostatic regulation of food intake. Recent findings have helped demonstrate the remarkable interaction between the homeostatic and reward circuits of feeding behaviors. Human studies strikingly demonstrate the importance of reward systems, in particular the DA system, in controlling eating behavior and obesity. Based on known genetic susceptibilities and regulation of the D2 receptor in food reward studies, it is clear that D2 receptor function is critical for food motivation and brain signaling in obesity. However, it remains difficult to define a framework of the involved brain circuits that includes the molecular substrates relevant for controlling food addiction. Recent studies from our laboratory demonstrated that the D2 receptor is not required for the acquisition of drug addiction, but it plays a key role in regulating synaptic modifications triggered by experiences such as stress. Therefore, the D2 receptor functions rather as a mediator of experience-induced, drug-seeking, and relapse behaviors (75), indicating its specific role in addictive behaviors.
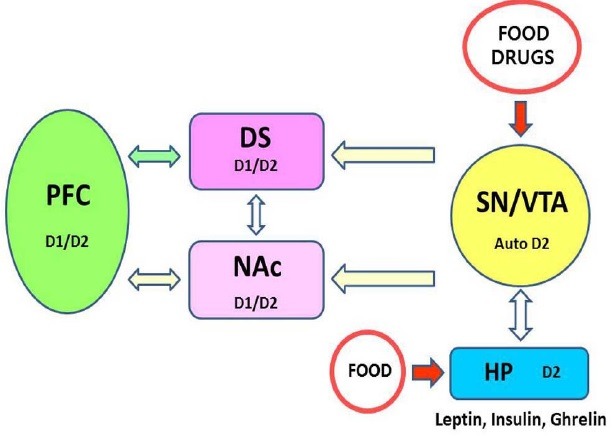
As for drug addiction, it appears that food stimuli activate the VTA-NAc dopaminergic mesolimbic circuit, with the phenotypic importance of feeding behaviors translated through signaling in the caudate putamen and dorsal striatum, which interact with the prefrontal cortex for decision making and executing eating behaviors. The aforementioned homeostatic regulators, such as leptin, insulin, and ghrelin, exert their impact on the midbrain DA systemby regulating the connection between the homeostatic and hedonic systems of food intake, (6,9,76) (Fig. 2). There is no doubt that these lines of investigation have provided a foundation for future studies on the neural circuitry of the DA system, which will assist in the elucidation of the underlying pathophysiology of food addiction. Recent breakthroughs in tools such as optogenetics and DREADDs (designer receptors exclusively activated by designer drugs) will facilitate these studies through allowing access to specific neuronal cells or circuits that control specific reward-related behaviors.
Fig. 2. Food reward circuit involving DA system and D2 receptors. As the drug addiction, it appears that food stimuli activate VTA-NAc DA mesolimbic circuit with phenotypic importance of feeding behaviors translated through signaling in caudate putamen, dorsal striatum (DS), interacting with prefrontal cortex (PFC) for decision making and execution of eating behaviors. As well, the homeostatic regulators such as leptin, insulin and ghrelin exert their input to midbrain DA system for connection between homeostatic and hedonic system of food intake. Expression of D1 and D2 receptors in each area is shown as D1, D2 or autoD2 (presynaptic D2 autoreceptor). Other mesolimbic circuit areas which control also eating behavior and food reward such as amygdala and hippocampus are not presented here. VTA, ventral tegmental area; SN, substanita nigra; NAc, nucleus accumbens; DS, dorsal striatum; PFC, Prefrontal cortex; HP, hypothalamus.
Acknowledgments
This work was supported by the grant of the Korean Health Technology R&D Project (A111776) from the Ministry of Health & Welfare, and partly by the Brain Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (2013056101), Republic of Korea.
References
- 1.Hornykiewicz O. Dopamine (3-hydroxytyramine) and brain function. Pharmacol. Rev. (1966);18:925–964. [PubMed] [Google Scholar]
- 2.Björklund A., Dunnett S. B. Dopamine neuron systems in the brain: an update. Trends Neurosci. (2007);30:194–202. doi: 10.1016/j.tins.2007.03.006. [DOI] [PubMed] [Google Scholar]
- 3.Beaulieu J. M., Gainetdinov R. R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. (2011);63:182–217. doi: 10.1124/pr.110.002642. [DOI] [PubMed] [Google Scholar]
- 4.Tritsch N. X., Sabatini B. L. Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron. (2012);76:33–50. doi: 10.1016/j.neuron.2012.09.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Morton G. J., Cummings D. E., Baskin D. G., Barsh G. S., Schwartz M. W. Central nervous system control of food intake and body weight. Nature. (2006);443:289–295. doi: 10.1038/nature05026. [DOI] [PubMed] [Google Scholar]
- 6.Palmiter R. D. Is dopamine a physiologically relevant mediator of feeding behavior? Trends Neurosci. (2007);30:375–381. doi: 10.1016/j.tins.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 7.Nestler E. J., Carlezon W. A. Jr. The mesolimbic dopamine reward circuit in depression. Biol. Psychiatry. (2006);59:1151–1159. doi: 10.1016/j.biopsych.2005.09.018. [DOI] [PubMed] [Google Scholar]
- 8.Steketee J. D., Kalivas P. W. Drug wanting: behavioral sensitization and relapse to drug-seeking behavior. Pharmacol. Rev. (2011);63:348–365. doi: 10.1124/pr.109.001933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kenny P. J. Common cellular and molecular mechanisms in obesity and drug addiction. Nat. Rev. Neurosci. (2011);12:638–651. doi: 10.1038/nrn3105. [DOI] [PubMed] [Google Scholar]
- 10.Schultz W. Predictive reward signal of dopamine neurons. J. Neurophysiol. (1998);80:1–27. doi: 10.1152/jn.1998.80.1.1. [DOI] [PubMed] [Google Scholar]
- 11.Schultz W. Behavioral dopamine signals. Trends Neurosci. (2007);30:203–210. doi: 10.1016/j.tins.2007.03.007. [DOI] [PubMed] [Google Scholar]
- 12.Schultz W. Updating dopamine reward signals. Curr. Opin. Neurobiol. (2012);23:229–238. doi: 10.1016/j.conb.2012.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wise R. A. Dopamine, learning and motivation. Nat. Rev. Neurosci. (2004);5:483–494. doi: 10.1038/nrn1406. [DOI] [PubMed] [Google Scholar]
- 14.Dearry A., Gingrich J. A., Falardeau P., Fremeau R. T., Jr., Bates M. D., Caron M. G. Molecular cloning and expression of the gene for a human D1 dopamine receptor. Nature. (1990);347:72–76. doi: 10.1038/347072a0. [DOI] [PubMed] [Google Scholar]
- 15.Zhou Q. Y., Grandy D. K., Thambi L., Kushner J. A., Van Tol H. H., Cone R., Pribnow D., Salon J., Bunzow J. R., Civelli O. Cloning and expression of human and rat D1 dopamine receptors. Nature. (1990);347:76–80. doi: 10.1038/347076a0. [DOI] [PubMed] [Google Scholar]
- 16.Grandy D. K., Zhang Y. A., Bouvier C., Zhou Q. Y., Johnson R. A., Allen L., Buck K., Bunzow J. R., Salon J., Civelli O. Multiple human D5 dopamine receptor genes: a functional receptor and two pseudogenes. Proc. Natl. Acad. Sci. U. S. A. (1991);88:9175–9179. doi: 10.1073/pnas.88.20.9175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sunahara R. K., Guan H. C., O’Dowd B. F., Seeman P., Laurier L. G., Ng G., George S. R., Torchia J., Van Tol H. H., Niznik H. B. Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than D1. Nature. (1991);350:614–619. doi: 10.1038/350614a0. [DOI] [PubMed] [Google Scholar]
- 18.Bunzow J. R., Van Tol H. H., Grandy D. K., Albert P., Salon J., Christie M., Machida C. A., Neve K. A., Civelli O. Cloning and expression of a rat D2 dopamine receptor cDNA. Nature. (1988);336:783–787. doi: 10.1038/336783a0. [DOI] [PubMed] [Google Scholar]
- 19.Dal Toso R., Sommer B., Ewert M., Herb A., Pritchett D. B., Bach A., Shivers B. D., Seeburg P. H. The dopamine D2 receptor: two molecular forms generated by alternative splicing. EMBO J. (1989);8:4025–4034. doi: 10.1002/j.1460-2075.1989.tb08585.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sokoloff P., Giros B., Martres M. P., Bouthenet M. L., Schwartz J. C. Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature. (1990);347:146–151. doi: 10.1038/347146a0. [DOI] [PubMed] [Google Scholar]
- 21.Van Tol H. H., Bunzow J. R., Guan H. C., Sunahara R. K., Seeman P., Niznik H. B., Civelli O. Cloning of the gene for a human dopamine D4 receptor with high affinity for the antipsychotic clozapine. Nature. (1991);350:610–614. doi: 10.1038/350610a0. [DOI] [PubMed] [Google Scholar]
- 22.Montmayeur J. P., Bausero P., Amlaiky N., Maroteaux L., Hen R., Borrelli E. Differential expression of the mouse D2 dopamine receptor isoforms. FEBS Lett. (1991);278:239–243. doi: 10.1016/0014-5793(91)80125-M. [DOI] [PubMed] [Google Scholar]
- 23.Baik J. H., Picetti R., Saiardi A., Thiriet G., Dierich A., Depaulis A., LeMeur M., Borrelli E. Parkinsonian-like locomotor impairment in mice lacking dopamine D2 receptors. Nature. (1995);377:424–428. doi: 10.1038/377424a0. [DOI] [PubMed] [Google Scholar]
- 24.Usiello A., Baik J. H., Rouge-Pont F., Picetti R., Dierich A., LeMeur M., Piazza P. V., Borrelli E. Distinct functions of the two isoforms of dopamine D2 receptors. Nature. (2000);408:199–202. doi: 10.1038/35041572. [DOI] [PubMed] [Google Scholar]
- 25.Wang Y., Xu R., Sasaoka T., Tonegawa S., Kung M. P., Sankoorikal E. B. Dopamine D2 long receptor-deficient mice display alterations in striatum-dependent functions. J. Neurosci. (2000);20:8305–8314. doi: 10.1523/JNEUROSCI.20-22-08305.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Moyer R. A., Wang D., Papp A. C., Smith R. M., Duque L., Mash D. C., Sadee W. Intronic polymorphisms affecting alternative splicing of human dopamine D2 receptor are associated with cocaine abuse. Neuropsychopharmacology. (2011);36:753–762. doi: 10.1038/npp.2010.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gorwood P., Le Strat Y., Ramoz N., Dubertret C., Moalic J. M., Simonneau M. Genetics of dopamine receptors and drug addiction. Hum Genet. (2012);131:803–822. doi: 10.1007/s00439-012-1145-7. [DOI] [PubMed] [Google Scholar]
- 28.Sesack S. R., Aoki C., Pickel V. M. Ultrastructural localization of D2 receptor-like immunoreactivity in midbrain dopamine neurons and their striatal targets. J. Neurosci. (1994);14:88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chiodo L. A., Kapatos G. Membrane properties of identified mesencephalic dopamine neurons in primary dissociated cell culture. Synapse. (1992);11:294–309. doi: 10.1002/syn.890110405. [DOI] [PubMed] [Google Scholar]
- 30.Lacey M. G., Mercuri N. B., North R. A. Dopamine acts on D2 receptors to increase potassium conductance in neurones of the rat substantia nigra zona compacta. J. Physiol (Lond). (1987);392:397–416. doi: 10.1113/jphysiol.1987.sp016787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Onali P., Oliansa M. C., Bunse B. Evidence that adenosine A2 and dopamine autoreceptors antagonistically regulate tyrosine hydroxylase activity in rat striatal synaptosomes. Brain. Res. (1988);456:302–309. doi: 10.1016/0006-8993(88)90232-6. [DOI] [PubMed] [Google Scholar]
- 32.Pothos E. N, Davila V., Sulzer D. Presynaptic recording of quanta from midbrain dopamine neurons and modulation of the quantal size. J. Neurosci. (1998);18:4106–4118. doi: 10.1523/JNEUROSCI.18-11-04106.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cass W. A., Zahniser N. R. Potassium channel blockers inhibit D2 dopamine, but not A1 adenosine, receptor-mediated inhibition of striatal dopamine release. J. Neurochem. (1991);57:147–152. doi: 10.1111/j.1471-4159.1991.tb02109.x. [DOI] [PubMed] [Google Scholar]
- 34.Kennedy R. T., Jones S. R., Wightman R. M. Dynamic observation of dopamine autoreceptor effects in rat striatal slices. J. Neurochem. (1992);59:449–455. doi: 10.1111/j.1471-4159.1992.tb09391.x. [DOI] [PubMed] [Google Scholar]
- 35.Congar P., Bergevin A., Trudeau L. E. D2receptors inhibit the secretory process downstream from calcium influx in dopaminergic neurons: implication of K+ channels. J. Neurophysiol. (2002);87:1046–1056. doi: 10.1152/jn.00459.2001. [DOI] [PubMed] [Google Scholar]
- 36.Kim S. Y., Choi K. C., Chang M. S., Kim M. H., Kim S. Y., Na Y. S., Lee J. E., Jin B. K., Lee B. H., Baik J. H. The dopamine D2 receptor regulates the development of dopaminergic neurons via extracellular signal- regulated kinase and Nurr1 activation. J. Neurosci. (2006);26:4567–4576. doi: 10.1523/JNEUROSCI.5236-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yoon S., Choi M. H., Chang M. S., Baik J. H. Wnt5a-dopamine D2 receptor interactions regulate dopamine neuron development via extracellular signal-regulated kinase (ERK) activation. J. Biol. Chem. (2011);286:15641–15651. doi: 10.1074/jbc.M110.188078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yoon S., Baik J. H. Dopamine D2 receptormediated epidermal growth factor receptor transactivation through a disintegrin and metalloprotease regulates dopaminergic neuron development via extracellular signal-related kinase activation. J. Biol. Chem. (2013);288:28435–28446. doi: 10.1074/jbc.M113.461202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bello E. P., Mateo Y., Gelman D. M., Noain D., Shin J. H., Low M. J., Alvarez V. A., Lovinger D. M., Rubinstein M. Cocaine supersensitivity and enhanced motivation for reward in mice lacking dopamine D(2) autoreceptors. Nat. Neurosci. (2011);14:1033–1038. doi: 10.1038/nn.2862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang G. J., Volkow N. D., Logan J., Pappas N. R., Wong C. T., Zhu W., Netusil N., Fowler J. S. Brain dopamine and obesity. Lancet. (2001);357:354–357. doi: 10.1016/S0140-6736(00)03643-6. [DOI] [PubMed] [Google Scholar]
- 41.Small D. M., Zatorre R. J., Dagher A., Evans A. C., Jones-Gotman M. Changes in brain activity related to eating chocolate: from pleasure to aversion. Brain. (2001);124:1720–1733. doi: 10.1093/brain/124.9.1720. [DOI] [PubMed] [Google Scholar]
- 42.Small D. M., Jones-Gotman M., Dagher A. Feeding-induced dopamine release in dorsal striatum correlates with meal pleasantness ratings in healthy human volunteers. Neuroimage. (2003);19:1709–1715. doi: 10.1016/S1053-8119(03)00253-2. [DOI] [PubMed] [Google Scholar]
- 43.Volkow N. D., Wang G. J., Baler R. D. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn. Sci. (2011);15:37–46. doi: 10.1016/j.tics.2010.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Di Chiara G., Imperato A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc. Natl. Acad. Sci. U. S. A. (1988);85:5274–5278. doi: 10.1073/pnas.85.14.5274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bassareo V., Di Chiara G. Differential influence of associative and nonassociative learning mechanisms on the responsiveness of prefrontal and accumbal dopamine transmission to food stimuli in rats fed ad libitum. J. Neurosci. (1997);17:851–861. doi: 10.1523/JNEUROSCI.17-02-00851.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hernandez L., Hoebel B. G. Food reward and cocaine increase extracellular dopamine in the nucleus accumbens as measured by microdialysis. Life Sci. (1988);42:1705–1712. doi: 10.1016/0024-3205(88)90036-7. [DOI] [PubMed] [Google Scholar]
- 47.Roitman M. F., Stuber G. D., Phillips P. E., Wightman R. M., Carelli R. M. Dopamine operates as a subsecond modulator of food seeking. J. Neurosci. (2004);24:1265–1271. doi: 10.1523/JNEUROSCI.3823-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Beninger R. J., Ranaldi R. Microinjections of flupenthixol into the caudate-putamen but not the nucleus accumbens, amygdala or frontal cortex of rats produce intra- session declines in food-rewarded operant responding. Behav. Brain Res. (1993);55:203–212. doi: 10.1016/0166-4328(93)90116-8. [DOI] [PubMed] [Google Scholar]
- 49.Szczypka M. S., Kwok K., Brot M. D., Marck B. T., Matsumoto A. M., Donahue B. A., Palmiter R. D. Dopamine production in the caudate putamen restores feeding in dopamine-deficient mice. Neuron. (2001);30:819–828. doi: 10.1016/S0896-6273(01)00319-1. [DOI] [PubMed] [Google Scholar]
- 50.Hnasko T. S., Perez F. A., Scouras A. D., Stoll E. A., Gale S. D., Luquet S., Phillips P. E., Kremer E. J., Palmiter R. D. Cre recombinase-mediated restoration of nigrostriatal dopamine in dopamine-deficient mice reverses hypophagia and bradykinesia. Proc. Natl. Acad. Sci. U. S. A. (2006);103:8858–8863. doi: 10.1073/pnas.0603081103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Salamone J. D., Mahan K., Rogers S. Ventrolateral striatal dopamine depletions impair feeding and food handling in rats. Pharmacol. Biochem. Behav. (1993);44:605–610. doi: 10.1016/0091-3057(93)90174-R. [DOI] [PubMed] [Google Scholar]
- 52.Baldo B. A., Sadeghian K., Basso A. M., Kelley A. E. Effects of selective dopamine D1 or D2 receptor blockade within nucleus accumbens subregions on ingestive behavior and associated motor activity. Behav. Brain Res. (2002);137:165–177. doi: 10.1016/S0166-4328(02)00293-0. [DOI] [PubMed] [Google Scholar]
- 53.Huang X. F., Zavitsanou K., Huang X., Yu Y., Wang H., Chen F., Lawrence A. J., Deng C. Dopamine transporter and D2 receptor binding densities in mice prone or resistant to chronic high fat diet-incuded obesity. Behav Brain Res. (2006);175:415–419. doi: 10.1016/j.bbr.2006.08.034. [DOI] [PubMed] [Google Scholar]
- 54.Davis J. F., Tracy A. L., Schurdak J. D., Tschop M. H., Lipton J. W., Clegg D. J., Benoit S. C. Exposure to elevated levels of dietary fat attenuates psychostimulant reward and mesolimbic dopamine turnover in the rat. Behav Neurosci. (2008);122:1257–1263. doi: 10.1037/a0013111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Halpern C. H., Tekriwal A., Santollo J., Keating J. G., Wolf J. A., Daniels D., Bale T. L. Amelioration of binge eating by nucleus accumbens shell deep brain stimulation in mice involves D2 receptor modulation. J. Neurosci. (2013);33:7122–7129. doi: 10.1523/JNEUROSCI.3237-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Johnson P. M., Kenney P. J. Dopamine D2 receptors in addictionlike reward dysfunction and compulsive eating in obese rats. Nat. Neurosci. (2010);13:635–641. doi: 10.1038/nn.2519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Könner A. C., Hess S., Tovar S., Mesaros A., Sánchez-Lasheras C., Evers N., Verhagen L. A., Brönneke H. S., Kleinridders A., Hampel B., Kloppenburg P., Brüning J. C. Role for insulin signaling in catecholaminergic neurons in control of energy homeostasis. Cell Metab. (2011);13:720–728. doi: 10.1016/j.cmet.2011.03.021. [DOI] [PubMed] [Google Scholar]
- 58.Kim K. S., Yoon Y. R., Lee H. J., Yoon S., Kim S. Y., Shin S. W., An J. J., Kim M. S., Choi S. Y., Sun W., Baik J. H. Enhanced hypothalamic leptin signaling in mice lacking dopamine D2 receptors. J. Biol. Chem. (2010);285:8905–8917. doi: 10.1074/jbc.M109.079590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Stice E., Yokum S., Zald D., Dagher A. Dopamine-based reward circuitry responsivity, genetics, and overeating. Curr. Top Behav. Neurosci. (2011);6:81–93. doi: 10.1007/7854_2010_89. [DOI] [PubMed] [Google Scholar]
- 60.Salamone J. D., Correa M. Dopamine and food addiction: lexicon badly needed. Biol. Psychiatry. (2013);73:e15–24. doi: 10.1016/j.biopsych.2012.09.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wang G. J., Volkow N. D., Thanos P. K., Fowler J. S. Imaging of brain dopamine pathways: Implications for understanding obesity. J. Addict Med. (2009);3:8–18. doi: 10.1097/ADM.0b013e31819a86f7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Volkow N. D., Fowler J. S., Wang G. J., Baler R., Telang F. Imaging dopamine’s role in drug abuse and addiction. Neuropharmacology. (2009);56:3–8. doi: 10.1016/j.neuropharm.2008.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Volkow N. D., Wang G. J., Telang F., Fowler J. S., Thanos P. K., Logan J., Alexoff D., Ding Y. S., Wong C., Ma Y., Pradhan K. Low dopamine striatal D2 receptors are associated with prefrontal metabolism in obese subjects: possible contributing factors. Neuroimage. (2008);42:1537–1543. doi: 10.1016/j.neuroimage.2008.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ritchie T., Noble E. P. Association of seven polymorphisms of the D2 dopamine receptor gene with brain receptor-binding characteristics. Neurochem. Res. (2003);28:73–82. doi: 10.1023/A:1021648128758. [DOI] [PubMed] [Google Scholar]
- 65.Fossella J., Green A. E., Fan J. Evaluation of a structural polymorphism in the ankyrin repeat and kinase domain containing 1 (ANKK1) gene and the activation of executive attention networks. Cogn. Affect. Behav. Neurosci. (2006);6:71–78. doi: 10.3758/CABN.6.1.71. [DOI] [PubMed] [Google Scholar]
- 66.Noble E. P. D2 dopamine receptor gene in psychiatric and neurologic disorders and its phenotypes. Am. J. Med. Genet. B. Neuropsychiatr. Genet. (2003);116B:103–125. doi: 10.1002/ajmg.b.10005. [DOI] [PubMed] [Google Scholar]
- 67.Epstein L. H., Wright S. M., Paluch R. A., Leddy J. J., Hawk L. W., Jaroni J. L., Saad F. G., Crystal-Mansour S., Shields P. G., Lerman C. Relation between food reinforcement and dopamine genotypes and its effect on food intake in smokers. Am. J. Clin. Nutr. (2004);80:82–88. doi: 10.1093/ajcn/80.1.82. [DOI] [PubMed] [Google Scholar]
- 68.Epstein L. H., Temple J. L., Neaderhiser B. J., Salis R. J., Erbe R. W., Leddy J. J. Food reinforcement, the dopamine D2 receptor genotype, and energy intake in obese and nonobese humans. Behav. Neurosci. (2007);121:877–886. doi: 10.1037/0735-7044.121.5.877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Stice E., Spoor S, Bohon C., Small D. M. Relation between obesity and blunted striatal response to food is moderated by TaqIA A1 allele. Science. (2008);322:449–452. doi: 10.1126/science.1161550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Stice E., Spoor S., Bohon C., Veldhuizen M., Small D. M. Relation of reward from food intake and anticipated intake to obesity: a functional magnretic resonance imaging study. J. Abnorm Psychol. (2008);117:924–935. doi: 10.1037/a0013600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Stice E., Yokum S., Bohon C., Marti N., Smolen A. Reward circuitry responsivity to food predicts future increases in body mass: moderating effects of DRD2 and DRD4. Neuroimage. (2010);50:1618–1625. doi: 10.1016/j.neuroimage.2010.01.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Davis C., Levitan R. D., Yilmaz Z., Kaplan A. S., Carter J. C., Kennedy J. L. Binge eating disorder and the dopamine D2 receptor: Genotypes and sub-phenotypes. Prog. Neuro-psychopharmacol. Biol. Psychiatry. (2012);38:328–335. doi: 10.1016/j.pnpbp.2012.05.002. [DOI] [PubMed] [Google Scholar]
- 73.Caravaggio F, Raitsin S, Gerretsen P, Nakajima S, Wilson A., Graff-Guerrero A. Ventral striatum binding of a dopamine D2/3 receptor agonist but not antagonist predicts normal body mass index. Biol. Psychiatry. (2013) doi: 10.1016/j.biopsych.2013.02.017. doi:pii:S0006-3223(13)00185-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Martinez D., Broft A., Foltin R. W., Slifstein M., Hwang D. R., Huang Y., Perez A., Frankle W. G., Cooper T., Kleber H. D., Fischman M. W., Laruelle M. Cocaine dependence and d2 receptor availability in the functional subdivisions of the striatum: relationship with cocaine-seeking behavior. Neuropsychopharmacology. (2004);29:1190–1202. doi: 10.1038/sj.npp.1300420. [DOI] [PubMed] [Google Scholar]
- 75.Sim H. R., Choi T. Y, Lee H. J., Kang E. Y., Yoon S., Han P. L., Choi S. Y., Baik J. H. Role of dopamine D2 receptors in plasticity of stress-induced addictive behaviours. Nat Commu. (2013);4:1579. doi: 10.1038/ncomms2598. [DOI] [PubMed] [Google Scholar]
- 76.Baik J. H. Dopamine Signaling in reward-related behaviors. Front. Neural. Circuits. (2013);7:152. doi: 10.3389/fncir.2013.00152. [DOI] [PMC free article] [PubMed] [Google Scholar]