

Abstract

Polyhydroxybutyrate (PHB) synthases catalyze the polymerization of 3-(R)-hydroxybutyrate coenzyme A (HBCoA) to produce polyoxoesters of 1–2 MDa. A substrate analogue HBCH2CoA, in which the S in HBCoA is replaced with a CH2 group, was synthesized in 13 steps using a chemoenzymatic approach in a 7.5% overall yield. Kinetic studies reveal it is a competitive inhibitor of a class I and a class III PHB synthases, with Kis of 40 and 14 μM, respectively. To probe the elongation steps of the polymerization, HBCH2CoA was incubated with a synthase acylated with a [3H]-saturated trimer-CoA ([3H]-sTCoA). The products of the reaction were shown to be the methylene analogue of [3H]-sTCoA ([3H]-sT-CH2-CoA), saturated dimer-([3H]-sD-CO2H), and trimer-acid ([3H]-sT-CO2H), distinct from the expected methylene analogue of [3H]-saturated tetramer-CoA ([3H]-sTet-CH2-CoA). Detection of [3H]-sT-CH2-CoA and its slow rate of formation suggest that HBCH2CoA may be reporting on the termination and repriming process of the synthases, rather than elongation.
Polyhydroxyalkanoates (PHAs) are carbon and energy storage polymers synthesized by a variety of bacteria when they find themselves limited in an essential nutrient such as nitrogen or phosphorus but with an abundant carbon source available.1 Under conditions of maximum accumulation, PHAs can constitute up to 90% of the cell dry weight.2 When the limiting nutrient is restored, the bacteria degrade the PHAs to release energy and monomers for other biological processes.3,4 PHAs are of general interest as they can provide a biodegradable alternative to environmentally unfriendly, petroleum-based plastics.5,6
PHA synthases (PhaCs) catalyze the polymerization of 3-(R)-hydroxyalkanoate coenzyme A (CoA) to form PHA. They have been divided into four classes based on their substrate specificity, subunit composition, and molecular weight.7 Class I and III PhaCs utilize short-chain-length monomers such as 3-(R)-hydroxybutyrate (HB) coenzyme A (HBCoA) to generate polyhydroxybutyrate (PHB) polymers. The synthase from Ralstonia eutropha (PhaCRe) is the prototypical class I enzyme and is a dimer, composed of two 65 kDa subunits.8 The synthase from Allochromatium vinosum (PhaCAv) is the prototypical class III synthase and is a tetramer of two ∼40 kDa subunits: PhaC (the synthase) and PhaE (a protein of unknown function that is essential for activity).9,10 Studying the mechanism of polymerization (the priming and initiation, elongation, and termination) has been challenging because the elongation process is much faster than the initiation process.8,11 Since PhaCs are largely responsible for determining the molecular weight and polydispersity of the PHA, understanding the PhaC mechanism can contribute to the ultimate goal of producing PHA products in an economically competitive fashion.
Two models have been proposed for the polymerization catalyzed by PhaCs and involve the covalent catalysis by a Cys-His dyad and general base catalysis of an Asp to activate the 3-hydroxyl of a second HBCoA for ester bond formation.1,12 The model shown in Scheme 1A is based on analogy to fatty acid synthases13 and predicts that the growing PHB chain is always covalently attached to the protein and alternates between the two PhaC monomers.8,14−16 This model requires that the active site sits on the interface of two monomers in order to facilitate chain transfer, which is contradictory to the results from structural study that indicated the active site should be deeply buried.17 Thus, an alternative model based on the type III polyketide synthases18 is shown in Scheme 1B. It predicts that addition of the second HBCoA will generate a noncovalently bound (HB)2CoA that then rapidly reacylate the Cys.1,12 Detection of noncovalent intermediates will provide an effective way to distinguish between the two models.
Scheme 1. Two Mechanistic Models for Chain Elongation Catalyzed by PhaCs.

Our recent investigation with a mutant of PhaECAv in which the active site cysteine was replaced with a serine revealed noncovalent intermediates (dimers and trimers) providing evidence in support of the model in Scheme 1B.11 However, the turnover of this mutant was 5 × 10 –4 the rate of the wild-type (wt) PhaECAv. The slow rate suggests that additional mechanistic links to the wt enzyme need to be established. To obtain further evidence for the model shown in Scheme 1B, HBCH2CoA 1 (Scheme 2), an analogue of HBCoA in which the S of the thioester is replaced with a CH2 group, was designed. Our hypothesis was 1 could be incorporated into the growing PHB chain, reporting on the chain elongation process and resulting in chain termination.
Scheme 2. Chemoenzymatic Synthesis of Methylene Analogue HBCH2CoA 1.

Results and Discussion
Chemoenzymatic Synthesis of HBCH2CoA
Preparation of HBCH2CoA 1 using a chemoenzymatic approach is shown in Scheme 2. Synthesis of the acid 5 has been previously reported.19 Although the method is convenient, traces of byproduct tert-butyldimethylsilyl (TBDMS) alcohol lowered the yield of the next coupling reaction. Therefore, a different procedure of protecting carboxylic and hydroxyl groups, respectively, with benzyl (Bn) and TBDMS followed by hydrogenation was developed for acid 5. Formation of the Weinreb amide 6 was achieved using a standard coupling condition.20 The terminal alcohol 7 was obtained through the addition of Grignard reagent generated in situ from trimethylene chlorohydrin.21−23 Treatment of 7 with methansulfonyl chloride (MsCl) followed by sodium azide resulted in the azide 9.24 Various methods have been reported to reduce the organic azides to amines, in which the Staudinger reduction is the most widely used one.25−27 However, efforts to purify the amine produced in high yield based on TLC were not successful, presumably because during concentration of 10, it underwent intramolecular cyclization to form a cyclic imine. Therefore, we decided to perform the reduction without purification and concentration using Pd/C-catalyzed hydrogenation in methylene chloride.28 After filtration of the catalyst, the mixture was directly reacted with protected pantothenic acid 11 to give HB-carba(dethio)pantetheine 12. The precursor 13 for enzymatic transformation was obtained after treating 12 with 4% HF in CH3CN for 15 min at 0 °C. It has to be pointed out that the yield from 12 to 13 was very low if the diol in 12 was protected with benzylidene. Compound 13 was obtained in 10 steps in a total 15% yield from 2.
Preparation of HBCH2CoA 1 was achieved by use of three enzymes involved in CoA biosynthesis: a pantothenate kinase from Staphylococcus aureus (SaPanK),29 a phosphopantetheine adenylyltransferase (EcCoaD),30 and a dephospho-CoA kinase (EcCoaE)31 from Escherichia coli. Although our knowledge of the substrate specificity of these enzymes remains incomplete, previous work has established that they can accept a wide spectrum of pantothenate-based substrates.32−36 It has been demonstrated that the key regulatory enzyme in CoA biosynthesis from E. coli is the pantothenate kinase (EcPanK) due to its feedback inhibition by CoA and/or its thioesters.37 This feedback inhibition will limit the conversion of pantothenate analogues to CoA analogues and thus cause low yields. However, in contrast to all known pantothenate kinases, SaPanK is not feedback-regulated by CoA and/or its thioesters,29 which will allow the accumulation of CoA. Therefore, in order to maximize the conversion yield of enzymatic precursor 13 to HBCH2CoA 1, SaPanK was employed for the first phosphorylation step in our reaction. An HPLC method was developed to monitor the progress of enzymatic conversions as depicted in Supplementary Figure S1. The retention times for enzymatic precursor 13, 4′-phospho derivative 14, 3′-dephospho-CoA derivative 15, and HBCH2CoA 1 are at 48, 33, 39, and 32 min, respectively. Their identities have been confirmed by mass spectroscopy (MS). Compound 1 was obtained in a 50% yield from 13 after isolation by semipreparative HPLC. Its identity was confirmed by NMR and HRMS (see Supporting Information).
Inhibition Study of HBCH2CoA with Class I and III Synthases
HBCH2CoA was evaluated as an inhibitor to wt-PhaCRe and wt-PhaECAv. The kinetics of recombinant PhaCRe and PhaECAv, monitoring CoA release via a discontinuous 5,5′-dithiobis(2-nitrobenzoic acid) (DTNB) assay, are distinct from each other and are multiphasic. PhaCRe exhibits a lag phase followed by a linear phase,38 and PhaECAv exhibits a fast linear phase followed by a slower phase.8,39 In the former case the lag phase was ignored, and in the latter case only the fast phase was examined. The data (Supplementary Figures S4 and S5) were fit to the Michaelis–Menten equation, and the results in both cases showed competitive inhibition as summarized in Table 1.
Table 1. Summary of Kinetic Constants.
Lag phase ignored.
From ref (39).
Although the KM of HBCoA with PhaCRe has been reported to be 103–381 μM,40−42 the assays were carried out in high concentrations of fructose or detergents. Therefore, the kinetic parameters of PhaCRe were determined in the presence of 0.05% Hecameg (Supplementary Figure S6), providing a kcat similar to the previously reported value (188 s–1),41 but a KM that is 10-fold higher. Neither synthase was found to have any time-dependent component to the observed inhibition that might, for example, be associated with hemithioketal formation between the active site cysteine of PhaC and the carbonyl group in HBCH2CoA (data not shown). The tight binding of 1 relative to the KM for HBCoA suggests that it might be a reasonable probe of the elongation process.
Predicted Products from Reactions between Acylated Synthase and HBCH2CoA
Differentiation of two mechanistic models in Scheme 1 has been difficult as with both class I and III synthases the rate of chain elongation is much faster than that of chain initiation.8,43 Thus, even at very low substrate to enzyme ratios, a small amount of protein is modified with high molecular weight PHB polymer, while most of the protein remains unmodified.43 In order to overcome this problem, we developed a method to uniformly load PhaC, so that the elongation process could be examined.38 Specifically, an artificial primer, [3H]-saturated trimeric-CoA ([3H]-sTCoA, 16 in Scheme 3), is used to acylate the enzyme, which can then be incubated with 1. Our hypothesis is illustrated in Scheme 3. If the model in Scheme 1A operates, 1 cannot acylate the active site cysteine. However, if the model in Scheme 1B is operative, HBCH2CoA would extend the sT chain to generate a methylene analogue of [3H]-saturated tetrameric-CoA ([3H]-sTet-CH2-CoA, 17, Scheme 3), and further elongation would not be possible. Moreover, it was expected that the rate of elongation would be fast, given the Kis for 1 and rate of elongation with HBCoA.
Scheme 3. Differentiation of Two Models Using HBCH2CoA 1.
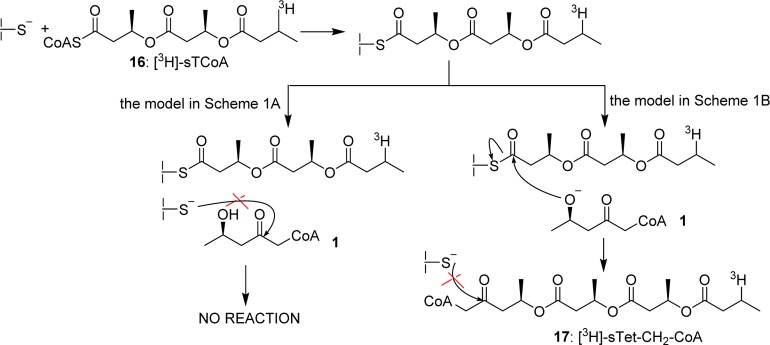
Products from Reactions between Acylated Synthase and HBCH2CoA
Although both the PhaCRe and PhaECAv can be labeled with [3H]-sTCoA with similar stoichiometry (0.5 labels monomer–1),19 the stability of the covalent linkage in PhaCRe is much higher. Thus, PhaCRe was acylated with [3H]-sTCoA, purified by Sephadex G-50 chromatography, and then reacted with HBCH2CoA. Aliquots of the reaction mixture were removed as a function of time, the reaction was stopped in 10% trichloroacetic acid, and the protein was separated from the small molecules. The supernatant was neutralized and analyzed by reverse phase HPLC by A260 nm, and the fractions were monitored by scintillation counting. A typical elution profile in which radioactivity and A260 nm are monitored is shown in Figure 1. In the experiment fractions 19–22 (sample A), 26–30 (sample B), and 31–34 min (sample C) were pooled based on radioactivity, concentrated, and analyzed by either ESI-MS (samples A and C) or MALDI-TOF MS (sample B). The results are shown in Figure 2 and Supplementary Figures S7 and S8. The material in samples A and C correspond to [3H]-sD-CO2H 18 and [3H]-sT-CO2H 19, respectively (Scheme 4). No absorption at 260 nm is apparent in either sample (Figure 1). Sample B, on the other hand, has absorbance at 260 nm associated with CoA that accounts for 90% of the sample. The MALDI-TOF MS of this sample is consistent with [3H]-sT-CH2-CoA 20 (Figure 2). Finally, the retention times of species 18, 19, and 20 are identical to the standards prepared synthetically. The kinetics of formation of these species is shown in Supplementary Figure S9, and the rate constants from these data are summarized in Table 2. A control experiment in the absence of HBCH2CoA was also carried out, and the results are shown in Figure 1 as a black solid line. The rate constants are again summarized in Table 2. The rate of hydrolysis to form [3H]-sT-CO2H 19 is 5-fold faster than that of [3H]-sD-CO2H 18. When HBCH2CoA is present, hydrolysis is suppressed. However, the rate of formation of [3H]-sT-CH2-CoA 20 is slow and only 3-fold faster than the total rate of formation of [3H]-sD-CO2H 18 and [3H]-sT-CO2H 19.
Figure 1.
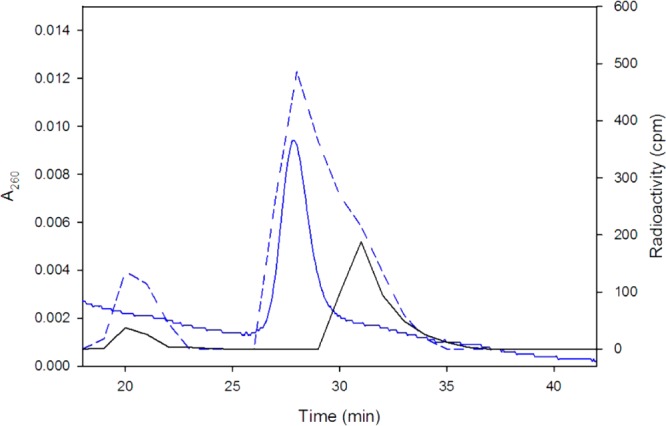
Typical HPLC and radioactivity profiles of the reaction between [3H]-sT-PhaCRe (20.0 μM) and HBCH2CoA and a control reaction. The experiment in the presence of HBCH2CoA at 15.0 mM is shown in blue solid and dashed lines for A260 nm (left Y-axis) and radioactivity (right Y-axis), respectively. The control in the absence of HBCH2CoA is shown as a black solid line for radioactivity.
Figure 2.

Negative mode of MALDI-TOF MS spectrum of sample B that is assigned as [3H]-sT-CH2-CoA 20 [(M – H)−: calcd 990.2669, found 989.9526].
Scheme 4. Model of Product Formation.
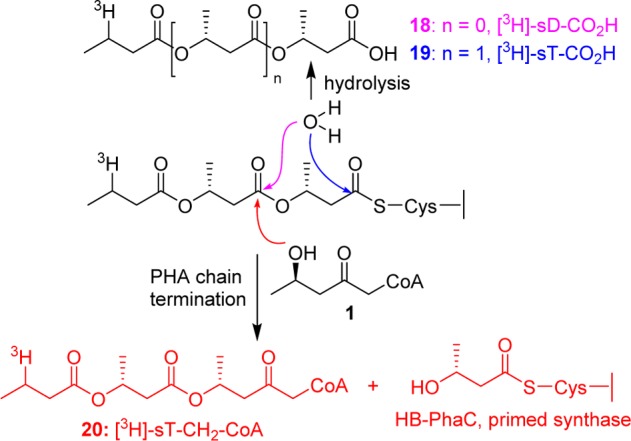
Table 2. Rate Constants of Product Formation.
| product |
|||
|---|---|---|---|
| 18 | 19 | 20 | |
| hydrolysis (control) (min–1) | 0.09 | 0.44 | |
| with HBCH2CoA (min–1) | 0.025 | 0.26 | 0.78 |
Mechanistic Implications
Several unexpected results were obtained from this study. First was the identification of [3H]-sT-CH2-CoA 20 (Scheme 4) rather than [3H]-sTet-CH2-CoA 17 (Scheme 3) as the major product. Second was the very slow rate constant for its formation relative to kcat. Our interpretation of these results is that 1 is not reporting on chain elongation as we intended, and as an alternative model we suggest that it may be reporting on chain termination and PhaC repriming. We know that the size of the PHB polymers are 1–2 MDa1 and the rate constant for polymerization is 196 s–1 (this study). Thus, chain termination occurs every 1–2 × 104 turnovers, proceeding with a rate constant of 0.6–1.2 min–1. This number is very similar to the rate constant of [3H]-sT-CH2-CoA formation of 0.8 min–1. Thus, we propose that 1 binds and dissociates from the active site of PhaC many times before ester formation occurs. Detection of [3H]-sT-CH2-CoA suggests that the chain termination occurs by reaction at the penultimate HB unit (Scheme 4) leaving an acylated-HB unit. We suggest that this HB-PhaC (Scheme 4) may function as a primer of the polymerization process. Although a number of studies have reported on chain termination events,44−48 the actual chain terminator and termination site in the biological systems remain unknown. The proposed formation of HB-PhaC is in accord with our previous study of mutant class III D302A-PhaECAv, in which proteins modified with (HB)n (n = 3–10) were identified.48 Furthermore, a recent study on class I Aeromonas caviae PhaC has suggested that HBCoA causes conversion of the monomeric form of the synthase into its active, dimeric form.49 We have also shown previously that acylation of PhaCRe with a (HB)nCoA (n = 2 or 3) decreases the lag phase of the reaction, increases the amount of dimeric PhaC, and increases the activity of the enzyme.38 The similarity in structure between HBCH2CoA and HBCoA suggests that HBCoA itself could function as a chain terminator in vivo. The observation of [3H]-sD-CO2H 18 and [3H]-sT-CO2H 19 (Scheme 4) is also interesting as water may function as a chain terminator as well. The observed ratio of [3H]-sT-CO2H/[3H]-sD-CO2H of 5 (calculated from the rate constants), rather than 100 based on the differences in chemical reactivity of an oxo vs thioester,50 suggests again that there is something special about the penultimate HB unit. Thus, these results may have unexpectedly given us new insight about the termination and repriming processes and are in line with other published results described above. Additional studies with this analogue are in progress and will be reported in a due course.
Conclusion
A methylene analogue, HBCH2CoA 1, was prepared through a chemoenzymatic approach. Inhibition studies with class I and III synthases reveals that 1 is a competitive inhibitor of HBCoA with tight binding relative to KM for HBCoA. The unexpected product ([3H]-sT-CH2-CoA) identified when HBCH2CoA was incubated with the [3H]-sT-acylated PhaC and its slow rate of formation suggest that HBCH2CoA may be reporting on the chain termination process rather than elongation based on our original design. The detection of [3H]-sT-CH2-CoA and [3H]-sD-CO2H not only highlight the importance of penultimate HB unit in PHB chain but may also shed light on the possible termination site and chain terminator in vivo. Further investigations are needed in order to better understand the challenging reactions catalyzed by PHA/PHB synthases.
Methods
Materials and Enzymes
All commercial chemicals were purchased at the highest purity grade. All solvents were anhydrous. All reactions were performed under argon atmosphere unless otherwise specified. Radiolabeled [3H]-sTCoA was synthesized according to our previously developed method.19 Thin layer chromatography (TLC) was performed using 60 mesh silica gel plates and visualization was performed using short wavelength UV light (254 nm) and/or basic KMnO4 staining.
His-tagged pantothenate kinase from Staphylococcus aureus (SaCoaA) and phosphopantetheine adenylyltransferase (EcCoaD) and dephospho-CoA kinase (EcCoaE) from E. coli were purified according to the published methods.29,34 The activity of each enzyme was measured following the reported procedure. The specific activities (SA) were measured at 45, 27, and 20 μmol min–1 mg–1 at 25 °C for SaCoaA, EcCoaD, and EcCoaE, respectively. Synthases of wt-PhaCRe and wt-PhaECAv were purified following the published procedures.10,39 The SA were measured at 50 (25 °C) and 120 μmol min–1mg–1 (37 °C) for PhaCRe and PhaECAv, respectively.
Enzymatic Synthesis of HBCH2CoA 1
A 2 mL reaction mixture consisted of 13 (20.0 mM), ATP (50.0 mM), MgCl2 (10.0 mM), SaCoaA (80.0 μg), EcCoaD (80.0 μg), and EcCoaE (80.0 μg) in 100 mM Tris-HCl (pH 7.60). The reaction was initiated by addition of the enzymes and incubated at 37 °C for 3 h. The reaction was stopped by heating the reaction mixture in a 95 °C water bath for 5 min, and the precipitated protein was removed by centrifugation (14,000 rpm × 5 min). The supernatant was loaded onto a semipreparative HPLC column (Luna C18-2, 5 μm, 10 mm × 250 mm) that was eluted at 3.00 mL min–1 using a linear gradient from 0 to 30% methanol in 10.0 mM ammonium acetate (pH 5) over 60 min. The fractions containing the product were pooled, concentrated in vacuo, and lyophilized to give a white powder (15.0 mg, 50% yield, HPLC: tR = 33 min). The final product was desalted by HPLC using methanol and water as the eluents.
Inhibition Studies
The reaction was carried out at 25 and 30 °C for wt-PhaCRe and wt-PhaECAv, respectively, in a final volume of 300 μL consisting of buffer (for wt-PhaCRe: 150 mM KPi, pH 7.20 and 0.05% Hecameg; for wt-PhaECAv: 20 mM Tris-HCl, pH 7.80 and 50 mM NaCl), wt synthase (140 and 25 nM for PhaCRe and PhaECAv, respectively), HBCoA, and HBCH2CoA. The reaction was initiated by the addition of enzyme. At defined time points, 20 μL aliquots were removed from the reaction mixture and quenched with 20 μL of 10% TCA. Each sample was centrifuged to remove the precipitated protein, then 38 μL of the quenched reaction was added to 382 μL of 0.25 mM DTNB in 500 mM KPi (pH 7.80), and A412 was measured. The rates of the reaction were determined by the slope of the initial fast phase in the case of PhaECAv and ignoring lag phase in the case of PhaCRe. Each point was done in triplicate. The data were fit to Michaelis–Menten equation for the best inhibition mode (competitive, uncompetitive, and noncompetitive) using SigmaPlot.
Reaction of *sT-PhaCRe and HBCH2CoA
In a final volume of 50 μL, 500 μM wt-PhaCRe and 5.00 mM *sT-CoA (3120 cpm nmol–1) in buffer A consisting of 150 mM KPi (pH 7.20) and 0.05% Hecameg were incubated at 25 °C for 1 min before loading the mixture onto a Sephadex G-50 column (5 mm × 150 mm) pre-equilibrated with buffer A at 4 °C. The acylated protein [3H]-sT-PhaCRe was isolated in 97% yield and labeled as 0.5 equiv [3H] per monomer. The protein was then divided equally into two portions. One portion was reacted with HBCH2CoA. The reaction was carried out at 25 °C in a final volume of 500 μL containing 20 μM [3H]-sT-PhaCRe and 15.0 mM HBCH2CoA. At defined time points, 100 μL aliquots were removed from the reaction mixture. The reaction was then stopped by addition of 50 μL of 10% TCA followed by centrifugation at 14,000 rpm for 15 min at 4 °C. The supernatant was removed, and the protein precipitate was washed three times with 100 μL of ddH2O. The supernatant and washes were combined, and the pH was adjusted to 7 using 1 M NaOH. The mixture was then analyzed by HPLC and scintillation counting. HPLC was performed using a reverse-phase C18 column (10 μm, 250 mm × 4.6 mm, Alltech) eluting with 50 mM KPi (pH 6.72) (A) and methanol (B) and at a flow rate of 1 mL min–1. The gradient (B) was from 5% to 40% from 0 to 20 min, 40% to 70% from 20 to 55 min, 70% to 95% from 55 to 75 min, and to 95% from 75 to 85 min; 1 min fractions were collected and analyzed by scintillation counting. The protein precipitate from the each reaction was dissolved in 10% SDS and analyzed by scintillation counting. The inventory of radioactivity is summarized in Supplementary Table S3. A control experiment (hydrolysis) was performed in parallel with the other portion of [3H]-sT-PhaCRe, in which HBCH2CoA was replaced with buffer A. The inventory of radioactivity is summarized in Supplementary Table S4.
The rates of product formation were obtained by fitting the kinetic data to the equation y = a × (1 – e–bx) using SigmaPlot.
Sample Preparation for MS Analysis
For the above reaction, fractions eluted at 19–22 (sample A), 26–30 (sample B), and 31–34 min (sample C) were combined and concentrated to dryness using a speedvac. Each sample was then dissolved in 80 μL of ddH2O and subjected to A260 nm measurement. Quantitation by A260 nm and radioactivity showed that sample B contained more than 90% CoA analogue. Samples A/C and B were analyzed by ESI-MS and MALDI-TOF MS, respectively.
Acknowledgments
This work was supported by funds from Kansas State University and Johnson Cancer Research Center to P.L. and National Institute of Health grant to J.S. (GM49171). J.J. was supported by the ASSURE program of the Department of Defense in partnership with the National Science Foundation REU Site program under grant number 1004991.
Supporting Information Available
Experimental details and spectral data. This material is available free of charge via the Internet at http://pubs.acs.org.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
Supplementary Material
References
- Stubbe J.; Tian J. (2003) Polyhydroxyalkanoate (PHA) homeostasis: the role of PHA synthase. Nat. Prod. Rep. 20, 445–457. [DOI] [PubMed] [Google Scholar]
- Madison L. L.; Huisman G. W. (1999) Metabolic engineering of poly(3-hydroxyalkanoates): From DNA to plastic. Microbiol. Mol. Biol. Rev. 63, 21–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doi Y. (1990) Microbial Polyesters, VCH, New York, NY. [Google Scholar]
- Steinbüchel A. (1991) in Biomaterials: Novel Materials from Biological Sources (Byrom D., Ed.) p 123, Macmillan Publishers Ltd., Basingstoke, U.K.. [Google Scholar]
- Steinbuchel A.; Hein S. (2001) Biochemical and molecular basis of microbial synthesis of polyhydroxyalkanoates in microorganisms. Adv. Biochem. Eng. Biotechnol. 71, 81–123. [DOI] [PubMed] [Google Scholar]
- Sudesh K.; Iwata T. (2008) Sustainability of biobased and biodegradable plastics. Clean-Soil Air Water 36, 433–442. [Google Scholar]
- Rehm B. H. A. and Steinbüchel A. (2001) Polyhydroxyalkanoate (PHA) Synthases: The Key Enzymes of PHA Synthesis. In Biopolymers (Steinbüchel A. and Doi Y., Ed.), pp 173, Wiley-VCH, Heidelberg, Germany. [Google Scholar]
- Gerngross T. U.; Snell K. D.; Peoples O. P.; Sinskey A. J.; Csuhai E.; Masamune S.; Stubbe J. (1994) Overexpression and purification of the soluble polyhydroxyalkanoate synthase from Alcaligenes eutrophus: evidence for a required posttranslational modification for catalytic activity. Biochemistry 33, 9311–9320. [DOI] [PubMed] [Google Scholar]
- Liebergesell M.; Steinbuchel A. (1992) Cloning and nucleotide sequences of genes relevant for biosynthesis of poly(3-hydroxybutyric acid) in Chromatium vinosum strain D. Eur. J. Biochem. 209, 135–150. [DOI] [PubMed] [Google Scholar]
- Muh U.; Sinskey A. J.; Kirby D. P.; Lane W. S.; Stubbe J. (1999) PHA synthase from chromatium vinosum: cysteine 149 is involved in covalent catalysis. Biochemistry 38, 826–837. [DOI] [PubMed] [Google Scholar]
- Li P.; Chakraborty S.; Stubbe J. (2009) Detection of covalent and nNoncovalent intermediates in the polymerization reaction catalyzed by a C149S class III polyhydroxybutyrate synthase. Biochemistry 48, 9202–9211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stubbe J.; Tian J. M.; He A. M.; Sinskey A. J.; Lawrence A. G.; Liu P. H. (2005) Nontemplate-dependent polymerization processes: Polyhydroxyalkanoate synthadses as a paradigms. Annu. Rev. Biochem. 74, 433–480. [DOI] [PubMed] [Google Scholar]
- Smith S. (1994) The animal fatty acid synthase: one gene, one polypeptide, seven enzymes. FASEB J. 8, 1248–1259. [PubMed] [Google Scholar]
- Griebel R.; Smith Z.; Merrick J. M. (1968) Metabolism of poly-beta-hydroxybutyrate. I. Purification composition and properties of native poly-beta-hydroxybutyrate granules from Bacillus megaterium. Biochemistry 7, 3676–3681. [DOI] [PubMed] [Google Scholar]
- Griebel R. J.; Merrick J. M. (1971) Metabolism of poly-beta-hydroxybutyrate - Effect of mild alkaline extraction on native poly-beta-hydroxybutyrate granules. J. Bacteriol. 108, 782–789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witkowski A.; Joshi A. K.; Smith S. (1997) Characterization of the interthiol acyltransferase reaction catalyzed by the beta-ketoacyl synthase domain of the animal fatty acid synthase. Biochemistry 36, 16338–16344. [DOI] [PubMed] [Google Scholar]
- Jia Y.; Kappock T. J.; Frick T.; Sinskey A. J.; Stubbe J. (2000) Lipases provide a new mechanistic model for polyhydroxybutyrate (PHB) synthases: Characterization of the functional residues in Chromatium vinosum PHB synthase. Biochemistry 39, 3927–3936. [DOI] [PubMed] [Google Scholar]
- Austin M. B.; Noel J. P. (2003) The chalcone synthase superfamily of type III polyketide synthases. Nat. Prod. Rep. 20, 79–110. [DOI] [PubMed] [Google Scholar]
- Jia Y.; Yuan W.; Wodzinska J.; Park C.; Sinskey A. J.; Stubbe J. (2001) Mechanistic studies on class I polyhydroxybutyrate (PHB) synthase from Ralstonia eutropha: class I and III synthases share a similar catalytic mechanism. Biochemistry 40, 1011–1019. [DOI] [PubMed] [Google Scholar]
- Dinh T. Q.; Armstrong R. W. (1996) Synthesis of ketones and aldehydes via reactions of Weinreb-type amides on solid support. Tetrahedron Lett. 37, 1161–1164. [Google Scholar]
- Cahiez G.; Alexakis A.; Normant J. F. (1978) Organomagnesium derivatives of terminal Alcohols. Tetrahedron Lett. 3013–3014. [Google Scholar]
- Chen B.; Ko R. Y. Y.; Yuen M. S. M.; Cheng K. F.; Chiu P. (2003) A tandem metal carbene cyclization-cycloaddition approach to the pseudolaric acids. J. Org. Chem. 68, 4195–4205. [DOI] [PubMed] [Google Scholar]
- Stanton S. A.; Felman S. W.; Parkhurst C. S.; Godleski S. A. (1983) Alkoxides as nucleophiles in (pi-allyl)Palladium chemistry - synthetic and mechanistic studies. J. Am. Chem. Soc. 105, 1964–1969. [Google Scholar]
- Li Y.; Zhang Q.; Wittlin S.; Jin H. X.; Wu Y. K. (2009) Synthesis and in vitro antimalarial activity of spiro-analogues of peroxyplakoric acids. Tetrahedron 65, 6972–6985. [Google Scholar]
- Gololobov Y. G.; Zhmurova I. N.; Kasukhin L. F. (1981) 60 years of Staudinger reaction. Tetrahedron 37, 437–472. [Google Scholar]
- Staudinger H.; Meyer J. (1919) Über neue organische phosphorverbindungen III. phosphinmethylenderivate and phosphinimine. Helv. Chim. Acta 2, 635–646. [Google Scholar]
- van Kalkeren H. A.; Bruins J. J.; Rutjes F.; van Delft F. L. (2012) Organophosphorus-catalysed staudinger reduction. Adv. Synth. Catal. 354, 1417–1421. [Google Scholar]
- Chhen A.; Vaultier M.; Carrie R. (1989) Stereospecific synthesis of Z-olefins bearing an omega-azido group. Tetrahedron Lett. 30, 4953–4956. [Google Scholar]
- Leonardi R.; Chohnan S.; Zhang Y. M.; Virga K. G.; Lee R. E.; Rock C. O.; Jackowski S. (2005) A pantothenate kinase from Staphylococcus aureus refractory to feedback regulation by coenzyme A. J. Biol. Chem. 280, 3314–3322. [DOI] [PubMed] [Google Scholar]
- Geerlof A.; Lewendon A.; Shaw W. V. (1999) Purification and characterization of phosphopantetheine adenylyltransferase from Escherichia coli. J. Biol. Chem. 274, 27105–27111. [DOI] [PubMed] [Google Scholar]
- Mishra P. K.; Park P. K.; Drueckhammer D. G. (2001) Identification of yacE (coaE) as the structural gene for dephosphocoenzyme a kinase in Escherichia coli K-12. J. Bacteriol. 183, 2774–2778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke K. M.; Mercer A. C.; La Clair J. J.; Burkart M. D. (2005) In vivo reporter labeling of proteins via metabolic delivery of coenzyme A analogues. J. Am. Chem. Soc. 127, 11234–11235. [DOI] [PubMed] [Google Scholar]
- Dai M.; Feng Y. G.; Tonge P. J. (2001) Synthesis of crotonyl-OxyCoA: A mechanistic probe of the reaction catalyzed by enoyl-CoA hydratase. J. Am. Chem. Soc. 123, 506–507. [DOI] [PubMed] [Google Scholar]
- Strauss E.; Begley T. P. (2002) The antibiotic activity of N-pentylpantothenamide results from its conversion to ethyldethia-coenzyme A, a coenzyme A antimetabolite. J. Biol. Chem. 277, 48205–48209. [DOI] [PubMed] [Google Scholar]
- Tosin M.; Spiteller D.; Spencer J. B. (2009) Malonyl carba(dethia)- and malonyl oxa(dethia)-coenzyme A as tools for trapping polyketide intermediates. ChemBioChem. 10, 1714–1723. [DOI] [PubMed] [Google Scholar]
- van der Westhuyzen R.; Strauss E. (2010) Michael acceptor-containing coenzyme A analogues as inhibitors of the atypical coenzyme a disulfide reductase from Staphylococcus aureus. J. Am. Chem. Soc. 132, 12853–12855. [DOI] [PubMed] [Google Scholar]
- Rock C. O.; Park H. W.; Jackowski S. (2003) Role of feedback regulation of pantothenate kinase (CoaA) in control of coenzyme A levels in Escherichia coli. J. Bacteriol. 185, 3410–3415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wodzinska J.; Snell K. D.; Rhomberg A.; Sinskey A. J.; Biemann K.; Stubbe J. (1996) Polyhydroxybutyrate synthase: Evidence for covalent catalysis. J. Am. Chem. Soc. 118, 6319–6320. [Google Scholar]
- Yuan W.; Jia Y.; Tian J. M.; Snell K. D.; Muh U.; Sinskey A. J.; Lambalot R. H.; Walsh C. T.; Stubbe J. (2001) Class I and III polyhydroxyalkanoate synthases from Ralstonia eutropha and Allochromatium vinosum: Characterization and substrate specificity studies. Arch. Biochem. Biophys. 394, 87–98. [DOI] [PubMed] [Google Scholar]
- Normi Y. M.; Hiraishi T.; Taguchi S.; Abe H.; Sudesh K.; Najimudin N.; Doi Y. (2005) Characterization and properties of G4X mutants of Ralstonia eutropha PHA synthase for poly (3-hydroxybutyrate) biosynthesis in Escherichia coli. Macromol. Biosci. 5, 197–206. [DOI] [PubMed] [Google Scholar]
- Ushimaru K.; Sangiambut S.; Thomson N.; Sivaniah E.; Tsuge T. (2013) New insights into activation and substrate recognition of polyhydroxyalkanoate synthase from Ralstonia eutropha. Appl. Microbiol. Biotechnol. 97, 1175–1182. [DOI] [PubMed] [Google Scholar]
- Zhang S. M.; Yasuo T.; Lenz R. W.; Goodwin S. (2000) Kinetic and mechanistic characterization of the polyhydroxybutyrate synthase from Ralstonia eutropha. Biomacromolecules 1, 244–251. [DOI] [PubMed] [Google Scholar]
- Tian J. M.; Sinskey A. J.; Stubbe J. (2005) Class III polyhydroxybutyrate synthase: Involvement in chain termination and reinitiation. Biochemistry 44, 8369–8377. [DOI] [PubMed] [Google Scholar]
- Lawrence A. G.; Choi J.; Rha C.; Stubbe J.; Sinskey A. J. (2005) In vitro analysis of the chain termination reaction in the synthesis of poly-(R)-beta-hydroxybutyrate by the class III synthase from Allochromatium vinosum. Biomacromolecules 6, 2113–2119. [DOI] [PubMed] [Google Scholar]
- Madden L. A.; Anderson A. J.; Shah D. T.; Asrar J. (1999) Chain termination in polyhydroxyalkanoate synthesis: involvement of exogenous hydroxy-compounds as chain transfer agents. Int. J. Biol. Macromol. 25, 43–53. [DOI] [PubMed] [Google Scholar]
- Shi F. Y.; Ashby R.; Gross R. A. (1996) Use of poly(ethylene glycol)s to regulate poly(3-hydroxybutyrate) molecular weight during Alcaligenes eutrophus cultivations. Macromolecules 29, 7753–7758. [Google Scholar]
- Shi F. Y.; Gross R. A.; Rutherford D. R. (1996) Microbial polyester synthesis: Effects of poly(ethylene glycol) on product composition, repeat unit sequence, and end group structure. Macromolecules 29, 10–17. [Google Scholar]
- Tian J.; Sinskey A. J.; Stubbe J. (2005) Detection of intermediates from the polymerization reaction catalyzed by a D302A mutant of class III polyhydroxyalkanoate (PHA) synthase. Biochemistry 44, 1495–1503. [DOI] [PubMed] [Google Scholar]
- Numata K.; Motoda Y.; Watanabe S.; Tochio N.; Kigawa T.; Doi Y. (2012) Active intermediates of polyhydroxyalkanoate synthase from Aeromonas caviae in polymerization reaction. Biomacromolecules 13, 3450–3455. [DOI] [PubMed] [Google Scholar]
- Jencks W. P.; Cordes S.; Carriuolo J. (1960) The free energy of thiol ester hydrolysis. J. Biol. Chem. 235, 3608–3614. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.