

Abstract
LexA and two structurally related regulators, PrtR and PA0906, coordinate the Pseudomonas aeruginosa SOS response. RecA-mediated autocleavage of LexA induces the expression of a protective set of genes that increase DNA damage repair and tolerance. In contrast, RecA-mediated autocleavage of PrtR induces antimicrobial pyocin production and a program that lyses cells to release the newly synthesized pyocin. Recently, PrtR-regulated genes were shown to sensitize P. aeruginosa to quinolones, antibiotics that elicit a strong SOS response. Here, we investigated the mechanisms by which PrtR-regulated genes determine antimicrobial resistance and genotoxic stress survival. We found that induction of PrtR-regulated genes lowers resistance to clinically important antibiotics and impairs the survival of bacteria exposed to one of several genotoxic agents. Two distinct mechanisms mediated these effects. Cell lysis genes that are induced following PrtR autocleavage reduced resistance to bactericidal levels of ciprofloxacin, and production of extracellular R2 pyocin was lethal to cells that initially survived UV light treatment. Although typically resistant to R2 pyocin, P. aeruginosa becomes transiently sensitive to R2 pyocin following UV light treatment, likely because of the strong downregulation of lipopolysaccharide synthesis genes that are required for resistance to R2 pyocin. Our results demonstrate that pyocin production during the P. aeruginosa SOS response carries both expected and unexpected costs.
INTRODUCTION
Bacteria respond to DNA damage and replication fork stress by activating the SOS response. Much of our understanding of the SOS response stems from seminal research with Escherichia coli, where this response functions primarily to increase the cell's ability to cope with DNA damage and replication stress (1, 2). The SOS response of E. coli is coordinated by the LexA protein, which directly represses the expression of genes that bear a LexA-binding motif. During growth under nonstressed conditions, LexA keeps the expression of target genes at a minimum level. Upon DNA damage and/or replication fork stalling, the nucleoprotein filament formed by RecA binding to single-stranded DNA stimulates the latent serine protease of LexA, resulting in LexA autocleavage and derepression of LexA-regulated genes. These LexA-regulated genes code for proteins that enhance nucleotide excision repair, DNA damage tolerance, recombinational repair, and cell cycle control (1, 2).
Autocleavage of LexA during the SOS response can also derepress genes on accessory plasmids, including genes that code for translesion polymerases, antibiotic resistance determinants, and antimicrobial colicins (3–6). Colicins are particularly interesting from the viewpoint of the SOS response because induced colicin production causes cell death, a stark contrast to the protective effects of chromosomal genes regulated by LexA. Colicin synthesis causes death because a coexpressed protein mediates cell lysis (6). Colicins are intraspecies antimicrobial bacteriocins that target nonrelated individuals. Related individuals are immune to colicin because of an immunity protein encoded by a plasmid.
The SOS response in Pseudomonas aeruginosa is more complex than that of E. coli, as it is coordinated not only by LexA but also by two structurally related regulators, PrtR and PA0906 (7–11). These regulators are encoded by the chromosome, and they are found in most P. aeruginosa strains and in related Pseudomonas spp. (12). As in E. coli, DNA damage leads to RecA-mediated autocleavage of LexA and derepression of the protective LexA regulon (8, 11, 13). RecA single-stranded DNA nucleoprotein filaments also stimulate the latent serine protease of PrtR and PA0906, resulting in autocleavage and derepression of their target genes (8, 14–16). PrtR directly and indirectly controls the expression of 43 genes involved in the synthesis and release of antimicrobial pyocins, and PA0906 is predicted to control 6 genes whose functions have not yet been elucidated (8, 14).
Pyocins are antimicrobial bacteriocins that target other strains of the same species, although some pyocin types have been found to target other species in addition to P. aeruginosa (14). However, pyocin production comes with a cost, because cells that produce pyocin during the SOS response lyse and die. Pyocin has been reported to accumulate to low levels in standard cultures after overnight incubation (17), and the addition of genotoxic agents such as mitomycin C (MMC) during the exponential phase strongly induces pyocin production (14, 17). Pyocin production is initiated during the SOS response when PrtR autocleavage derepresses the expression of prtN. In turn, PrtN binds to the P-box in the promoters of R-, F-, and S-type pyocin genes to activate their expression (7, 18). In addition to pyocin biosynthetic genes, PrtN activates the expression of a lysis cassette coding for several proteins, including holin-like and chitinase-like enzymes (7). After pyocin synthesis, the lysis cassette mediates cell lysis, killing the cell and releasing pyocin into the environment.
P. aeruginosa infections are often treated with fluoroquinolone antibiotics that inhibit DNA gyrase and topoisomerase IV and strongly induce the SOS response and pyocin production. Research has shown that PrtR-regulated genes increase the sensitivity of P. aeruginosa to bacteriostatic and bactericidal concentrations of ciprofloxacin (9, 19). It has been hypothesized that in wild-type cells, pyocin synthesis makes cells more susceptible to fluoroquinolones because cell lysis enzymes that accompany pyocin production mediate death (9, 19). Whether the production of pyocins influences the sensitivity of P. aeruginosa to other genotoxic agents is not known.
In this study, we investigated how induced expression of PrtR-regulated genes determines antimicrobial resistance and genotoxic stress survival. We created a strain in which pyocin synthesis genes were no longer induced along with LexA and PA0906 target genes during the SOS response and found that the absence of pyocin production and cell lysis increased resistance to several antibiotics and enhanced the survival of bacteria exposed to one of several genotoxic agents. Further genetic experiments supported the hypothesis that the death of ciprofloxacin-treated wild-type cells was mediated, in part, by cell lysis genes. In addition, we discovered that cell lysis and R2 pyocin were both required to mediate the death of UV-treated cells. Surprisingly, although P. aeruginosa is typically resistant to R2 pyocin, we found that UV light-treated P. aeruginosa transiently loses its R2 pyocin resistance, most likely because of the strong and rapid downregulation of genetic determinants of R2 pyocin susceptibility during the SOS response. This created a situation where cells that initially survived exposure to UV irradiation were killed by extracellular R2 pyocin that was produced by some cells during their SOS response. Thus, the induced production of pyocins during the P. aeruginosa SOS responses carries both expected and unexpected costs. We discuss the implications of these findings for the SOS response of P. aeruginosa and related bacteria.
MATERIALS AND METHODS
Bacterial strains and plasmids.
All of the bacterial strains used in this study are listed in Table 1. Constructs used to genetically manipulate strains were generated by PCR-based methods. Unmarked deletions and point mutations were generated in the wild-type PAO1 strain by using the pEX18GM vector and previously published methods (20). Strains with the prtRS162A and PA0906S153A mutations were created by replacing the codon of the catalytic serine residue with the codon for an alanine residue. The lysis-defective strain Δ0614 Δ0629 was constructed by deleting codons 8 to 142 and 12 to 203 of PA0614 and PA0629, respectively. ΔS5 (PA0985) was generated by deleting codons 102 to 466. For complementation of the Δ0614 Δ0629 mutant strain, a construct consisting of the 575 bp upstream of the PA0614 translation start site, the PA0614 coding region, an in-frame fusion of the first 110 codons of PA0615 with the last 87 codons of PA0628, and the PA0629 coding region was cloned into mini-CTX2 and then integrated into the chromosome at the neutral attB site according to previously published methods (21). A PA0614-lacZ translational reporter gene was constructed by fusing the 575 bp upstream of the PA0614 translation start site plus three codons to promoterless lacZ in mini-CTX2 (22). This reporter gene was then integrated into the chromosome according to previously published methods (21).
TABLE 1.
Bacterial strains and plasmids used in this study
| P. aeruginosa strain or plasmid | Relevant characteristic(s) | Source or reference |
|---|---|---|
| Strains | ||
| PAO1 | Wild type (parent strain) | B. Iglewski |
| prtRS162A mutant | Endogenous prtR replaced with prtRS162A, producing proteolytically inactive transcriptional regulator | This study |
| prtRS162A PA0906S153A mutant | Endogenous PA0906 replaced with PA0906S153A, producing proteolytically inactive transcriptional regulator | This study |
| prtRS162A lexAG86V mutant | LexAG86V resistant to autoproteolytic cleavage | This study |
| prtRS162A lexAG86V PA0906S153A mutant | Wild-type alleles replaced | This study |
| S2-mutant | S2 pyocin transposon insertion mutant with PA1150-H12::ISlacZ/hah allele from PW3083, Tcr | 53 |
| S4-mutant | S4 pyocin transposon insertion mutant with PA3866-F01::ISlacZ/hah allele from PW7522, Tcr | 53 |
| F2-mutant | F2 pyocin transposon insertion mutant with PA0633-E12::ISlacZ/hah allele from PW2150 | 53 |
| ΔS5 mutant | Unmarked deletion of PA0985 | This study |
| R2-mutant | R1 pyocin transposon insertion mutant with PA0625-H05::ISlacZ/hah allele from PW2138, Tcr | 53 |
| ΔR2 mutant | Unmarked deletion of PA0615-PA0628 | 32 |
| ΔPA0614 ΔPA0629 mutant | Clean deletion of codons 8-142 and 12-203 from PA0614 and PA0629, respectively | This study |
| ΔPA0614/+ ΔPA0629/+ mutant | Genetic complementation of lysis, PA0614p::PA0614-PA0629 placed at attB site, Tcr | This study |
| PML1516d | Indicator for pyocin S1, S2, and AP41 | 25 |
| NIH5 | Indicator for pyocin F1, F2, and F3 | 25 |
| 13s | Indicator strain for R1 to R5 pyocins | 24 |
| Plasmids | ||
| PA0614-lacZ | PA0614-lacZ translational fusion in CTX2, Tcr | This study |
| recAp::GFP | recA promoter in front of promoterless gene for GFP in pUCP18 | 37 |
Bacterial growth conditions and treatment.
Bacteria were grown at 37°C in LB on a roller drum or on LB agar unless otherwise noted. Log-phase cultures with an optical density at 600 nm of 0.5 to 0.6 were used for zone-of-inhibition assays to determine ciprofloxacin (2 μg/ml) and mitomycin (15 μg/ml) susceptibility, UV irradiation experiments, and profiling reporter gene expression during the SOS response. The drug concentrations used for zone-of-inhibition assays were as follows: ciprofloxacin, 5 μg; paraquat, 10 μl of a 100 mM solution; H2O2, 10 μl of a 6% solution; mitomycin, 20 μg. UV treatment was done by using a germicidal lamp (General Electric) to irradiate serial dilutions of cells spotted onto agar plates. In experiments with R2 pyocin and UV light treatment, 1 μl of purified R2 pyocin was mixed with 10 μl of each serial dilution prior to spotting onto agar plates. MICs were determined in Mueller-Hinton broth as previously described (23).
Measurement of reporter gene expression.
Expression of PA0614-lacZ was induced by the addition of 1 μg/ml of ciprofloxacin to broth cultures, and LacZ activity was measured with the Galacto-Light kit at the indicated times as previously described (22). Expression of recAp::GFP was induced by the addition of ciprofloxacin (1 μg/ml) to cultures grown in tryptic soy broth, and GFP fluorescence was quantified at the times indicated.
Pyocin induction, isolation, and assays.
Pyocin was induced in exponential-phase cultures as previously described (24). Briefly, 3 μg/ml of MMC was added to a culture with an optical density at 600 nm of 0.25 to 0.3 and the culture was further incubated for 3 to 4 h unless noted otherwise. The culture was then treated for 30 min with DNase I (2 U/ml) to reduce its viscosity. Nonlysed cells and debris from lysed cells were pelleted by centrifugation, and the resulting supernatant containing all of the pyocin types was either used to purify R2 pyocin as previously described (24) or filter sterilized and spotted onto a lawn of the pyocin indicator strain (24, 25). To generate a lawn of the indicator strain, a 100-μl solution of a 1,000-fold dilution of a saturated culture was plated and allowed to dry prior to the addition of pyocin. To detect the activity of pyocin that accumulated in saturated cultures after overnight incubation, cultures were centrifuged to pellet cells and the supernatant was filter sterilized and applied to the indicator strains. Agar plates containing indicator strains were then incubated overnight. Clearing in the lawn corresponding to the spotted pyocin preparation is indicative of pyocin activity. Indicator strain 13s is sensitive to only R2 pyocin produced by wild-type PAO1 (Table 1; see Fig. S1A in the supplemental material) (24). Indicator strain PML1516d is sensitive to S2 pyocin in addition to one or more other pyocins produced by wild-type PAO1 (Table 1; see Fig. S1A) (25, 26). Indicator strain NIH5 is sensitive to F2 pyocin in addition to possibly one or more of the other pyocins produced by wild-type PAO1 (Table 1; see Fig. S1A) (25, 27).
Quantitative PCR (qPCR).
UV-treated cells were collected from the agar plate at the indicated times and mixed 1:2 (vol/vol) with RNAprotect Bacteria Reagent (Qiagen). Fixed cells were then pelleted and lysed in Tris-EDTA buffer (pH 8.0) containing 15 mg/ml of lysozyme and 15 μl of proteinase K (Qiagen). RNA was then isolated with the RNeasy Plus minikit (Qiagen).
cDNA for qPCR experiments was generated as previously described (28). qPCR analysis of cDNA of the indicated genes and the internal control gene proC (29) was performed with Power SYBR green PCR master mix (Life Technologies) on a StepOnePlus real-time PCR machine (Life Technologies). The 2−ΔΔCT method was used to determine the expression levels of the indicated genes relative to that of proC. Relative gene expression data from three biological replicates were normalized, log transformed, mean centered, and autoscaled (30). The sequences of the oligonucleotide primers used are available upon request.
RESULTS
Autoproteolytic activity of PrtR is required for induction of pyocins during standard culture and genotoxic stress.
The autoproteolytic cleavage of PrtR that is mediated by RecA during DNA damage or replication stress is likely critical for pyocin production in P. aeruginosa (15, 18). To better understand how pyocins affect genotoxic stress resistance, we constructed a prtRS162A mutant in order to unlink pyocin production from the LexA- and PA0906-regulated components of the RecA-mediated SOS response. The prtRS162A mutation replaces the codon for the catalytic serine residue of the PrtR autoprotease domain with a codon for a catalytically inactive alanine residue. This strategy has been widely used to inactivate serine proteases, including the serine autoprotease of LexA (8, 31). We verified that the expression of PA0614p::lacZ, the first gene of the R2 pyocin biosynthetic operon (7), was not induced in prtRS162A mutant cells exposed to ciprofloxacin (Fig. 1A). We also demonstrated that MMC treatment failed to elicit pyocin production in exponential-phase prtRS162A mutant cultures, as MMC treatment does to wild-type cultures (Fig. 1B; see Fig. S1A in the supplemental material) (14, 17, 18, 24). In contrast, induced expression of the LexA-regulated recAp::GFP reporter gene was normal in prtRS162A mutant cells exposed to genotoxic stress (Fig. 1C), which showed that the prtRS162A mutation had no effect on the induction of the LexA regulon during the SOS response.
FIG 1.
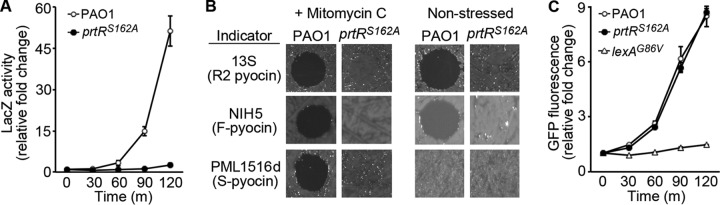
Autoproteolytic activity of PrtR is required for pyocin production. (A) Expression of the PA0614p::lacZ translational reporter gene in the prtRS162A mutant and the parent strain treated with 1 μg/ml ciprofloxacin. Error bars indicate standard deviations. (B) Production of pyocin by the prtRS162A mutant and the parent strain. Filtered supernatant from cultures treated with MMC (3 μg/ml) and from liquid cultures incubated overnight were spotted onto lawns of the indicator strains 13s, PML1516d, and NIH5. Clearing of the bacterial lawn is indicative of pyocin-mediated death (see Materials and Methods; see Fig. S1A in the supplemental material for information on the specificity of indicator strains for different pyocin types). (C) Expression of the recAp::GFP transcriptional reporter gene in the prtRS162A mutant, the lexAG86V mutant, and the parent strain treated with 1 μg/ml ciprofloxacin. LexAG86V protein is resistant to autoproteolytic cleavage (37). Error bars indicate standard deviations.
Although pyocin production in exponential-phase cultures is strongly induced by genotoxic agents (14, 17, 18, 24), observations by a number of groups have shown that P. aeruginosa accumulates low levels of R pyocin in cultures incubated overnight under nonstressed conditions (17, 32, 33). However, it has not been clear whether this accumulation of pyocin under nonstressed conditions is due to PrtR autoproteolysis. We used our prtRS162A mutant strain to determine if the accumulation of pyocin during standard culture under nonstressed conditions is dependent on PrtR autoproteolysis and found no detectable pyocin activity in the supernatant of prtRS162A mutant cultures (Fig. 1B). Importantly, the wild-type and prtRS162A mutant strains exhibited similar growth dynamics and similar cell yields after overnight incubation (see Fig. S2 in the supplemental material). Thus, PrtR autoproteolysis is required for the production and accumulation of pyocin under standard culture conditions, as well as for pyocin production during genotoxic stress.
Pyocin production affects resistance to antibiotics and genotoxic agents.
Previous research showed that transposon insertion mutations in the P. aeruginosa pyocin biosynthetic locus increase resistance to ciprofloxacin (9, 19). These studies have proposed that the sensitivity of a wild-type strain is due in part to genotoxic stress eliciting the induction of pyocin production and cell lysis proteins. Consistent with this idea, we found that the prtRS162A mutant strain was more resistant than the wild type to bacteriostatic (see Table S1 and Fig. S3A in the supplemental material) and bactericidal (Fig. 2A) levels of ciprofloxacin. To better understand the role of pyocin production in genotoxic stress resistance, we determined how the prtRS162A mutation affects resistance to other genotoxic agents. In addition, we explored whether the prtRS162A mutation affects resistance to other antimicrobial stresses.
FIG 2.
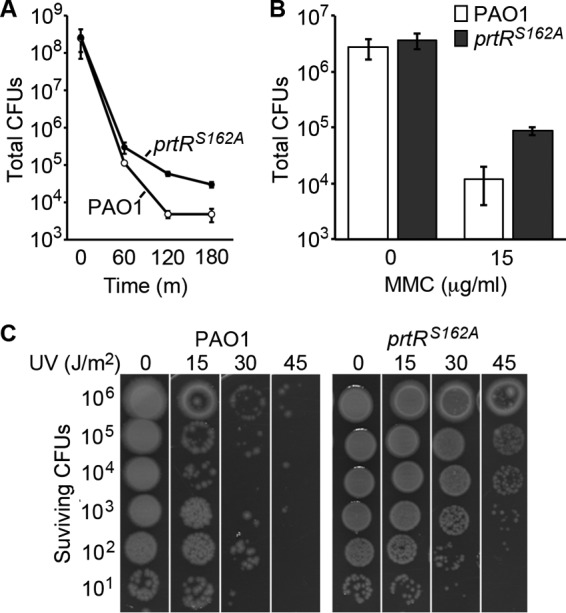
Autoproteolytic activity of PrtR reduces survival during genotoxic stress. (A) Total CFU counts in cultures of wild-type PAO1 and the prtRS162A mutant treated with a bactericidal concentration of ciprofloxacin (2 μg/ml). Error bars indicate standard deviations. (B) Total CFU counts of wild-type PAO1 and the prtRS162A mutant on agar plates containing 0 and 15 μg/ml of MMC after overnight incubation. Error bars indicate standard deviations. (C) Surviving CFU counts of wild-type PAO1 and the prtRS162A mutant after UV treatment. Cultures were serially diluted 10-fold, spotted onto agar, and treated with UV light at the indicated doses. Surviving CFU counts were determined after overnight incubation.
We focused on the genotoxic agents MMC and UV light because both of them had previously been shown to induce pyocin production (14). We found that the prtRS162A mutant strain was slightly more resistant to MMC than the parent strain in zone-of-inhibition assays (see Fig. S3B in the supplemental material) and that the prtRS162A mutant strain exhibited a modest improvement in survival on agar containing bactericidal concentrations of MMC (Fig. 2B). To compare the survival of wild-type PAO1 and prtRS162A mutant cells exposed to UV light, we treated serial dilutions of each strain spotted onto agar plates with increasing amounts of UV radiation and incubated the agar plates overnight to determine the level of survival. We observed a striking increase in the survival of the prtRS162A mutant relative to that of the wild-type parent strain (Fig. 2C). Treatment of wild-type P. aeruginosa with UV light (30 J/m2) reduced the CFU count by 5 orders of magnitude, whereas the CFU count of the prtRS162A mutant strain was reduced by only 1 order of magnitude (Fig. 2C). Thus, blocking pyocin production in P. aeruginosa improves survival under conditions of genotoxic stress, such as UV irradiation treatment.
The relationship between pyocin production and resistance to other antimicrobial agents proved to be complex. We found that while the prtRS162A mutant and the wild-type strain were similarly sensitive to growth inhibition by carbenicillin and tetracycline, the prtRS162A mutant strain exhibited a 2-fold higher MIC of every aminoglycoside antibiotic tested (see Table S1 in the supplemental material). The prtRS162A mutant strain was also more resistant than the wild-type strain to paraquat, a superoxide generator (see Fig. S3C in the supplemental material). This increase in paraquat resistance was not due to an enhanced generalized oxidative stress response because prtRS162A mutant cells exhibited a normal level of sensitivity to hydrogen peroxide (see Fig. S3D). It could be that under our conditions, aminoglycoside and paraquat treatment of P. aeruginosa elicited an SOS response, as occurs in aminoglycoside-treated Vibrio cholerae (34, 35) and paraquat-treated E. coli (36), and that pyocin production has a negative effect. However, arguing against this possibility is the observation that P. aeruginosa treated with hydrogen peroxide activates an SOS response and induces pyocin gene synthesis (10). An alternative explanation for the role of PrtR in aminoglycoside and paraquat resistance is that the basal expression of genes regulated directly and indirectly by PrtR could have cryptic functions that impact aminoglycoside and paraquat resistance.
PrtR-regulated lysis is a determinant of ciprofloxacin resistance and UV irradiation survival.
We further pursued a mechanistic understanding of how pyocin production affects P. aeruginosa genotoxic stress resistance. Prior work has shown that RecA-mediated PrtR autoproteolysis derepresses prtN expression, and PrtN then massively upregulates pyocin gene expression (18, 19). In addition, PrtN activates the expression of a lysis cassette that mediates cellular lysis and releases the newly synthesized pyocin into the environment (7). Previous research hypothesized that induction of this lysis cassette sensitizes P. aeruginosa to genotoxic stress (9). Indeed, artificial overexpression of these lysis genes reduces cell viability (30). We tested this hypothesis by assaying the sensitivity of a lysis-impaired strain to UV irradiation and ciprofloxacin.
We constructed a lysis-impaired strain by deleting the holin-like PA0614 and lysozyme-like PA0629 genes thought to mediate lysis (7) and confirmed that the Δ0614 Δ0629 mutant strain produced less extracellular pyocin than wild-type cells during genotoxic stress (Fig. 3A). We found that, relative to the wild type, the Δ0614 Δ0629 mutant strain exhibited a striking increase in survival following UV irradiation treatment (Fig. 3B). Genetic complementation of the mutations in the Δ0614 Δ0629 mutant strain restored extracellular pyocin production (Fig. 3A) and killing by UV irradiation to levels near those of the wild-type strain (Fig. 3B). In zone-of-inhibition assays, the level of ciprofloxacin resistance of the Δ0614 Δ0629 mutant strain was intermediate between those of the wild type (more sensitive) and the prtRS162A mutant strain (more resistant) (see Fig. S4 in the supplemental material). However, Δ0614 Δ0629 and prtRS162A mutant cells were equally resistant to bactericidal concentrations of ciprofloxacin (Fig. 3C). These results indicate that PA0614 and PA0629 are genetic determinants of genotoxic stress sensitivity and support the hypothesis that PrtR-regulated cell lysis sensitizes P. aeruginosa to genotoxic agents.
FIG 3.
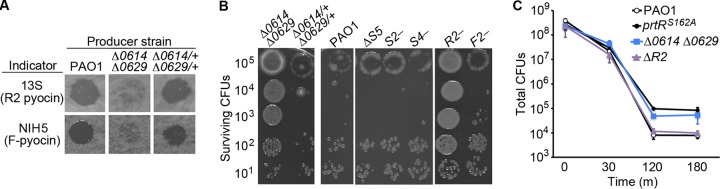
PrtR-regulated lysis and R2 pyocin genes are determinants of UV irradiation survival. (A) Production of extracellular pyocin in cultures treated with 3 μg/ml of MMC for 2 h. Clearing of the bacterial lawn of the indicator strains is indicative of pyocin-mediated death. (B) Surviving CFU counts of the lysis-defective Δ0614 Δ0629 mutant strain and pyocin mutant strains after UV treatment (30 J/m2). Cultures were serially diluted 10-fold, spotted onto agar, and treated with UV light. Surviving CFU counts were determined after overnight incubation. S2-, PA1150-H12::ISlacZ/hah allele; S4-, PA3866-F01::ISlacZ/hah allele; F2-, PA0633-E12::ISlacZ/hah allele; R1-, PA0625-H05::ISlacZ/hah allele. (C) Total CFU counts in cultures of wild-type PAO1 and prtRS162A, Δ0614 Δ0629, and ΔR2 mutant cells treated with a bactericidal concentration of ciprofloxacin (2 μg/ml). Error bars indicate standard deviations.
R2 pyocin is a strong determinant of UV irradiation survival.
The marked effect of the prtRS162A and Δ0614 Δ0629 mutations on UV irradiation survival led us to wonder whether a particular toxic pyocin might also contribute to the high level of cell death observed in the wild-type strain following UV treatment. We tested for this possibility by assaying the UV irradiation survival of pyocin mutants. Strikingly, we found that a transposon insertion in the R2 pyocin biosynthetic locus, but not in genes that synthesize S2, S4, S5, and F2 pyocins, substantially increased survival after UV irradiation treatment (Fig. 3B). An R2 pyocin deletion mutant exposed to UV light also exhibited a striking increase in resistance to killing by UV irradiation similar to that of the prtRS162A mutant (Fig. 4A below). Taken together, these observations indicate that the poor survival of P. aeruginosa treated with UV light requires both R2 pyocin production and cell lysis functions.
FIG 4.
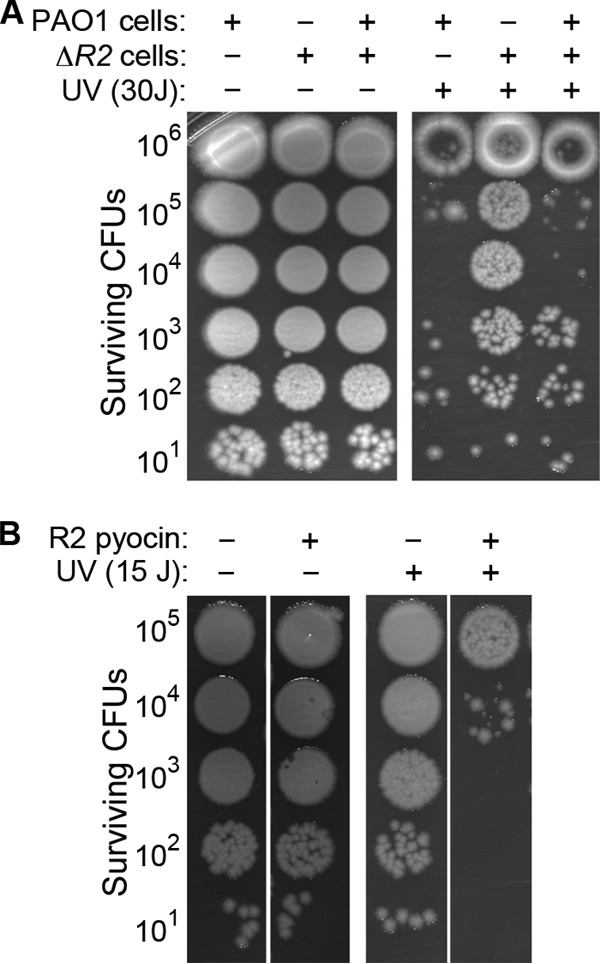
Extracellular R2 pyocin is lethal to cells that initially survive UV treatment. (A) Survival of the wild type, the ΔR2 mutant, and a 1:1 mixture of the wild type and the ΔR2 mutant after UV treatment (30 J/m2). (B) Survival of UV-irradiated ΔR2 mutant cells with or without purified R2 pyocin. Cultures were serially diluted 10-fold, mixed 1:1 with saline or saline with purified R2 pyocin, and spotted onto agar prior to UV treatment. Surviving CFU counts were determined after overnight incubation. Note that ΔR2 mutant cells not treated with UV light were resistant to purified R2 pyocin.
We also assayed the ciprofloxacin resistance of the ΔR2 mutant strain and found that ΔR2 mutant cells exposed to a bactericidal concentration of ciprofloxacin did not exhibit an increase in survival (Fig. 3C). Thus, R2 pyocin synthesis genes have little to no effect on ciprofloxacin resistance in wild-type P. aeruginosa.
Self-produced R2 pyocin kills P. aeruginosa after UV irradiation.
How could R2 pyocin synthesis and cell lysis reduce the survival of UV-irradiated P. aeruginosa? R2 pyocin is an antimicrobial that targets other strains, and typically the producer strain is resistant (14). However, recent research showed that self-produced R2 pyocin can be lethal under certain conditions (32), raising the possibility that R2 pyocin produced by some cells during their SOS response was toxic to the remaining population that survived UV light treatment. We tested for this possibility by using two approaches. In one approach, we mixed ΔR2 mutant cells with wild-type cells prior to treatment with UV irradiation. We found that the addition of wild-type cells significantly reduced the survival of the ΔR2 mutant strain (Fig. 4A), indicating that R2 pyocin produced and released in response to UV irradiation can act in trans between cells to kill other members of the UV-irradiated population. In the second approach, we directly added purified R2 pyocin to the serial dilutions of ΔR2 mutant cells prior to treatment with UV light and found that purified R2 pyocin also reduced the survival of ΔR2 mutant cells (Fig. 4B). Importantly, purified R2 pyocin had no effect on the growth and survival of nontreated control cells (Fig. 4B). These results indicate that self-produced R2 pyocin was lethal to P. aeruginosa cells that survived UV light exposure.
Sensitivity to R2 pyocin is a transient phenotype not controlled by PrtR, PA0906, and LexA.
Colonies that form from ΔR2 mutant and wild-type cells that survive UV irradiation are R2 pyocin resistant, indicating that the loss of R2 pyocin resistance following UV light treatment is a transient physiological response. We tested whether induction of the LexA, PA0906, and PrtR regulons during the P. aeruginosa SOS response mediates the loss of R2 pyocin resistance. We used the PA0906S153A, lexAG86V, and prtRS162A mutations to create double and triple mutants in which two or three regulons of the SOS response could not be induced during genotoxic stress. The LexAG86V protein is resistant to autoproteolytic cleavage, and the LexA regulon cannot be derepressed in a lexAG86V mutant (37). Importantly, none of these mutations had an effect on the R2 pyocin resistance of P. aeruginosa under nonstress conditions (see Fig. S1B in the supplemental material). Following UV light exposure, we observed R2 pyocin-sensitive cells in double mutants (prtRS162A PA0906S153A and prtRS162A lexAG86V mutants) and in a triple mutant (PA0906S153A lexAG86V prtRS162A mutant), indicating that induction of genes regulated by PrtR, PA0906, and LexA is not required for the loss of R2 pyocin resistance after UV irradiation treatment in P. aeruginosa cells (Fig. 5). Intriguingly, these data suggest that an as-yet-unidentified system that regulates pyocin resistance is activated during the SOS response in P. aeruginosa.
FIG 5.
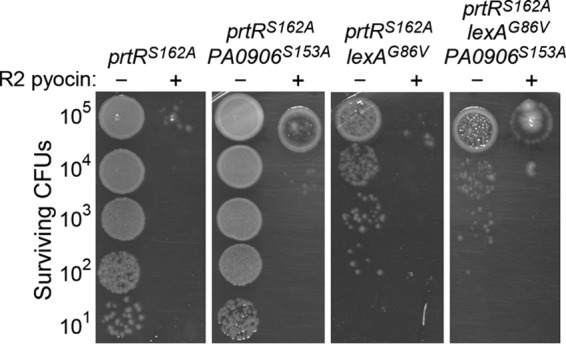
Loss of R2 pyocin resistance after UV treatment is not regulated by RecA-stimulated cleavage of PrtR, LexA, and PA0906 during the SOS response. Cultures were serially diluted 10-fold, mixed 1:1 with saline or saline with purified R2 pyocin, and spotted onto agar prior to treatment with UV (10 J/m2). Surviving CFU counts were determined after overnight incubation.
Loss of R2 pyocin resistance after UV irradiation correlates with B-band LPS downregulation.
In P. aeruginosa, the B-band O-antigen confers resistance to R2 pyocin by masking a receptor that lies within the core polysaccharide of lipopolysaccharide (LPS) (32, 33). Mutations that eliminate B-band O-antigen synthesis and unmask this receptor have been shown to confer sensitivity to self-produced R2 pyocin (32). We hypothesized that in UV-irradiated P. aeruginosa, the R2 pyocin receptor was transiently unmasked and therefore became accessible to self-produced R2 pyocin. We tested this hypothesis by profiling the expression of genes involved in each distinct step of LPS synthesis in nontreated and UV light-treated ΔR2 mutant cells. Strikingly, we found that the B-band O-antigen synthesis genes wbpJ, wbpA, and wzx were rapidly and significantly downregulated 30 min after UV treatment (Fig. 6A). At 60 min after UV treatment, the expression of wbpJ and wzm was downregulated ∼10-fold and that of wbpA was downregulated 5-fold relative to their expression in nontreated control cultures (Fig. 6A). In contrast, the expression of the A-band O-antigen synthesis genes wbpZ and wzm was unchanged (Fig. 6A). The expression of genes involved in lipid A (lpxA) and the core polysaccharide (waaP) also decreased following UV irradiation (Fig. 6B), although not as rapidly as that of B-band LPS biosynthesis genes. These data clearly show a surprising differential response in the expression of B-band and A-band O-antigen biosynthesis genes following UV treatment.
FIG 6.

Relative expression of B-band and A-band O-antigen synthesis genes after treatment of wild-type cells with UV (15 J/m2). (A) Relative expression of A-band synthesis genes (wbpZ, rmd) and B-band synthesis genes (wbpJ, wbpA, wzx) in wild-type cells on agar. Nontreated control cells were incubated on agar for 15 min. Error bars indicate 95% confidence intervals. (B) Relative expression of lpxA and waaP in wild-type cells on agar. Error bars indicate 95% confidence intervals. (C) Relative expression of A-band synthesis genes (wbpZ, rmd) and B-band synthesis genes (wbpJ, wbpA) in PA0906S153A lexAG86V prtRS162A triple mutant cells on agar. Nontreated control cells were incubated on agar for 15 min. Error bars indicate 95% confidence intervals.
We assayed the expression of O-antigen biosynthesis genes in the PA0906S153A lexAG86V prtRS162A triple mutant after UV light exposure and found that B-band LPS O-antigen genes were significantly downregulated (Fig. 6C). This result indicates that the downregulation of B-band LPS O-antigen genes after UV light exposure is not dependent on RecA-mediated autocleavage of LexA, PrtR, or PA0906.
Previous research has shown that mutation of wbpA and wbpJ eliminates B-band O-antigen synthesis and confers sensitivity to self-produced R2 pyocin (32). Lower expression of wpbA and wpbJ likely lowers B-band O-antigen synthesis after UV light treatment. In addition, because both B- and A-band O antigens compete for the same site on the core polysaccharide during LPS synthesis (38), reduced B-band O-antigen synthesis caused by wpbA and wpbJ downregulation was likely compounded by the relatively high expression of A-band O-antigen synthesis genes. We propose that reduced B-band LPS synthesis after UV treatment produces patches of LPS where R2 pyocin can access its receptor.
DISCUSSION
Pyocins are produced by nearly all P. aeruginosa strains (14), an indication of their importance for this species. However, our results clearly show that pyocin biosynthesis carries a significant cost for P. aeruginosa. Induction of PrtR-regulated genes impairs survival during genotoxic stress, lowers resistance to clinically important antibiotics, and lowers resistance to an agent that causes oxidative stress. Our study has shown that there are at least two distinct mechanisms by which PrtR-regulated pyocin production affects resistance to genotoxic stress. (i) Lysis genes induced following PrtR autocleavage reduce resistance to bactericidal levels of ciprofloxacin, and (ii) production of extracellular R2 pyocin is lethal to cells that initially survive UV treatment. It is likely that the many bacterial species that produce bacteriocins as part of their SOS response (39) incur a biological cost similar to the one we described here during severe genotoxic stress.
LexA and PrtR are conserved in P. aeruginosa, and it is likely that each regulator simultaneously interacts with the RecA coprotease during the SOS response. The fact that genes induced by LexA autocleavage enhance survival and genes induced by PrtR autocleavage reduce survival raises the question of how P. aeruginosa balances the induction of these two opposing responses during genotoxic stress. On the basis of research with E. coli on the autocleavage of LexA and the CI phage repressor, we speculate that the rates of LexA and PrtR autocleavage during genotoxic stress differ and that the different rates of LexA and PrtR autocleavage determine which response is induced during a genotoxic event. In E. coli lysogens that harbor lambda prophage, the RecA coprotease also mediates the autoproteolysis of two regulators, LexA and CI. However, despite both being targeted by the RecA coprotease, LexA undergoes autoproteolysis at a higher rate than the CI repressor (40). The different rates of autoproteolytic cleavage in vivo reflect the strength of interaction between the site of cleavage and the protease-active site, and the strength of this interaction is greater in LexA than it is in the CI repressor (40, 41). Since the PrtR regulator is orthologous to a phage repressor, these findings may have some relevance to the P. aeruginosa SOS response. If PrtR has retained the lower rate of self-cleavage found in its ancestors, then PrtR autocleavage would be slower than LexA autocleavage. With this system, only the LexA regulon would be induced during an acute genotoxic event whereas the PrtR-regulated genes would be strongly induced only if genotoxic stress were sustained. It would be interesting to carry out a study of the relative rates of autocleavage of P. aeruginosa LexA, PrtR, and PA0906 and to analyze the kinetics of the induction of their regulons at different levels of genotoxic stress.
Unexpectedly, we found that downregulation of B-band O-antigen genes after UV light treatment is not dependent on the cleavage of LexA, PrtR, and PA0906, suggesting that another pathway that responds to UV light (or the effects of UV light) regulates B-band O-antigen synthesis genes. Interestingly, ciprofloxacin and hydrogen peroxide treatments also cause a modest downregulation of some B-band O-antigen synthesis genes (8–10), suggesting that this putative pathway responds to other agents that cause DNA damage. Thus, this as-yet-unidentified regulator could represent an important new DNA damage response pathway in P. aeruginosa. Moreover, because B-band O-antigen is required for complement resistance in humans (42, 43), understanding how B-band O-antigen genes are regulated in response to stress could potentially have clinical and therapeutic implications. Determining how UV light treatment affects the expression of B-band O-antigen genes will be the subject of future work.
Our surprising discovery that P. aeruginosa loses resistance to R2 pyocin following UV treatment indicates that there is a risk for bacteria that produce high-molecular-weight (HMW) or phage tail-like bacteriocins. B-band O-antigen synthesis confers resistance to R2 pyocin, and B-band O-antigen loss severely impairs growth (32). Unlike the immunity genes for low-molecular-weight S-type pyocins, which are genetically linked and coexpressed, the genes that code for B-band O-antigen synthesis are genetically distinct from the genes that code for R2 pyocin synthesis (14). Our work demonstrates that these genes are also transcriptionally regulated by distinct mechanisms and that R2 pyocin can be lethal under conditions that downregulate B-band O-antigen synthesis genes. This raises the possibility that R2 pyocin might restrict the conditions under which P. aeruginosa can grow because the expression of B-band O-antigen synthesis genes must be maintained to resist R2 pyocin-mediated killing. Many bacteria produce HMW or phage tail-like bacteriocins (44–52), and it is likely that the genes that confer resistance and synthesize the HMW bacteriocin are not genetically linked. These species may also have to balance the benefits of producing HMW bacteriocins with the risks that may pose under certain conditions.
Supplementary Material
ACKNOWLEDGMENTS
We thank Colin Manoil for transposon mutant strains; John Govan, Dean Scholl, and Suphan Bakkal for pyocin indicator strains; and the Walker lab for helpful discussion.
This work was supported by NIH GM31010 to G.C.W.; by grants from the NIH (R01HL110879, R01AI101307, and K24HL102246), the Cystic Fibrosis Foundation, and the Burroughs Wellcome Fund to P.K.S.; by P30 ES002109 to the MIT Center for Environmental Health Sciences; and by P30-CA14051 to the Cancer Center Support (core facilities). G.C.W. is an American Cancer Society Professor. J.P. was supported by a postdoctoral NIH National Research Service Award.
We have no conflict of interest to declare.
Footnotes
Published ahead of print 14 July 2014
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JB.01889-14.
REFERENCES
- 1.Friedberg EC, Walker GC, Siede W, Wood RD, Schultz RA, Ellenberger T. 2006. DNA repair and mutagenesis. ASM Press, Washington, DC [Google Scholar]
- 2.Kreuzer KN. 2013. DNA damage responses in prokaryotes: regulating gene expression, modulating growth patterns, and manipulating replication forks. Cold Spring Harb. Perspect. Biol. 5(11):a012674. 10.1101/cshperspect.a012674 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Walker GC. 1995. SOS-regulated proteins in translesion DNA synthesis and mutagenesis. Trends Biochem. Sci. 20:416–420. 10.1016/S0968-0004(00)89091-X [DOI] [PubMed] [Google Scholar]
- 4.Jacoby GA, Walsh KE, Mills DM, Walker VJ, Oh H, Robicsek A, Hooper DC. 2006. qnrB, another plasmid-mediated gene for quinolone resistance. Antimicrob. Agents Chemother. 50:1178–1182. 10.1128/AAC.50.4.1178-1182.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang M, Jacoby GA, Mills DM, Hooper DC. 2009. SOS regulation of qnrB expression. Antimicrob. Agents Chemother. 53:821–823. 10.1128/AAC.00132-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cascales E, Buchanan SK, Duche D, Kleanthous C, Lloubes R, Postle K, Riley M, Slatin S, Cavard D. 2007. Colicin biology. Microbiol. Mol. Biol. Rev. 71:158–229. 10.1128/MMBR.00036-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nakayama K, Takashima K, Ishihara H, Shinomiya T, Kageyama M, Kanaya S, Ohnishi M, Murata T, Mori H, Hayashi T. 2000. The R-type pyocin of Pseudomonas aeruginosa is related to P2 phage, and the F-type is related to lambda phage. Mol. Microbiol. 38:213–231. 10.1046/j.1365-2958.2000.02135.x [DOI] [PubMed] [Google Scholar]
- 8.Cirz RT, O'Neill BM, Hammond JA, Head SR, Romesberg FE. 2006. Defining the Pseudomonas aeruginosa SOS response and its role in the global response to the antibiotic ciprofloxacin. J. Bacteriol. 188:7101–7110. 10.1128/JB.00807-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brazas MD, Hancock RE. 2005. Ciprofloxacin induction of a susceptibility determinant in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 49:3222–3227. 10.1128/AAC.49.8.3222-3227.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chang W, Small DA, Toghrol F, Bentley WE. 2005. Microarray analysis of Pseudomonas aeruginosa reveals induction of pyocin genes in response to hydrogen peroxide. BMC Genomics 6:115. 10.1186/1471-2164-6-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sanders LH, Rockel A, Lu H, Wozniak DJ, Sutton MD. 2006. Role of Pseudomonas aeruginosa dinB-encoded DNA polymerase IV in mutagenesis. J. Bacteriol. 188:8573–8585. 10.1128/JB.01481-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Winsor GL, Lam DK, Fleming L, Lo R, Whiteside MD, Yu NY, Hancock RE, Brinkman FS. 2011. Pseudomonas Genome Database: improved comparative analysis and population genomics capability for Pseudomonas genomes. Nucleic Acids Res. 39:D596–D600. 10.1093/nar/gkq869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mo CY, Birdwell LD, Kohli RM. 2014. Specificity determinants for autoproteolysis of LexA, a key regulator of bacterial SOS mutagenesis. Biochemistry 53:3158–3168. 10.1021/bi500026e [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Michel-Briand Y, Baysse C. 2002. The pyocins of Pseudomonas aeruginosa. Biochimie 84:499–510. 10.1016/S0300-9084(02)01422-0 [DOI] [PubMed] [Google Scholar]
- 15.Früh R, Watson JM, Haas D. 1983. Construction of recombination-deficient strains of Pseudomonas aeruginosa. Mol. Gen. Genet. 191:334–337. 10.1007/BF00334835 [DOI] [PubMed] [Google Scholar]
- 16.Sano Y, Kageyama M. 1987. The sequence and function of the recA gene and its protein in Pseudomonas aeruginosa PAO. Mol. Gen. Genet. 208:412–419. 10.1007/BF00328132 [DOI] [PubMed] [Google Scholar]
- 17.Morse SA, Vaughan P, Johnson D, Iglewski BH. 1976. Inhibition of Neisseria gonorrhoeae by a bacteriocin from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 10:354–362. 10.1128/AAC.10.2.354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Matsui H, Sano Y, Ishihara H, Shinomiya T. 1993. Regulation of pyocin genes in Pseudomonas aeruginosa by positive (prtN) and negative (prtR) regulatory genes. J. Bacteriol. 175:1257–1263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sun Z, Shi J, Liu C, Jin Y, Li K, Chen R, Jin S, Wu W. 2014. PrtR homeostasis contributes to Pseudomonas aeruginosa pathogenesis and resistance against ciprofloxacin. Infect. Immun. 82:1638–1647. 10.1128/IAI.01388-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hoang TT, Karkhoff-Schweizer RR, Kutchma AJK, Schweizer HP. 1998. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 212:77–86. 10.1016/S0378-1119(98)00130-9 [DOI] [PubMed] [Google Scholar]
- 21.Hoang TT, Kutchma AJ, Becher A, Schweizer HP. 2000. Integration-proficient plasmids for Pseudomonas aeruginosa: site-specific integration and use for engineering of reporter and expression strains. Plasmid 43:59–72. 10.1006/plas.1999.1441 [DOI] [PubMed] [Google Scholar]
- 22.Irie Y, Starkey M, Edwards AN, Wozniak DJ, Romeo T, Parsek MR. 2010. Pseudomonas aeruginosa biofilm matrix polysaccharide Psl is regulated transcriptionally by RpoS and post-transcriptionally by RsmA. Mol. Microbiol. 78:158–172. 10.1111/j.1365-2958.2010.07320.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yarwood JM, Bartels DJ, Volper EM, Greenberg EP. 2004. Quorum sensing in Staphylococcus aureus biofilms. J. Bacteriol. 186:1838–1850. 10.1128/JB.186.6.1838-1850.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Williams SR, Gebhart D, Martin DW, Scholl D. 2008. Retargeting R-type pyocins to generate novel bactericidal protein complexes. Appl. Environ. Microbiol. 74:3868–3876. 10.1128/AEM.00141-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bakkal S, Robinson SM, Ordonez CL, Waltz DA, Riley MA. 2010. Role of bacteriocins in mediating interactions of bacterial isolates taken from cystic fibrosis patients. Microbiology 156:2058–2067. 10.1099/mic.0.036848-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Seo Y, Galloway DR. 1990. Purification of the pyocin S2 complex from Pseudomonas aeruginosa PAO1: analysis of DNase activity. Biochem. Biophys. Res. Commun. 172:455–461. 10.1016/0006-291X(90)90694-I [DOI] [PubMed] [Google Scholar]
- 27.Kuroda K, Kageyama M. 1981. Comparative study of F-type pyocins of Pseudomonas aeruginosa. J. Biochem. 89:1721–1736 [DOI] [PubMed] [Google Scholar]
- 28.Penterman J, Abo RP, De Nisco NJ, Arnold MF, Longhi R, Zanda M, Walker GC. 2014. Host plant peptides elicit a transcriptional response to control the Sinorhizobium meliloti cell cycle during symbiosis. Proc. Natl. Acad. Sci. U. S. A. 111:3561–3566. 10.1073/pnas.1400450111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Savli H, Karadenizli A, Kolayli F, Gundes S, Ozbek U, Vahaboglu H. 2003. Expression stability of six housekeeping genes: a proposal for resistance gene quantification studies of Pseudomonas aeruginosa by real-time quantitative RT-PCR. J. Med. Microbiol. 52:403–408. 10.1099/jmm.0.05132-0 [DOI] [PubMed] [Google Scholar]
- 30.Willems E, Leyns L, Vandesompele J. 2008. Standardization of real-time PCR gene expression data from independent biological replicates. Anal. Biochem. 379:127–129. 10.1016/j.ab.2008.04.036 [DOI] [PubMed] [Google Scholar]
- 31.Slilaty SN, Little JW. 1987. Lysine-156 and serine-119 are required for LexA repressor cleavage: a possible mechanism. Proc. Natl. Acad. Sci. U. S. A. 84:3987–3991. 10.1073/pnas.84.12.3987 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Penterman J, Nguyen D, Anderson E, Staudinger BJ, Greenberg EP, Lam JS, Singh PK. 2014. Rapid evolution of culture-impaired bacteria during adaptation to biofilm growth. Cell Rep. 6:293–300. 10.1016/j.celrep.2013.12.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Köhler T, Donner V, van Delden C. 2010. Lipopolysaccharide as shield and receptor for R-pyocin-mediated killing in Pseudomonas aeruginosa. J. Bacteriol. 192:1921–1928. 10.1128/JB.01459-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Baharoglu Z, Babosan A, Mazel D. 2014. Identification of genes involved in low aminoglycoside-induced SOS response in Vibrio cholerae: a role for transcription stalling and Mfd helicase. Nucleic Acids Res. 42:2366–2379. 10.1093/nar/gkt1259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Baharoglu Z, Mazel D. 2011. Vibrio cholerae triggers SOS and mutagenesis in response to a wide range of antibiotics: a route towards multiresistance. Antimicrob. Agents Chemother. 55:2438–2441. 10.1128/AAC.01549-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brawn MK, Fridovich I. 1985. Increased superoxide radical production evokes inducible DNA repair in Escherichia coli. J. Biol. Chem. 260:922–925 [PubMed] [Google Scholar]
- 37.Boles BR, Singh PK. 2008. Endogenous oxidative stress produces diversity and adaptability in biofilm communities. Proc. Natl. Acad. Sci. U. S. A. 105:12503–12508. 10.1073/pnas.0801499105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rahim R, Burrows LL, Monteiro MA, Perry MB, Lam JS. 2000. Involvement of the rml locus in core oligosaccharide and O polysaccharide assembly in Pseudomonas aeruginosa. Microbiology 146:2803–2814 [DOI] [PubMed] [Google Scholar]
- 39.Riley MA, Wertz JE. 2002. Bacteriocins: evolution, ecology, and application. Annu. Rev. Microbiol. 56:117–137. 10.1146/annurev.micro.56.012302.161024 [DOI] [PubMed] [Google Scholar]
- 40.Little JW. 1993. LexA cleavage and other self-processing reactions. J. Bacteriol. 175:4943–4950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kim B, Little JW. 1993. LexA and lambda Cl repressors as enzymes: specific cleavage in an intermolecular reaction. Cell 73:1165–1173. 10.1016/0092-8674(93)90645-7 [DOI] [PubMed] [Google Scholar]
- 42.Lam JS, Taylor VL, Islam ST, Hao Y, Kocincova D. 2011. Genetic and functional diversity of Pseudomonas aeruginosa lipopolysaccharide. Front. Microbiol. 2:118. 10.3389/fmicb.2011.00118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pier GB. 2007. Pseudomonas aeruginosa lipopolysaccharide: a major virulence factor, initiator of inflammation and target for effective immunity. Int. J. Med. Microbiol. 297:277–295. 10.1016/j.ijmm.2007.03.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liu J, Chen P, Zheng C, Huang YP. 2013. Characterization of maltocin P28, a novel phage tail-like bacteriocin from Stenotrophomonas maltophilia. Appl. Environ. Microbiol. 79:5593–5600. 10.1128/AEM.01648-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gebhart D, Williams SR, Bishop-Lilly KA, Govoni GR, Willner KM, Butani A, Sozhamannan S, Martin D, Fortier LC, Scholl D. 2012. Novel high-molecular-weight, R-type bacteriocins of Clostridium difficile. J. Bacteriol. 194:6240–6247. 10.1128/JB.01272-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jabrane A, Sabri A, Compere P, Jacques P, Vandenberghe I, Van Beeumen J, Thonart P. 2002. Characterization of serracin P, a phage-tail-like bacteriocin, and its activity against Erwinia amylovora, the fire blight pathogen. Appl. Environ. Microbiol. 68:5704–5710. 10.1128/AEM.68.11.5704-5710.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fischer S, Godino A, Quesada JM, Cordero P, Jofre E, Mori G, Espinosa-Urgel M. 2012. Characterization of a phage-like pyocin from the plant growth-promoting rhizobacterium Pseudomonas fluorescens SF4c. Microbiology 158:1493–1503. 10.1099/mic.0.056002-0 [DOI] [PubMed] [Google Scholar]
- 48.Morales-Soto N, Gaudriault S, Ogier JC, Thappeta KR, Forst S. 2012. Comparative analysis of P2-type remnant prophage loci in Xenorhabdus bovienii and Xenorhabdus nematophila required for xenorhabdicin production. FEMS Microbiol. Lett. 333:69–76. 10.1111/j.1574-6968.2012.02600.x [DOI] [PubMed] [Google Scholar]
- 49.Smarda J, Benada O. 2005. Phage tail-like (high-molecular-weight) bacteriocins of Budvicia aquatica and Pragia fontium (Enterobacteriaceae). Appl. Environ. Microbiol. 71:8970–8973. 10.1128/AEM.71.12.8970-8973.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Strauch E, Kaspar H, Schaudinn C, Dersch P, Madela K, Gewinner C, Hertwig S, Wecke J, Appel B. 2001. Characterization of enterocoliticin, a phage tail-like bacteriocin, and its effect on pathogenic Yersinia enterocolitica strains. Appl. Environ. Microbiol. 67:5634–5642. 10.1128/AEM.67.12.5634-5642.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zink R, Loessner MJ, Scherer S. 1995. Characterization of cryptic prophages (monocins) in Listeria and sequence analysis of a holin/endolysin gene. Microbiology 141(Pt 10):2577–2584. 10.1099/13500872-141-10-2577 [DOI] [PubMed] [Google Scholar]
- 52.McDonnell GE, Wood H, Devine KM, McConnell DJ. 1994. Genetic control of bacterial suicide: regulation of the induction of PBSX in Bacillus subtilis. J. Bacteriol. 176:5820–5830 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Held K, Ramage E, Jacobs M, Gallagher L, Manoil C. 2012. Sequence-verified two-allele transposon mutant library for Pseudomonas aeruginosa PAO1. J. Bacteriol. 194:6387–6389. 10.1128/JB.01479-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.