

Abstract
Objective
Mononuclear cell infiltration in valvular tissue is one of the characteristics in calcific aortic valve disease. The inflammatory responses of aortic valve interstitial cells (AVICs) play an important role in valvular inflammation. However, it remains unclear what may evoke AVIC inflammatory responses. Accumulation of biglycan has been found in diseased aortic valve leaflets. Soluble biglycan can function as a danger-associated molecular pattern to induce the production of pro-inflammatory mediators in cultured macrophages. We tested the hypothesis that soluble biglycan induces AVIC production of pro-inflammatory mediators involved in mononuclear cell infiltration through Toll-like receptor (TLR)-dependent signaling pathways.
Methods
Human AVICs isolated from normal aortic valve leaflets were treated with specific siRNA and neutralizing antibody against TLR2 or TLR4 before biglycan stimulation. The production of ICAM-1 and MCP-1 were assessed. To determine the signaling pathway involved, phosphorylation of ERK1/2 and p38 MAPK was analyzed, and specific inhibitors of ERK1/2 and p38 MAPK were applied.
Results
Soluble biglycan induced ICAM-1 expression and MCP-1 release in human AVICs, but had no effect on IL-6 release. TLR4 blockade and knockdown reduced ICAM-1 and MCP-1 production induced by biglycan, while knockdown and neutralization of TLR2 resulted in greater suppression of the inflammatory responses. Biglycan induced the phosphorylation of ERK1/2 and p38 MAPK, but ICAM-1 and MCP-1 production was reduced only by inhibition of the ERK1/2 pathway. Further, inhibition of ERK1/2 attenuated NF-κB activation following biglycan treatment.
Conclusions
Soluble biglycan induces the expression of ICAM-1 and MCP-1 in human AVICs through TLR2 and TLR4, and requires activation of the ERK1/2 pathway. AVIC inflammatory responses induced by soluble biglycan may contribute to the mechanism of chronic inflammation associated with calcific aortic valve disease.
Keywords: Biglycan, AVIC, Cytokines, TLR2, TLR4
INTRODUCTION
Chronic inflammation is a prominent feature of calcific aortic valve disease (CAVD). Increased levels of proinflammatory cytokines and inflammatory cell infiltration, including T lymphocytes and monocytes, are observed in diseased aortic valves [1, 2]. Leukocytes migrate into the interstitial space of the heart valve in response to locally produced chemoattractant molecules [3]. Aortic valve interstitial cells (AVICs) are capable of expressing proinflammatory mediators that may initiate and promote the inflammatory process [4]. Thus, AVICs are believed to play an important role in the progression of CAVD.
Molecular imaging in inflammation-driven disease reveals increased expression of proinflammatory mediators (eg, monocyte chemotactic protein 1 [MCP-1] and intercellular adhesion molecule 1 [ICAM-1]). MCP-1, acting through its receptor CCR2, is critical for the recruitment of leukocytes to the sites of inflammation [5]. ICAM-1 is an immunoglobulin (Ig)-like cell adhesion molecule [6]. Following stimulation or injury, ICAM-1 expression is up-regulated in conjunction with local inflammation and increased leukocyte infiltration [7]. We have reported that stimulation of human AVICs with lipopolysaccharide induces the release of MCP-1 and the expression of ICAM-1 [8, 9]. However, the role of endogenous agents in the induction of AVIC inflammatory response is not well understood.
As a critical determinant of valve function and durability, AVICs have close contact with the extracellular matrix (ECM) and regulates ECM components. In this regard, AVICs are known to synthesize ECM proteins, including biglycan, which are crucial to maintaining the quality and quantity of the ECM in aortic valves [4]. Biglycan, a member of the family of small proteoglycans, is a stationary component of the ECM [10]. However, when biglycan is secreted by cells or is detached from the ECM, it becomes available in a soluble form [11-13]. Soluble biglycan has been found to induce proinflammatory cytokine production in macrophages through TLR2 and TLR4 [14, 15]. We have previously reported that human AVICs express functional TLR2 and TLR4 and activation of these innate immune receptors in human AVICs induces the expression of proinflammatory mediators [16]. However, endogenous ligands to TLR2 and TLR4 in the aortic valve remain to be identified. Interestingly, biglycan accumulation is observed in calcific, stenotic areas of human aortic valves [17, 18]. Further, soluble biglycan is capable of inducing AVIC expression of a phospholipid transfer protein through TLR2 [17]. Key questions arise: does soluble biglycan induce the expression of proinflammatory mediators in human AVICs and what is the signaling mechanism is if it does? Therefore, we aimed to determine the effect of biglycan on the expression of MCP-1 and ICAM -1 in human AVICs and to identify the signaling pathway involved.
We hypothesized that soluble biglycan induces the production of MCP-1 and ICAM-1 in human AVICs via TLR2 and/or TLR4. The purpose of this study was to determine: 1) the effect of soluble biglycan on MCP-1 and ICAM-1 production in human AVICs, 2) the impact of knockdown and neutralization of TLR2 and TLR4 on the effect of biglycan, and 3) the signaling pathway that is required for biglycan to exert an effect.
MATERIALS AND METHODS
Materials
Tissue culture medium (M199) was purchased from Lonza (Walkersville, MD). Antibiotics and collagenase were from GIBCO Laboratories (Grand Island, N.Y.). Fetal bovine serum was from Alekenbio (Nash, TX). Recombinant human biglycan and MCP-1 ELISA kit were purchased from R&D System (Minneapolis, MN). Antibodies against ICAM-1, phosphorylated extracellular signal-regulated kinase 1/2 (p-ERK1/2), total ERK1/2 (t-ERK1/2), phosphorylated p38 mitogen-activated protein kinase (p-MAPK, pp38), total p38 MAPK (tp38), phosphorylated nuclear factor-κB (pNF-κB) and total NF-κB (tNF-κB) were purchased from Cell Signaling, Inc. (Beverly, MA). Antibodies against TLR2 and TLR4, specific siRNA for human TLR2 or TLR4, and scrambled siRNA were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). HiPerFect® transfection reagent and other transfection-related reagents were purchased from Qiagen (Valencia, CA). All other chemicals and reagents were from Sigma-Aldrich Chemical Co. (St Louis, MO).
Cell isolation and treatment
Human AVICs were derived from normal aortic valve leaflets from the explanted hearts of 6 patients (4 males and 2 females; age 59.0±8.1 years; all with cardiomyopathy) undergoing heart transplantation at the University of Colorado Hospital. These valve leaflets were thin and did not exhibit histological abnormalities. All patients gave informed consent for the use of their valves for this study. This study was approved by the University of Colorado Denver Institutional Review Board.
AVICs were isolated and cultured as previously described [19] with some modifications [16]. Briefly, valve leaflets were subjected to sequential digestions with collagenase, and cells were collected by centrifugation. Cells were cultured in M199 growth medium supplemented with penicillin G, streptomycin, amphotericin B and 10% fetal bovine serum. Cells from passages 3 to 6 were used for this study. Experiments were performed when cells reached 80 to 90% confluence on plates.
The effect of biglycan on the inflammatory response was determined following stimulation of cells with varied concentrations of biglycan (0.05, 0.10 and 0.20 μg/ml) for 48 h. Levels of MCP-1 in culture medium and ICAM-1 in cell lysate were assessed using ELISA and immunoblotting, respectively.
To determine the effects of biglycan on phosphorylation of ERK1/2 and p38 MAPK, cells were stimulated with biglycan (0.10 μg/ml) for 60 to 240 min. To determine the role of ERK1/2 and p38 MAPK in the effect of biglycan on the inflammatory response, specific ERK1/2 inhibitor (PD98059, 10-40 μmol/L) and p38 MAPK inhibitor (SB203580, 5-20 μmol/L) were added to culture medium 30 min before adding biglycan.
Immunoblotting
Immunoblotting was performed to analyze ICAM-1, TLR2, TLR4, phosphorylated and total ERK1/2, phosphorylated and total p38 MAPK, and phosphorylated and total NF-κB. Beta-actin was utilized as a loading control. After stimulation, human AVICs were lysed in a sample buffer (100 mM Tris-HCl, pH 6.8, 2% SDS, 0.02% bromophenol blue and 10% glycerol). The cell lysates were then run on 4-20% SDS-PAGE and the proteins were transferred onto PVDF membranes. After being blocked with 5% skim milk solution, membranes were incubated with primary antibodies, followed by peroxidase-linked secondary antibodies specific to the primary antibodies. Protein bands were revealed using the enhanced chemiluminescence system. The band density was determined using the National Institutes of Health Image J software (Wayne Rasband, National Institutes of Health, Bethesda, MD).
ELISA
The levels of MCP-1 in cell culture supernatants were measured using a commercially available ELISA kit according to the manufacturer's instructions. MCP-1 levels in the culture supernatant were measured in triplicate with an automatic microplate reader (BIO-RAD680, Hercules, CA) at a wavelength of 450 nm.
Gene knockdown
To knockdown TLR2 and TLR4, cells were transfected with siRNAs using HiPerFect® transfection reagent. Briefly, cells (60-70% confluent) in 24-well plates were incubated with a mixture of siRNA (60 nM) and transfection reagent (6 μl per ml of medium) for 48 h. Control cells were treated with scrambled siRNA and transfection reagent. Knockdown of a specified gene was confirmed by immunoblot analysis for protein expression.
Statistical analysis
Data were analyzed using StatView software (Abacus Concepts, Calabasas, CA). All values are expressed as mean ± standard error (SE). Comparisons between groups were performed by one-way ANOVA, complimented by Tukey's HSD/Dunn test when appropriate. A difference was considered significant at P< 0.05.
RESULTS
Biglycan induces ICAM-1 and MCP-1 production in human AVICs
To examine AVIC inflammatory response to soluble biglycan, we analyzed the expression of ICAM-1 and the release of MCP-1 and IL-6 by cells stimulated with recombinant human biglycan, 0.05-0.20 μg/ml, for 48 hours. As shown in Figure 1A, biglycan at 0.10 or 0.20 μg/ml up-regulated ICAM-1 protein levels. In addition, biglycan at these two concentrations induced MCP-1 secretion (Figure 1B). However, biglycan had no effect on the release of IL-6 (Figure 1B). These results demonstrate that soluble biglycan induces the inflammatory response characterized by expression of ICAM-1 and MCP-1 in human AVICs.
Figure 1. Biglycan up-regulates ICAM-1 expression and MCP-1 release in human AVICs.

Human AVICs from normal valves were treated with biglycan (BGN; 0.05, 0.10 and 0.20 μg/mL) for 48 h. A. Representative immunoblots of four separate experiments and densitometric data show dose-dependent induction of ICAM-1 by biglycan. B. Biglycan promoted the release of MCP-1, but had no effect on IL-6 release. Values are means ± SE. n = 4; *P<0.05 vs. untreated control.
Both TLR2 and TLR4 are involved in biglycan-induced inflammatory response in human AVICs
We determined the effects of neutralizing antibodies against TLR2 and TLR4 on ICAM-1 and MCP-1 production. Neutralization of TLR2 markedly reduced ICAM-1 protein levels following biglycan stimulation (Figure 2A). MCP-1 production was reduced by 75.58% (P<0.05; Figure 2A) in AVICs treated with TLR2-neutralizing antibody before biglycan stimulation. Meanwhile, neutralization of TLR4 resulted in a moderate reduction in ICAM-1 and MCP-1 production in response to biglycan (Figure 2A).
Figure 2. TLR2 and TLR4 are required for ICAM-1 and MCP-1 production induced by biglycan.

A. Representative immunoblots and densitometric data show that neutralization of TLR2 markedly reduced cellular levels of ICAM-1 and MCP-1 release following treatment with biglycan (BGN, 0.10 μg/mL for 48 h), while neutralization of TLR4 had a moderate effect. B. Treatment with specific siRNAs reduced cellular TLR2 and TLR4 levels. Silencing TLR2 markedly reduced cellular ICAM-1 levels and MCP-1 release after biglycan stimulation, while silencing TLR4 had a moderate effect. Values are means ± SE. n = 3; *P<0.05, vs. untreated control; #P<0.05 vs. BGN alone; †P<0.05 vs. BGN + non-immune IgG; ‡P<0.05 vs. BGN + scrambled siRNA.
To further determine the role of TLR2 and TLR4 in biglycan-induced inflammatory response, TLR2 siRNA and TLR4 siRNA were applied before biglycan stimulation. As shown in Figure 2B, treatment with a specific siRNA for TLR2 or TLR4, but not a control siRNA, markedly reduced the protein levels of TLR2 or TLR4. TLR4 knockdown reduced ICAM-1 and MCP-1 production induced by biglycan, while knockdown of TLR2 resulted in greater suppression of the inflammatory responses (Figure 2B). Knockdown of TLR2 markedly reduced production of ICAM-1 and MCP-1 following biglycan stimulation. The results suggest that biglycan induces the inflammatory response in human AVICs through both TLR2 and TLR4. Between these two receptors, TLR2 has a more important role in mediating biglycan-induced production of ICAM-1 and MCP-1.
Inhibition of ERK1/2 suppresses ICAM-1 and MCP-1 production induced by biglycan
ERK1/2 and p38 MAPK signaling pathways play a central role in proinflammatory signaling downstream of TLR2 and TLR4 [20]. ERK1/2 and p38 MAPK activation was determined in AVICs by analysis of their phosphorylation. As shown in Figure 3A, biglycan stimulation resulted in p38 MAPK phosphorylation. However, inhibition of p38 MAPK with SB203580 had no effect on the production of ICAM-1 and MCP-1 (Figure 3B). Phosphorylation of ERK1/2 was also induced by biglycan (Figure 4A). Inhibition of ERK1/2 with PD98059 (10-40 μmol/L) markedly decreased biglycan-induced ICAM-1 expression. Similarly, inhibition of ERK1/2 with varied concentration of PD98059 reduced MCP-1 levels by 58.97% - 61.43% (P<0.05; Figure 4B).
Figure 3. Inhibition of p38 MAPK has a minor effect on biglycan-induced ICAM-1 and MCP-1 production in human AVICs.

A. Representative immunoblots show that biglycan (BGN, 0.10 μg/mL) induced rapid phosphorylation of p38 MAPK. B. Inhibition of p38 MAPK with SB203580 (5, 10 and 20 μmol/L) resulted in a minimal reduction in ICAM-1 and MCP-1 production following biglycan stimulation. Values are means ± SE. n = 3; *P<0.05 vs. untreated control.
Figure 4. Inhibition of ERK1/2 suppresses ICAM-1 and MCP-1 production in human AVICs.

A. Representative immunoblots show that biglycan (BGN, 0.10 μg/mL) induced rapid phosphorylation of ERK1/2, and PD98059 at 20 μmol/L attenuated BGN-induced ERK1/2 phosphorylation. B. Inhibition of ERK1/2 markedly reduced ICAM-1 and MCP-1 production following biglycan stimulation. Values are means ± SE. n = 3; *P<0.05 vs. untreated control; #P<0.05 vs BGN alone.
ERK1/2 is involved in mediating biglycan-induced NF-κB phosphorylation
NF-κB is important for AVIC production of MCP-1 and ICAM-1 [21, 22]. We sought to determine the relationship between ERK1/2 activation and NF-κB phosphorylation in human AVICs following biglycan stimulation. Cells treated with biglycan exhibited increases in the phosphorylated form of NF-κB. Inhibition of ERK1/2 with PD98059 (20 μmol/L) markedly reduced biglycan-induced NF-κB phosphorylation (Figure 5). These results together indicate that the ERK1/2-NF-κB cascade is involved in the mechanism of TLR2/4-dependent AVIC inflammatory response to biglycan stimulation.
Figure 5. The ERK1/2 pathway is involved in biglycan-induced NF-κB activation in human AVICs.
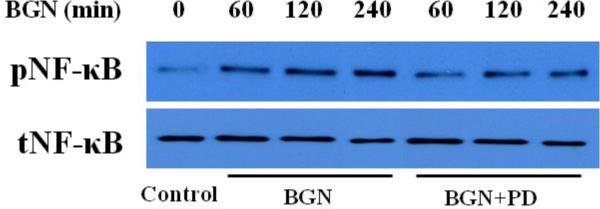
Cells were treated with biglycan (BGN, 0.10 μg/mL) for 60 to 240 min in the presence of PD98059 (20 μmol/L). A representative immunoblot shows that inhibition of ERK1/2 reduced NF-κB phosphorylation induced by biglycan.
DISCUSSION
CAVD occurs commonly in people over 65 years of age [23]. Unfortunately, mechanisms underlying the pathogenesis of CAVD are not well understood. The scientific consensus is that chronic inflammatory changes are important events responsible for progression of this disease. However, the molecular mechanism underlying aortic valve inflammation remains to be elucidated. We observed in the present study that soluble biglycan induces ICAM-1 expression and MCP-1 release in human AVICs. Knockdown of TLR2 and TLR4 significantly reduced the induction of ICAM-1 and MCP-1 by biglycan. Further, inhibition of ERK1/2 essentially abolished biglycan-induced production of these two proinflammatory mediators. Notably, we demonstrated that activation of NF-κB by biglycan is ERK1/2 dependent. These results indicate a novel paradigm that biglycan induces the production of ICAM-1 and MCP-1 in the aortic valve mainly via the TLR/ERK1/2/NF-κB pathway. This study is the first to demonstrate endogenous protein biglycan can induce the expression of proinflammatory mediators in human AVICs through immunoreceptors.
Soluble biglycan induces the expression of ICAM-1 and MCP-1 in human AVICs via TLR2 and TLR4
Biglycan accumulation is observed in calcific, stenotic areas of human aortic valves [17, 18]. Our previous studies and the work by others demonstrate that TLR2 is required for mediating the effects of soluble biglycan in the induction of an osteogenic response and the expression of phospholipid transfer protein in AVICs [17, 24]. These studies suggest that biglycan functions as a pathogen-associated molecular pattern. Several studies suggest that biglycan modulates the inflammatory process [12, 13, 15, 25]. In a murine unilateral ureteral obstruction model, biglycan up-regulation is sustained in the renal interstitium [26]. In addition, the expression of biglycan is found to be elevated in human fibroblasts from granulation tissue and bronchial mucosa of asthmatic patients [27]. Such elevated expression suggests a role for biglycan in the development of chronic inflammatory lesions. In murine primary peritoneal macrophages, biglycan induces the production of proinflammatory cytokines, such as TNF-α, pro–IL-1β, MIP-2, MIP-1α, MCP-1, CXCL13, and RANTES [11, 14].
We found in this study that soluble biglycan up-regulates cellular ICAM-1 levels and MCP-1 release in human AVICs. Particularly, biglycan at 0.10 μg/ml has a robust effect on ICAM-1 expression and MCP-1 release. It is well known that ICAM-1 and MCP-1 are two important inflammatory mediators that mediate leukocyte infiltration [5, 7]. It is likely that biglycan modulates aortic valve inflammatory response and contributes to the mechanism underlying CAVD progression.
The “early lesions” associated with CAVD are characterized by inflammatory changes, including the accumulation of T lymphocytes, mononuclear cells and macrophages in valvular tissue, and an inflammatory milieu appears to promote aortic valve calcification [28]. In this regard, AVICs are recognized as the most important cell type involved in aortic valve inflammation and calcification [22, 29]. Further, AVICs are found to differentiate to osteoblast-like cells when exposed to pro-inflammatory mediators [16, 22, 30]. The results of the present study show that biglycan up-regulates the expression of ICAM-1 and MCP-1 in human AVICs. Since these two proinflammatory mediators are important for leukocyte adherence to endothelial cells and to migrate to the interstitial spaces, up-regulation of their expression by biglycan may contribute to the mechanism of chronic inflammation associated with CAVD progression. In addition, these two inflammatory mediators may contribute to the mechanism of aortic valve calcification by paracrine effect on AVIC pro-osteogenic differentiation. Indeed, we have observed that prolonged stimulation of human AVICs with biglycan induces the expression of osteogenic biomarkers in the absence of leukocytes [24]. Nevertheless, up-regulation of the expression of ICAM-1 and MCP-1 in human AVICs may contribute to the pathogenesis of CAVD through leukocyte -dependent and -independent mechanisms. Further studies are needed, however, to determine the role of ICAM-1 and MCP-1 expressed by AVIC in aortic valve inflammation and calcification.
Previous studies found that biglycan induces production of proinflammatory mediators in cultured macrophages via TLR2 and TLR4 [14, 25]. Human AVICs express functional TLR2 and TLR4 [16]. We have previously found that activation of TLR2 or TLR4 up-regulates ICAM-1 expression and MCP-1 release in human AVICs [16]. Further, our previous work shows that biglycan can interact with TLR2 and TLR4 on human AVICs [24]. In the present study, we determined the role of TLR2 and TLR4 in the induction of ICAM-1 and MCP-1 by biglycan. We found that TLR4 blockade reduced ICAM-1 and MCP-1 production induced by biglycan, while TLR2 blockade resulted in greater suppression of the inflammatory responses. By utilizing a knockdown approach, we confirmed that induction of the AVIC inflammatory response by biglycan involves both TLR2 and TLR4 while TLR2 has a more important role.
ERK1/2- NF-κB pathway is involved in post-receptor signaling
TLR2 and TLR4 activates multiple proinflammatory signaling pathways [31-33]. In the present study, we found that biglycan induces rapid phosphorylation of p38 MAPK and ERK1/2. Inhibition of ERK1/2 with PD98059 markedly decreased biglycan-induced ICAM-1 expression and MCP-1 release in human AVICs while inhibition of p38 MAPK had no effect. The results suggest that ERK1/2 is the factor downstream of TLR2 and TLR4 in mediating the induction of the proinflammatory mediators by biglycan in human AVICs.
NF-κB plays a central role in TLR-mediated inflammatory response [34, 35]. In the present study, we observed that phosphorylation of NF-κB is induced in human AVICs following biglycan stimulation. Inhibition of ERK1/2 with PD98059 markedly reduces biglycan-induced NF-κB activation. Taken together, our results indicate that the ERK1/2-NF-κB cascade is an important mechanism in the TLR2- and TLR4-dependent induction of the proinflammatory mediators by biglycan.
Collectively, our findings demonstrate that biglycan induces the production of ICAM-1 and MCP-1 in human AVICs through TLR2 and TLR4. TLR2 plays a more important role. Further, biglycan activates the ERK1/2-NF-κB cascade that is proinflammatory. Biglycan accumulation is evident in diseased aortic valves [17, 18, 36]. Therefore, activation of the immunoreceptor-mediated inflammatory response in AVICs by soluble biglycan may contribute to the mechanism of chronic inflammation associated with CAVD.
Acknowledgements
a) Acknowledgement: None
b) Sources of Funding: This study was supported in part by National Institutes of Heart, Lung and Blood Grant HL106582.
Footnotes
c) Disclosure: None
References
- 1.Freeman RV, Otto CM. Spectrum of calcific aortic valve disease: pathogenesis, disease progression, and treatment strategies. Circulation. 2005;111(24):3316–26. doi: 10.1161/CIRCULATIONAHA.104.486738. [DOI] [PubMed] [Google Scholar]
- 2.Parolari A, Loardi C, Mussoni L, Cavallotti L, Camera M, Biglioli P, et al. Nonrheumatic calcific aortic stenosis: an overview from basic science to pharmacological prevention. Eur J Cardiothorac Surg. 2009;35(3):493–504. doi: 10.1016/j.ejcts.2008.11.033. [DOI] [PubMed] [Google Scholar]
- 3.Charo IF, Taubman MB. Chemokines in the pathogenesis of vascular disease. Circ Res. 2004;95(9):858–66. doi: 10.1161/01.RES.0000146672.10582.17. [DOI] [PubMed] [Google Scholar]
- 4.Taylor PM, Batten P, Brand NJ, Thomas PS, Yacoub MH. The cardiac valve interstitial cell. Int J Biochem Cell Biol. 2003;35(2):113–8. doi: 10.1016/s1357-2725(02)00100-0. [DOI] [PubMed] [Google Scholar]
- 5.Schober A. Chemokines in vascular dysfunction and remodeling. Arterioscler Thromb Vasc Biol. 2008;28(11):1950–9. doi: 10.1161/ATVBAHA.107.161224. [DOI] [PubMed] [Google Scholar]
- 6.New SE, Aikawa E. Molecular imaging insights into early inflammatory stages of arterial and aortic valve calcification. Circ Res. 2011;108(11):1381–91. doi: 10.1161/CIRCRESAHA.110.234146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Galkina E, Ley K. Vascular adhesion molecules in atherosclerosis. Arterioscler Thromb Vasc Biol. 2007;27(11):2292–301. doi: 10.1161/ATVBAHA.107.149179. [DOI] [PubMed] [Google Scholar]
- 8.Song Y, Fullerton DA, Mauchley D, Su X, Ao L, Yang X, et al. Microfilaments Facilitate TLR4-Mediated ICAM-1 Expression in Human Aortic Valve Interstitial Cells. J Surg Res. 2009;166:52–58. doi: 10.1016/j.jss.2009.03.101. [DOI] [PubMed] [Google Scholar]
- 9.Zeng Q, Jin C, Ao L, Cleveland JC, Jr., Song R, Xu D, et al. Cross-talk between the toll-like receptor 4 and notch1 pathways augments the inflammatory response in the interstitial cells of stenotic human aortic valves. Circulation. 2012;126(11 Suppl 1):S222–S230. doi: 10.1161/CIRCULATIONAHA.111.083675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Schaefer L, Iozzo RV. Biological functions of the small leucine-rich proteoglycans: from genetics to signal transduction. J Biol Chem. 2008;283(31):21305–9. doi: 10.1074/jbc.R800020200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Moreth K, Brodbeck R, Babelova A, Gretz N, Spieker T, Zeng-Brouwers J, et al. The proteoglycan biglycan regulates expression of the B cell chemoattractant CXCL13 and aggravates murine lupus nephritis. J Clin Invest. 2010;120(12):4251–72. doi: 10.1172/JCI42213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Popovic ZV, Wang S, Papatriantafyllou M, Kaya Z, Porubsky S, Meisner M, et al. The proteoglycan biglycan enhances antigen-specific T cell activation potentially via MyD88 and TRIF pathways and triggers autoimmune perimyocarditis. J Immunol. 2011;187(12):6217–26. doi: 10.4049/jimmunol.1003478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schaefer L, Iozzo RV. Small leucine-rich proteoglycans, at the crossroad of cancer growth and inflammation. Curr Opin Genet Dev. 2012;22(1):56–7. doi: 10.1016/j.gde.2011.12.002. [DOI] [PubMed] [Google Scholar]
- 14.Schaefer L, Babelova A, Kiss E, Hausser HJ, Baliova M, Krzyzankova M, et al. The matrix component biglycan is proinflammatory and signals through Toll-like receptors 4 and 2 in macrophages. J Clin Invest. 2005;115(8):2223–33. doi: 10.1172/JCI23755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Moreth K, Iozzo RV, Schaefer L. Small leucine-rich proteoglycans orchestrate receptor crosstalk during inflammation. Cell Cycle. 2012;11(11):2084–91. doi: 10.4161/cc.20316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Meng X, Ao L, Song Y, Babu A, Yang X, Wang M, et al. Expression of functional Toll-like receptors 2 and 4 in human aortic valve interstitial cells: potential roles in aortic valve inflammation and stenosis. Am J Physiol Cell Physiol. 2008;294(1):C29–C35. doi: 10.1152/ajpcell.00137.2007. [DOI] [PubMed] [Google Scholar]
- 17.Derbali H, Bosse Y, Cote N, Pibarot P, Audet A, Pepin A, et al. Increased biglycan in aortic valve stenosis leads to the overexpression of phospholipid transfer protein via Toll-like receptor 2. Am J Pathol. 2010;176(6):2638–45. doi: 10.2353/ajpath.2010.090541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stephens EH, Saltarrelli JG, Baggett LS, Nandi I, Kuo JJ, Davis AR, et al. Differential proteoglycan and hyaluronan distribution in calcified aortic valves. Cardiovasc Pathol. 2010;20(6):334–42. doi: 10.1016/j.carpath.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Messier RH, Jr., Bass BL, Aly HM, Jones JL, Domkowski PW, Wallace RB, et al. Dual structural and functional phenotypes of the porcine aortic valve interstitial population: characteristics of the leaflet myofibroblast. J Surg Res. 1994;57(1):1–21. doi: 10.1006/jsre.1994.1102. [DOI] [PubMed] [Google Scholar]
- 20.Moresco EM, LaVine D, Beutler B. Toll-like receptors. Curr Biol. 2011;21(13):R488–93. doi: 10.1016/j.cub.2011.05.039. [DOI] [PubMed] [Google Scholar]
- 21.Zeng Q, Song R, Ao L, Weyant MJ, Lee J, Xu D, et al. Notch1 promotes the pro-osteogenic response of human aortic valve interstitial cells via modulation of ERK1/2 and nuclear factor-kappaB activation. Arterioscler Thromb Vasc Biol. 2013;33(7):1580–90. doi: 10.1161/ATVBAHA.112.300912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yang X, Fullerton DA, Su X, Ao L, Cleveland JC, Meng X. Pro-osteogenic phenotype of human aortic valve interstitial cells is associated with higher levels of Toll-like receptors 2 and 4 and enhanced expression of bone morphogenetic protein 2. J Am Coll Cardiol. 2009;53:491–500. doi: 10.1016/j.jacc.2008.09.052. [DOI] [PubMed] [Google Scholar]
- 23.Stewart BF, Siscovick D, Lind BK, Gardin JM, Gottdiener JS, Smith VE, et al. Clinical factors associated with calcific aortic valve disease. Cardiovascular Health Study. J Am Coll Cardiol. 1997;29(3):630–4. doi: 10.1016/s0735-1097(96)00563-3. [DOI] [PubMed] [Google Scholar]
- 24.Song R, Zeng Q, Ao L, Yu JA, Cleveland JC, Zhao KS, et al. Biglycan Induces the Expression of Osteogenic Factors in Human Aortic Valve Interstitial Cells via Toll-Like Receptor-2. Arterioscler Thromb Vasc Biol. 2012;32:2711–2720. doi: 10.1161/ATVBAHA.112.300116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Babelova A, Moreth K, Tsalastra-Greul W, Zeng-Brouwers J, Eickelberg O, Young MF, et al. Biglycan, a danger signal that activates the NLRP3 inflammasome via toll-like and P2X receptors. J Biol Chem. 2009;284(36):24035–48. doi: 10.1074/jbc.M109.014266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schaefer L, Macakova K, Raslik I, Micegova M, Grone HJ, Schonherr E, et al. Absence of decorin adversely influences tubulointerstitial fibrosis of the obstructed kidney by enhanced apoptosis and increased inflammatory reaction. Am J Pathol. 2002;160(3):1181–91. doi: 10.1016/S0002-9440(10)64937-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.de Kluijver J, Schrumpf JA, Evertse CE, Sont JK, Roughley PJ, Rabe KF, et al. Bronchial matrix and inflammation respond to inhaled steroids despite ongoing allergen exposure in asthma. Clin Exp Allergy. 2005;35(10):1361–9. doi: 10.1111/j.1365-2222.2005.02334.x. [DOI] [PubMed] [Google Scholar]
- 28.Otto CM, Kuusisto J, Reichenbach DD, Gown AM, O'Brien KD. Characterization of the early lesion of ‘degenerative’ valvular aortic stenosis. Histological and immunohistochemical studies. Circulation. 1994;90(2):844–53. doi: 10.1161/01.cir.90.2.844. [DOI] [PubMed] [Google Scholar]
- 29.Osman L, Yacoub MH, Latif N, Amrani M, Chester AH. Role of human valve interstitial cells in valve calcification and their response to atorvastatin. Circulation. 2006;114(1 Suppl):I547–52. doi: 10.1161/CIRCULATIONAHA.105.001115. [DOI] [PubMed] [Google Scholar]
- 30.Kaden JJ, Kilic R, Sarikoc A, Hagl S, Lang S, Hoffmann U, et al. Tumor necrosis factor alpha promotes an osteoblast-like phenotype in human aortic valve myofibroblasts: a potential regulatory mechanism of valvular calcification. Int J Mol Med. 2005;16(5):869–72. [PubMed] [Google Scholar]
- 31.Oak S, Mandrekar P, Catalano D, Kodys K, Szabo G. TLR2- and TLR4-mediated signals determine attenuation or augmentation of inflammation by acute alcohol in monocytes. J Immunol. 2006;176(12):7628–35. doi: 10.4049/jimmunol.176.12.7628. [DOI] [PubMed] [Google Scholar]
- 32.Hu X, Chen J, Wang L, Ivashkiv LB. Crosstalk among Jak-STAT, Toll-like receptor, and ITAM-dependent pathways in macrophage activation. J Leukoc Biol. 2007;82(2):237–43. doi: 10.1189/jlb.1206763. [DOI] [PubMed] [Google Scholar]
- 33.Huang S, Rutkowsky JM, Snodgrass RG, Ono-Moore KD, Schneider DA, Newman JW, et al. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J Lipid Res. 2012;53(9):2002–13. doi: 10.1194/jlr.D029546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dillon S, Agrawal A, Van Dyke T, Landreth G, McCauley L, Koh A, et al. A Toll-like receptor 2 ligand stimulates Th2 responses in vivo, via induction of extracellular signal-regulated kinase mitogen-activated protein kinase and c-Fos in dendritic cells. J Immunol. 2004;172(8):4733–43. doi: 10.4049/jimmunol.172.8.4733. [DOI] [PubMed] [Google Scholar]
- 35.Medvedev AE, Kopydlowski KM, Vogel SN. Inhibition of lipopolysaccharide-induced signal transduction in endotoxin-tolerized mouse macrophages: dysregulation of cytokine, chemokine, and Toll-like receptor 2 and 4 gene expression. J Immunol. 2000;164:5564–5574. doi: 10.4049/jimmunol.164.11.5564. [DOI] [PubMed] [Google Scholar]
- 36.Lehmann S, Walther T, Kempfert J, Rastan A, Garbade J, Dhein S, et al. Mechanical strain and the aortic valve: influence on fibroblasts, extracellular matrix, and potential stenosis. Ann Thorac Surg. 2009;88(5):1476–83. doi: 10.1016/j.athoracsur.2009.07.025. [DOI] [PubMed] [Google Scholar]