

Abstract
Rotaviruses (RVs) are a leading cause of childhood diarrhea. Current oral vaccines are not effective in impoverished countries where the vaccine is needed most. Therefore, alternative affordable strategies are urgently needed. Probiotics can alleviate diarrhea in children and enhance specific systemic and mucosal Ab responses, but the T cell responses are undefined. In this study, we elucidated the T cell and cytokine responses to attenuated human RV (AttHRV) and virulent human RV (HRV) in gnotobiotic pigs colonized with probiotics (Lactobacillus rhamnosus strain GG [LGG] and Bifidobacterium lactis Bb12 [Bb12]), mimicking gut commensals in breastfed infants. Neonatal gnotobiotic pigs are the only animal model susceptible to HRV diarrhea. Probiotic colonized and nonvaccinated (Probiotic) pigs had lower diarrhea and reduced virus shedding postchallenge compared with noncolonized and nonvaccinated pigs (Control). Higher protection in the Probiotic group coincided with higher ileal T regulatory cells (Tregs) before and after challenge, and higher serum TGF-β and lower serum and biliary proinflammatory cytokines postchallenge. Probiotic colonization in vaccinated pigs enhanced innate serum IFN-α, splenic and circulatory IFN-γ−producing T cells, and serum Th1 cytokines, but reduced serum Th2 cytokines compared with noncolonized vaccinated pigs (Vac). Thus, LGG+Bb12 induced systemic Th1 immunostimulatory effects on oral AttHRV vaccine that coincided with lower diarrhea severity and reduced virus shedding postchallenge in Vac+Pro compared with Vac pigs. Previously unreported intestinal CD8 Tregs were induced in vaccinated groups postchallenge. Thus, probiotics LGG+Bb12 exert divergent immunomodulating effects, with enhanced Th1 responses to oral AttHRV vaccine, whereas inducing Treg responses to virulent HRV.
Rotaviruses (RVs) are a major cause of acute gastroenteritis in infants and children. Annually, RV diarrhea is responsible for ∼450,000 deaths in children <5 years old (1). The two licensed oral RV vaccines have higher efficacy against severe RV gastroenteritis in developed countries than in developing countries, where the burden of RV infection is higher (2). Probiotics are alternative low-cost treatments to moderate infectious diarrhea, and they may also enhance oral vaccine efficacy in impoverished countries (3, 4). In children, initial microbial colonization depends on the mode of delivery and the type of feeding (breast versus formula fed). Lactobacilli (belongs to Firmicutes) and Bifidobacteria (belongs to Actinobacteria) spp. are dominant Gram-positive commensals in breastfed infants, and contribute to immune- and health-related benefits (5–7). Moreover, randomized clinical trials have shown that certain Lactobacilli and Bifidobacteria spp. reduced RV diarrhea in children (8, 9). Colonization by commensals (selected probiotics) results in induction of different subsets of Th cells including Th1, Th2, Th17, and T regulatory cells (Tregs) through interactions with dendritic cells, which may modulate immune responses to vaccines and infections (10–13). Commensal/probiotic colonization and their associated beneficial effects play a critical role early in life that may result in lifelong benefits (14). Thus, early colonization by probiotic species (Lactobacilli rhamnosus strain GG [LGG], Bifidobacterium animalis subsp. lactis Bb12 [Bb12]) may mature gut immunity to enhance oral vaccine efficacy and also moderate the severity of enteric infections.
RV induces diarrhea, dehydration, and vomiting in infants and gnotobiotic (Gn) pigs (15). Human RV (HRV)–induced diarrhea in both infected children and Gn pigs is characterized by upregulated proinflammatory cytokines in the blood, which correlates with diarrhea severity and viremia (16–18). Acute RV infection in children is associated with higher expression of TLR2, TLR3, TLR4, TLR7, and TLR8 on blood mononuclear cells (MNCs) (19). Similarly, Gn pigs infected with RV had increased numbers of TLR3 expressing APCs in blood and spleen (20). The similar immunopathogenesis of RV disease in children and Gn pigs indicates that Gn pigs are a relevant animal model for evaluating protection induced by HRV vaccines with or without adjuvants (probiotics). Previously, we and others showed that L. acidophilus, colonized in multiple increasing doses, modulate B and T cell responses to attenuated HRV (AttHRV) in Gn pigs, but no significant differences in protection against virulent HRV (VirHRV)–induced diarrhea were observed compared with vaccinated only pigs (21, 22). Moreover, supplementation with higher doses of the same probiotics in vaccinated pigs did not enhance protection from diarrhea and fecal HRV shedding. However, it resulted in differential immunomodulation as compared with low-dose L. acidophilus supplementation (21). A combination of L. acidophilus and L. reuteri given in multiple low doses also did not enhance protection against diarrhea and fecal shedding post-VirHRV infection in a Gn pig model, which coincided with no significant differences in HRV-specific Ab responses between probiotic colonized and noncolonized groups postchallenge (23). In children, probiotics and HRV studies have focused primarily on B cell responses, with limited studies investigating cell-mediated immune responses (8, 9, 24). We focused on comprehensively investigating how cell-mediated immune responses can be modulated by early colonization with probiotics (combination of LGG and Bb12), mimicking colonization kinetics of commensal species (Lactobacilli and Bifidobacteria) in breastfed infants.
In this study, we investigated the role of selected probiotics (LGG +Bb12), mimicking colonization patterns in breastfed infants, on T cell and cytokine responses to AttHRV Wa (G1P[8]) vaccine (genotypically similar to Rotarix, G1P[8]) and to VirHRV Wa infection. We hypothesize that initial cocolonization by LGG and Bb12 will enhance immune maturation and responses (adjuvant properties) to an oral AttHRV vaccine and reduce VirHRV diarrhea (prophylactic properties). We show that probiotics LGG+Bb12 enhanced immunostimulatory Th1 responses to oral AttHRV vaccine and induced immunoregulatory responses to VirHRV infection.
Materials and Methods
Virus and probiotic strains
Cell culture adapted AttHRV Wa was used as vaccine at a dose of 5 × 107 fluorescent foci-forming units (FFU) (23). The VirHRV Wa was used for challenge at a dose of 1 × 105 FFU as previously described (15). The probiotics LGG strain ATCC 53103 (American Type Culture Collection, Manassas, VA) and Bb12 (Christian Hansen, Horsholm, Denmark) were used to colonize Gn pigs as previously described (25).
Animals and experimental design
All experimental procedures were approved by The Ohio State University Institutional Animal Care and Use Committee (no. 2010A0088) and were conducted in compliance with local and federal guidelines. Piglets were surgically derived from near-term sows (Landrace × Yorkshire × Duroc crossbred) and maintained in sterile isolators as previously described (26). All pigs were seronegative for HRVAbs and were fed ultra-high-temperature processed commercial cow milk (Parmalat, Parma, Italy) throughout the trial. Gn piglets were randomly assigned to one of four treatment groups: 3XAttHRV Wa vaccinated and probiotic colonized (Vac+Pro: prechallenge n = 6, postchallenge n = 7), 3XAttHRV Wa vaccinated only (Vac: prechallenge n = 6, postchallenge n = 6), probiotic colonized only (Probiotics: prechallenge n = 6, postchallenge n = 5), and unvaccinated and nonprobiotic colonized negative controls (Control: prechallenge n = 5, postchallenge n = 4). Probiotic-fed groups were sequentially colonized orally at 3 d of age with Bb12 and at 5 d of age with LGG+Bb12 (1:1) at a dose of 105 CFUs (CFU)/pig/time point (Fig. 1). Vaccinated pigs were orally inoculated with the first vaccine dose at 6 d of age and then received two additional doses at 10-d intervals (postinoculation day [PID] 10 and PID20; Fig. 1). A subset of pigs was euthanized at PID27/postchallenge day (PCD) 0 to assess prechallenge responses. Remaining pigs were challenged with VirHRV and euthanized at PID34/PCD7 (Fig. 1).
Figure 1.
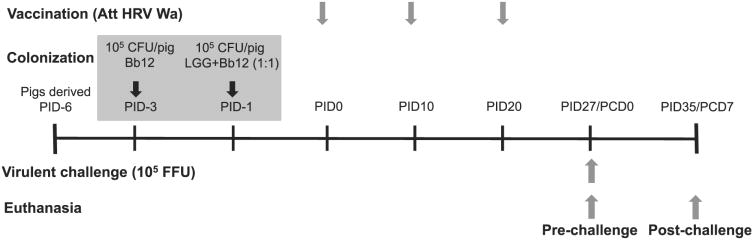
Schematic for experimental design showing time points for probiotic colonization, vaccination (AttHRV Wa), VirHRV Wa challenge, and euthanasia.
Detection of HRV and probiotic shedding and assessment of clinical signs
Rectal swabs were collected daily postchallenge to detect and quantitate infectious HRV by cell culture immunofluorescence assay as previously described (15). Pigs were observed daily for diarrhea post-VirHRV challenge. Probiotic intestinal colonization and fecal shedding was determined as previously described prechallenge and postchallenge (23, 25). In brief, sections (2–3 g) of small intestinal (duodenum, ileum and jejunum) and large intestinal (cecum, colon, and rectum) tissues were collected prechallenge and postchallenge, and were rinsed and homogenized in 0.1% peptone water. Enumeration of probiotics in rectal swab fluids and gut tissues was done by plating dilutions on MRS agar containing 0.05% cysteine hydrochloride, which were incubated in sealed BBL Gaspak jars (Fisher, Hanover Park, IL) containing Anaerogen packs (BD) for 24 h at 37°C. The combined LGG and Bb12 colonies in rectal swab fluids and tissues were enumerated collectively as total CFU and were expressed as CFU/ml for rectal swabs and CFU/g for gut tissues. Colonization by both LGG and Bb12 was confirmed using specific primers by quantitative PCR on DNA extracted from representative pigs as previously described (25).
Isolation of MNCs and flow cytometry
The MNCs were isolated from blood, spleen, duodenum, and ileum as previously described (22, 27). In brief, lymphoid tissues were collected aseptically and placed in ice-cold wash medium (RPMI 1640 with 10 mM HEPES, 200 μg/ml gentamicin, and 20 μg/ml ampicillin). Spleens were cut into small pieces and pressed through a stainless-steel 80-mesh screen of cell collector (E-C Apparatus, St. Petersburg, FL) to obtain single-cell suspensions. Intestinal tissues (duodenum = 15 cm and Ileum = 18 cm in length) were rinsed twice with Ca2+ and Mg2+ free HBSS and were then incubated with HBSS containing 10 mM DTT (Sigma, St. Louis, MO), 5 mM EDTA, 40 mM HEPES, and antibiotics on shaker at 37°C for 15 min to dislodge and remove mucus, epithelial cells, and intraepithelial cells. Subsequently, these segments were minced and further digested for 20 min at 37°C with RPMI 1640 containing 400 U/ml collagenase type II (Invitrogen/Life Technologies, New York, NY), 10 mM DTT, 5 mM EDTA, 20 mM HEPES, 8% FBS, and antibiotics. The supernatants (containing MNCs) and digested tissue were pressed through 80-mesh screens and single-cell suspensions were collected. The MNCs in cell suspensions from spleen and intestinal tissues were isolated by using 43–70% Percoll solution (Sigma) gradients. The MNCs from blood were isolated by using Ficoll-Paque solution (GE Healthcare, Uppsala, Sweden). After isolation, cells were resuspended in RPMI containing 8% FBS, 2 mM l-glutamine, 1 mM sodium pyruvate, 0.1 mM nonessential amino acids, 20 mM HEPES, and antibiotics (E-RPMI). The viability of each MNC preparation was determined by trypan blue dye exclusion (all were >90%). Fresh MNCs on the same day were stained for determining T cells (CD3+CD4+/CD3+CD8+), natural (CD4+CD25+Foxp3+/CD8+CD25+Foxp3+) and inducible (CD4+CD25−Foxp3+/CD8+CD25−Foxp3+) Treg subsets, and activated T cells (CD4+CD25+Foxp3−/CD8+CD25+Foxp3−).
T cells
To determine Th cells (CD3+CD4+) and cytotoxic T cells (CD3+CD8+), we stained each sample with anti-porcine CD3-FITC (clone PPT3; Southern Biotech, Birmingham, AL), CD4-PE (clone 74-12-4; BD Biosciences, San Jose, CA), and CD8-SPRD (clone 76-2-11; BD Biosciences) for 15 min at 4°C. The frequency of T cells was expressed as percentage among lymphocytes (determined based on forward and side-scatter characteristics).
Tregs and activated T cells
MNCs were stained with either CD4-PE or CD8-PE, and anti-porcine CD25 (Clone K231.3B2; Serotec, Raleigh, NC), followed by allophycocyanin-conjugated rat anti-mouse (RAM) IgG1 Ab (clone ×56; BD Biosciences). All samples were fixed and permeabilized with Cytofix/Cytoperm buffer (BD Biosciences) per manufacturer's instructions. RAM Foxp3-FITC Ab (clone FJK-16s; eBioscience, San Diego, CA) was incubated with MNCs for 45 min at 4°C. Appropriate isotype and secondary Ab controls were used for each reaction. After resuspending in perm/wash buffer (BD Biosciences), 100,000 events were acquired on Accuri C6 flow cytometer (BD, Franklin Lakes, NJ), and data were analyzed using C6 flow sampler software (BD). Frequency of Tregs was expressed as percentage among respective T cell subsets (CD4 or CD8) and among total MNCs (Fig. 3A, Supplemental Fig. 1A). Frequency of activated T cells was expressed as percentage among respective T cell subsets. The absolute number of Tregs (CD8+CD25+Foxp3+/CD8+CD25+Foxp3+) was determined by multiplying frequencies of Tregs among MNCs with total MNCs isolated from 50 ml blood, complete spleen, and measured sections of ileum and duodenum.
Figure 3.

Probiotic colonization enhanced systemic IFN-γ−producing T cells pre- and post-VirHRV challenge in vaccinated pigs. Representative dot plot of frequencies of CD3+CD4+IFN-γ+ and CD3+CD8+IFN-γ+ T cells among respective T cell subsets in RV and mock-stimulated (18 h at 37°C) splenic MNCs from the Vac+Pro group (A). For each cell population and tissue sample, IFN-γ−producing cells from mocks were subtracted from HRV Ag-stimulated samples. Mean frequencies (n = 4–7/group prechallenge and postchallenge) of IFN-γ−producing T cells (± SEM) prechallenge (B, CD3+CD4+ IFN-γ+; C, CD3+CD8+IFN-γ+) and post-VirHRV challenge (D, CD3+CD4+IFN-γ+; E, CD3+CD8+IFN-γ+) in blood, spleen, and ileum. Significant differences between groups are indicated by *p < 0.05, as determined by nonparametric Kruskal–Wallis rank sum test. Arrow indicates numerical increase (not significant) in CD8 IFN-γ−producing T cells in vaccinated groups compared with controls postchallenge. Control, Nonvaccinated and noncolonized control; Probiotics, probiotic colonized only.
HRV-specific IFN-γ−producing CD4 and CD8 T cells
Freshly processed MNCs from blood, spleen, and ileum were restimulated in vitro with semipurified HRV Ag (12 μg/ml) and porcine reactive anti-human CD49d mAb (0.5 μg/ml; clone 9F10; BD Pharmingen) for 18 h at 37°C in E-RPMI media as previously described (22, 28). Brefeldin A (10 μg/ml; Sigma), a protein transport inhibitor, was added for the last 5 h to prevent extracellular transfer of cytokines produced by the MNCs. Subsequently, stimulated MNCs were stained with CD3, CD8-FITC, and CD4-biotin followed by RAM IgG1-allophycocyanin and streptavidin PE-Cy7 (BD Biosciences) as secondary Abs. Samples were stained intracellularly with anti-porcine IFN-γ−PE Ab (clone P2G10; BD Biosciences). CD3+CD4+IFN-γ+ and CD3+CD8+IFN-γ+ cells were expressed as percentage of CD3+CD4+ and CD3+CD8+ T cells, respectively (Fig. 2A).
Figure 2.

Probiotics colonization enhanced intestinal Th cells and activated CD4 T cells prechallenge and systemic cytotoxic T cells postchallenge in vaccinated pigs. Mean frequencies (n = 4–7/group prechallenge and postchallenge) of CD3+CD4+ and CD3+CD8+ T cells among the lymphocytes (± SEM) in blood, spleen, ileum, and duodenum prechallenge and postchallenge (A–D). Mean frequencies of CD4+CD25+Foxp3− (prechallenge) (E) and CD8+ CD25+Foxp3− (postchallenge) (F) T cells among CD4+ and CD8+ T cells subsets (± SEM), respectively. Significant differences between groups are indicated by *p < 0.05, as determined by nonparametric Kruskal–Wallis rank sum test. Control, Nonvaccinated and noncolonized control; Probiotics, probiotic colonized only.
Detection of cytokines in serum, intestinal contents, and bile by ELISA
Blood was collected at multiple time points; PIDs 0, 2, 6, 14, 20, 27 (PCD0), 29 (PCD2), and 34 (PCD7), and intestinal contents (IC) were collected at euthanasia. Blood was spun at 1850 × g for 15 min, and serum was collected and stored at −20°C until tested. The IC collected were diluted 1:1 in MEM (Invitrogen/Life Technologies). Protease inhibitor mixture (Sigma) was added to the diluted IC and bile samples to prevent cytokine degradation. Bile and IC samples were centrifuged at 2000 × g, and supernatants were collected and stored at −20°C until tested. TGF-β, IL-4, IL-12, IFN-γ, and IL-6 were measured as previously described (18, 25). Porcine TNF-α ELISA Kit was used per manufacturer's recommendations (Thermoscientific Pierce, Rockford, IL). Nunc MaxiSorp 96-well plates were coated with anti-porcine IL-10 (4 μg/ml; Thermoscientific Fisher), IL-8 (0.2 μg/ml; Bethyl Laboratories, Montgomery, TX), IL-17 (0.5 μg/ml; Kingfisher Biotechnologies, St. Paul, MN), and IFN-α (clone K9, 2.5 μg/ml; PBL IFN Source, Piscataway, NJ). All coated plates were incubated overnight at 4°C except IFN-α (37°C). Biotinylated anti-porcine IL-10 (1 μg/ml), IL-8 (0.02 μg/ml), IL-17 (0.1 μg/ml), and IFN-α (clone F17, 2 μg/ml) Abs were used for detection. Porcine IFN-α detection Ab was biotinylated using a commercial kit (Thermoscientific Pierce) as per manufacturer's recommendations. Plates were developed and cytokine concentrations were calculated as previously described (18).
Statistical analysis
Statistical analysis was done by SAS version 9.3 (SAS Institute, Cary, NC). Mean RV fecal shedding, diarrhea scores, and duration of diarrhea were compared by one-way ANOVA (ANOVA-general linear model), followed by Duncan's multiple range test. The area under the curve (AUC) for diarrhea severity was calculated from diarrhea score-time graph of individual animals (29). Frequencies of cells determined by flow cytometry, absolute number of MNCs and Tregs, cytokine concentrations, and AUC for diarrhea severity between and within groups were compared using Kruskal–Wallis rank sum test.
Results
Probiotics reduce diarrhea severity and HRV fecal shedding post-VirHRV challenge
Diarrhea scores and HRV fecal shedding were recorded daily post-VirHRV challenge (Fig. 1). Vac+Pro group had significantly lower mean AUC (2.9 versus 5.0; Table I) and mean diarrhea scores at PCD3 and PCD5 compared with the Vac group, indicating increased clinical protection in Vac+Pro pigs post-VirHRV challenge. All 100% of Vac+Pro pigs were protected from virus shedding compared with 83% of Vac pigs (Table I). Probiotics only pigs had significantly lower mean duration of diarrhea (1.2 versus 3 d) and AUC (5.7 versus 8.5; Table I), and lower mean diarrhea scores from PCD4 through PCD7 compared with Control group, suggesting that early colonization by LGG+Bb12 moderates severity of VirHRV infection. Lower diarrhea scores coincided with delayed onset and significantly lower mean fecal HRV shedding from PCD4 through PCD7, and significantly lower average peak titers shed (977.5 versus 3343.8 FFU/ml; Table I) in the Probiotics only group compared with Control group. In both probiotic groups, LGG and Bb12 colonized small and large intestines both prechallenge and postchallenge (Supplemental Table I) and were shed fecally as reported previously (25). Postchallenge probiotic counts were significantly lower in cecum and colon of probiotic colonized groups compared with the prechallenge bacterial counts in the counterpart groups (Supplemental Table I).
Table I. Summary of HRV shedding and diarrhea scores in the four different treatment groups.
| Treatment | N | Virus Sheddinga | Protection Rate (%) against HRV Shedding | Diarrheab | ||
|---|---|---|---|---|---|---|
|
|
|
|||||
| Mean Days to Onset of Sheddingc | Average Peak Titer Shed (FFU/ml)c | Mean Duration, dc | AUCd | |||
| Vac+Pro | 7 | NAe | 12.5f,* | 100 | 0 | 2.9* |
| Vac | 6 | NA g | 32.3* | 83 | 0 | 5.0** |
| Probiotics | 5 | 5* | 977.5** | 0 | 1.2* | 5.7** |
| Control | 4 | 3.5** | 3343.8*** | 0 | 3** | 8.5*** |
Determined by cell culture immunofluorescence assay and expressed as FFU/ml.
Pigs with fecal score >1 were considered diarrheic. Fecal consistency was scored as follows: 0, normal; 1, pasty/semiliquid; and 2, liquid.
Means with different asterisks in the same column differ significantly (determined by one-way ANOVA followed by Duncan's multiple range test, p < 0.05).
AUC indicates diarrhea severity. Means in the same column, with different asterisks, differ significantly (determined by Kruskal–Wallis rank sum test, p < 0.05).
No virus shedding was observed.
Samples negative for HRV detection (<25) were assigned a titer value of 12.5 for statistical analysis.
Only 16% (1/6) of piglets shed virus, and determination of mean days to onset of virus shedding is not applicable (NA).
LGG+Bb12 colonization increased intestinal Th cells prechallenge and systemic cytotoxic T cells postchallenge in vaccinated pigs
The Vac+Pro group had significantly higher frequencies of duodenal and higher, although not significant (p = 0.07), ileal Th cells (CD3+CD4+) prechallenge compared with the Vac group (Fig. 2A). No differences in T cell subsets were observed between Vac and Vac+Pro groups in the intestinal tissues postchallenge. However, the Probiotics only group had higher duodenal Th cells prechallenge and significantly higher ileal cytotoxic T cells (CD3+ CD8+) postchallenge compared with Control pigs, which coincided with lower shedding and diarrhea scores in the LGG+Bb12 colonized only group (Fig. 2B, Table I). Vac+Pro had significantly higher systemic CD3+CD8+ (blood and spleen) and CD3+CD4+ T cells (spleen) postchallenge compared with the Vac group (Fig. 2D). These findings suggest that colonization by LGG+Bb12 may prime for cell-mediated immune responses to AttHRV vaccine and HRV infection.
Vaccination increased activated (CD25+Foxp3−) intestinal CD4 T cells prechallenge and decreased systemic activated CD8 T cells postchallenge
Vaccinated groups had higher frequencies of CD4+CD25+Foxp3− T cells in blood and ileum compared with unvaccinated groups at PID27 (Fig. 2E), suggesting that peripheral blood may reflect increase of intestinal activated CD4 T cells. Interestingly, probiotic colonization significantly enhanced CD4+CD25+Foxp3− T cells prechallenge in duodenum of the Vac+Pro group compared with the Vac group (Fig. 2E), which coincided with higher numbers of duodenal IgA Ab-secreting cells and intestinal IgA Ab titers in the Vac+Pro group compared with the Vac group postchallenge (Kandasamy, Saif et al., unpublished observations). No significant differences were observed for activated CD8+ T cells prechallenge in blood and tissues, and activated CD4+ T cells postchallenge in intestinal tissues among groups (Supplementary Fig. 1). Vaccinated pigs had significantly reduced systemic (blood and spleen) CD8+CD25+Foxp3− T cells compared with unvaccinated pigs at PID34/PCD7, indicating localized control of infection in intestinal tissues without systemic dissemination (Fig. 2F).
Probiotics colonization in vaccinated pigs increased systemic IFN-γ−producing T cells pre- and post-VirHRV challenge
IFN-γ−producing T cells were investigated pre- and post-VirHRV challenge. Vac+Pro group had higher frequencies of systemic (blood and spleen) CD3+CD4+IFN-γ+ and CD3+CD8+IFN-γ+ T cells at PID27/PCD0 compared with the Vac group (Fig. 3B, 3C), suggesting Th1-enhancing effects of probiotics on AttHRV vaccine. At PID34/PCD7, the Vac+Pro group had significantly higher frequencies of splenic CD3+CD4+IFN-γ+ (4.6 fold) and CD3+CD8+IFN-γ+ (10.8-fold), and a trend for higher frequencies of circulatory CD3+CD8+IFN-γ+ T cells compared with the Vac group (Fig. 3D, 3E). Interestingly, in ileum, probiotic colonization resulted in slight decrease in CD3+CD4+IFN-γ+ T cells in vaccinated pigs, suggesting different modulatory effects of probiotics on systemic versus mucosal responses.
Vaccination and/or probiotic colonization–induced intestinal CD25+Foxp3+ Tregs post-VirHRV challenge
CD25+Foxp3+ T cells. Mean frequencies of CD4+CD25+Foxp3+ T cells were higher in ileum and blood of Probiotics only group compared with Control group prechallenge suggesting a role of LGG+Bb12 in induction of CD4 Tregs (Fig. 4B). Interestingly, AttHRV also significantly enhanced CD4 Tregs in blood prechallenge, irrespective of probiotic colonization (Fig. 4B), which was independent of increase in intestinal CD4 Tregs.
Figure 4.

Probiotic colonization and/or vaccination increased ileal CD4+CD25+Foxp3+ Tregs post-VirHRV infection. Representative dot plot of frequencies of ileal CD4+CD25+Foxp3+ T cells among CD4 T cells from the Vac+Pro group (A). Mean frequencies (n = 4-7/group prechallenge and post-challenge) of CD4+CD25+Foxp3+ (B, prechallenge; C, postchallenge), CD8+CD25+Foxp3+ (D, prechallenge; E, postchallenge), and CD4+CD25−Foxp3+ T cells (F, prechallenge and postchallenge) ± SEM in blood, spleen, ileum, and duodenum. Significant differences between groups are indicated with *p < 0.05, as determined by nonparametric Kruskal−Wallis rank sum test. Control, Nonvaccinated and noncolonized control; Probiotics, probiotic colonized only.
Postchallenge, vaccinated groups had significantly higher frequencies of CD4+CD25+Foxp3+ and CD8+CD25+Foxp3+ cells in ileum and CD8+CD25+Foxp3+ cells in blood, as compared with the Control group (Fig. 4C, 4E, Supplementary Fig. 2C, 2E). The AttHRV vaccine-induced Treg responses post-VirHRV challenge coincided with lower diarrhea severity scores in these groups compared with the Control group. The absolute number of CD4+ CD25+Foxp3+ and CD8+CD25+Foxp3+ cells in blood and tissues followed the general trend observed for mean frequencies of CD4 and CD8 Tregs among MNCs and among respective CD4 and CD8 T cell subsets (Fig. 4C, 4E, Supplementary Figs. 2C, 2E, 3B, 3C). Similarly, the Probiotics only group also had higher frequencies of CD4 Tregs among ileal MNCs postchallenge as compared with the Control group (Fig. 4C, Supplemental Fig. 2C), suggesting the role of probiotics in inducing Treg responses after VirHRV infection to restore gut homeostasis and alleviate HRV-induced diarrhea. Contrary to the ileum, lower CD4 and CD8 Treg frequencies among MNCs were observed in spleen (systemically) of the Probiotics only group compared with the Control group (Supplemental Fig. 2C, 2E), coinciding with higher CD3+CD8+ IFN-γ+ T cell frequencies in blood (4.4-fold) and spleen (2.9-fold) at PID34/PCD7 in the former group (Fig. 3E).
Probiotic colonization in vaccinated pigs (Pro+Vac) enhanced systemic effector Th1 cells, but not CD4 and CD8 Tregs (Figs. 3D, 4C). The Pro+Vac group had higher systemic effector T cell (CD3+CD4+IFN-γ+ T cells)/Treg (CD4+CD25+Foxp3+) ratios prechallenge and postchallenge compared with the Vac group, suggesting the immunostimulating (Th1) potential of LGG+Bb12 for AttHRV vaccine (Fig. 5).
Figure 5.

Probiotics colonized and vaccinated (Vac+Pro) pigs had higher ratios of systemic effector/Treg responses compared with vaccinated group (Vac). Mean ratios (n = 4–7/group prechallenge and postchallenge) of CD3+CD4+IFN-γ+ cells as subset of CD4 T cell (effector T cells) and CD4+CD25+Foxp3+ cells (Tregs) among CD4 T cells prechallenge and postchallenge in blood, spleen, and ileum for different treatment groups. Control, Nonvaccinated and noncolonized control; Probiotics, probiotic colonized only.
CD4+CD25−Foxp3+ T cells in intestinal tissues
Probiotic only pigs had significantly higher frequencies of ileal CD4+CD25− Foxp3+ T cells in ileum prechallenge as compared with Control pigs, similar to that of CD4+CD25+Foxp3+ cells (Fig. 4F). Postchallenge, vaccinated pigs irrespective of probiotic colonization had lower mean frequencies of CD4+CD25−Foxp3+ T cells among CD4 T cells (Fig. 4F) and among MNCs (data not shown) in duodenum (effector site) as compared with control pigs, contrary to no effect of vaccination on duodenal CD4+CD25+Foxp3+ T cells. No association was found between reduced inducible CD4+CD25−Foxp3+ T cells and activated CD4 and CD8 T cells in duodenum.
Probiotics increased serum innate (IFN-α), Th1 (IFN-γ and IL-12), and reduced Th2 (IL-4) cytokine responses to oral AttHRV vaccine
Innate, Th1, Th2, Th17, and Treg cytokines were measured to determine type of immune response and its modulation by LGG+Bb12 colonization. Vaccinated groups had significantly higher serum IFN-γ, IL-12, and IFN-α cytokines at PID2, and IFN-γ at PID6 as compared with unvaccinated groups (Fig. 6A–C). The Pro+Vac group had significantly higher IFN-α (3-fold) at PID2, IFN-γ at PID14 (after second AttHRV inoculation), and IL-12 at PID6 as compared with Vac group, suggesting an adjuvant effect of probiotics on AttHRV vaccine. Moreover, the Pro+Vac group had significantly lower IL-4 at PID2 (after first vaccine) and PID20 (during third vaccine dose) as compared with the Vac group (Fig. 6D), suggesting deviation toward Th1 responses. Pro+Vac pigs reached mean peak for IL-4 levels at PID20, whereas Vac pigs reached peak earlier at PID6. Vaccinated pigs had lower serum Treg cytokine TGF-β (PID2) as compared with controls irrespective of probiotic colonization (Fig. 6E). Fourteen of 22 (64%) vaccinated pigs had decreased TGF-β at PID2 compared with PID0.
Figure 6.

LGG+Bb12 enhanced innate and Th1 cytokines, and reduced Th2 cytokine responses in vaccinated pigs prechallenge. Mean concentrations (± SEM) of IFN-γ and IL-12 (both Th1), IFN-α (innate), IL-4 (Th2), TGF-β (Treg), and IL-17 (Th17) in serum of pigs from different groups (A–F). Arrows indicate vaccine time points. Different alphabetical letters indicate significant differences (derived by nonparametric Kruskal–Wallis rank sum test, p < 0.05) at the same time point in cytokine concentrations among treatment groups. Cont, Nonvaccinated and noncolonized control; Pro, probiotic colonized only.
Probiotics decreased proinflammatory (TNF-α and IL-8) and Th17 (IL-17), and increased Treg (TGF-β) cytokine responses post-VirHRV challenge compared with noncolonized control pigs
Postchallenge, no significant differences were observed between Vac+Pro and Vac groups for most cytokines in serum except IL-12, which was significantly higher in the Vac+Pro group (PID34/PCD7; Fig. 7A–E). Probiotics only and Control groups had higher IFN-α, IL-12, and IL-8 (PCD2), and IL-17 (PCD7) compared with vaccinated pigs postchallenge. The Probiotics only group had significantly lower IL-8 (PCD2 and PCD7), IL-17 (PCD7), and higher TGF-β (PCD2) compared with the Control group (Fig. 7B–E), suggesting downregulation of inflammatory cytokines and induction of regulatory cytokines by LGG+Bb12. Higher IL-12 and IFN-α at PCD2 coincided with higher HRV shedding (replication) in Control as compared with Probiotics groups.
Figure 7.

LGG+Bb12 colonization reduced proinflammatory and innate cytokines, and maintained regulatory cytokine responses post-VirHRV challenge in nonvaccinated pigs. Mean concentrations (± SEM) of IL-12 (Th1), IFN-α, and IL-8 (innate), TGF-β (Treg), and IL-17 (Th17) in serum of pigs from different groups (A–E). Mean concentrations (± SEM) of TNF-α (proinflammatory), IL-12, and IL-17 in bile and small intestinal contents (SIC) of pigs from different groups (F–H). Different alphabetical letters in (A)–(E) indicate significant differences (derived by nonparametric Kruskal–Wallis rank sum test, p < 0.05) at the same time point among treatment groups. The * in (F)–(H) indicate significant differences (derived by nonparametric Kruskal–Wallis rank sum test, p < 0.05) between treatment groups for bile and SIC. Cont, Nonvaccinated and noncolonized control; Pro, probiotic colonized only.
We measured cytokine concentrations in the IC and bile to characterize local (gut) cytokine microenvironments. Most of the cytokines were unstable and thus undetectable as previously described (18). Vaccinated pigs had lower biliary TNF-α and IL-12, and significantly lower biliary IL-17 as compared with unvaccinated pigs at PID34/PCD7 (Fig. 7F–H). The Probiotics group had significantly lower IL-12 and lower TNF-α as compared with Controls (Fig. 7F, 7G), suggesting a role of LGG+Bb12 in reducing local (gut) inflammation caused by RV infection and pathology.
Discussion
Probiotic studies conducted in infants and children using vaccines have primarily focused on Ab responses (24), and few studies have addressed T cell responses. Using the relevant neonatal piglet model of HRV diarrhea, to our knowledge, ours is the first study to investigate modulation of T cell responses to RV vaccine and infection, by probiotics LGG and Bb12 that belong to commensal groups (Lactobacilli and Bifidobacteria) that are dominant in breastfed infants. Commensal colonization showed divergent effects on AttHRV vaccine and VirHRV infection: oral administration of LGG and Bb12 moderated RV infection because of their ability to increase intestinal Tregs, whereas they enhanced vaccine efficacy by increasing systemic Th1 and innate immune responses.
A major finding of our study is that early dual colonization with LGG and Bb12 can moderate the severity of RV infection. Similarly, previous studies in children have shown that LGG, when used for treatment of RV diarrhea, resulted in reduced duration of diarrhea (30–33). Moreover, prophylactic supplementation of certain probiotics (LGG, Bifidobacterium bifidum, etc.) in higher doses and over a period of time in children resulted in reduced incidence of RV infection (8) and RV gastroenteritis (9), although the mechanisms were not defined. Probiotics may directly reduce enteric infections and ailments by various mechanisms including competitively inhibiting binding by pathogens, stimulating gut epithelial cell proliferation and improving gut integrity, acidifying luminal microenvironment, modulating immune responses, and so forth (4, 34). In our study, early LGG+Bb12 colonization induced intestinal Tregs (CD25+Foxp3+) both pre- and post-VirHRV challenge in the Gn pig model for HRV infection. Increased Tregs and the associated higher serum TGF-β might have resulted in reduced proinflammatory serum IL-8 and IL-17, and biliary TNF-α and IL-12 cytokines, inducing an anti-inflammatory microenvironment, thereby reducing RV induced pathology or the ensuing exacerbated host immune responses (16–18). Recently, Rodrigues et al. (35) showed that pretreatment of neonatal mice with Lactobacillus helveticus and LGG induced Tregs and higher expression of regulatory and anti-inflammatory cytokines and lower expression of proinflammatory cytokines, which coincided with higher protection in these mice against the murine enteric pathogen Citrobacter rodentium. A combination of Lactobacilli and Bifidobacterium spp. prevented chemically induced colitis in mice (36) and alleviated clinical symptoms in patients with inflammatory bowel disease (37). In mice, protection was associated with induction of Tregs and reduction of inflammatory cytokines (IL-12, IFN-γ, TNF-α) (36). Moreover, in a randomized clinical trial involving healthy adults, probiotics LGG and Bb12 induced an anti-inflammatory environment indicated by lower serum C-reactive protein in vivo and lower TNF-α and IL-2 in vitro (38). Thus, early colonization by LGG+Bb12 in breastfed infants may establish gut homeostasis and reduce the severity of RV infection by induction of intestinal Tregs without suppressing systemic effector T cells (higher CD3+CD8+IFN-γ+ T cells in Probiotics only group versus Control, postchallenge). Similar to our study, induction of Tregs also reduced inflammation in other infectious conditions (39, 40).
The combination of LGG+Bb12 likely enhanced AttHRV vaccine efficacy by enhancing the frequency of intestinal CD4 T cells (prechallenge), systemic CD8 T cells (postchallenge), systemic CD4 and CD8 IFN-γ−producing T cells (prechallenge and postchallenge), and activated duodenal CD4 T cells prechallenge, all of which coincided with lower diarrhea severity scores in the Vac+Pro compared with the Vac group. IFN-γ−producing T cells are a correlate of protection to RV infection in pigs, and IFN-γsecreted by T cells after vaccination has been shown to have anti-RV activity in mice (22, 28, 41). Similar to pigs, HRV infection in children and adults is also known to increase IFN-γ−producing T cells in the peripheral blood (42, 43). Increased systemic IFN-γ−producing T cells and higher effector T cell/Treg ratios both prechallenge and postchallenge coincided with higher Th1 cytokines (prechallenge and postchallenge) and lower Th2 cytokine prechallenge, suggesting a systemic Th1 bias induced by the selected probiotics. The ratio of effector T cells to Tregs reflects IL-12/IL-10 cytokine ratios, which have been previously used to designate the type of immune response (Th1, Th2, or Treg) (44, 45). However, in this study, we did not investigate Th2 cytokine-producing cells per se and because of a technical limitation, we could not definitively type the gut immune responses based on cytokine levels (18). The early increased IFN-γ, IL-12, and IFN-α in the Vac+Pro compared with the Vac group coincides with higher stimulation of innate responses in this group, which affects subsequent development of adaptive T cell and Ab responses (46). Thus, the probiotics LGG+Bb12 increased effectiveness of AttHRV vaccine by increasing both innate and adaptive immune responses. Moreover, LGG+Bb12 colonization may prime for development of Th1 responses in infants, which are needed for protection from viral and intracellular bacterial infections. This is important because infants exhibit a bias toward Th2 responses (47).
In our study, probiotics mediated differential immunomodulation of the mucosal versus systemic immune system in vaccinated animals. In contrast with systemic tissues, no significant increase in intestinal IFN-γ−producing T cells and intestinal IL-12 was observed in the Vac+Pro group compared with the Vac group, although the former group had increased intestinal IgA Ab responses and IgA Ab-secreting cells, suggesting that intestinal B cell responses synergistically with increased systemic T cell (Th1) responses may have a played a role in clearing RV infection. The increased intestinal natural Tregs, along with slightly increased intestinal IFN-γ−producing effector T cells in vaccinated animals (irrespective of probiotics) compared with controls suggests that simultaneous expansion of these two subsets of T cells in the intestine may have been necessary to control RV infection and/or the associated tissue inflammation/pathology caused by RV. Similar to our study, IL-2–dependent expansion of Tregs and effector IFN-γ−producing CD4 and CD8 T cells occurred in lungs and was associated with resistance from tuberculosis infection in macaques (48). Recently, Wen et al. (49) suggested an important role for inducible Tregs in determining protection from HRV infection in the Gn pig model and showed that lower numbers of inducible Tregs coincided with higher protection rates. Similarly, we also show reduced inducible Tregs in vaccinated animals in duodenum postchallenge, which coincided with higher protection rates in vaccinated groups. However, in contrast with our findings, Wen et al. (49) showed lower numbers of intestinal natural Tregs in vaccinated animals compared with control animals postchallenge. The reasons for this discrepancy is unknown, but in our study, we vaccinated pigs with three doses of AttHRV (mimicking HRV vaccine [Rotarix] doses in children), whereas Wen et al. (49) used only two doses. Moreover, difference in pig breed and age of animals may have influenced these observations.
Modulation of the immune system by probiotics is bacterial strain and type specific, and can be attributed to differences in microbe-associated molecular patterns expressed by these bacteria (50). In vitro studies have shown differential modulation of cytokine responses and dendritic cell maturation by Lactobacilli and Bifidobacterium spp. and their products; however, these functions are not always replicated or comparable in vivo (50–52). This can be attributed to various contributing factors including host genetics, individual immune function, stress, age, species, nutritional status, diet, gut microbiome, experimental conditions, among others. Both LGG and Bb12 individually exhibited proinflammatory as well as anti-inflammatory responses under different conditions (44, 53–56). In our study, we showed differential effects of dual LGG+Bb12 cocolonization in HRV vaccinated and infected pigs, and suggest that interactions between probiotics and vaccine and/or infection and host immune response together derive different outcomes in vivo.
The role of CD8 Tregs in RV infection is unknown and, to our knowledge, this is the first study showing induction of these cells in AttHRV vaccinated pigs post-VirHRV challenge. CD8 Tregs were shown previously to be induced by both viral and bacterial infections (57, 58). Moreover, they have been shown to control SIV infection and type I diabetes (57, 59). Thus, in our study, both CD4 and CD8 Tregs may play synergistic roles in moderating RV infection in LGG+Bb12 colonized, vaccinated, and unvaccinated pigs. However, further studies are needed to elucidate the functional role of these Treg subsets.
We observed higher serum IL-17 in control animals, irrespective of probiotic colonization compared with vaccinated groups. A recent study of mice showed induction of IL-17–producing memory CD4 cells after mucosal administration of RV viral protein 6 with E. coli heat labile toxin or cholera toxin derivative, but IL-17 did not exhibit direct anti-RV activity in vitro (41, 60). Considering these findings and our results from control animals, it appears that higher IL-17 levels may indicate either recent exposure to RV infection or more severe clinical signs (diarrhea) similar to observations for influenza virus and respiratory syncytial virus infections (61, 62). IL-17 is not secreted exclusively by Th17 cells, but also by other innate cells such as γδ T cells, neutrophils, and NKT cells, which may have contributed to the increased serum IL-17 in our study (63).
Whether the findings from this study using Gn animals can be generalized to conventional pigs or other species containing normal microflora is speculative because of the reduced immune responses in germfree, monocolonized, or dual-colonized versus conventional animals (64, 65). Although the magnitude of the immune responses is lower in Gn compared with conventional pigs, the overall mucosal immune responses and their kinetics are similar. Neonatal Gn, like conventional pigs and infants, are immunocompetent at birth, and although immune responses are less than in adults, they develop protective immunity after recovery from HRV infection. Furthermore, the cytokine profile and correlates of protection to HRV diarrhea between Gn pigs and infants are similar (18, 66, 67). Ongoing studies in our laboratory will delineate the impact of these probiotics in pigs colonized with a commensal mixture and vaccinated and/or challenged with VirHRV. Previous studies of germfree mice monocolonized with a single dose of B. bifidum and conventional mice supplemented with multiple and higher doses of B. bifidum showed similar survival and histological lesions after Salmonella enteritidis subspecies typhimurium challenge, suggesting that supplementation of a combination of LGG and Bb12 may be effective in the presence of commensals (68) as documented in RV-infected infants (30– 33). Our previous studies and those of others have shown that Gn animal models are advantageous to investigate the mechanisms underlying host-commensal/probiotic strain-specific interactions such as antagonism and synergism with HRV or other infections, and then after deciphering their immunostimulatory or immunoregulatory effects, tailor them to improve therapies or vaccine efficacies in humans (21–23, 68). In our study, use of LGG and Bb12 combined may have masked their differential immunomodulating effects on gut and systemic immune responses. Further studies are needed to investigate how these probiotics individually affect HRV-specific protective immune responses. In summary, we show that early neonatal colonization by LGG+Bb12 not only moderated HRV infection, but also was immunostimulatory for an oral AttHRV vaccine. These prophylactic and adjuvant properties are at least partially mediated through induction of innate and adaptive immune responses as described.
Supplementary Material
Acknowledgments
This work was supported by the National Center for Complementary and Alternative Medicine, National Institutes of Health (Grant R21 AT004716 to L.J.S.); National Institute of Allergy and Infectious Diseases, National Institutes of Health (Grant R01 A1099451 to L.J.S.); and federal funds appropriated to the Ohio Agricultural Research and Development Center, The Ohio State University.
We thank Dr. Juliette Hanson, Rich McCormick, and Lindsey Good for technical assistance. We thank Ozkan Timurkan, Joshua Amimo, Kyle T. Scheuer, Ning Chen, and Zhe Liu for assistance with animal work, sample collection, and processing. We also thank Dr. Gloria Solano-Aguilar, U.S. Department of Agriculture, for providing primers and probes specific for LGG.
Abbreviations
- AttHRV
attenuated human rotavirus
- AUC
area under the curve
- Bb12
Bifidobacterium lactis Bb12
- FFU
fluorescent foci forming unit
- Gn
gnotobiotic
- HRV
human rotavirus
- IC
intestinal contents
- LGG
Lactobacillus rhamnosus strain GG
- MNC
mononuclear cell
- PCD
postchallenge day
- PID
postinoculation day
- Pro+Vac
probiotic colonization in vaccinated pigs
- RAM
rat anti-mouse
- RV
rotavirus
- Treg
T regulatory cell
- Vac
3XAttHRV Wa vaccinated only group
- Vac+Pro
3XAttHRV Wa vaccinated and probiotic colonized group
- VirHRV
virulent human rotavirus
Footnotes
The online version of this article contains supplemental material.
Disclosures: The authors have no financial conflicts of interest.
References
- 1.Tate JE, Burton AH, Boschi-Pinto C, Steele AD, Duque J, Parashar UD. WHO-coordinated Global Rotavirus Surveillance Network. 2012. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: a systematic review and meta-analysis. Lancet Infect Dis. 12:136–141. doi: 10.1016/S1473-3099(11)70253-5. [DOI] [PubMed] [Google Scholar]
- 2.Gray J. Rotavirus vaccines: safety, efficacy and public health impact. J Intern Med. 2011;270:206–214. doi: 10.1111/j.1365-2796.2011.02409.x. [DOI] [PubMed] [Google Scholar]
- 3.Licciardi PV, Tang ML. Vaccine adjuvant properties of probiotic bacteria. Discov Med. 2011;12:525–533. [PubMed] [Google Scholar]
- 4.Preidis GA, Hill C, Guerrant RL, Ramakrishna BS, Tannock GW, Versalovic J. Probiotics, enteric and diarrheal diseases, and global health. Gastroenterology. 2011;140:8–14. doi: 10.1053/j.gastro.2010.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fanaro S, Chierici R, Guerrini P, Vigi V. Intestinal microflora in early infancy: composition and development. Acta Paediatr Suppl. 2003;91:48–55. doi: 10.1111/j.1651-2227.2003.tb00646.x. [DOI] [PubMed] [Google Scholar]
- 6.Rinne M, Kalliomaki M, Arvilommi H, Salminen S, Isolauri E. Effect of probiotics and breastfeeding on the bifidobacterium and lactobacillus/enterococcus microbiota and humoral immune responses. J Pediatr. 2005;147:186–191. doi: 10.1016/j.jpeds.2005.03.053. [DOI] [PubMed] [Google Scholar]
- 7.Sjogren YM, Tomicic S, Lundberg A, Bottcher MF, Bjorksten B, Sverremark-Ekstrom E, Jenmalm MC. Influence of early gut microbiota on the maturation of childhood mucosal and systemic immune responses. Clin Exp Allergy. 2009;39:1842–1851. doi: 10.1111/j.1365-2222.2009.03326.x. [DOI] [PubMed] [Google Scholar]
- 8.Saavedra JM, Bauman NA, Oung I, Perman JA, Yolken RH. Feeding of Bifidobacterium bifidum and Streptococcus thermophilus to infants in hospital for prevention of diarrhoea and shedding of rotavirus. Lancet. 1994;344:1046–1049. doi: 10.1016/s0140-6736(94)91708-6. [DOI] [PubMed] [Google Scholar]
- 9.Szajewska H, Kotowska M, Mrukowicz JZ, Armańska M, Mikołajczyk W. Efficacy of Lactobacillus GG in prevention of nosocomial diarrhea in infants. J Pediatr. 2001;138:361–365. doi: 10.1067/mpd.2001.111321. [DOI] [PubMed] [Google Scholar]
- 10.Delcenserie V, Martel D, Lamoureux M, Amiot J, Boutin Y, Roy D. Immunomodulatory effects of probiotics in the intestinal tract. Curr Issues Mol Biol. 2008;10:37–54. [PubMed] [Google Scholar]
- 11.Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science. 2012;336:1268–1273. doi: 10.1126/science.1223490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, Cheng G, Yamasaki S, Saito T, Ohba Y, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 2011;331:337–341. doi: 10.1126/science.1198469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ivanov II, Atarashi K, Manel N, Brodie EL, Shima T, Karaoz U, Wei D, Goldfarb KC, Santee CA, Lynch SV, et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139:485–498. doi: 10.1016/j.cell.2009.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cho I, Yamanishi S, Cox L, Methé BA, Zavadil J, Li K, Gao Z, Mahana D, Raju K, Teitler I, et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature. 2012;488:621–626. doi: 10.1038/nature11400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ward LA, Rosen BI, Yuan L, Saif LJ. Pathogenesis of an attenuated and a virulent strain of group A human rotavirus in neonatal gnotobiotic pigs. J Gen Virol. 1996;77:1431–1441. doi: 10.1099/0022-1317-77-7-1431. [DOI] [PubMed] [Google Scholar]
- 16.Azim T, Ahmad SM, Sefat-E-Khuda, Sarker MS, Unicomb LE, De S, Hamadani JD, Salam MA, Wahed MA, Albert MJ. Immune response of children who develop persistent diarrhea following rotavirus infection. Clin Diagn Lab Immunol. 1999;6:690–695. doi: 10.1128/cdli.6.5.690-695.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jiang B, Snipes-Magaldi L, Dennehy P, Keyserling H, Holman RC, Bresee J, Gentsch J, Glass RI. Cytokines as mediators for or effectors against rotavirus disease in children. Clin Diagn Lab Immunol. 2003;10:995–1001. doi: 10.1128/CDLI.10.6.995-1001.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Azevedo MS, Yuan L, Pouly S, Gonzales AM, Jeong KI, Nguyen TV, Saif LJ. Cytokine responses in gnotobiotic pigs after infection with virulent or attenuated human rotavirus. J Virol. 2006;80:372–382. doi: 10.1128/JVI.80.1.372-382.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Xu J, Yang Y, Sun JE, Ding YZ, Su LY, Shao CH. Expression of Toll-like receptors in mononuclear cells from children with acute rotavirus diarrhea. Chinese Journal of experimental and clinical virology. 2007;21:38–40. [PubMed] [Google Scholar]
- 20.Wen K, Azevedo MS, Gonzalez A, Zhang W, Saif LJ, Li G, Yousef A, Yuan L. Toll-like receptor and innate cytokine responses induced by lactobacilli colonization and human rotavirus infection in gnotobiotic pigs. Vet Immunol Immunopathol. 2009;127:304–315. doi: 10.1016/j.vetimm.2008.10.322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wen K, Li G, Bui T, Liu F, Li Y, Kocher J, Lin L, Yang X, Yuan L. High dose and low dose Lactobacillus acidophilus exerted differential immune modulating effects on T cell immune responses induced by an oral human rotavirus vaccine in gnotobiotic pigs. Vaccine. 2012;30:1198–1207. doi: 10.1016/j.vaccine.2011.11.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang W, Azevedo MS, Wen K, Gonzalez A, Saif LJ, Li G, Yousef AE, Yuan L. Probiotic Lactobacillus acidophilus enhances the immunogenicity of an oral rotavirus vaccine in gnotobiotic pigs. Vaccine. 2008;26:3655–3661. doi: 10.1016/j.vaccine.2008.04.070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang W, Azevedo MS, Gonzalez AM, Saif LJ, Van Nguyen T, Wen K, Yousef AE, Yuan L. Influence of probiotic Lactobacilli colonization on neonatal B cell responses in a gnotobiotic pig model of human rotavirus infection and disease. Vet Immunol Immunopathol. 2008;122:175–181. doi: 10.1016/j.vetimm.2007.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Maidens C, Childs C, Przemska A, Dayel IB, Yaqoob P. Modulation of vaccine response by concomitant probiotic administration. Br J Clin Pharmacol. 2013;75:663–670. doi: 10.1111/j.1365-2125.2012.04404.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chattha KS, Vlasova AN, Kandasamy S, Esseili MA, Siegismund C, Rajashekara G, Saif LJ. Probiotics and colostrum/milk differentially affect neonatal humoral immune responses to oral rotavirus vaccine. Vaccine. 2013;31:1916–1923. doi: 10.1016/j.vaccine.2013.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Meyer RC, Bohl EH, Kohler EM. Procurement and maintenance of germ-free seine for microbiological investigations. Appl Microbiol. 1964;12:295–300. doi: 10.1128/am.12.4.295-300.1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yuan L, Ward LA, Rosen BI, To TL, Saif LJ. Systematic and intestinal antibody-secreting cell responses and correlates of protective immunity to human rotavirus in a gnotobiotic pig model of disease. J Virol. 1996;70:3075–3083. doi: 10.1128/jvi.70.5.3075-3083.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yuan L, Wen K, Azevedo MS, Gonzalez AM, Zhang W, Saif LJ. Virus-specific intestinal IFN-gamma producing T cell responses induced by human rotavirus infection and vaccines are correlated with protection against rotavirus diarrhea in gnotobiotic pigs. Vaccine. 2008;26:3322–3331. doi: 10.1016/j.vaccine.2008.03.085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pérez-Cano FJ, Marín-Gallén S, Castell M, Rodríguez-Palmero M, Rivero M, Castellote C, Franch A. Supplementing suckling rats with whey protein concentrate modulates the immune response and ameliorates rat rotavirus-induced diarrhea. J Nutr. 2008;138:2392–2398. doi: 10.3945/jn.108.093856. [DOI] [PubMed] [Google Scholar]
- 30.Guandalini S, Pensabene L, Zikri MA, Dias JA, Casali LG, Hoekstra H, Kolacek S, Massar K, Micetic-Turk D, Papadopoulou A, et al. Lactobacillus GG administered in oral rehydration solution to children with acute diarrhea: a multicenter European trial. J Pediatr Gastroenterol Nutr. 2000;30:54–60. doi: 10.1097/00005176-200001000-00018. [DOI] [PubMed] [Google Scholar]
- 31.Guarino A, Canani RB, Spagnuolo MI, Albano F, Di Benedetto L. Oral bacterial therapy reduces the duration of symptoms and of viral excretion in children with mild diarrhea. J Pediatr Gastroenterol Nutr. 1997;25:516–519. doi: 10.1097/00005176-199711000-00005. [DOI] [PubMed] [Google Scholar]
- 32.Isolauri E, Juntunen M, Rautanen T, Sillanaukee P, Koivula T. A human Lactobacillus strain (Lactobacillus casei sp strain GG) promotes recovery from acute diarrhea in children. Pediatrics. 1991;88:90–97. [PubMed] [Google Scholar]
- 33.Rosenfeldt V, Michaelsen KF, Jakobsen M, Larsen CN, Møller PL, Tvede M, Weyrehter H, Valerius NH, Paerregaard A. Effect of probiotic Lactobacillus strains on acute diarrhea in a cohort of nonhospitalized children attending day-care centers. Pediatr Infect Dis J. 2002;21:417–419. doi: 10.1097/00006454-200205000-00013. [DOI] [PubMed] [Google Scholar]
- 34.Sherman PM, Ossa JC, Johnson-Henry K. Unraveling mechanisms of action of probiotics. Nutr Clin Pract. 2009;24:10–14. doi: 10.1177/0884533608329231. [DOI] [PubMed] [Google Scholar]
- 35.Rodrigues DM, Sousa AJ, Johnson-Henry KC, Sherman PM, Gareau MG. Probiotics are effective for the prevention and treatment of Citrobacter rodentium-induced colitis in mice. J Infect Dis. 2012;206:99–109. doi: 10.1093/infdis/jis177. [DOI] [PubMed] [Google Scholar]
- 36.Roselli M, Finamore A, Nuccitelli S, Carnevali P, Brigidi P, Vitali B, Nobili F, Rami R, Garaguso I, Mengheri E. Prevention of TNBS-induced colitis by different Lactobacillus and Bifidobacterium strains is associated with an expansion of gammadeltaT and regulatory T cells of intestinal intraepithelial lymphocytes. Inflamm Bowel Dis. 2009;15:1526–1536. doi: 10.1002/ibd.20961. [DOI] [PubMed] [Google Scholar]
- 37.Kajander K, Hatakka K, Poussa T, Färkkilä M, Korpela R. A probiotic mixture alleviates symptoms in irritable bowel syndrome patients: a controlled 6-month intervention. Aliment Pharmacol Ther. 2005;22:387–394. doi: 10.1111/j.1365-2036.2005.02579.x. [DOI] [PubMed] [Google Scholar]
- 38.Kekkonen RA, Lummela N, Karjalainen H, Latvala S, Tynkkynen S, Jarvenpaa S, Kautiainen H, Julkunen I, Vapaatalo H, Korpela R. Probiotic intervention has strain-specific anti-inflammatory effects in healthy adults. World J Gastroenterol. 2008;14:2029–2036. doi: 10.3748/wjg.14.2029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.O'Mahony C, Scully P, O'Mahony D, Murphy S, O'Brien F, Lyons A, Sherlock G, MacSharry J, Kiely B, Shanahan F, O'Mahony L. Commensal-induced regulatory T cells mediate protection against pathogen-stimulated NF-kappaB activation. PLoS Pathog. 2008;4:e1000112. doi: 10.1371/journal.ppat.1000112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhong Y, Cantwell A, Dube PH. Transforming growth factor beta and CD25 are important for controlling systemic dissemination following Yersinia enterocolitica infection of the gut. Infect Immun. 2010;78:3716–3725. doi: 10.1128/IAI.00203-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.McNeal MM, Stone SC, Basu M, Clements JD, Choi AH, Ward RL. IFN-gamma is the only anti-rotavirus cytokine found after in vitro stimulation of memory CD4+ T cells from mice immunized with a chimeric VP6 protein. Viral Immunol. 2007;20:571–584. doi: 10.1089/vim.2007.0055. [DOI] [PubMed] [Google Scholar]
- 42.Jaimes MC, Rojas OL, González AM, Cajiao I, Charpilienne A, Pothier P, Kohli E, Greenberg HB, Franco MA, Angel J. Frequencies of virus-specific CD4(+) and CD8(+) T lymphocytes secreting gamma interferon after acute natural rotavirus infection in children and adults. J Virol. 2002;76:4741–4749. doi: 10.1128/JVI.76.10.4741-4749.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mesa MC, Gutiérrez L, Duarte-Rey C, Angel J, Franco MA. A TGF-beta mediated regulatory mechanism modulates the T cell immune response to rotavirus in adults but not in children. Virology. 2010;399:77–86. doi: 10.1016/j.virol.2009.12.016. [DOI] [PubMed] [Google Scholar]
- 44.Dong H, Rowland I, Yaqoob P. Comparative effects of six probiotic strains on immune function in vitro. Br J Nutr. 2012;108:459–470. doi: 10.1017/S0007114511005824. [DOI] [PubMed] [Google Scholar]
- 45.Kaji R, Kiyoshima-Shibata J, Nagaoka M, Nanno M, Shida K. Bacterial teichoic acids reverse predominant IL-12 production induced by certain lactobacillus strains into predominant IL-10 production via TLR2-dependent ERK activation in macrophages. J Immunol. 2010;184:3505–3513. doi: 10.4049/jimmunol.0901569. [DOI] [PubMed] [Google Scholar]
- 46.Medzhitov R, Janeway CA., Jr Innate immunity: impact on the adaptive immune response. Curr Opin Immunol. 1997;9:4–9. doi: 10.1016/s0952-7915(97)80152-5. [DOI] [PubMed] [Google Scholar]
- 47.Zaghouani H, Hoeman CM, Adkins B. Neonatal immunity: faulty T-helpers and the shortcomings of dendritic cells. Trends Immunol. 2009;30:585–591. doi: 10.1016/j.it.2009.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chen CY, Huang D, Yao S, Halliday L, Zeng G, Wang RC, Chen ZW. IL-2 simultaneously expands Foxp3+ T regulatory and T effector cells and confers resistance to severe tuberculosis (TB): implicative Treg-T effector cooperation in immunity to TB. J Immunol. 2012;188:4278–4288. doi: 10.4049/jimmunol.1101291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wen K, Li G, Yang X, Bui T, Bai M, Liu F, Kocher J, Yuan L. CD4+ CD25- FoxP3+ regulatory cells are the predominant responding regulatory T cells after human rotavirus infection or vaccination in gnotobiotic pigs. Immunology. 2012;137:160–171. doi: 10.1111/j.1365-2567.2012.03617.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Meijerink M, Wells JM. Probiotic modulation of dendritic cells and T cell responses in the intestine. Benef Microbes. 2010;1:317–326. doi: 10.3920/BM2010.0029. [DOI] [PubMed] [Google Scholar]
- 51.Flinterman AE, Knol EF, van Ieperen-van Dijk AG, Timmerman HM, Knulst AC, Bruijnzeel-Koomen CA, Pasmans SG, van Hoffen E. Probiotics have a different immunomodulatory potential in vitro versus ex vivo upon oral administration in children with food allergy. Int Arch Allergy Immunol. 2007;143:237–244. doi: 10.1159/000099467. [DOI] [PubMed] [Google Scholar]
- 52.O'Flaherty S, Saulnier DM, Pot B, Versalovic J. How can probiotics and prebiotics impact mucosal immunity? Gut Microbes. 2010;1:293–300. doi: 10.4161/gmic.1.5.12924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Imaoka A, Shima T, Kato K, Mizuno S, Uehara T, Matsumoto S, Setoyama H, Hara T, Umesaki Y. Anti-inflammatory activity of probiotic Bifidobacterium: enhancement of IL-10 production in peripheral blood mononuclear cells from ulcerative colitis patients and inhibition of IL-8 secretion in HT-29 cells. World J Gastroenterol. 2008;14:2511–2516. doi: 10.3748/wjg.14.2511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.López P, González-Rodríguez I, Gueimonde M, Margolles A, Suárez A. Immune response to Bifidobacterium bifidum strains support Treg/Th17 plasticity. PLoS ONE. 2011;6:e24776. doi: 10.1371/journal.pone.0024776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Pohjavuori E, Viljanen M, Korpela R, Kuitunen M, Tiittanen M, Vaarala O, Savilahti E. Lactobacillus GG effect in increasing IFN-gamma production in infants with cow's milk allergy. J Allergy Clin Immunol. 2004;114:131–136. doi: 10.1016/j.jaci.2004.03.036. [DOI] [PubMed] [Google Scholar]
- 56.Ruiz PA, Hoffmann M, Szcesny S, Blaut M, Haller D. Innate mechanisms for Bifidobacterium lactis to activate transient pro-inflammatory host responses in intestinal epithelial cells after the colonization of germ-free rats. Immunology. 2005;115:441–450. doi: 10.1111/j.1365-2567.2005.02176.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lu W, Chen S, Lai C, Guo W, Fu L, Andrieu JM. Induction of CD8(+) regulatory T cells protects macaques against SIV challenge. Cell Rep. 2012;2:1736–1746. doi: 10.1016/j.celrep.2012.11.016. [DOI] [PubMed] [Google Scholar]
- 58.Silva-Campa E, Mata-Haro V, Mateu E, Hernández J. Porcine reproductive and respiratory syndrome virus induces CD4+CD8+CD25+Foxp3+regulatory T cells (Tregs) Virology. 2012;430:73–80. doi: 10.1016/j.virol.2012.04.009. [DOI] [PubMed] [Google Scholar]
- 59.Jiang H, Canfield SM, Gallagher MP, Jiang HH, Jiang Y, Zheng Z, Chess L. HLA-E-restricted regulatory CD8(+) T cells are involved in development and control of human autoimmune type 1 diabetes. J Clin Invest. 2010;120:3641–3650. doi: 10.1172/JCI43522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.McNeal MM, Basu M, Bean JA, Clements JD, Lycke NY, Ramne A, Löwenadler B, Choi AH, Ward RL. Intrarectal immunization of mice with VP6 and either LT(R192G) or CTA1-DD as adjuvant protects against fecal rotavirus shedding after EDIM challenge. Vaccine. 2007;25:6224–6231. doi: 10.1016/j.vaccine.2007.05.065. [DOI] [PubMed] [Google Scholar]
- 61.Crowe CR, Chen K, Pociask DA, Alcorn JF, Krivich C, Enelow RI, Ross TM, Witztum JL, Kolls JK. Critical role of IL-17RA in immunopathology of influenza infection. J Immunol. 2009;183:5301–5310. doi: 10.4049/jimmunol.0900995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ryzhakov G, Lai CC, Blazek K, To KW, Hussell T, Udalova I. IL-17 boosts proinflammatory outcome of antiviral response in human cells. J Immunol. 2011;187:5357–5362. doi: 10.4049/jimmunol.1100917. [DOI] [PubMed] [Google Scholar]
- 63.Gaffen SL. Structure and signalling in the IL-17 receptor family. Nat Rev Immunol. 2009;9:556–567. doi: 10.1038/nri2586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Round JL, Mazmanian SK. The gut microbiota shapes intestinal immune responses during health and disease. Nat Rev Immunol. 2009;9:313–323. doi: 10.1038/nri2515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Smith K, McCoy KD, Macpherson AJ. Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin Immunol. 2007;19:59–69. doi: 10.1016/j.smim.2006.10.002. [DOI] [PubMed] [Google Scholar]
- 66.Desselberger U, Huppertz HI. Immune responses to rotavirus infection and vaccination and associated correlates of protection. J Infect Dis. 2011;203:188–195. doi: 10.1093/infdis/jiq031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Yuan L, Saif LJ. Induction of mucosal immune responses and protection against enteric viruses: rotavirus infection of gnotobiotic pigs as a model. Vet Immunol Immunopathol. 2002;87:147–160. doi: 10.1016/S0165-2427(02)00046-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Silva AM, Bambirra EA, Oliveira AL, Souza PP, Gomes DA, Vieira EC, Nicoli JR. Protective effect of bifidus milk on the experimental infection with Salmonella enteritidis subsp. typhimurium in conventional and gnotobiotic mice. J Appl Microbiol. 1999;86:331–336. doi: 10.1046/j.1365-2672.1999.00674.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.