

ABSTRACT
The factors that determine CD4+ T cell (TCD4+) specificities, functional capacity, and memory persistence in response to complex pathogens remain unclear. We explored these parameters in the C57BL/6 mouse through comparison of two highly related (>92% homology) poxviruses: ectromelia virus (ECTV), a natural mouse pathogen, and vaccinia virus (VACV), a heterologous virus that nevertheless elicits potent immune responses. In addition to elucidating several previously unidentified major histocompatibility complex class II (MHC-II)-restricted epitopes, we observed many qualitative and quantitative differences between the TCD4+ repertoires, including responses not elicited by VACV despite complete sequence conservation. In addition, we observed functional heterogeneity between ECTV- and VACV-specific TCD4+ at both a global and individual epitope level, particularly greater expression of the cytolytic marker CD107a from TCD4+ following ECTV infection. Most striking were differences during the late memory phase where, in contrast to ECTV, VACV infection failed to elicit measurable epitope-specific TCD4+ as determined by intracellular cytokine staining. These findings illustrate the strong influence of epitope-extrinsic factors on TCD4+ responses and memory.
IMPORTANCE Much of our understanding concerning host-pathogen relationships in the context of poxvirus infections stems from studies of VACV in mice. However, VACV is not a natural mouse pathogen, and therefore, the relevance of results obtained using this model may be limited. Here, we explored the MHC class II-restricted TCD4+ repertoire induced by mousepox (ECTV) infection and the functional profile of the responding epitope-specific TCD4+, comparing these results to those induced by VACV infection under matched conditions. Despite a high degree of homology between the two viruses, we observed distinct specificity and functional profiles of TCD4+ responses at both acute and memory time points, with VACV-specific TCD4+ memory being notably compromised. These data offer insight into the impact of epitope-extrinsic factors on the resulting TCD4+ responses.
INTRODUCTION
Through their recognition of pathogen-derived peptides presented by major histocompatibility complex class II (MHC-II), CD4+ T cells (TCD4+) play important roles in shaping cellular (1, 2) and humoral immunity (3, 4) and in establishing immunological memory (5–7). Additionally, TCD4+ can suppress viral replication through the secretion of antiviral cytokines, such as gamma interferon (IFN-γ), and less frequently, through cytotoxic granule-mediated killing of infected cells (5, 8, 9).
Smallpox, caused by the Variola virus poxvirus, plagued mankind for millennia and continues to be a concern due to the threat of weaponization (10–13). Other poxviruses are equally lethal to their natural hosts, including ectromelia virus (ECTV), a poxvirus that causes smallpox-like symptoms in mice. Due to the threat it poses to mouse colonies, ECTV has not been widely investigated, and our understanding of host-poxvirus interplay and the resulting TCD4+ response stems mainly from studies in mice with vaccinia virus (VACV), a poxvirus of unknown origin and the centuries-old vaccine against smallpox. Moreover, these poxviruses have distinct courses of infection after intradermal infection in mice. ECTV multiplies rapidly at the site of infection before disseminating into the lymphatics and bloodstream, where it leads to a systemic infection that affects both the liver and spleen (14, 15), whereas VACV remains relatively localized after intradermal infection and does not lead to systemic infection (16). Importantly, because VACV is not a natural mouse pathogen, despite a high degree of homology with ECTV, the relevance of results from the widely studied VACV murine infection model may be limited. For example, distinct innate responses (17–21) that can alter the array of immunogenic peptides (22), which can profoundly affect TCD4+ responses, can differ substantially even with highly related viruses due to host cell tropism and host-specific immunomodulatory factors, such as viral cytokine mimics and/or receptors (23–30). These epitope-extrinsic factors can dramatically alter the course of infection and the resulting host immune response. For instance, it has been previously reported that Toll-like receptor 9 (TLR9) is critical for resistance against ECTV but not VACV (31). Indeed, low-dose footpad infection of C57BL/6 mice with ECTV usually results in loss of the infected limb, while much higher doses of VACV cause no discernible long-term effects. Thus, a comparative analysis of ECTV and VACV infection in mice provides an excellent opportunity to reveal the character of the ensuing virus-specific TCD4+ responses through the examination of specificity and functionality.
The primary aim of the present study was to compare the reactivity, magnitude, and functionality of ECTV- and VACV-specific TCD4+. By screening a large number of 12- to 15-mer peptides, we identified a total of 14 ECTV-specific TCD4+ epitopes and observed both quantitative and qualitative differences between the TCD4+ epitope repertoires elicited by ECTV and VACV. Subsequently, we probed differences in virus-mediated imprinting on TCD4+ function and found that the resulting profiles of epitope-specific TCD4+ are distinct and that long-term TCD4+ memory to ECTV is substantially stronger. In total, these data offer insight into the degree to which the alignment of host and pathogen can affect the specificity and functionality of responding virus-specific TCD4+.
MATERIALS AND METHODS
Ethics statement.
All experimental procedures involving mice were approved by the Institutional Animal Care and Use Committee at Thomas Jefferson University (Philadelphia, PA) and carried out in a humane manner.
Viruses.
The VACV WR strain was obtained from Bernard Moss (National Institute of Allergy and Infectious Diseases) and grown in 143 TK– cells. The ECTV Moscow strain was grown in BSC1 cells. The Δevm0158 ECTV (Fox Chase Cancer Center) was generated by homologous recombination, like other mutant viruses, (32, 33) and grown in BSC1 cells.
Mice.
Six- to 8-week-old female C57BL/6 mice were primed via footpad with 3,000 PFU ectromelia virus (Moscow strain), 3,000 PFU or 3 × 106 PFU vaccinia virus (Western Reserve), or 3,000 PFU Δevm0158 ECTV. C57BL/6 mice were obtained from The Jackson Laboratory and were used between 6 and 8 weeks of age according to the National Institutes of Health guidelines and Institutional Animal Care and Use Committee-approved animal protocols.
Infection and immunizations.
Six- to 8-week-old female C57BL/6 female mice were infected in the left hind footpad with either 3,000 PFU ectromelia virus (Moscow strain), 3,000 PFU or 3 × 106 PFU vaccinia virus (Western Reserve), or 3,000 PFU Δevm0158 ECTV. At various time points postinfection, the mice were observed and/or sacrificed and the inguinal lymph nodes, liver, and spleen were harvested. The splenocytes were used in either ex vivo IFN-γ enzyme-linked immunosorbent spot assay (ELISpot) or intracellular cytokine staining (ICS) assay as described below.
Epitope mapping.
The 1,022 peptides used for mapping were a subset of a previously described library (34). Briefly, peptides were synthesized as crude material by Pepscan Systems and mimotopes ranging from 12 to 15 amino acids in length were used previously to identify VACV epitopes (34). The peptides were screened for reactivity against splenocytes from ECTV-primed mice at various time points postinfection. Splenocytes from naive mice were used as antigen-presenting cells. Naive splenocytes were incubated with peptide (final concentration, 2 μg/ml) at 37°C and 5% CO2. Peptide-primed splenocytes were then coincubated overnight with either ECTV-specific whole splenocytes or TCD4+ isolated from splenocytes (Dynal mouse CD4 negative isolation kit; Invitrogen). IFN-γ-positive T cell responses were assayed by IFN-γ ELISpot (BD). Spots were counted using ImmunoSpot software (Cellular Technology Limited). To account for varying signal-to-noise ratios, we also calculated a stimulation index (SI), defined as (SFC [spot-forming cell] experiment results)/(SFC background) (34). Peptides with average spot numbers of ≥20, means of 1 × 106 effector T cells, P values of <0.05, and SI values of >2 in three independent experiments were considered positive.
ICS assay.
ICS assays were performed as previously described (15). Briefly, bone marrow-derived dendritic cells (1 × 106) generated using previously published methods (35) were either pulsed with peptides (3 μg/ml) for 1 h in a 96-well plate or were infected with VACV WR (multiplicity of infection [MOI] of 5) for between 10 and 18 h before the addition of 1 × 106 to 2 × 106 splenocytes (pooled from two to five mice that were immunized with VACV WR for 10 days). Two hours later, brefeldin A (10 μg/ml) was added, and cells were cultured for another 6 h before staining according to the protocol of the BD Fix/Perm solution kit (BD Biosciences). At least 1.5 × 106 to 2 × 106 events per sample were collected using an LSRII fluorescence-activated cell sorting (FACS) system (BD Biosciences) and were analyzed with FlowJo software (Tree Star). Background values were determined from samples pulsed with dimethyl sulfoxide (DMSO) only (no peptide) and were subtracted from the experimental values. At least three independent experiments were performed for each peptide or peptide pool. A peptide was considered positive if the average of the individual experiments was at least 1 standard deviation above the background.
RESULTS
TCD4+ response magnitude after infection with ECTV or VACV.
The distinct pathogenesis of ECTV compared with that of VACV in C57BL/6 mice, despite >92% genetic identity (36), could be appreciated following footpad inoculation of these viruses (Fig. 1A). VACV-induced inflammation was often detectable by day 3 postinfection, while the onset of ECTV-induced swelling was typically not appreciable until day 6. This can be ascribed, at least in part, to greater subversion of early immune responses by ECTV than by VACV (14, 37). However, while VACV-induced inflammation remained relatively mild and eventually subsided, ECTV-associated swelling became considerable, ultimately leading to necrosis and loss of the limb within ∼21 days of infection in most cases, consistent with previous reports (38). In both infections, we found day 10 postinfection to be the time point at which virus-specific TCD4+ responses could be discriminately measured, providing an optimal signal-to-noise ratio (data not shown). The divergent pathogenesis of ECTV and VACV in mice, despite a high degree of genetic similarity, provides an ideal experimental system to explore the influence of virulence and infectivity on the resulting TCD4+ responses following poxvirus infection.
FIG 1.
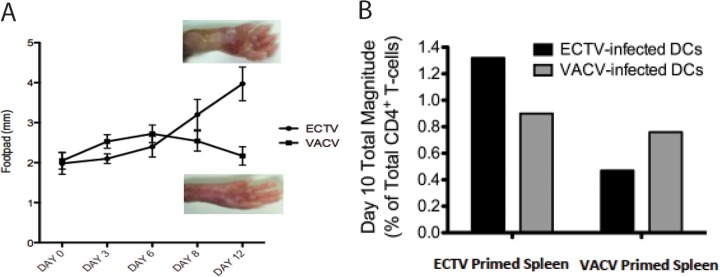
Divergent pathogenesis and magnitude of TCD4+ response to ECTV and VACV. (A) C57BL/6 mice were infected with either VACV or ECTV (3,000 PFU per footpad). Footpad swelling was measured using a caliper and recorded in millimeters at the indicated days postinfection. Representative photos of footpads at day 12 postinfection with either ECTV or VACV are shown. (B) TCD4+ response magnitude, represented as the frequency of total responding TCD4+ from mice primed with either VACV and ECTV (3,000 PFU per footpad). Responses were measured by intracellular cytokine staining after splenocytes were stimulated with BMDCs infected with ECTV or VACV (MOI = 1). Total magnitude was calculated after background subtraction by summing across all combinations of cells producing at least one of the following functions: CD107a, IFN-γ, IL-2, and TNF-α.
The global TCD4+ responses to ECTV and VACV were initially compared to determine differences in the overall magnitudes of the responses to these two distinct poxviruses. We primed C57BL/6 mice with ECTV or VACV, and 7 days later, cocultured splenocytes with bone marrow-derived dendritic cells (BMDCs) infected with either ECTV or VACV. Using flow cytometry, we measured the magnitude of the TCD4+ responses by examining several T cell functional outputs (Fig. 1B). Heterologous stimulation produced approximately 62% (VACV TCD4+ to ECTV BMDC) to 67% (ECTV TCD4+ to VACV BMDC) of the numbers of activated TCD4+ produced by homologous stimulation. This was 25 to 30% lower than what would be anticipated based upon the degree of sequence identity shared by the proteomes of VACV and ECTV (36, 39). This suggested a disparity in the TCD4+ responses to the two poxviruses extending beyond sequence heterogeneity, a possibility that was first investigated by assessing responses to individual epitopes.
TCD4+ specificities elicited by VACV and ECTV.
Previous mapping of the MHC-II-restricted C57BL/6 response to intraperitoneal VACV infection was accomplished with a library of synthetic peptides, comprising ∼30% of the predicted transcriptome of VACV (34). That study identified 14 specificities, with late-phase antigens predominating among the list of parent proteins (34). Utilizing 1,022 of those peptides (comprising ∼15% of the predicted transcriptome of VACV), we compared the reactivities elicited by ECTV and VACV in C57BL/6 mice by ELISpot analysis (Fig. 2) (34). Since the common route of entry for ECTV is through abrasions on the skin (38, 40–42), we performed footpad injection at the standard dose (3,000 PFU) as the route of infection for this study. Importantly, for the ELISpot screening, both the route and dose of VACV were matched to allow for direct comparison with the results obtained following ECTV priming. Spleens were harvested 10 days after infection, and the frequency of reproducible ECTV epitopes (Fig. 3A) measured was relatively low, on the order of 1 to 2% (Fig. 3B), consistent with the previous VACV screen (34).
FIG 2.

Comparative epitope specificities and magnitudes of responses to VACV- and ECTV-primed splenocytes. Naive splenocytes were incubated with synthetic poxvirus peptides ranging from 12 to 15 amino acids in length and screened for reactivity against splenocytes from ECTV-primed mice 10 days postinfection. Naive splenocytes were incubated with peptide (final concentration, 2 μg/ml). Peptide-primed splenocytes were coincubated overnight with whole splenocytes from ECTV-primed mice (3,000 PFU per footpad [f.p.]) 10 days postinfection. IFN-γ-positive T cell responses were assayed by IFN-γ ELISpot. To determine the level of statistical significance, Student's t test was performed using the mean of triplicate values of the response. #, number; *, P < 0.05; limits, means ± standard deviations (SD); APC, antigen-presenting cell. Data are representative of 3 independent experiments.
FIG 3.

Determination of ECTV-specific TCD4+epitopes in C57BL/6 mice. A synthetic VACV peptide library of 1,022 peptides, ranging from 12 to 15 amino acids in length, was screened for reactivity against splenocytes from ECTV-primed mice (3,000 PFU per footpad) 10 days postinfection. Naive splenocytes were incubated with peptide (final concentration, 2 μg/ml). Peptide-primed splenocytes were coincubated overnight with ECTV-specific whole splenocytes. (A) IFN-γ-positive T cell responses were assayed by IFN-γ ELISpot. Stars indicate previously unreported poxvirus epitopes, and MHC class I or MHC class II epitopes are indicated below. (B) Frequency map of the positive peptides identified in panel A. (C) Confirmation that the epitopes identified in panel A are recognized by TCD4+. A screen was performed as described in the legend to panel A with splenocytes or purified TCD4+ from ECTV-primed mice. Limits, means ± SD. MHC class I or MHC class II epitopes are indicated below.
The majority of peptides screened were fully conserved between VACV and ECTV, consistent with the high degree of homology. Fourteen distinct and reproducible MHC-II-restricted specificities were identified in response to ECTV infection (Fig. 3A and C and Table 1). Eleven of these had been previously identified in the VACV screen, with two common epitopes (residues 46 to 60 of A18R [A18R46–60] and I1L7–21) differing by a single, apparently neutral, amino acid (Table 2). Thus, three epitopes (I4L632–646, B13R14–28, and E2L426–440) were novel (Table 3). Interestingly, when we screened VACV-specific TCD4+ (Fig. 2, solid bars), these three specificities were also elicited, although B13R14–28 and E2L426–440 were just above the limit of detection. Three other specificities identified in the original VACV screen (D8L238–252, A28L10–24, and A24R399–413) were not elicited by ECTV. This could not be attributed to sequence heterogeneity for any of the three because A28L10–24 and A24R399–413 are 100% conserved. And while the ECTV sequence differs from D8L238–252 by two residues (A244→V244 and A249→V249), the ECTV counterpart was also nonreactive (Fig. 4).
TABLE 1.
Identification of ECTV-specific epitopesa
| Peptide hit | Sequence | VACV protein | Time of expression | Function | MHC class |
|---|---|---|---|---|---|
| B8R20–27 | TSYKFESV | B8R (VACWR190) [A190] | Early | Virulence | I |
| M1L424–438 | KSIIIPFIAYFVLMH | M1L (VACWR030) [A030] | Early | Unknown | I |
| A18R46–60 | PKGFYASPSVKTSLV | A18R (VACWR138) | Early | Regulation | II |
| A20R230–244 | GDNIFIPSVITKSGK | A20R (VACWR141) [A141] | Early | Regulation | II |
| E9L176–190 | PSVFINPISHTSYCY | E9L (VACWR065) | Early | Regulation | II |
| I4L632–646 | EFQVVNPHLLRVLTE | I4L (VACWR073) [A073] | Early | Regulation | II |
| J4R75–89 | DDDYGEPIIITSYLQ | J4R (VACWR096) | Early | Regulation | II |
| B13R14–28 | ENVFISPASISSVLT | B13R (VACWR195) [A195] | Early | Virulence | II |
| E2L426–440 | RLMFEYPLTKEASDH | E2L (VACWR058) [A058] | Early | Unknown | II |
| E1L114–128 | VLTIKAPNVISSKIS | E1L (VACWR057) [A057] | Late | Regulation | II |
| D13L483–497 | PKIFFRPTTITANVS | D13L (VACWR118) [A118] | Late | Structural | II |
| I1L7–21 | QLVFNSISARALKAY | VACWR070 (I1L) | Late | Structural | II |
| I1L18–33 | LKAYFTAKINEMVDE | I1L (VACWR070) | Late | Structural | II |
| H3L269–283 | PGVMYAFTTPLISFF | H3L (VACWR101) | Late | Structural | II |
| L4R173–187 | ISKYAGINILNVYSP | L4R (VACWR091) [A091] | Late | Structural | II |
| F15L41–56 | TPRYIPSTSISSSNI | F15L (VACWR054) | Late | Unknown | II |
Boldface indicates previously unreported epitopes.
TABLE 2.
TCD4+ poxvirus-specific epitopes
| Peptide hit | Reactivitya | % Conservation of: |
Time of expression | MHC class | ||
|---|---|---|---|---|---|---|
| Epitope (differing residue[s]) | Flanking regionb | Parent protein | ||||
| ECTV > VACV | ||||||
| A20R230–244 | ECTV and VACV | 100 | 100 | 97 | Early | II |
| F15L41–56 | ECTV and VACV | 100 | 100 | 99 | Late | II |
| VACV > ECTV | ||||||
| I1L7–21 | ECTV and VACV | 93.3 (V9→I9) | 100 | 99 | Late | II |
| I1L18–33 | ECTV and VACV | 100 | 100 | 99 | Late | II |
| L4R173–187 | ECTV and VACV | 100 | 100 | 99 | Late | II |
| A24R399–413 | VACV only | 100 | 100 | 99 | Early | II |
| A28L7–21 | VACV only | 100 | 100 | 99 | Late | II |
| D8L235–249 | VACV only | 86.7 (A244→V244, A249→V249) | 100 | 95 | Late | II |
| ECTV ≈ VACV | ||||||
| A18R46–60 | ECTV and VACV | 93.3 (A54→S54) | 100 | 97 | Early | II |
| E9L176–190 | ECTV and VACV | 100 | 100 | 98 | Early | II |
| J4R75–89 | ECTV and VACV | 100 | 100 | 99 | Early | II |
| E1L114–128 | ECTV and VACV | 100 | 100 | 99 | Late | II |
| D13L483–497 | ECTV and VACV | 100 | 100 | 99 | Late | II |
| H3L269–283 | ECTV and VACV | 100 | 100 | 96 | Late | II |
Stimulation index (SI) of >2.
Includes 5 amino acid residues upstream and downstream from the epitope.
TABLE 3.
Previously unreported poxvirus-specific epitopes
| Peptide hit | Reactivitya | % Conservation of: |
Time of expression | MHC class | ||
|---|---|---|---|---|---|---|
| Epitope (differing residue) | Flanking regionb | Parent protein | ||||
| M1L424–438 | ECTV and VACV | 100 | 96 | 96 | Early | I |
| I4L632–646 | ECTV and VACV | 100 | 100 | 97 | Early | II |
| B13R14–28 | ECTV only | 93.3 (P20→S20) | 100 | 95 | Early | II |
| E2L426–440 | ECTV only | 100 | 100 | 98 | Early | II |
Stimulation index (SI) of >2.
Includes 5 amino acid residues upstream and downstream from the epitope.
FIG 4.
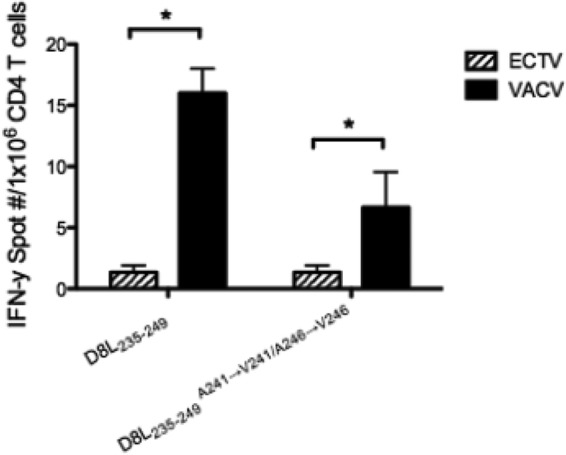
Comparative epitope reactivities to D8L238–252 between VACV- and ECTV-primed splenocytes. Naive splenocytes were incubated with synthetic D8L238–252 (VACV sequence) or D8L238–252A244→V244/A249→V249 (ECTV sequence) at a final concentration of 2 μg/ml. Peptide-primed splenocytes were then coincubated overnight with whole splenocytes from ECTV-primed mice or VACV-primed mice (3,000 PFU per footpad) 10 days postinfection. IFN-γ-positive T cell responses were assayed by IFN-γ ELISpot. To determine the level of statistical significance, Student's t test was performed using the mean of triplicate values of the response. *, P < 0.05.
At the same time, we expanded the screen to include CD8+ T cell (TCD8+) specificities and identified a novel and relatively potent MHC class I (MHC-I)-restricted epitope, M1L424–438, that elicited TCD8+ responses after both ECTV and VACV infections (Fig. 2 and 3C). The minimal H2-Kb-restricted epitope within the M1L424–338 peptide was defined via a series of truncated peptides as M1L426–434 (IIIPFIAYF) (data not shown).
The sum of the differences in TCD4+ specificities depicted in Fig. 2, 3, and 4 do not, on their own, account for the unexpected deficit in cross-reactivity shown by the results in Fig. 1. Particularly striking are the unequal responses to homologous epitopes. For example, despite the sequence identity, ECTV elicits significantly greater responses to the A20R230–244, E2L426–440, and F15L41–55 epitopes than VACV. Conversely, VACV elicits far greater responses to the I1L7–21, I1L18–33, and L4R173–187 epitopes. These differences in both directions indicate qualitative differences in the TCD4+ responses to the two viruses that are independent of epitope sequence. This led us to assess additional attributes of the resulting TCD4+ responses.
Minimal influence of the ECTV IFN-γ binding protein on assay results.
One factor we needed to address at an early stage was the IFN-γ binding protein expressed by both ECTV and VACV, since only the ECTV version has specificity for murine IFN-γ (43). Thus, the comparative ELISpot assays might have been compromised by this selective activity. To address this, we compared the responses of mice to wild-type (WT) ECTV and a recombinant strain that lacks the soluble IFN-γ receptor (B8R in VACV). The TCD4+ responses to WT and Δevm0158 ECTV were generally quite similar, with only a few significant differences (Fig. 5). For example, the E2L426–440 and F15L41–55 responses were reduced and the I1L18–33 response elevated in comparison to the responses of these epitopes to WT ECTV. Collectively, these data indicate that Δevm0158 ECTV does not significantly affect virus-specific TCD4+ reactivity or IFN-γ production. Compatible with this finding, Δevm0158 ECTV was not appreciably attenuated in vivo in our hands (unpublished data).
FIG 5.

Comparative epitope specificities and magnitudes of responses between WT ECTV- and Δevm0158 ECTV-primed splenocytes. Naive splenocytes were incubated with synthetic poxvirus peptides ranging from 12 to 15 amino acids in length (final concentration, 2 μg/ml). Peptide-primed splenocytes were coincubated overnight with whole splenocytes from WT ECTV- or Δevm0158 ECTV-primed mice (3,000 PFU per footpad) 10 days postinfection. IFN-γ-positive T cell responses were assayed by IFN-γ ELISpot. To determine the level of statistical significance, Student's t test was performed using the mean of triplicate values of the response. *, P < 0.05; limits, mean ± SD. Data are representative of 3 independent experiments.
Comparative functional profiles of epitope-specific TCD4+from ECTV- or VACV-primed mice.
To assess functionality using intracellular cytokine staining (ICS) and polychromatic flow cytometry, we pooled six peptides (Table 4, boldface) that consistently elicited robust TCD4+ responses in ELISpot assays, five being completely conserved between ECTV and VACV. Three cytokines were measured: IFN-γ, interleukin-2 (IL-2), and tumor necrosis factor alpha (TNF-α). In general, these molecules play well-described roles in antiviral immunity (44–47), and both IFN-γ and TNF-α have been found to be particularly important for protection following poxvirus infection (48, 49). In order to identify TCD4+ with cytotoxic potential, we also assessed degranulation by measuring the surface expression of CD107a after stimulation (50). In agreement with our prior study (15), the average TCD4+ response profile across all six epitopes was comparable between the two viruses (Fig. 6A). However, there were significant differences in the overall frequency of cytokine production when individual functions were assessed (Fig. 6B), and it became clear that ECTV-specific TCD4+ released significantly more cytotoxic granules after peptide stimulation.
TABLE 4.
Selected TCD4+/MHC class II poxvirus-specific epitopesa
| Peptide | Amino acid sequence |
|---|---|
| A20R230–244 | GDNIFIPSVITKSGK |
| I4L632–646 | EFQVVNPHLLRVLTE |
| F15L41–55 | TPRYIPSTSISSSNI |
| E1L114–128 | VLTIKAPNVISSKIS |
| I1L7–21 | QLVFNSISARALKAY |
| I1L18–33 | LKAYFTAKINEMVDE |
| D13L483–497 | PKIFFRPTTITANVS |
| E9L176–190 | PSVFINPISHTSYCY |
| H3L269–283 | PGVMYAFTTPLISFF |
| L4R173–187 | ISKYAGINILNVYSP |
Boldface indicates peptides that consistently elicited robust TCD4+ responses in ELISpot assays and were pooled to assess functionality using intracellular cytokine staining (ICS) and polychromatic flow cytometry.
FIG 6.

Comparative functional profiles of epitope-specific TCD4+ from ECTV- or VACV-primed mice. (A) TCD4+ responses to a poxvirus-specific class II peptide pool (Table 4) at day 10 were divided according to the relative contribution of each functional combination. Permutations that did not contribute significantly to the functional profile are not shown due to space constraints. Responses are grouped according to the degree of positivity and matched to the colors in the pie graphs for responses to the peptide pool at day 10. (B) Proportion of poxvirus-specific TCD4+ at day 10 postinfection for each specific function measured after stimulation. (C and D) Functional profiles of TCD4+ responsive to a selected cohort of poxvirus epitopes at day 10 (C) and day 100 (D) postinfection are shown for ECTV-infected (3,000 PFU per footpad) and VACV-infected (3,000 PFU per footpad or 3 ×106 PFU per footpad) mice. (A to D) All depicted data were from pooled cells of three mice at each time point and represent the average of two independent experiments. Bars represent the means, and error bars indicate the standard deviations. P values are defined where applicable; N.S., not significant. Data are representative of 3 independent experiments.
Next, we examined the functional profiles of the individual specificities. Despite the functional similarity between ECTV- and VACV-specific TCD4+ at a global (peptide pool) level, we observed heterogeneity in the individual response profiles (Fig. 6C). Of note, VACV-infected mice did not yield a response above the background for two specificities, I4L632–646 and H3L269–283 (Fig. 6C). Of the specificities that could be compared, E9L176–190-specific TCD4+ displayed the greatest functional divergence.
Additionally, due to the importance and relevance of long-lasting poxvirus immunization strategies (51, 52, 113), we examined epitope-specific TCD4+ functionality and persistence into the memory phase (≥100 days postinfection). We found that five of the six ECTV specificities were detectable (greater than or equal to 0.05% of total TCD4+) at 100 days postinfection (reactivity to I4L632–646 was undetectable by ICS), with minor degradation of functionality over time in each case. In striking contrast, no individual VACV-specific epitopes were detectable at 100 days postinfection, even though a 1,000-fold high dose of VACV was used for priming (Fig. 6D). In the context of this study, these data suggest that ECTV and VACV infections mediate distinct priming and functional imprinting of epitope-specific TCD4+ that affect persistence into the memory phase.
A greater frequency of effector TCD4+ is elicited by ECTV during acute infection.
Having examined CD107a expression among the poxvirus-specific responses (Fig. 6), we looked in more detail at the effector status of TCD4+ responses after infection. Our recent work points to the importance of cytolytic function by TCD4+ during acute ECTV infection of mice (9). Here, we asked whether cytolytic function is a poxvirus-specific phenomenon or unique to ECTV. Using flow cytometric analysis, we observed significant differences in the surface mobilization of cytotoxic granules by TCD4+ that were dependent upon both the epitope specificity and identity of the infecting poxvirus. The TCD4+ responses to two conserved epitopes, D13L483–497 and E9L176–190, serve as an illustration of these points, with ECTV infection inducing E9L176–190-specific TCD4+ with greater degranulation capacity than TCD4+ with other specificities (Fig. 7A). Additionally, compared with VACV, ECTV infection consistently elicited a higher proportion of responding TCD4+ with discernible CD107a expression after stimulation (Fig. 7A). The frequency of degranulation typically declined over time but was better maintained for some epitope specificities (Fig. 7B).
FIG 7.

ECTV-specific TCD4+ degranulate and express larger amounts of granzyme B during acute infection. (A) Proportions of D13L- or E9L-specific TCD4+ splenocytes coexpressing CD107a and IFN-γ from ECTV- or VACV-primed mice (3,000 PFU per footpad) at 10 days postinfection. Percentages represent the fraction of overlaid cells that fall within each quadrant. (B) Percentages of D13L-, E9L-, H3L-, I1L-, or L4R-specific TCD4+ splenocytes expressing CD107a from ECTV-primed mice (3,000 PFU per footpad) at acute (10 days postinfection) and memory (75 to 100 days postinfection) time points. (C) Percentages of grzB-positive (grzB+) cells from bulk TCD4+ collected from the spleen, inguinal lymph nodes (LN), or liver of ECTV- or VACV-primed mice (3,000 PFU per footpad) at 10 days postinfection. (D) Quantification of the average percentages of grzB+ from bulk TCD4+ collected from the spleen, inguinal LN, or liver of ECTV- or VACV-primed mice (3,000 PFU per footpad) at acute time points postinfection. Depicted data are from pooled cells of three mice and represent the average and SD of two independent experiments. P values are defined where applicable. Data are representative of 2 independent experiments.
Granzyme B (gzmB) is a major proapoptotic mediator stored within cytotoxic granules. As an additional way to assess cytotoxic potential, we measured the global levels of this molecule within total TCD4+ at acute time points postinfection with both viruses in the liver, inguinal lymph nodes, and spleen. We found that total TCD4+ in all three locations within ECTV-infected mice expressed 2- to 3-fold-higher levels of grzB than were observed with VACV (Fig. 7C and D). The enhanced grzB expression observed from ECTV TCD4+ suggests that the induction of cytolytic TCD4+ is characteristic of the murine host response to ECTV and not generally associated with murine poxvirus infection, as it is absent in VACV-immunized animals.
To further explore differential effector phenotypes between ECTV and VACV TCD4+, we examined the expression levels of macrophage inflammatory protein 1α (MIP1α) from ECTV- and VACV-specific TCD4+. MIP1α is a proinflammatory chemokine involved in the recruitment of immune cells, and its expression by T cells is associated with a more robust effector profile (53–55). We found greater expression of MIP1α from ECTV-specific TCD4+ than from VACV TCD4+ (Fig. 8), offering additional evidence that, in general, ECTV-specific TCD4+ display a greater effector-like profile than their VACV counterparts.
FIG 8.
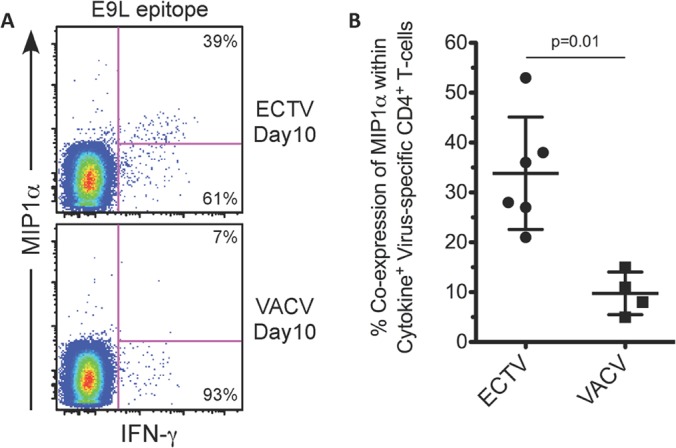
ECTV-specific TCD4+ express larger amounts of MIP1α during acute infection. (A) Representative staining showing higher expression of MIP1α by ECTV-specific TCD4+. E9L-specific TCD4+ responses from ECTV- or VACV-primed mice (3,000 PFU per footpad) at 10 days postinfection are shown. Percentages represent the fractions of IFN-γ+ cells that were either positive or negative for MIP1α after peptide stimulation. (B) Quantification of MIP1α expression among several different peptide-specific TCD4+ responses. Total percentages of virus-specific TCD4+ splenocytes coexpressing MIP1α and at least one additional cytokine from ECTV- or VACV-primed mice (3,000 PFU per footpad) at 10 days postinfection are shown. Depicted data are representative of four independent experiments. P value was determined using the Mann-Whitney test. Bars and whiskers represent the means and SD.
DISCUSSION
This study has revealed several ways in which host responses to the natural murine poxvirus, ECTV, differ considerably from those to VACV, the predominant model for examining poxvirus virulence and immunity in mice (16, 56–59). The initial experiments revealed a level of cross-reactivity, ∼62 to 67%, that was far lower than that expected by the degree of homology (92%). The basis for this became clear when we examined individual specificities with a 12- to 15-mer peptide library. Eleven of 14 previously identified VACV-induced specificities were elicited by ECTV (34), with the remaining three specificities (D8L238–252, A28L10–24, and A24R399–413) failing to develop in response to ECTV. The screen also uncovered four novel poxvirus epitopes for both ECTV and VACV, three of which were MHC class II (I4L632–646, B13R14–28, and E2L426–440) and one of which was MHC class I (M1L426–434, the precise boundaries being subsequently determined by a truncated-peptide series). Several factors may explain why the three MHC-II epitopes were not discovered in an earlier screen (33). Different routes of infection can affect TCD4+ differentiation, since they determine the initial cell types that interact with the virus (60), and prior screens utilized an intraperitoneal challenge (34), whereas here, we employed a dermal footpad challenge to mimic the natural infection route of ECTV (42). Additionally, prior VACV epitope screens utilized B cells to present synthetic peptides to TCD4+ (34), and differential costimulatory molecule expression by unique antigen-presenting cell types can alter signaling at the immunological synapse, resulting in shifted TCD4+ activation thresholds (61).
The novel MHC-I epitope, M1L426–434, induced relatively equivalent responses from both ECTV and VACV splenocytes that were nearly as potent as the immunodominant B8R20–27 for both VACV and ECTV splenocytes (Fig. 2), independent of infection route and dose (data not shown). In silico analyses that utilized algorithm-based predictions for MHC-I binders within the VACV transcriptome identified a 10-mer H-2Db class I epitope (TSNVITDQTV/M1L291–300) within the M1L parent protein (56) but not at the 426–434 location.
The lack of ECTV TCD4+ reactivity to the three specificities identified in the original VACV screen could not be accounted for by sequence heterogeneity, as A28L10–24 and A24R399–413 are both 100% conserved. And although there were two amino acid differences in the D8L238–252 epitope (A244→V244 and A249→V249), the possibility that these changes prevent proper processing of the antigen or prevent binding to the I-Ab molecule was discounted by the observation that VACV TCD4+ responds to the ECTV homolog. Thus, in all three cases, factors extrinsic to epitope composition are at work. One clear difference between the two viruses is the course of infection. ECTV productively infects a wider range of murine cell types, including dendritic cells, epidermal T cells, and keratinocytes (14, 41, 62). Thus, different sets of antigen-presenting cells with differing processing capabilities will be engaged. Furthermore, ECTV produces species-specific factors that allow for evasion and subversion of host responses and far greater replication in mice, as well as a greater antigen load, which will affect the levels of epitope display. Virulence factors and antigen load can also affect the cytokine milieu, which is markedly different in the two infections (15, 63–67) and which can strongly influence antigen-processing capabilities (68, 69). Chief among the cytokines of interest is IFN-γ, which, in addition to influencing the expression of antigen-processing components (68–71), drives upregulation of MHC-II (72). This was especially true for these investigations since ECTV but not VACV encodes a soluble IFN-γ receptor (B8R) that binds to murine IFN-γ (25, 73, 74). However, deletion of B8R did not have a substantial impact on overall TCD4+ magnitude or on the individual TCD4+ reactivities, consistent with our observation that Δevm0158 is not attenuated in vivo (unpublished data).
Differences in participating antigen-presenting cells and antigen load can also affect the functional character of the TCD4+ response (75–79). Indeed, we found that for the majority of the epitopes tested (5 of 6), ECTV-specific TCD4+ demonstrated a higher effector capacity; most notably enhanced were cytolytic potential and MIP1α expression. Furthermore, VACV TCD4+ exhibited responses above the background for just 4 of the 6 epitopes examined at 10 days postinfection, while ECTV-specific TCD4+ reacted strongly to all 6. We also identified higher percentages of grzB-positive cells from the tissues of ECTV-infected mice (Fig. 7C and D). This was true of all three tissues examined at acute time points postinfection. This result demonstrates that the generation of robust cytolytic TCD4+ is not a general property of all poxvirus infections and is consistent with earlier reports that increased antigen loads drive the development of cytolytic TCD4+ in both acute and chronic viral infections (80, 81).
Previous work by Fang et al. demonstrated that cytolytic TCD4+ contribute to the suppression and host control of ECTV replication (9), consistent with the well-documented pleiotropic antiviral effects of grzB expression. In addition to triggering apoptosis during ECTV infection, despite the production of antiapoptotic proteins such as B13R (82, 83), grzB has been shown to directly suppress VACV replication via cleavage of eukaryotic initiation factor 4 gamma 3 (eIF4G3), a protein essential for the initiation of protein translation (84). Increased grzB production may also play a role in the greater inflammation shown with ECTV at the site of infection (footpad) (Fig. 1) by inducing apoptosis of endothelial cells (anoikis) via granule exocytosis (85) and remodeling of extracellular matrix through cleavage of vitronectin, fibronectin, and laminin (86).
We also assessed the functional profiles of several peptide-specific TCD4+ at memory (day 100) time points using flow cytometry (Fig. 6D). While ECTV infection stimulated robust long-term TCD4+ memory, none of the 6 epitopes examined yielded a response above the background at day 100 postinfection with VACV, even when the input dose was 1,000-fold-greater for VACV than for ECTV. This was unexpected, since prior studies have shown persistence of VACV-specific TCD4+ in both humans (52) and mice (15, 66). In our own earlier study (15), we detected both TCD4+ memory responses at day 75 postinfection, utilizing a more comprehensive peptide pool than the one examined here, and VACV-infected presenting cells. Thus, while long-term TCD4+ memory to VACV may not be completely absent, it is significantly compromised.
The considerably greater difference between ECTV- and VACV-specific TCD4+ at the memory phase is consistent with the earlier demonstration that differences in the initial antiviral TCD4+ characters can become exaggerated as populations transition to central and/or effector memory cells (87, 88). Several factors might contribute to this amplification effect. Early expression of IL-15 by phagocytes and innate immune cells has been found to induce both a cytolytic TCD4+ character and TCD4+ effector memory cells (89–91) and has also been associated with NK cell recruitment, shown to be essential for natural resistance to ECTV (92). In addition, the above-mentioned parameters of antigen exposure and persistence can greatly affect the initial and long-term effectorlike properties of responding TCD4+ (77, 87, 93). At the same time, excessive antigen exposure can also lead to exhaustion in some settings (94). A greater understanding of the factors that set the balance between long-lasting TCD4+ effector-memory and exhaustion has been elusive (87, 94) but will likely be critical for insight into the profound differences in TCD4+ memory to two such homologous viruses.
The long-term protection against smallpox conferred by VACV is well known (52, 95–105). In a recent study of smallpox vaccinees, the levels of VACV-specific antibody, generally held to be the measure of protection (52), remained stable over many years postimmunization. In contrast and in line with our findings, VACV-specific TCD4+ memory declined over time (52). Due to the complexity of poxviruses, which clearly contain an abundance of MHC-II-restricted epitopes (this study and reference 34), this decline may not meaningfully impact protection. Alternatively or in addition, memory TCD4+ may not play a critical role in protection from smallpox. The importance of robust TCD4+ memory populations for protection from agents such as influenza virus and hepatitis A and C viruses is more apparent (106–111). Whether this is due to their relative simplicity and the limited number of TCD4+ epitopes they encode or other factors remains to be determined.
ECTV is proving to be an excellent model both in terms of providing an opportunity to study a natural host-pathogen relationship under defined conditions and in being highly relevant to other poxviruses (112), including smallpox and monkeypox (11, 12). The overall results of this study highlight interesting distinctions between the TCD4+ epitope reactivity profiles of ECTV and VACV. A better understanding of the mediators that influence TCD4+ reactivity and function is critical to designing better vaccines and antiviral therapeutics. Factors such as tropism and replication efficiency are likely major reasons for the functional differences observed between ECTV and VACV TCD4+ responding to common epitopes. Moreover, distinct host-pathogen relationships may ultimately play a predominant role in both shaping the TCD4+ repertoire and influencing the functional imprinting and differentiation of poxvirus-specific TCD4+. The striking cytolytic character of TCD4+ induced by ECTV and the inability of VACV to drive the development of this protective TCD4+ subset (89) or a substantial memory TCD4+ population demonstrate the impact of epitope-extrinsic mediators on TCD4+ repertoire, function, and persistence. Further study in this comparative poxvirus model may yield additional insights into the design of vaccine strategies that lead to more robust and long-lived TCD4+ responses.
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health grant U19-AI083008.
Research in this publication includes work carried out at the Kimmel Cancer Center Flow Cytometry Facility, which is supported in part by NCI Cancer Center support grant P30CA56036.
Footnotes
Published ahead of print 25 June 2014
REFERENCES
- 1.Janssen EM, Lemmens EE, Wolfe T, Christen U, von Herrath MG, Schoenberger SP. 2003. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature 421:852–856. 10.1038/nature01441 [DOI] [PubMed] [Google Scholar]
- 2.Nakanishi Y, Lu B, Gerard C, Iwasaki A. 2009. CD8(+) T lymphocyte mobilization to virus-infected tissue requires CD4(+) T-cell help. Nature 462:510–513. 10.1038/nature08511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yin L, Calvo-Calle JM, Cruz J, Newman FK, Frey SE, Ennis FA, Stern LJ. 2013. CD4+ T cells provide intermolecular help to generate robust antibody responses in vaccinia virus-vaccinated humans. J. Immunol. 190:6023–6033. 10.4049/jimmunol.1202523 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Crotty S, Kersh EN, Cannons J, Schwartzberg PL, Ahmed R. 2003. SAP is required for generating long-term humoral immunity. Nature 421:282–287. 10.1038/nature01318 [DOI] [PubMed] [Google Scholar]
- 5.Darrah PA, Patel DT, De Luca PM, Lindsay RW, Davey DF, Flynn BJ, Hoff ST, Andersen P, Reed SG, Morris SL, Roederer M, Seder RA. 2007. Multifunctional TH1 cells define a correlate of vaccine-mediated protection against Leishmania major. Nat. Med. 13:843–850. 10.1038/nm1592 [DOI] [PubMed] [Google Scholar]
- 6.Zhu J, Paul WE. 2008. CD4 T cells: fates, functions, and faults. Blood 112:1557–1569. 10.1182/blood-2008-05-078154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wan YY, Flavell RA. 2009. How diverse—CD4 effector T cells and their functions. J. Mol. Cell Biol. 1:20–36. 10.1093/jmcb/mjp001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Soghoian DZ, Jessen H, Flanders M, Sierra-Davidson K, Cutler S, Pertel T, Ranasinghe S, Lindqvist M, Davis I, Lane K, Rychert J, Rosenberg ES, Piechocka-Trocha A, Brass AL, Brenchley JM, Walker BD, Streeck H. 2012. HIV-specific cytolytic CD4 T cell responses during acute HIV infection predict disease outcome. Sci. Transl. Med. 4:123ra25. 10.1126/scitranslmed.3003165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fang M, Siciliano NA, Hersperger AR, Roscoe F, Hu A, Ma X, Shamsedeen AR, Eisenlohr LC, Sigal LJ. 2012. Perforin-dependent CD4+ T-cell cytotoxicity contributes to control a murine poxvirus infection. Proc. Natl. Acad. Sci. U. S. A. 109:9983–9988. 10.1073/pnas.1202143109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mahy BW, Almond JW, Berns KI, Chanock RM, Lvov DK, Pettersson RF, Schatzmayr HG, Fenner F. 1993. The remaining stocks of smallpox virus should be destroyed. Science 262:1223–1224. 10.1126/science.8235651 [DOI] [PubMed] [Google Scholar]
- 11.Anderson PD, Bokor G. 2012. Bioterrorism: pathogens as weapons. J. Pharm. Pract. 25:521–529. 10.1177/0897190012456366 [DOI] [PubMed] [Google Scholar]
- 12.Folio LR, Yao EF. 2007. US military smallpox vaccination program: occupational impact of immunizations on aircrew in Air Mobility Command, US Air Force. J. Am. Osteopath. Assoc. 107:547–553 [PubMed] [Google Scholar]
- 13.Whitby M, Street AC, Ruff TA, Fenner F. 2002. Biological agents as weapons 1: smallpox and botulism. Med. J. Aust. 176:431–433 [DOI] [PubMed] [Google Scholar]
- 14.Esteban DJ, Buller RM. 2005. Ectromelia virus: the causative agent of mousepox. J. Gen. Virol. 86:2645–2659. 10.1099/vir.0.81090-0 [DOI] [PubMed] [Google Scholar]
- 15.Hersperger AR, Siciliano NA, Eisenlohr LC. 2012. Comparable polyfunctionality of ectromelia virus- and vaccinia virus-specific murine T cells despite markedly different in vivo replication and pathogenicity. J. Virol. 86:7298–7309. 10.1128/JVI.00038-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tscharke DC, Smith GL. 1999. A model for vaccinia virus pathogenesis and immunity based on intradermal injection of mouse ear pinnae. J. Gen. Virol. 80(Pt 10):2751–2755 [DOI] [PubMed] [Google Scholar]
- 17.Jenkins MK, Khoruts A, Ingulli E, Mueller DL, McSorley SJ, Reinhardt RL, Itano A, Pape KA. 2001. In vivo activation of antigen-specific CD4 T cells. Annu. Rev. Immunol. 19:23–45. 10.1146/annurev.immunol.19.1.23 [DOI] [PubMed] [Google Scholar]
- 18.Tao X, Constant S, Jorritsma P, Bottomly K. 1997. Strength of TCR signal determines the costimulatory requirements for Th1 and Th2 CD4+ T cell differentiation. J. Immunol. 159:5956–5963 [PubMed] [Google Scholar]
- 19.Steinman RM, Hawiger D, Nussenzweig MC. 2003. Tolerogenic dendritic cells. Annu. Rev. Immunol. 21:685–711. 10.1146/annurev.immunol.21.120601.141040 [DOI] [PubMed] [Google Scholar]
- 20.Iwasaki A, Medzhitov R. 2004. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 5:987–995. 10.1038/ni1112 [DOI] [PubMed] [Google Scholar]
- 21.Zhu J, Yamane H, Paul WE. 2010. Differentiation of effector CD4 T cell populations. Annu. Rev. Immunol. 28:445–489. 10.1146/annurev-immunol-030409-101212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Khan S, van den Broek M, Schwarz K, de Giuli R, Diener PA, Groettrup M. 2001. Immunoproteasomes largely replace constitutive proteasomes during an antiviral and antibacterial immune response in the liver. J. Immunol. 167:6859–6868. 10.4049/jimmunol.167.12.6859 [DOI] [PubMed] [Google Scholar]
- 23.Smith VP, Alcami A. 2000. Expression of secreted cytokine and chemokine inhibitors by ectromelia virus. J. Virol. 74:8460–8471. 10.1128/JVI.74.18.8460-8471.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Alcami A, Smith GL. 1992. A soluble receptor for interleukin-1 beta encoded by vaccinia virus: a novel mechanism of virus modulation of the host response to infection. Cell 71:153–167. 10.1016/0092-8674(92)90274-G [DOI] [PubMed] [Google Scholar]
- 25.Alcami A, Smith GL. 1995. Vaccinia, cowpox, and camelpox viruses encode soluble gamma interferon receptors with novel broad species specificity. J. Virol. 69:4633–4639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Alcami A, Symons JA, Smith GL. 2000. The vaccinia virus soluble alpha/beta interferon (IFN) receptor binds to the cell surface and protects cells from the antiviral effects of IFN. J. Virol. 74:11230–11239. 10.1128/JVI.74.23.11230-11239.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Spriggs MK, Hruby DE, Maliszewski CR, Pickup DJ, Sims JE, Buller RM, VanSlyke J. 1992. Vaccinia and cowpox viruses encode a novel secreted interleukin-1-binding protein. Cell 71:145–152. 10.1016/0092-8674(92)90273-F [DOI] [PubMed] [Google Scholar]
- 28.Born TL, Morrison LA, Esteban DJ, VandenBos T, Thebeau LG, Chen N, Spriggs MK, Sims JE, Buller RM. 2000. A poxvirus protein that binds to and inactivates IL-18, and inhibits NK cell response. J. Immunol. 164:3246–3254. 10.4049/jimmunol.164.6.3246 [DOI] [PubMed] [Google Scholar]
- 29.Bowie A, Kiss-Toth E, Symons JA, Smith GL, Dower SK, O'Neill LA. 2000. A46R and A52R from vaccinia virus are antagonists of host IL-1 and toll-like receptor signaling. Proc. Natl. Acad. Sci. U. S. A. 97:10162–10167. 10.1073/pnas.160027697 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Loparev VN, Parsons JM, Knight JC, Panus JF, Ray CA, Buller RM, Pickup DJ, Esposito JJ. 1998. A third distinct tumor necrosis factor receptor of orthopoxviruses. Proc. Natl. Acad. Sci. U. S. A. 95:3786–3791. 10.1073/pnas.95.7.3786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Samuelsson C, Hausmann J, Lauterbach H, Schmidt M, Akira S, Wagner H, Chaplin P, Suter M, O'Keeffe M, Hochrein H. 2008. Survival of lethal poxvirus infection in mice depends on TLR9, and therapeutic vaccination provides protection. J. Clin. Invest. 118:1776–1784. 10.1172/JCI33940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Xu RH, Cohen M, Tang Y, Lazear E, Whitbeck JC, Eisenberg RJ, Cohen GH, Sigal LJ. 2008. The orthopoxvirus type I IFN binding protein is essential for virulence and an effective target for vaccination. J. Exp. Med. 205:981–992. 10.1084/jem.20071854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rubio D, Xu RH, Remakus S, Krouse TE, Truckenmiller ME, Thapa RJ, Balachandran S, Alcami A, Norbury CC, Sigal LJ. 2013. Crosstalk between the type 1 interferon and nuclear factor kappa B pathways confers resistance to a lethal virus infection. Cell Host Microbe 13:701–710. 10.1016/j.chom.2013.04.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Moutaftsi M, Bui HH, Peters B, Sidney J, Salek-Ardakani S, Oseroff C, Pasquetto V, Crotty S, Croft M, Lefkowitz EJ, Grey H, Sette A. 2007. Vaccinia virus-specific CD4+ T cell responses target a set of antigens largely distinct from those targeted by CD8+ T cell responses. J. Immunol. 178:6814–6820. 10.4049/jimmunol.178.11.6814 [DOI] [PubMed] [Google Scholar]
- 35.Siciliano NA, Skinner JA, Yuk MH. 2006. Bordetella bronchiseptica modulates macrophage phenotype leading to the inhibition of CD4+ T cell proliferation and the initiation of a Th17 immune response. J. Immunol. 177:7131–7138. 10.4049/jimmunol.177.10.7131 [DOI] [PubMed] [Google Scholar]
- 36.Chen N, Danila MI, Feng Z, Buller RM, Wang C, Han X, Lefkowitz EJ, Upton C. 2003. The genomic sequence of ectromelia virus, the causative agent of mousepox. Virology 317:165–186. 10.1016/S0042-6822(03)00520-8 [DOI] [PubMed] [Google Scholar]
- 37.O'Gorman WE, Sampath P, Simonds EF, Sikorski R, O'Malley M, Krutzik PO, Chen H, Panchanathan V, Chaudhri G, Karupiah G, Lewis DB, Thorne SH, Nolan GP. 2010. Alternate mechanisms of initial pattern recognition drive differential immune responses to related poxviruses. Cell Host Microbe 8:174–185. 10.1016/j.chom.2010.07.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Marchal J. 1930. Infectious ectromelia: a hitherto undescribed virus disease of mice. J. Pathol. Bacteriol. 33:713–728. 10.1002/path.1700330317 [DOI] [Google Scholar]
- 39.Gubser C, Hue S, Kellam P, Smith GL. 2004. Poxvirus genomes: a phylogenetic analysis. J. Gen. Virol. 85:105–117. 10.1099/vir.0.19565-0 [DOI] [PubMed] [Google Scholar]
- 40.Fenner F. 1981. Mousepox (infectious ectromelia): past, present, and future. Lab. Anim. Sci. 31:553–559 [PubMed] [Google Scholar]
- 41.Roberts JA. 1962. Histopathogenesis of mousepox. II. Cutaneous infection. Br. J. Exp. Pathol. 43:462–468 [PMC free article] [PubMed] [Google Scholar]
- 42.Fenner F. 1947. Studies in infectious ectromelia in mice; natural transmission; the portal of entry of the virus. Aust. J. Exp. Biol. Med. Sci. 25:275–282. 10.1038/icb.1947.39 [DOI] [PubMed] [Google Scholar]
- 43.Sakala IG, Chaudhri G, Buller RM, Nuara AA, Bai H, Chen N, Karupiah G. 2007. Poxvirus-encoded gamma interferon binding protein dampens the host immune response to infection. J. Virol. 81:3346–3353. 10.1128/JVI.01927-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Karupiah G. 1998. Type 1 and type 2 cytokines in antiviral defense. Vet. Immunol. Immunopathol. 63:105–109. 10.1016/S0165-2427(98)00086-5 [DOI] [PubMed] [Google Scholar]
- 45.Isaacs A, Lindenmann J. 1957. Virus interference. I. The interferon. Proc. R. Soc. Lond. B Biol. Sci. 147:258–267. 10.1098/rspb.1957.0048 [DOI] [PubMed] [Google Scholar]
- 46.D'Souza WN, Schluns KS, Masopust D, Lefrancois L. 2002. Essential role for IL-2 in the regulation of antiviral extralymphoid CD8 T cell responses. J. Immunol. 168:5566–5572. 10.4049/jimmunol.168.11.5566 [DOI] [PubMed] [Google Scholar]
- 47.Seo SH, Webster RG. 2002. Tumor necrosis factor alpha exerts powerful anti-influenza virus effects in lung epithelial cells. J. Virol. 76:1071–1076. 10.1128/JVI.76.3.1071-1076.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ruby J, Bluethmann H, Peschon JJ. 1997. Antiviral activity of tumor necrosis factor (TNF) is mediated via p55 and p75 TNF receptors. J. Exp. Med. 186:1591–1596. 10.1084/jem.186.9.1591 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chaudhri G, Panchanathan V, Buller RM, van den Eertwegh AJ, Claassen E, Zhou J, de Chazal R, Laman JD, Karupiah G. 2004. Polarized type 1 cytokine response and cell-mediated immunity determine genetic resistance to mousepox. Proc. Natl. Acad. Sci. U. S. A. 101:9057–9062. 10.1073/pnas.0402949101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Betts MR, Brenchley JM, Price DA, De Rosa SC, Douek DC, Roederer M, Koup RA. 2003. Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation. J. Immunol. Methods 281:65–78. 10.1016/S0022-1759(03)00265-5 [DOI] [PubMed] [Google Scholar]
- 51.Fenner F. 1989. Risks and benefits of vaccinia vaccine use in the worldwide smallpox eradication campaign. Res. Virol. 140:465–466. 10.1016/S0923-2516(89)80126-8 [DOI] [PubMed] [Google Scholar]
- 52.Hammarlund E, Lewis MW, Hansen SG, Strelow LI, Nelson JA, Sexton GJ, Hanifin JM, Slifka MK. 2003. Duration of antiviral immunity after smallpox vaccination. Nat. Med. 9:1131–1137. 10.1038/nm917 [DOI] [PubMed] [Google Scholar]
- 53.Casazza JP, Betts MR, Price DA, Precopio ML, Ruff LE, Brenchley JM, Hill BJ, Roederer M, Douek DC, Koup RA. 2006. Acquisition of direct antiviral effector functions by CMV-specific CD4+ T lymphocytes with cellular maturation. J. Exp. Med. 203:2865–2877. 10.1084/jem.20052246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Casazza JP, Brenchley JM, Hill BJ, Ayana R, Ambrozak D, Roederer M, Douek DC, Betts MR, Koup RA. 2009. Autocrine production of beta-chemokines protects CMV-Specific CD4 T cells from HIV infection. PLoS Pathog. 5:e1000646. 10.1371/journal.ppat.1000646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hersperger AR, Pereyra F, Nason M, Demers K, Sheth P, Shin LY, Kovacs CM, Rodriguez B, Sieg SF, Teixeira-Johnson L, Gudonis D, Goepfert PA, Lederman MM, Frank I, Makedonas G, Kaul R, Walker BD, Betts MR. 2010. Perforin expression directly ex vivo by HIV-specific CD8 T-cells is a correlate of HIV elite control. PLoS Pathog. 6:e1000917. 10.1371/journal.ppat.1000917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Moutaftsi M, Peters B, Pasquetto V, Tscharke DC, Sidney J, Bui HH, Grey H, Sette A. 2006. A consensus epitope prediction approach identifies the breadth of murine T(CD8+)-cell responses to vaccinia virus. Nat. Biotechnol. 24:817–819. 10.1038/nbt1215 [DOI] [PubMed] [Google Scholar]
- 57.Bauer S, Bathke B, Lauterbach H, Patzold J, Kassub R, Luber CA, Schlatter B, Hamm S, Chaplin P, Suter M, Hochrein H. 2010. A major role for TLR8 in the recognition of vaccinia viral DNA by murine pDC? Proc. Natl. Acad. Sci. U. S. A. 107:E139. 10.1073/pnas.1008626107 (Reply, 107:E140, ) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Symons JA, Adams E, Tscharke DC, Reading PC, Waldmann H, Smith GL. 2002. The vaccinia virus C12L protein inhibits mouse IL-18 and promotes virus virulence in the murine intranasal model. J. Gen. Virol. 83:2833–2844 [DOI] [PubMed] [Google Scholar]
- 59.Moutaftsi M, Salek-Ardakani S, Croft M, Peters B, Sidney J, Grey H, Sette A. 2009. Correlates of protection efficacy induced by vaccinia virus-specific CD8+ T-cell epitopes in the murine intranasal challenge model. Eur. J. Immunol. 39:717–722. 10.1002/eji.200838815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pepper M, Linehan JL, Pagan AJ, Zell T, Dileepan T, Cleary PP, Jenkins MK. 2010. Different routes of bacterial infection induce long-lived TH1 memory cells and short-lived TH17 cells. Nat. Immunol. 11:83–89. 10.1038/ni.1826 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kleindienst P, Brocker T. 2005. Concerted antigen presentation by dendritic cells and B cells is necessary for optimal CD4 T-cell immunity in vivo. Immunology 115:556–564. 10.1111/j.1365-2567.2005.02196.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Spohr de Faundez I, Gierynska M, Niemialtowski MG, Malicka E, Popis A. 1995. Ectromelia virus establishes a persistent infection in spleen dendritic cells and macrophages of BALB/c mice following the acute disease. Adv. Exp. Med. Biol. 378:257–261. 10.1007/978-1-4615-1971-3_57 [DOI] [PubMed] [Google Scholar]
- 63.Briody BA. 1959. Response of mice to ectromelia and vaccinia viruses. Bacteriol. Rev. 23:61–95 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fenner F. 1948. The pathogenesis of the acute exanthems; an interpretation based on experimental investigations with mousepox; infectious ectromelia of mice. Lancet ii:915–920 [DOI] [PubMed] [Google Scholar]
- 65.Fenner F. 1949. Mouse-pox; infectious ectromelia of mice; a review. J. Immunol. 63:341–373 [PubMed] [Google Scholar]
- 66.Harrington LE, van der Most R, Whitton JL, Ahmed R. 2002. Recombinant vaccinia virus-induced T-cell immunity: quantitation of the response to the virus vector and the foreign epitope. J. Virol. 76:3329–3337. 10.1128/JVI.76.7.3329-3337.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mahalingam S, Foster PS, Lobigs M, Farber JM, Karupiah G. 2000. Interferon-inducible chemokines and immunity to poxvirus infections. Immunol. Rev. 177:127–133. 10.1034/j.1600-065X.2000.17720.x [DOI] [PubMed] [Google Scholar]
- 68.Delvig AA, Lee JJ, Chrzanowska-Lightowlers ZM, Robinson JH. 2002. TGF-beta1 and IFN-gamma cross-regulate antigen presentation to CD4 T cells by macrophages. J. Leukoc. Biol. 72:163–166 [PubMed] [Google Scholar]
- 69.Collins T, Korman AJ, Wake CT, Boss JM, Kappes DJ, Fiers W, Ault KA, Gimbrone MA, Jr, Strominger JL, Pober JS. 1984. Immune interferon activates multiple class II major histocompatibility complex genes and the associated invariant chain gene in human endothelial cells and dermal fibroblasts. Proc. Natl. Acad. Sci. U. S. A. 81:4917–4921. 10.1073/pnas.81.15.4917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hastings KT. 2013. GILT: shaping the MHC class II-restricted peptidome and CD4 T cell-mediated immunity. Front. Immunol. 4:429. 10.3389/fimmu.2013.00429 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Li P, Gregg JL, Wang N, Zhou D, O'Donnell P, Blum JS, Crotzer VL. 2005. Compartmentalization of class II antigen presentation: contribution of cytoplasmic and endosomal processing. Immunol. Rev. 207:206–217. 10.1111/j.0105-2896.2005.00297.x [DOI] [PubMed] [Google Scholar]
- 72.Steimle V, Siegrist CA, Mottet A, Lisowska-Grospierre B, Mach B. 1994. Regulation of MHC class II expression by interferon-gamma mediated by the transactivator gene CIITA. Science 265:106–109. 10.1126/science.8016643 [DOI] [PubMed] [Google Scholar]
- 73.Oseroff C, Kos F, Bui HH, Peters B, Pasquetto V, Glenn J, Palmore T, Sidney J, Tscharke DC, Bennink JR, Southwood S, Grey HM, Yewdell JW, Sette A. 2005. HLA class I-restricted responses to vaccinia recognize a broad array of proteins mainly involved in virulence and viral gene regulation. Proc. Natl. Acad. Sci. U. S. A. 102:13980–13985. 10.1073/pnas.0506768102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Symons JA, Tscharke DC, Price N, Smith GL. 2002. A study of the vaccinia virus interferon-gamma receptor and its contribution to virus virulence. J. Gen. Virol. 83:1953–1964 [DOI] [PubMed] [Google Scholar]
- 75.Igyarto BZ, Kaplan DH. 2013. Antigen presentation by Langerhans cells. Curr. Opin. Immunol. 25:115–119. 10.1016/j.coi.2012.11.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Duraes FV, Thelemann C, Sarter K, Acha-Orbea H, Hugues S, Reith W. 2013. Role of major histocompatibility complex class II expression by non-hematopoietic cells in autoimmune and inflammatory disorders: facts and fiction. Tissue Antigens 82:1–15. 10.1111/tan.12136 [DOI] [PubMed] [Google Scholar]
- 77.Harari A, Vallelian F, Meylan PR, Pantaleo G. 2005. Functional heterogeneity of memory CD4 T cell responses in different conditions of antigen exposure and persistence. J. Immunol. 174:1037–1045. 10.4049/jimmunol.174.2.1037 [DOI] [PubMed] [Google Scholar]
- 78.Harari A, Vallelian F, Pantaleo G. 2004. Phenotypic heterogeneity of antigen-specific CD4 T cells under different conditions of antigen persistence and antigen load. Eur. J. Immunol. 34:3525–3533. 10.1002/eji.200425324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Shortman K, Liu YJ. 2002. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2:151–161. 10.1038/nri746 [DOI] [PubMed] [Google Scholar]
- 80.Appay V. 2004. The physiological role of cytotoxic CD4(+) T-cells: the holy grail? Clin. Exp. Immunol. 138:10–13. 10.1111/j.1365-2249.2004.02605.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Brown DM. 2010. Cytolytic CD4 cells: direct mediators in infectious disease and malignancy. Cell. Immunol. 262:89–95. 10.1016/j.cellimm.2010.02.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Pardo J, Galvez EM, Koskinen A, Simon MM, Lobigs M, Regner M, Mullbacher A. 2009. Caspase-dependent inhibition of mousepox replication by gzmB. PLoS One 4:e7512. 10.1371/journal.pone.0007512 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Macen JL, Garner RS, Musy PY, Brooks MA, Turner PC, Moyer RW, McFadden G, Bleackley RC. 1996. Differential inhibition of the Fas- and granule-mediated cytolysis pathways by the orthopoxvirus cytokine response modifier A/SPI-2 and SPI-1 protein. Proc. Natl. Acad. Sci. U. S. A. 93:9108–9113. 10.1073/pnas.93.17.9108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Marcet-Palacios M, Duggan BL, Shostak I, Barry M, Geskes T, Wilkins JA, Yanagiya A, Sonenberg N, Bleackley RC. 2011. Granzyme B inhibits vaccinia virus production through proteolytic cleavage of eukaryotic initiation factor 4 gamma 3. PLoS Pathog. 7:e1002447. 10.1371/journal.ppat.1002447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Trapani JA, Sutton VR. 2003. Granzyme B: pro-apoptotic, antiviral and antitumor functions. Curr. Opin. Immunol. 15:533–543. 10.1016/S0952-7915(03)00107-9 [DOI] [PubMed] [Google Scholar]
- 86.Buzza MS, Zamurs L, Sun J, Bird CH, Smith AI, Trapani JA, Froelich CJ, Nice EC, Bird PI. 2005. Extracellular matrix remodeling by human granzyme B via cleavage of vitronectin, fibronectin, and laminin. J. Biol. Chem. 280:23549–23558. 10.1074/jbc.M412001200 [DOI] [PubMed] [Google Scholar]
- 87.Sallusto F, Geginat J, Lanzavecchia A. 2004. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu. Rev. Immunol. 22:745–763. 10.1146/annurev.immunol.22.012703.104702 [DOI] [PubMed] [Google Scholar]
- 88.Williams MA, Ravkov EV, Bevan MJ. 2008. Rapid culling of the CD4+ T cell repertoire in the transition from effector to memory. Immunity 28:533–545. 10.1016/j.immuni.2008.02.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Soghoian DZ, Streeck H. 2010. Cytolytic CD4(+) T cells in viral immunity. Exp. Rev. Vaccines 9:1453–1463. 10.1586/erv.10.132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ye W, Young JD, Liu CC. 1996. Interleukin-15 induces the expression of mRNAs of cytolytic mediators and augments cytotoxic activities in primary murine lymphocytes. Cell. Immunol. 174:54–62. 10.1006/cimm.1996.0293 [DOI] [PubMed] [Google Scholar]
- 91.Picker LJ, Reed-Inderbitzin EF, Hagen SI, Edgar JB, Hansen SG, Legasse A, Planer S, Piatak M, Jr, Lifson JD, Maino VC, Axthelm MK, Villinger F. 2006. IL-15 induces CD4 effector memory T cell production and tissue emigration in nonhuman primates. J. Clin. Invest. 116:1514–1524. 10.1172/JCI27564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Fang M, Lanier LL, Sigal LJ. 2008. A role for NKG2D in NK cell-mediated resistance to poxvirus disease. PLoS Pathog. 4:e30. 10.1371/journal.ppat.0040030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Kaech SM, Wherry EJ, Ahmed R. 2002. Effector and memory T-cell differentiation: implications for vaccine development. Nat. Rev. Immunol. 2:251–262. 10.1038/nri778 [DOI] [PubMed] [Google Scholar]
- 94.Han S, Asoyan A, Rabenstein H, Nakano N, Obst R. 2010. Role of antigen persistence and dose for CD4+ T-cell exhaustion and recovery. Proc. Natl. Acad. Sci. U. S. A. 107:20453–20458. 10.1073/pnas.1008437107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Barquet N, Domingo P. 1997. Smallpox: the triumph over the most terrible of the ministers of death. Ann. Intern. Med. 127:635–642. 10.7326/0003-4819-127-8_Part_1-199710150-00010 [DOI] [PubMed] [Google Scholar]
- 96.Baxby D. 1981. Jenner's smallpox vaccine: the riddle of the origin of vaccinia virus. Heinemann, London, England [Google Scholar]
- 97.Behbehani AM. 1983. The smallpox story: life and death of an old disease. Microbiol. Rev. 47:455–509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Bloch H. 1993. Edward Jenner (1749–1823). The history and effects of smallpox, inoculation, and vaccination. Am. J. Dis. Child. 147:772–774 [DOI] [PubMed] [Google Scholar]
- 99.Dunn PM. 1996. Dr Edward Jenner (1749–1823) of Berkeley, and vaccination against smallpox. Arch. Dis. Child. Fetal Neonatal Ed. 74:F77–F78. 10.1136/fn.74.1.F77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Fenner F. 1977. The eradication of smallpox. Prog. Med. Virol. 23:1–21 [PubMed] [Google Scholar]
- 101.Fenner F. 1980. The global eradication of smallpox. Med. J. Aust. 1:455. [DOI] [PubMed] [Google Scholar]
- 102.Fenner F. 1982. A successful eradication campaign. Global eradication of smallpox. Rev. Infect. Dis. 4:916–930 [DOI] [PubMed] [Google Scholar]
- 103.Fenner F. 1984. Smallpox, “the most dreadful scourge of the human species.” Its global spread and recent eradication. Med. J. Aust. 141:841–846 [PubMed] [Google Scholar]
- 104.Metzger W, Mordmueller BG. 2007. Vaccines for preventing smallpox. Cochrane Database Syst. Rev. 18:CD004913. 10.1002/14651858.CD004913.pub2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Willis NJ. 1997. Edward Jenner and the eradication of smallpox. Scott. Med. J. 42:118–121 [DOI] [PubMed] [Google Scholar]
- 106.Lechner F, Wong DK, Dunbar PR, Chapman R, Chung RT, Dohrenwend P, Robbins G, Phillips R, Klenerman P, Walker BD. 2000. Analysis of successful immune responses in persons infected with hepatitis C virus. J. Exp. Med. 191:1499–1512. 10.1084/jem.191.9.1499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Lee BO, Rangel-Moreno J, Moyron-Quiroz JE, Hartson L, Makris M, Sprague F, Lund FE, Randall TD. 2005. CD4 T cell-independent antibody response promotes resolution of primary influenza infection and helps to prevent reinfection. J. Immunol. 175:5827–5838. 10.4049/jimmunol.175.9.5827 [DOI] [PubMed] [Google Scholar]
- 108.Shoukry NH, Cawthon AG, Walker CM. 2004. Cell-mediated immunity and the outcome of hepatitis C virus infection. Annu. Rev. Microbiol. 58:391–424. 10.1146/annurev.micro.58.030603.123836 [DOI] [PubMed] [Google Scholar]
- 109.Thomas PG, Keating R, Hulse-Post DJ, Doherty PC. 2006. Cell-mediated protection in influenza infection. Emerg. Infect. Dis. 12:48–54. 10.3201/eid1201.051237 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Wilkinson TM, Li CK, Chui CS, Huang AK, Perkins M, Liebner JC, Lambkin-Williams R, Gilbert A, Oxford J, Nicholas B, Staples KJ, Dong T, Douek DC, McMichael AJ, Xu XN. 2012. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 18:274–280. 10.1038/nm.2612 [DOI] [PubMed] [Google Scholar]
- 111.Zhou Y, Callendret B, Xu D, Brasky KM, Feng Z, Hensley LL, Guedj J, Perelson AS, Lemon SM, Lanford RE, Walker CM. 2012. Dominance of the CD4(+) T helper cell response during acute resolving hepatitis A virus infection. J. Exp. Med. 209:1481–1492. 10.1084/jem.20111906 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Parker S, Siddiqui AM, Oberle C, Hembrador E, Lanier R, Painter G, Robertson A, Buller RM. 2009. Mousepox in the C57BL/6 strain provides an improved model for evaluating anti-poxvirus therapies. Virology 385:11–21. 10.1016/j.virol.2008.11.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Boyle DB. 1989. How do other poxviruses fit in as potential vectored vaccine substrates for animal immunizations? Res. Virol. 140:483–485; discussion, 487–491 [DOI] [PubMed] [Google Scholar]