

Abstract
In this study, we aimed to compare the morphogenetic and neuronal characteristics between monolayer cells and spheroids. For this purpose, we established spheroid formation by growing SH-SY5Y cells on the hydrophobic surfaces of thermally-collapsed elastin-like polypeptide. After 4 days of culture, the relative proliferation of the cells within spheroids was approximately 92% of the values for monolayer cultures. As measured by quantitative assays for mRNA and protein expressions, the production of synaptophysin and neuronspecific enolase (NSE) as well as the contents of cell adhesion molecules (CAMs) and extracellular matrix (ECM) proteins are much higher in spheroids than in monolayer cells. Under the all-trans-retinoic acid (RA)-induced differentiation condition, spheroids extended neurites and further up-regulated the expression of synaptophysin, NSE, CAMs, and ECM proteins. Our data indicate that RA-differentiated SH-SY5Y neurospheroids are functionally matured neuronal architectures. [BMB Reports 2013; 46(5): 276-281]
Keywords: Cell adhesion molecules, Extracellular matrix, Multicellular neurospheroids, Neuroblastoma cells, Neuronal differentiation
INTRODUCTION
Three-dimensional multicellular spheroids (MCS) have been regarded as morphologically and physiologically superior to monolayer cultures for recapitulating in vivo tissues (1). In particular, the MCS generated by human neuroblastoma cell lines are very useful tools to assess the efficacy of chemo-, immuno-, and radio-therapy in vitro (2-4). When cultured over a non-adhesive surface, SH-SY5Y cells spontaneously aggregate and ultimately form MCS (5). In such cases, N-cadherin, neural cell adhesion molecule-2 (NCAM2), intercellular cell adhesion molecule-1 (ICAM1), and connexin-26 have been known to regulate the spheroid formation and neuronal differentiation (6,7). Collagens and fibronectin were strongly produced in BT4C tumor spheroids (8). However, in case of SH-SY5Y cells, the changes in the expression levels of these proteins upon MCS organization have not been investigated.
The SH-SY5Y cell line is a subclone of SK-N-SH cells (9). They are typically locked in an undifferentiated state and can be differentiated in vitro by all-trans-retinoic acid (RA) to acquire more neuron-like characteristics (10). RA-differentiated cells can be readily judged by biochemical analysis of neuronal markers such as synaptophysin and NSE. Recently, there has been a great increase in the use of RA-differentiated SH-SY5Y cells as a model for neurodegenerative diseases such as Parkinson’s disease (11). However, the necessity of differentiating SH-SY5Y cells has been controversial since a study demonstrated no significant difference in neuronal properties between undifferentiated and RA-differentiated SK-N-SH cells (12). Moreover, the majority of researches regarding cell differentiation have been done on monolayer cultures, and therefore the morphological and neuronal characteristics of RA-differentiated SH-SY5Y spheroids remain unstudied.
To address the issues described above, we produced MCS by culturing SH-SY5Y cells on an elastin-like polypeptide, TGPG[VGRGD(VGVPG)6]20WPC (referred to as RGD-ELP), as an amphiphilic substrate (13). Initially, we examined the growth characteristics of spheroids with respect to their macroscopic morphology, viability, and proliferation. We also systematically compared the differences among monolayer cells and spheroids grown under undifferentiated and RA-differentiated conditions in terms of the expression levels of CAMs and ECM proteins, neurite extension, and the contents of neuronal markers, by quantitative RT-PCR (qRT-PCR) and Western blot immunoassay.
RESULTS
Formation of SH-SY5Y spheroids
The conventional polystyrene (PS) plate is uncharged, whereas the RED-ELP substrate presenting amphiphilic characteristics of hydrophilic RGD ligands and hydrophobic (VGVPG)6 pentapeptides has been shown to modulate cellular morphology and functions through receptor-mediated signal transduction (14). Cell growth patterns on PS plates or RGD-ELP substrates are displayed in Fig. 1A. On PS, cells spread out and grew into a net-like structure in a monolayer configuration, indicating prevailing cell-substrate adhesive interaction. In contrast, on the RGD-ELP-coated surfaces, SH-SY5Y cells formed partly compact aggregates within 24 h. After 48 h, these aggregates subsequently developed into MCS. The high density of RGD-ELP was more effective in promoting cell aggregation during the earlier stage of cell growth (> 24 h), while the size of the spheroid was not significantly affected by the surface density of RGD-ELP. The spheroids formed after 72 h displayed a broad size distribution ranging from 30-150 μm in diameter.
Fig. 1. Spheroid formation and the viability and proliferation assays for monolayer cells and spheroids. (A) Phase contrast microscopic observation of MCS formation. Wells of 6-well plates were treated with 1 ml of 0, 0.5, 1, 5, or 10 μM RGD-ELP for 1 h at 37℃. Surface density of RGD-ELP was 0, 2.9 ± 0.4, 6.1 ± 0.8, 28.8 ± 3.5, or 60.4 ± 5.7 μg/cm2, respectively. Cells (1 × 105 cells/well) were seeded and cultured for 4 days. (B) SEM images of monolayer cells (a) and MCS (b). (C) Fluorescence images of live (green) and dead (red) cell staining. Concentration of RGD-ELP was 1 μM. (D) Time dependent change in the total number of live cells counted by the Tryphan Blue exclusion method. Black and gray columns represent monolayer cultures and MCS, respectively. The scale bars in A and C are 50 μm. Data are the mean ± SEM of three independent measurements.
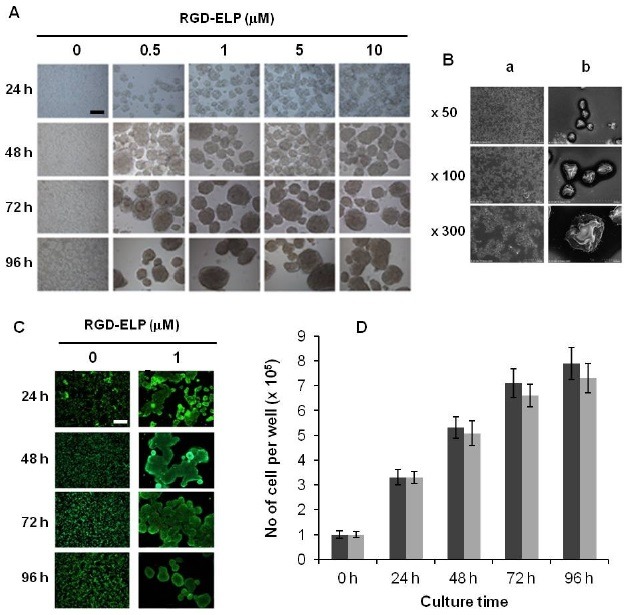
Scanning electron microscope (SEM) images confirmed that individual cells in the spheroids, generated after 48 h of over-lay culture, were in direct contact with each other to form dense aggregates (Fig. 1B).
Viability and proliferation of the cells within spheroids
With the fluorescence dye assay, almost no sign of cell death was detected during 4 days of cell growth on PS and RGD-ELP (Fig. 1C). To obtain the quantitative proliferation data, both the monolayer cells and spheroids were dissociated into a homogenous, single-cell population by trypsin-EDTA treatment. Live cell counting after 24, 48, 72, and 96 h by Tryphan blue showed an increase in the absolute numbers of cells from both monolayer culture and spheroids (Fig. 1D), indicating that SH-SY5Y cells proliferate within the spheroid structures. The relative values of cell proliferation obtained after 48, 72, and 96 h of spheroid culture were approximately 95%, 93% and 92% of the values for monolayer cells. This relatively low cell proliferation may be related to the limited oxygen/nutrient availability within the spheroidal structures.
Neurite outgrowth from RA-differentiated spheroids
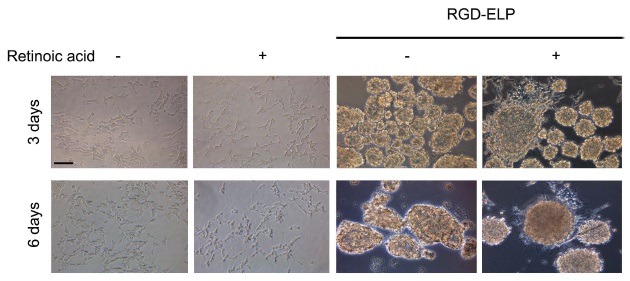
To evaluate the differentiability of monolayer cells and spheroids, we monitored neurite outgrowth under undifferentiated and RA-induced differentiation conditions (Fig. 2). In case of monolayer cells, weak and moderate neurite outgrowth occurred after 3 and 6 days of culture without RA treatment, while extensive neurite elongation from a single cell body was observed 3 days after RA treatment. For the spheroids, neurite formation was not observed in the absence of RA, whereas extensive neurite outgrowth occurred on the periphery of the spheres after 3 days of RA treatment.
Fig. 2. Neurite outgrowth from monolayer cells and spheroids cultured without (-) and with (+) 10 μM RA. After 2 days of cell growth, cells were treated with RA, and neuritogenesis was observed 3 and 6 days after RA treatment. Spheroids were formed using 1 μM RGD-ELP. Magnification is ×200. Scale bar is 50 μm.
Up-regulation of neuronal markers in RA-differentiated spheroids
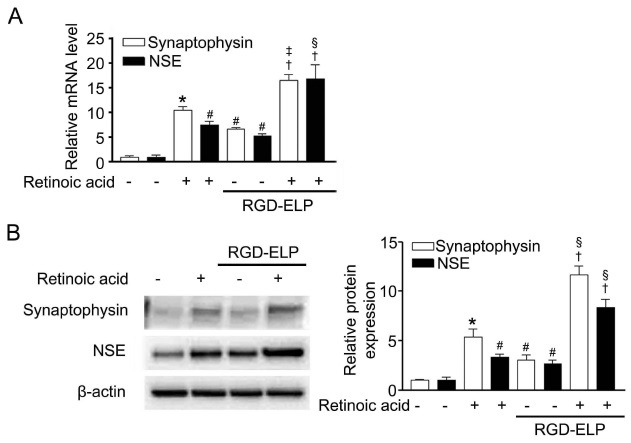
Production of neuronal markers synaptophysin and NSE were quantified at mRNA (Fig. 3A) and protein levels (Fig. 3B). A statistically (P < 0.01) significant increase in mRNA contents of synaptophysin (6.3-fold) and NSE (5.6-fold) were measured in the spheroids relative to the monolayer cells. In agreement with the mRNA expression results, a significant (P < 0.01) increase of synaptophysin (3-fold) and NSE (2.5-fold) proteins was measured in spheroids.
Synthesis of neuronal markers in monolayer cells and spheroids was examined under the RA-induced differentiation condition. For monolayer cells, RA-treatment increased both mRNA and protein contents of synaptophysin (10- and 5.4-fold for mRNA and protein) and NSE (7.5- and 3.3-fold for mRNA and protein). Likewise, the mRNA and protein contents of synaptophysin (2.6- and 4-fold for mRNA and protein) and NSE (3- and 2.8-fold for mRNA and protein) were much higher in RA-differentiated spheroids than in undifferentiated spheroids. Neural marker expression increased in the following order: monolayer cells < spheroids < RA-treated monolayer cells < RA-differentiated spheroids
Enhanced production of CAMs in RA-differentiated spheroids
As an effort to identify the molecular nature of cell aggregation, the mRNA contents of CAMs, known to be implicated in the control of MCS formation, were measured (Fig. 4A). The levels of N-cadherin (3-fold), NCAM2 (11-fold), ICAM1 (17-fold), and connexin-26 (13-fold) mRNAs were significantly (P<0.01) higher in spheroids compared to monolayer cells, which further increased 3.3-fold, 6.9-fold, 3.6-fold, and 6.9-fold after RA treatment. In consistent with enhanced mRNA expression, protein contents of all 4 CAMs were elevated upon spheroid formation (Fig. 4B and 4C). Spheroids treated with RA demonstrated a drastic increase in CAMs production; N-cadherin (6-fold), NCAM2 (5.7-fold), ICAM1 (2.9-fold), and connexin-26 (3.2-fold) relative to untreated spheroids.
Fig. 4. Comparison of the relative contents of CAMs and ECM proteins produced in monolayer cells and spheroids cultured in the absence (-) and presence (+) of 10 μM RA. (A) qRT-PCR mRNA analysis and (B and C) Western blot immunoassay of N-cadherin, NCAM2, ICAM1 and connexin-26. (D) qRT-PCR mRNA analysis of collagen type I and IV, laminin α4, and fibronectin. All data were expressed as fold increase relative to the monolayer cells grown without RA (=1). Data are the mean ± SEM of three separate experiments. #P < 0.01, *P < 0.001 compared with control, ∥P < 0.01, ¶P < 0.05, †P < 0.001 compared with RA alone and ‡P < 0.01, §P < 0.001 compared with RGD-ELP alone.
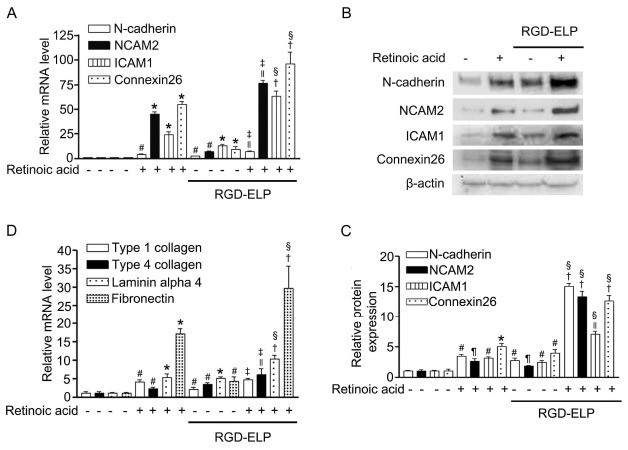
Fig. 3. Comparison of the expression levels of synaptophysin and NSE in monolayer cells and spheroids cultured without (-) and with (+) 10 μM RA analyzed by qRT-PCR (A) and Western blot immunoassay (B). All data were expressed as fold increase relative to the monolayer cells grown without RA (=1). Data are the mean ± SEM of three independent measurements. *P < 0.001, #P < 0.01 compared with control, †P < 0.001 compared with RA alone and ‡P < 0.01, §P < 0.001 compared with RGD-ELP alone.
Stronger expression of ECM proteins in RA-differentiated spheroids
Next, we compared the mRNA contents of ECM proteins among monolayer cells and spheroids (Fig. 4D). Upon the development of spheroids, the mRNA contents of type I (2-fold) and IV (3.7-fold) collagens, laminin (5.2-fold), and fibronectin (4-fold) were significantly (P < 0.05) increased. When SH-SY5Y cells were treated with RA, the levels of these proteins increased in both monolayer cells and spheroids. Specifically, the mRNA quantities of type I (2.1-fold) and IV (1.9-fold) collagens, laminin α4 (2.1-fold), and fibronectin (7.5-fold) in RA-treated spheroids were higher than those in untreated spheroids. After RA treatment, among the ECM proteins, the relative abundance of fibronectin was found to be the highest, followed by laminin α4 and collagens.
DISCUSSION
MCS have been produced from various types of primary, stem or tumor cell lines by exploiting a variety of culture techniques, including the hanging drop method, static culture on a substratum of polymers, and suspension culture in spinner flasks or wall vessels. Although there are no generalized points of reference, each method claims merit but exhibits weakness as well in terms of the ease of performance, size and shape of the produced structures, and homogeneity of the cell stage in MCS (15). As demonstrated in this study, the induced clustering of SH-SY5Y cells on the hydrophobic surfaces of elastin-like polypeptide is a simple but effective method to produce neuronal tumor spheroids in a short period of time.
In comparison with monolayer cultures, the contents of synaptophysin and NSE as well as N-cadherin and connexin-26 were higher in MCS culture. Neuronal markers and cell adhesion molecules were further up-regulated under the RA-induced differentiation condition. Aggregate formation was previously shown to be indispensable for the RA-induced neuronal differentiation of neuroectodermal progenitor cells (16). Based on such observations, the enhanced neuronal differentiation and neutrophic activity of RA on spheroids are apparently correlated with multicellular aggregation; however, further researches are necessary to provide the evidences supporting this proposal. The relatively higher expression of CAMs and ECM proteins in SH-SY5Y spheroids points to the importance of both cell-cell adhesion and cell-matrix interaction. Stronger N-cadherin expression seems to be related with the tight packing of multicellular associations through the development of N-cadherin-mediated adherens junctions (17). NCAM2 and ICAM1 are members of the integrin family of cell-cell and cell-matrix receptors (18), and thus they probably contribute to the reinforcement of spheroidal architectures by facilitating cell-cell cohesion among SH-SY5Y cells. Likewise, the connexin-26-mediated gap junction would improve the stability of SH-SY5Y spheroids, as the cases of connexin-43, by which assembly of the KGN granulosa spheroid was facilitated (19). Higher production of collagen I and IV, laminin α4, and fibronectin upon the clustering of SH-SY5Y cells into spheroids is consistent with the enhanced ECM biosynthesis by glioma BT4C spheroids (8). With regard to ECM protein production, the SH-SY5Y spheroids resemble in vivo neuroblastoma more closely than monolayer cells do.
In our experiments, in accordance with Cheung and co-workers (20), RA-induced differentiation showed drastic up-regulation of synaptophysin and NSE in SH-SY5Y cells. In contrast, Lombet et al., reported no significant change in the production of NSE after RA treatment on SK-N-SH cells (12). As suggested by Cheung et al., such discrepancy could be due to the low concentration of RA used for the SK-N-SH cell experiment (3 μM) compared to that (10 μM) used in this study. Our data obtained with monolayer cultures and MCS clearly demonstrate that RA regulates differentiation of SH-SY5Y cells and gives rise to neuron-like cells.
N-cadherin was shown to be involved in P19 neuron differentiation (21). Likewise, extensive neural differentiation and neurite outgrowth from RA-differentiated SH-SY5Y spheroids could be correlated with over-expression of NCAM and connexin-31 (22,23). Meanwhile, neurite extension by RA-treated SY5Y cells was more efficiently stimulated by laminin than by fibronectin or type I collagen (24). Therefore, extensive neuritogenesis occurred in RA-differentiated SH-SY5Y spheroids is likely to have resulted from the combined influences of CAMs and ECM proteins, rather than being the sole effect of a single cell adhesion molecule or matrix component.
There are increasing demands for in vitro model systems exhibiting a high resemblance to mature neural tissues. Accordingly, many attempts have been made to transform neuroblastoma SH-SY5Y cells into more neuron-like cells. Multicellular organization improves neuronal property as shown by enhanced amounts of neuron-specific markers in SH-SY5Y spheroids. However, spheroid culture alone may not be enough to induce sufficient neuronal differentiation. Most well differentiated neuronal characteristics were obtained by treatment of SH-SY5Y spheroids with RA. Morphologically, extensive neurite outgrowth, a typical neuronal phenotype in differentiating cells, occurred when the SH-SY5Y spheroids were cultured with RA. Physiologically, the production of synaptophysin and NSE was highly up-regulated in RA-treated SH-SY5Y spheroids. Therefore, our results indicated that SH-SY5Y spheroids would be more relevant with the in vivo brain tissues under the RA-induced differentiation condition.
In conclusion, a facile formation of SH-SY5Y spheroids was established by promoting cell-cell cohesion on thermally-collapsed elastin-like polypeptide. The SH-SY5Y spheroids express much higher amounts of CAMs and ECM proteins compared to monolayer cells. As evidenced by extensive neuritogenesis and up-regulated neuronal marker expression, the RA-differentiated SH-SY5Y spheroids are matured neuronal structures.
MATERIALS AND METHODS
Preparation of RGD-ELP
The RGD-ELP, TGPG[VGRGD(VGVPG)6]20WPC, consists of integrin-binding RGD motifs and hydrophobic VGVPG peptides and forms hydrophobic aggregates above a certain transition temperature. It was expressed using a pET-25b(+)-1 plasmid vector and purified from E. coli BLR(DE3) by reversible phase transition as previously described (13).
Maintenance of cells
The SH-SY5Y human neuroblastoma cell line (ATCC) was cultured with 10% complete medium (1:1 mixture of DMEM and Ham's F12 medium and 10% supplemental fetal bovine serum, 100 U/ml penicillin, and 100 μg/mL streptomycin) in a humidified, 5% CO2-95% air, 37℃ incubator.
Production and differentiation of spheroids
Wells of 6-well PS plates were treated with 1 ml of 0.5, 1, 5, or 10 μM RGD-ELP for 1 h at 37℃. After protein precipitation, the supernatant solution was carefully pipetted out. SH-SY5Y cells (1 × 105 cells/well) were seeded and cultured for 2 days. After 2 days, cells were treated with 10 μM all-trans-RA (Sigma) to differentiate the cells for 6 days. Cell growth patterns and morphology were recorded with a Leica DMI 3000B microscope.
Cell viability and proliferation assays
Monolayer cells and spheroids were gently washed 3 times in PBS and then treated with a Live/Dead cell staining reagent (Molecular Probes) containing 2 μM calcein AM and 4 μM EthD-1 at room temperature for 30 min. Fluorescence images were taken by a Leica DMI 3000B microscope. The numbers of live cells were counted by Tryphan Blue exclusion assay (Life technology). Briefly, 6-well plates were treated with 1 ml of 1 μM RGD-ELP and incubated at 37℃ for 1 h, SH-SY5Y cells (1 × 105 cells/well ) were seeded and cultured for the indicated time. Trypsin-EDTA was added to each well, and the cell viability, (No. of live cells in spheroids / No. of live cells in monolayer culture) × 100, was calculated by manual counting of live cells under the Leica DMI 3000B microscope.
Scanning electron microscope
The spheroids were formed on the glass plates coated using 1 μM RGD-ELP, fixed with 2% paraformaldehyde for 15 min at room temperature, and then washed with PBS twice. The cells were air-dried and photographed with a Hitachi S-4800 scanning electron microscope.
qRT-PCR
Total RNA was obtained from the cells by using a Trizol Reagent (Invitrogen) according to the manufacturer’s instructions. cDNA was synthesized using High-Capacity cDNA reverse transcription kits (Applied Biosystems). Quantitative real-time RT PCR was performed using the SYBR Green PCR master mix kit (Applied Biosystems) on the ABI 7500 Real Time PCR System under the following conditions: Cycling conditions were 2 min at 50℃, 10 min at 95℃, followed by 40 cycles of 95℃ for 15 s and 60℃ for 1 min. Primer sets were designed using primer express 3.0 software based on the human gene sequences from GenBank and are as follows: synaptophysin (sense: 5'-TCCTCGTCAGCCGAATTCTTT-3' and antisense: 5'-CTCGCTACTTGTTCTGCAGGAA-3'); gamma neuron specific enolase (NSE, sense: 5'-CGGGAACTGCCCCTGTATC-3' and antisense: 5'-CATGAGAGCCACCATTGATCA-3'); type 1 collagen (sense: 5'-GAGGGCCAAGACGAAGACATC-3' and antisense: 5'-GGTTGTCGCAGACGCAGAT-3'); type 4 collagen (sense: 5'-CGAGCTCTACCGCAACGACTA-3' and antisense: 5'-AGGCTTCGTGTTTCATGACCTT-3'); laminin α4 (sense: 5'-GC CTGCGGCCGAGAA-3' and antisense: 5'-TCCTGAGCCGTCC AAACACT-3'); fibronectin (sense: 5'-CCTCGGGCCCAGATAA CAG-3' and antisense: 5'-CGAGGGATACGGTGTACTCAGA T-3'); N-cadherin (sense: 5'-CACCATATGATCCCCTGTTAGTG TTT-3' and antisense: 5'-TCCTGCTCACCACCACTACTTG-3'); NCAM2 (sense: 5'-CTGCTGCTAATTCTTGTGGTAACAG-3' and antisense: 5'-CTGGAGCCACTTTTCTTTCCA-3'); ICAM1 (sense: 5'-GCCGGCCAGCTTATACACA-3' and antisense: 5'-TTCTGG CCACGTCCAGTTTC-3'); connexin-26 (sense: 5'-GTGAACAAACACTCCACCAGCAT-3' and antisense: 5'-CTCCCCACACCTCCTTTGC-3'); GAPDH (sense: 5'-ACCCACTCCTCCACCTTTGA-3' and antisense: 5'-TGTTGCTGTAGCCAAATTCGTT-3'). The housekeeping gene GAPDH was used as an internal standard. Reaction specificity was confirmed by melting curve analysis.
Western blot immunoassay
Cells were washed twice with PBS and suspended in a RIPA buffer (Sigma) containing a Complete Mini Protease Inhibitor Cocktail (Roche) and HaltTM Phosphatase Inhibitor Cocktail (Thermo), which were added according to the manufacturer’s instructions. The cells were lysed on ice for 30 min. The cell lysate was collected after centrifugation at 15,000 × g for 10 min. Protein quantification was performed using a Bio-Rad Protein Assay system (Bio-Rad). Cell lysates of 30 μg were electrophoresed by SDS-PAGE and then electrotransferred to polyvinylidine fluoride membranes (Millipore). After blocking with 5% skimmed milk in Tris-buffered saline containing Tween 20 (0.1%) for 1 h, the membrane was incubated with anti-synaptophysin (Millipore), anti-human gamma neuron-specific enolase (NSE; Chemicon Inc.), anti-N-cadherin (Abcam), anti-NCAM2 (Abcam), anti-ICAM1 (Abcam), and anti-Connexin-26 (Abcam) antibodies at 4℃ with gentle shaking overnight, and washed 3 times for 10 min in a washing buffer (Tris-buffered saline containing 0.1% Tween 20). Antibodies were detected with a horseradish peroxidase-linked secondary antibody by using the Super Signal West Dura Extended Duration substrate according to the manufacturer’s instructions (Thermo). The membrane was re-blotted with anti-β-actin antibodies (Sigma) to verify the equal loading of protein in each lane. Densitometric measurements of the bands were performed using the digitalized scientific program UN-SCAN-IT (Silk Scientific Corp.).
Statistical data analysis
Data were expressed as means ± SEM from at least three independent experiments. The statistical significance in regard to the differences between groups was determined by one-way analysis of variance (ANOVA) and the Tukey post-hoc test. All statistical analyses were performed using GraphPad InStat (Ver. 3.05). Values of P < 0.05 were considered statistically significant.
Acknowledgments
This study was supported by the Biodefense Program Fund (Project No. 11-BD-02) to WBJ from the Ministry of Education, Science and Technology of the Republic of Korea. Authors appreciate Mi-Ae Kwon for RGD-ELP purification.
References
- 1.Hirschhaeuser F., Menne H., Dittfeld C., West J., Mueller-Klieser W., Kunz-Schughart L. A. Multicellular tumor spheroids: an underestimated tool is catching up again. J. Biotechnol. (2010);148:3–15. doi: 10.1016/j.jbiotec.2010.01.012. [DOI] [PubMed] [Google Scholar]
- 2.Bilir A., Erguven M., Yazihan N., Aktas E., Oktem G., Sabanci A. Enhancement of vinorelbine-induced cytotoxicity and apoptosis by clomipramine and lithium chloride in human neuroblastoma cancer cell line SH-SY5Y. J. Neurooncol. (2010);100:385–395. doi: 10.1007/s11060-010-0209-6. [DOI] [PubMed] [Google Scholar]
- 3.Walker K. A., Murray T., Hilditch T. E., Wheldon T. E., Gregor A., Hann I. M. A tumour spheroid model for antibody-targeted therapy of micrometastases. Br. J. Cancer. (1988);58:13–16. doi: 10.1038/bjc.1988.152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Walker K. A., Mairs R., Murray T., Hilditch T. E., Wheldon T. E., Gregor A., Hann I. M. Tumor spheroid model for the biologically targeted radiotherapy of neuroblastoma micrometastases. Cancer Res. (1990);50:1000s–1002s. [PubMed] [Google Scholar]
- 5.Yuhas J. M., Li A. P., MartInez A. O., Ladman A. J. A simplified method for production and growth of multicellular tumor spheroids. Cancer Res. (1997);37:3639–3643. [PubMed] [Google Scholar]
- 6.Santini M. T., Rainaldi G., Indovina P. L. Apoptosis, cell adhesion and the extracellular matrix in the three-dimensional growth of multicellular tumor spheroids. Crit. Rev. Oncol. Hematol. (2000);36:75–87. doi: 10.1016/S1040-8428(00)00078-0. [DOI] [PubMed] [Google Scholar]
- 7.Neugebauer K. M., Tomaselli K. J., Lilien J., Reichardt L. E. N-cadherin, NCAM, and integrins promote retinal neurite outgrowth on astrocytes in vitro. J. Cell Biol. (1988);107:1177–1187. doi: 10.1083/jcb.107.3.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bjerkvig R., Lucrimi O. D., Rucklidge G. J. Immunocytochemical characterization of extracellular matrix proteins expressed by cultured glioma cells. Cancer Res. (1989);49:5424–5428. [PubMed] [Google Scholar]
- 9.Biedler J. L., Roffler-Tarlov S., Schachner M., Freedman L. S. Multiple neurotransmitter synthesis by human neuroblastoma cell lines and clones. Cancer Res. (1978);38:3751–3757. [PubMed] [Google Scholar]
- 10.Pahlman S., Ruusala A. I., Abrahamsson L., Mattsson M. E., Esscher T. Retinoic acid-induced differentiation of cultured human neuroblastoma cells: a comparison with phorbolester-induced differentiation. Cell Differ. (1984);14:135–144. doi: 10.1016/0045-6039(84)90038-1. [DOI] [PubMed] [Google Scholar]
- 11.Lopes F. M., Schröder R., da Frota M. L. Jr., Zanotto-Filho A., Müller C. B., Pires A. S., Meurer R. T., Colpo G. D., Gelain D. P., Kapczinski F., Moreira J. C., Fernandes M. C., Klamt F. Comparison between proliferative and neuron-like SH-SY5Y cells as an in vitro model for Parkinson disease studies. Brain Res. (2010);1337:85–94. doi: 10.1016/j.brainres.2010.03.102. [DOI] [PubMed] [Google Scholar]
- 12.Lombet A., Zujovic V., Kandouz M., Billardon C., Carvajal-Gonzalez S., Gompel A., William R. Resistance to induced apoptosis in the human neuroblastoma cell line SK-N-SH in relation to neuronal differentiation. Role of Bcl-2 protein family. Eur. J. Biochem. (2001);268:1352–1362. doi: 10.1046/j.1432-1327.2001.02002.x. [DOI] [PubMed] [Google Scholar]
- 13.Jeon W. B., Park B. H., Wei J., Park R. W. Stimulation of fibroblasts and neuroblasts on a biomimetic extracellular matrix consisting of tandem repeats of the elastic VGVPG domain and RGD motif. J. Biomed. Mater. Res. (2011);A97:2152–2157. doi: 10.1002/jbm.a.33041. [DOI] [PubMed] [Google Scholar]
- 14.Lee K. M., Jung G. S., Park J. K., Choi S. K., Jeon W. B. Effects of Arg-Gly-Asp-modified elastin-like polypeptide on pseudoislet formation via up-regulation of cell adhesion molecules and extracellular matrix proteins. Acta Biomater. (2013);9:5600–5608. doi: 10.1016/j.actbio.2012.10.036. [DOI] [PubMed] [Google Scholar]
- 15.Lin R. Z., Chang H. Y. Recent advances in three-dimensional multicellular spheroid culture for biomedical research. Biotechnol. J. (2008);3:1172–1184. doi: 10.1002/biot.200700228. [DOI] [PubMed] [Google Scholar]
- 16.Tárnok K., Pataki A., Kovács J., Schlett K., Madarász E. Stage-dependent effects of cell-to-cell connections on in vitro induced neurogenesis. Eur. J. Cell Biol. (2002);81:403–412. doi: 10.1078/0171-9335-00262. [DOI] [PubMed] [Google Scholar]
- 17.Ivascu A., Kubbies M. Diversity of cell-mediated adhesions in breast cancer spheroids. Int. J. Oncol. (2007);31:1403–1413. [PubMed] [Google Scholar]
- 18.Staunton D. E., Marlin S. D., Stratowa C., Dustin M. L., Springer T. A. Primary structure of ICAM-I demonstrates interaction between members of the immunoglobulin and integrin supergene families. Cell. (1988);52:925–933. doi: 10.1016/0092-8674(88)90434-5. [DOI] [PubMed] [Google Scholar]
- 19.Bao B., Jiang J., Yanase T., Nishi Y., Morgan J. R. Connexon-mediated cell adhesion drives microtissue self-assembly. FASEB J. 25:255–264. doi: 10.1096/fj.10-155291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cheung Y., Lau W. K., Yu M., Lai C. S., Yeung S., So K., Chang R. C. Effects of all-trans-retinoic acid on human SH-SY5Y neuroblastoma as in vitro model in neurotoxicity research. Neurotoxicology. (2009);30:127–135. doi: 10.1016/j.neuro.2008.11.001. [DOI] [PubMed] [Google Scholar]
- 21.Gao X., Bian W., Yang J., Tang K., Kitani H., Atsumi T., Jing N. A role of N-cadherin in neuronal differentiation of embryonic carcinoma P19 cells. Biochem. Biophys. Res. Commun. (2001);284:1098–1103. doi: 10.1006/bbrc.2001.5089. [DOI] [PubMed] [Google Scholar]
- 22.Seidenfaden R., Krauter A., Hildebrandt H. The neural cell adhesion molecule NCAM regulates neuritogenesis by multiple mechanisms of interaction. Neurochem. Int. (2006);49:1–11. doi: 10.1016/j.neuint.2005.12.011. [DOI] [PubMed] [Google Scholar]
- 23.Unsworth H. C., Aasen T., McElwaine S., Kelsell D. P. Tissue-specific effects of wild-type and mutant connexin 31: a role in neurite outgrowth. Hum. Mol. Genet. (2007);16:165–172. doi: 10.1093/hmg/ddl452. [DOI] [PubMed] [Google Scholar]
- 24.Rossino P., Defilippi P., Silengo L., Tarone G. Up-regulation of the integrin α1/β1 in human neuroblastoma cells differentiated by retinoic acid: correlation with increased neurite outgrowth response to laminin. Cell Regul. (1991);2:1021–1033. doi: 10.1091/mbc.2.12.1021. [DOI] [PMC free article] [PubMed] [Google Scholar]