

Abstract
Understanding patterns of larval dispersal is key in determining whether no-take marine reserves are self-sustaining, what will be protected inside reserves and where the benefits of reserves will be observed. We followed a multidisciplinary approach that merged detailed descriptions of fishing zones and spawning time at 17 sites distributed in the Midriff Island region of the Gulf of California with a biophysical oceanographic model that simulated larval transport at Pelagic Larval Duration (PLD) 14, 21 and 28 days for the most common and targeted predatory reef fish, (leopard grouper Mycteroperca rosacea). We tested the hypothesis that source–sink larval metapopulation dynamics describing the direction and frequency of larval dispersal according to an oceanographic model can help to explain empirical genetic data. We described modeled metapopulation dynamics using graph theory and employed empirical sequence data from a subset of 11 sites at two mitochondrial genes to verify the model predictions based on patterns of genetic diversity within sites and genetic structure between sites. We employed a population graph describing a network of genetic relationships among sites and contrasted it against modeled networks. While our results failed to explain genetic diversity within sites, they confirmed that ocean models summarized via graph and adjacency distances over modeled networks can explain seemingly chaotic patterns of genetic structure between sites. Empirical and modeled networks showed significant similarities in the clustering coefficients of each site and adjacency matrices between sites. Most of the connectivity patterns observed towards downstream sites (Sonora coast) were strictly asymmetric, while those between upstream sites (Baja and the Midriffs) were symmetric. The best-supported gene flow model and analyses of modularity of the modeled networks confirmed a pulse of larvae from the Baja Peninsula, across the Midriff Island region and towards the Sonoran coastline that acts like a larval sink, in agreement with the cyclonic gyre (anti-clockwise) present at the peak of spawning (May–June). Our approach provided a mechanistic explanation of the location of fishing zones: most of the largest areas where fishing takes place seem to be sustained simultaneously by high levels of local retention, contribution of larvae from upstream sites and oceanographic patterns that concentrate larval density from all over the region. The general asymmetry in marine connectivity observed highlights that benefits from reserves are biased towards particular directions, that no-take areas need to be located upstream of targeted fishing zones, and that some fishing localities might not directly benefit from avoiding fishing within reserves located adjacent to their communities. We discuss the implications of marine connectivity for the current network of marine protected areas and no-take zones, and identify ways of improving it.
Keywords: Biophysical models, Population genetics, Oceanography, Gulf of California, Fisheries, No-take zones, Marine reserves, Larval dispersal, Marine connectivity
Introduction
Knowledge of patterns of larval dispersal is essential to implement fully-protected marine reserves (no-take zones), a tool frequently used to enhance the conservation of biodiversity and the recovery of fisheries (Gaines et al., 2010). Reserves must either be large enough to be self-sufficient via local retention (larvae retained or returning to the reserve where they were produced) (Toonen et al., 2013), or need to be linked by a network of reserves for persistence (larval supply from reserves to other reserves/fished sites) (Hastings & Botsford, 2006; White et al., 2010a). However, the efficacy of networks of reserves has been hindered by a lack of knowledge regarding complex patterns of marine connectivity (Sale et al., 2005; Burgess et al., 2014). A multidisciplinary approach could best address the intricacy of connectivity by merging biophysical models of ocean currents that generate connectivity hypotheses using detailed biological information on the spatial and temporal distribution of propagules (larvae), followed by validation with empirical population genetics data (Gilg & Hilbish, 2003; Baums, Paris & Cherubin, 2006; White et al., 2010b; Alberto et al., 2011; Crandall, Treml & Barber, 2012; Foster et al., 2012; Soria et al., 2012; Feutry et al., 2013). Use of multiple methodologies could also help advance an increasing interest in incorporating genetic information into marine spatial planning, for example, by identifying sites with high genetic diversity that hold evolutionary potential under future environmental change (Beger et al., 2014).
Marine connectivity within a single species is influenced by multiple biological and physical factors including: spawning time and location, pelagic larval duration (PLD), their interaction with ocean current speed and direction, as well as the distribution of suitable habitat for settlement (reviewed by Cowen & Sponaugle, 2009). Additionally, many commercially exploited species of invertebrates and fishes display metapopulations that are connected via larval dispersal (Cowen et al., 2000). Few attempts have been done to establish multidisciplinary approaches to understand marine connectivity (e.g., Soria et al., 2012; Soria et al., 2014). A challenge is to find a key species that can be a relevant case study, for which this information can be gathered relatively easily, and can be used as an umbrella species to design marine reserves, although this approach might not be always effective (Bird et al., 2007; Toonen et al., 2011).
The Midriff Islands region includes 45 islands and islets located on the division between the northern and central Gulf of California (GC). The bathymetry of this region constitutes one of the most notable features of the GC, which is formed by a series of deep basins (e.g., Dolphin, Salsipuedes, San Pedro Martir) and sills, and which restrict the circulation between the northern and central GC (Hernandez-Ayon et al., 2013). The combination of this restricted circulation, with the weakening of stratification by the turbulent kinetic energy released during the passage of the semidiurnal tidal wave through the constrictions of this archipelago (Argote et al., 1995) results in an oceanographically complex body of water (Lavin & Marinone, 2003) and the presence of a surface thermal front (Paden & Abbott, 1991). These features have been documented to restrict larval distributions, generating differences between northern and central fish larval assemblages (Danell-Jiménez et al., 2009). This study was centered in the Northern GC, which is characterized by a large-scale seasonally reversing gyre that has been documented by different approaches, including satellite tracked drifters (Lavin et al., 1997), geostrophic calculations (Carrillo & Palacios-Hernández, 2002), current meters (Palacios-Hernandez et al., 2002) and numerical models (Marinone, 2003; Marinone, 2012). The basin-wide gyre is cyclonic (anti-clockwise) from May to September (Lavin et al., 1997), and reverses to an anti-cyclonic (clockwise) gyre from October to March. The forcing systems have been related to the annual-period monsoonal winds, the Pacific Ocean and the superficial heat flux (Beier & Ripa, 1999).
Small fisheries worldwide comprise most of the global catch, yet most lack formal assessments and are continuing to decline (Costello et al., 2012). The leopard grouper (Mycteroperca rosacea, Streets 1877) is the most common and heavily targeted grouper by small-scale and recreational fisheries in the GC (Sala et al., 2003; Craig, Sadovy de Mitcheson & Heemstra, 2011). Due to increased fishing pressure and observed declines in fisheries landings, sizes of harvested fish, and population abundances in some areas of the GC over the past few decades (Sala et al., 2004), the International Union for the Conservation of Nature (IUCN) currently lists M. rosacea as ‘Vulnerable’ (Craig & Sadovy, 2008). While commercial fishers are required to hold a finfish permit and record their landings of leopard grouper to their local fisheries offices, no specific regulations related to catch, size, or gear restrictions exist for this species. Currently, all M. rosacea catches are aggregated in a large group, registered as “finfish” with other 270 fish species according to the National Fisheries Chart (Anonymous, 2012). Marine Protected Areas (MPAs) represent the primary, current conservation and management strategy that has been implemented for this or any other reef fish in the GC (Cudney-Bueno et al., 2009; Aburto-Oropeza et al., 2011), and are mainly concentrated in the western coast of the GC and contain a few small no-take zones (Fig. 1).
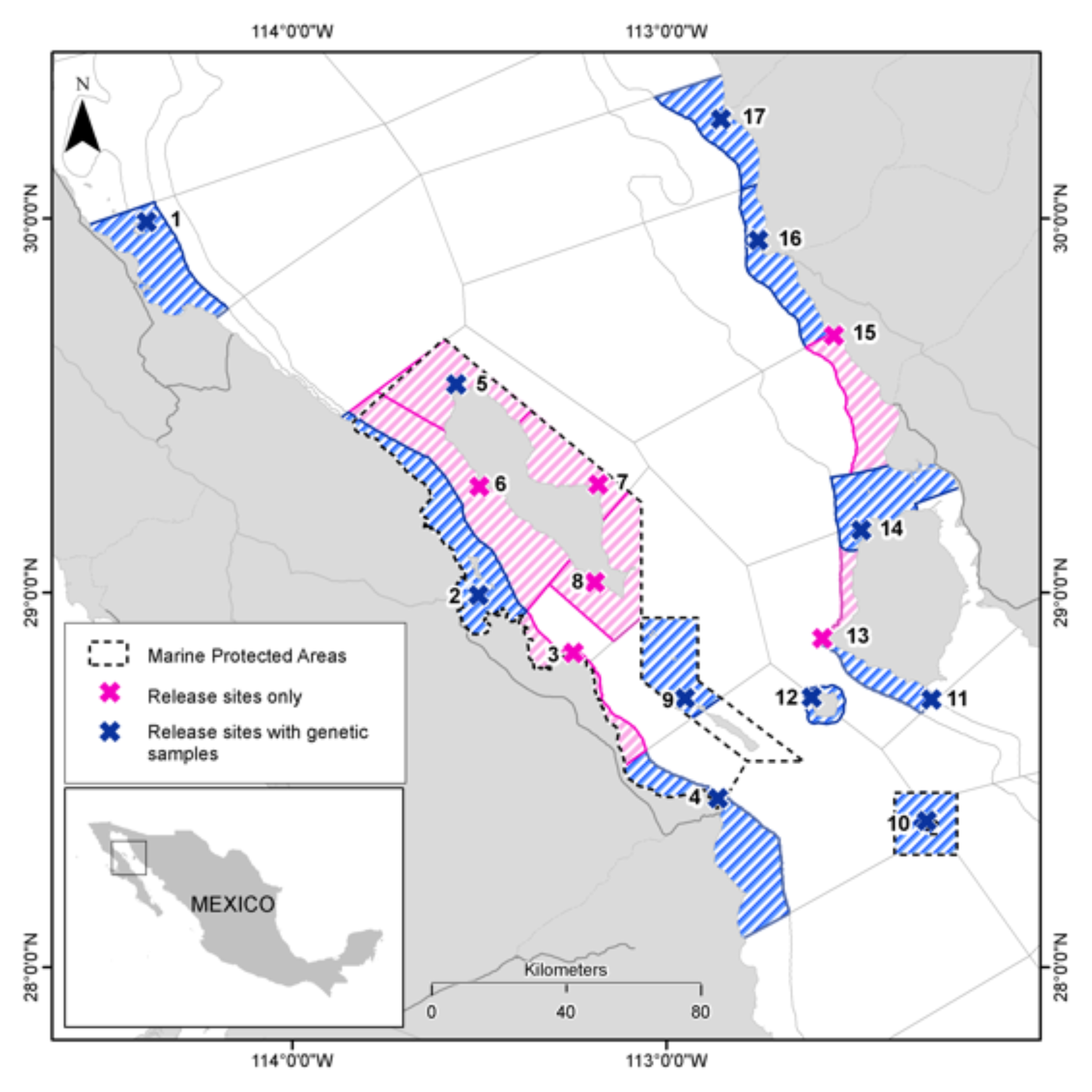
Figure 1. Area of study around the Midriff Islands in the Gulf of California.

Study sites, in which modeled and genetic connectivity was measured (release sites with genetic samples) and those only for modeled connectivity (release sites only), including marine protected areas, no-take zones and fishing zones identified by interviews with fishers (Moreno-Baez et al., 2010; Moreno-Báez et al., 2012). Note that no-take zones are small and dotted circles denote approximate location but not size.
Our main hypothesis is that source–sink larval metapopulation dynamics in the Midriff Island region, including the direction and frequency of larval dispersal described according to a numerical oceanographic model can help to significantly explain empirical genetic patterns and inform sustainable fisheries and the design of a network of marine reserves. We searched for agreement between models and empirical data at two levels: node-based analyses including genetic diversity within sites and matrix-based analyses including genetic structure between sites. To achieve this, we first determined the distinct spawning season for leopard grouper and identified the spatial distribution of spawning aggregation sites and fishing zones across the entire region. We modeled connectivity with a biophysical model and used graph theory to describe metapopulation dynamics. We contrasted distinct measures describing larval dispersal derived from graph theory against empirical estimates of genetic diversity and differentiation to corroborate model expectations. Our approach also aimed to identify sites that are likely self-sustaining and important sources and sinks for leopard grouper larvae, including locations that may lie inside or outside the borders of existing MPAs. Results of this study provide insights on validating biophysical models with empirical genetic data, and also on the benefits and limitations of the current network of MPAs to fishing communities in the Midriffs region that harvest M. rosacea. This work will help identify areas that may serve as ideal locations for refuges of spawning adults or juveniles of this economically important yet vulnerable species.
Methods
Spawning sites, season and period
Mycteroperca rosacea is a large predatory reef fish (Teleostei: Epinephelidae) endemic to the GC bioregion. It ranges from Bahía Magdalena in the Pacific coast of the Baja California Peninsula south to Bahía Banderas in Nayarit, Mexico, including all rocky-reefs within the interior of GC (Thomson, Findley & Kerstitch, 2000; Robertson & Cramer, 2009; Hastings, Findley & Van der heiden, 2010). Ecologically, it represents the most common and numerically abundant fish top predator on reefs in the entire GC. Individuals can reach 1 m in length and at least 22 years of age (Diaz-Uribe, Elorduy-Garay & Gonzalez-Valdovinos, 2001). Spawning occurs in the evening within groups of 6–40 individuals and is not correlated with the lunar cycle (Erisman, Buckhorn & Hastings, 2007). Histological and population data indicate gonochorism, with no evidence of post-maturational sex change found in adults caught in the wild (Erisman, Rosales-Casián & Hastings, 2007).
We used information from underwater and fisheries surveys conducted at locations throughout the Midriffs Islands region in the GC, Mexico (Fig. 1). This information helped to identify the sites to simulate the dispersal of leopard grouper eggs and larvae from spawning aggregation sites. Underwater surveys were performed at 33 sites throughout the Midriffs during the spawning season of M. rosacea (April–June) in 2008, 2009, and 2010. Evidence of the formation of spawning aggregations were based on standard protocols (Colin, Sadovy & Domeier, 2003) and those adapted for leopard grouper (Erisman, Rosales-Casián & Hastings, 2007). Direct evidence of spawning aggregations included observations of courtship or spawning behavior or the collection of females with hydrated or ovulated oocytes. Indirect evidence involved observations of putative females with enlarged abdomens indicative of imminent spawning, color patterns associated with courtship, the collection of males with ripe testes, and abundances and densities of fish that were markedly higher (e.g., 3-fold increases or greater) than observed during non-spawning months. Additional indirect evidence of spawning aggregations was acquired through interviews with commercial fishers at five fishing communities (Bahía de los Ángeles, Bahía de Kino, Desemboque Seri, Puerto Libertad and Punta Chueca) during 2005 and 2006 (Moreno-Baez et al., 2010; Moreno-Báez et al., 2012).
While the general spawning season for M. rosacea in the GC occurs from late April to June (Erisman, Buckhorn & Hastings, 2007), it was necessary to collect empirical data to narrow the specific spawning season from the Midriff Islands region. We acquired gonad samples of adult female leopard groupers (i.e., >30 cm TL; Erisman, Buckhorn & Hastings, 2007) from commercial fishers on a monthly basis from December 2008 to June 2010. Fish were captured by gill nets or handlines at various sites at or near San Pedro Martir and Tiburon islands, and Bahia Kino (Fig. 1). We processed tissue taken from the central portion from one gonad lobe for each sample using standard histological techniques (Humason, 1972) in order to determine sex and developmental stage. Classes of ovarian and testicular development were adapted from previous studies (Erisman, Rosales-Casián & Hastings, 2007), and stages of gametogenesis followed previously established definitions (Wallace & Selman, 1981).
We determined the duration of the spawning season using a combination of two methods. First, we examined the histological preparations of all gonad samples to identify the percentage of females capable of spawning or actively spawning during each month of sampling. Females categorized as ‘spawning capable’ included those with ovaries dominated by oocytes in advanced stages of vitellogenesis (e.g., primary to tertiary yolk stage), whereas those categorized as ‘actively spawning’ contained ovaries with oocytes in the migratory nucleus, hydrated, or ovulated stage or post-ovulatory follicles were present. Data were pooled by month to estimate the monthly proportion of spawning females over a calendar year. Collection dates of females classified as actively spawning based on histology were used as indicators to determine exact dates of spawning. A second estimate of the spawning season was obtained by calculating the mean monthly gonadosomatic index (GSI = 100 ∗ gonad weight/total body weight) of female M. rosacea over the study period. Changes in monthly GSI were used to assess reproductive activity, associating elevated levels with gonadal development and spawning. This information was used in the release dates for larvae in the oceanographic model (see below).
Supporting fishing knowledge
The central component to documenting the fishing grounds for the M. rosacea was captured through a series of interviews implemented across 17 fishing communities in the northern GC (Moreno-Baez et al., 2010; Moreno-Báez et al., 2012). The methodology entailed aggregating local knowledge of a representative set of individual fishers (captains) through semi-structured interviews conducted between December 2005 and July 2006 regarding what, where, when and how they fish. The interview included questions regarding the spatial and temporal distribution of fishing activities but also, their knowledge about locations of spawning aggregations and juvenile habitat. The maps were digitized, georeferenced, and integrated into a geographic information systems (GIS) using ArcGIS 9.2 (ESRI, 1999–2008).
These interviews indicated that fishing activity frequently overlapped spatially with spawning aggregation sites in three main regions: (1) the north end of Angel de la Guarda Island; (2) on the south, western and northern edge of Tiburon Island, and on sites in mainland Sonora north of Tiburon, around Las Cuevitas, Puerto Libertad and Puerto Lobos (yellow areas Fig. 1). Other important fishing zones were identified around Puerto Peñasco in northern Sonora. According to the interviews, the principal fishing season starts in November and ends in June.
Oceanographic model
Spatial units (Fig. S1) were established to evaluate spatial connectivity by combining physical and political boundaries, as well as local knowledge from fishers (Moreno-Baez et al., 2010; Moreno-Báez et al., 2012). We incorporated coastline and bathymetry developed by the National Geophysical Data Center (http://www.ngdc.noaa.gov/mgg/shorelines/shorelines.html) and the marine protected areas in Mexico (www.conanp.gob.mx), respectively. We used the spatial union function to integrate the different boundaries and define the spatial units, under ArcGIS 10.1 (ESRI) with the Spatial Analyst Extension and Model Builder tools. The size of the spatial units varied from 13 to 812 km2 (Fig. S1).
In a computer simulation exercise, four thousand particles were released at a depth of 5 m at each of 17 spawning aggregation sites (Fig. 1). Particles were tracked for 28 days, which is close to the maximum PLD for M. rosacea (Aburto-Oropeza et al., 2007). We released particles during eight dates representing the four neap and four spring tides during May and June (Fig. S2), which based on our observations covers the peak spawning period around the Midriff Islands region (see Results). Since seasonal oceanographic regimes are consistent across years in the GC (Marinone, 2003; Soria et al., 2014), the simulation year (2007) was chosen arbitrarily, and the phase of the spring-neap cycle is simply shifted from year to year.
We used the velocity field from the GC implementation of the three-dimensional baroclinic Hamburg Shelf Ocean Model (HAMSOM) (Backhaus, 1985) to calculate the particle trajectories. The model has been described in detail for the GC (Marinone, 2003; Marinone, 2008). Its domain has a mesh size of 2.5′/3 × 2.5′/3 (∼1.31 × 1.54 km) in the horizontal and 12 layers in the vertical with nominal lower levels at 10, 20, 30, 60, 100, 150, 200, 250, 350, 600, 1,000 and 4,000 m. The model equations are solved semi-implicitly with fully prognostic temperature and salinity fields, thus allowing time-dependent baroclinic motions. The model is started from rest with a 300 s time step and becomes periodically stable after three years. Results for this study were obtained from the fourth year of the model when it adequately reflects the main seasonal signals of surface temperature, heat balance, tidal elevation and tidal currents and surface circulation in the NGC (Lavin et al., 1997; Marinone, 2003). The forcing includes at the open boundary model tidal components (M2, S2, N2, K2, K1, O1, P1, Ssa, and the Sa), climatological hydrography historical data and at the sea surface climatological heat and fresh water fluxes. We used the seasonal climatology constructed from QUICKSCAT data as forcing for wind. The Lagrangian trajectories are due to the Eulerian velocity field plus a random-walk contribution related to turbulent eddy diffusion processes (Visser, 1997; Proehl et al., 2005). We obtained values of the diffusivities from the numerical model. A pseudo-advective term was introduced, since the vertical diffusivity is not constant, to prevent particles from walking away from areas of high to low diffusivities. The velocity at each particle position and the vertical eddy coefficients are calculated by bilinear interpolation of the instantaneous Eulerian velocity fields and the eddy coefficient from the numerical model, which were saved every hour. The horizontal diffusivity is taken as a constant (100 m2/s). We assumed that larvae are advected as passive particles and do not migrate vertically downward to deep depths, as in other studies (e.g., Watson et al., 2010), given that leopard grouper recruit to shallow Sargassum spp. beds of <5 m deep (Aburto-Oropeza et al., 2007).
Modeled connectivity
Hourly latitude and longitude data for each modeled particle were imported into MatLab (MATHWORKS). We estimated connectivity at different time intervals: 336 h (14 days), 504 h (21 days) and 672 h (28 days) respectively after the release dates. These PLDs were selected based on the average time of flexion in groupers that corresponds to the onset of larval behavior (14 days) (Cowen, 2002; Gracia-Lopez et al., 2005) and the maximum PLD (28 days) (Aburto-Oropeza et al., 2007). To identify the intersection between particles and the recruitment areas (spatial units), we used a selection by location function (in-polygon). We then generated connectivity matrices using the proportion of larvae that settled at each location relative to the total number of larvae released at each site. We constructed matrices averaging for the eight spawning dates within each PLD (i.e., days 14, 21, 28). The probability of local retention (i.e., diagonal in the connectivity matrix) was calculated as the proportion of particles produced locally that remained within the spatial unit at the end of the PLD (Burgess et al., 2014). The probabilities within each site were summarized with two statistics aimed at describing source–sink dynamics. Export probability was defined as the proportion of larvae produced within a site that successfully settled within any of the other 16 coastal areas at the end of the PLD. Import probability was defined as the proportion of all larvae produced among the 17 sites that settled within each site. This later metric is identical to self-recruitment as defined by Burgess et al. (2014).
Marine connectivity patterns were displayed using graph theory and a spatial network approach (Treml et al., 2008; Treml et al., 2012) with the software NODEXL (Smith et al., 2010). We calculated five statistics that describe the relationships among elements (i.e., sites or nodes) in complex networks (Newman, 2003), including: (1) graph size (the total number of directed links within a graph); (2) in-degree (number of links that enter a node); (3) out-degree (number of links that leave a node); (4) betweenness centrality, or the proportion of shortest paths between all node pairs that pass through a particular node; and (5) clustering coefficient, a measure of how close the neighbors of a node are to being a clique (complete graph). Betweenness centrality can be viewed as a measure of resilience by measuring how many paths will get longer when a node is removed (Newman, 2003). This measure highlights ‘most used’ dispersal pathways or stepping stones that act like gateways through which genes or individuals have to pass to spread to other nodes, emphasizing key sites for multigenerational connectivity (Andrello et al., 2013). The clustering coefficient varies from 1 (when every neighbor connected to the node is also connected to all the other nodes) to zero (if zero neighbors connected to the node are connected to other nodes connected to node in question).
We used the software GENETICSTUDIO (Dyer, 2009) to calculate two distinct types of symmetric matrices describing the distance separating pairs of nodes (sites) according to the topology of the network: (1) adjacency matrix, containing the actual edge weight (probability of larval dispersal, regardless of the direction) in the off-diagonal elements that attach the indexed nodes, while nodes that are not connected are denoted as zero; and (2) graph distance, or the length of the minimum topological distance (i.e., shortest geodesic path) between two nodes, calculated conditional to the entire data set of nodes and edges, and which has statistical properties including homoscedasticity and stability (Dyer, Nason & Garrick, 2010).
Genetic connectivity
We collected tissue samples from the pectoral fins of M. rosacea from 11 sites included in our modeling exercise around the Midriff Islands region (Fig. 1). Samples were acquired in fish markets or directly from fishermen at harbors between 2009 and 2012 under IACUC protocol Berng1101. We interviewed both fish vendors and fishermen to determine the approximate localities where fish were collected. Immediately after collection, samples were stored in 95% ethanol and kept at −20 ° C in the laboratory. Genomic DNA was extracted using standard chloroform extraction protocols (Sambrook, Fritsch & Maniatis, 1989). We amplified a 787 bp fragment of mitochondrial marker cytochrome b using primers Gludgl 5′-TGAYTTGAARAACCAYCGTTG-3′ and CB3H 5′-GGCAAATAGGAARTATCATTC-3′ (Palumbi et al., 1991). Thermocycler parameters were as follows: initial hold at 94 °C/5 min, 35 cycles of 94 °C/45 s, 45 °C/45 s, 72 °C/45 s, with a final extension of 72 °C/7 min. We developed species-specific primers for M. rosacea (MYCROS Forward: 5′-TTCTCCCACTACCCTGATTC-3′ and MYCROS Reverse: 5′-TACGTAGGCTTGGATCATTG-3′) to amplify a 726 bp fragment of mitochondrial marker ATPase. Thermocycler parameters were as follows: initial hold at 94 °C/5 min, 35 cycles of 94 °C/30 s, 54 °C/30 s, 72 °C/30 s, with a final extension of 72 °C/7 min. After purification of PCR products following ABI manufacturer’s protocols (ABI, Perkin-Elmer), we sequenced clean PCR products on an ABI 3730xl automated sequencer (Applied Biosystems, Foster City, CA).
We calculated molecular diversity indices including nucleotide diversity (π) and haplotype diversity (h). We corrected haplotype diversity using CONTRIB (Petit, El Mousadik & Pons, 1998) to account for differences in sample size between sites based on rarefaction to a minimum sample size of n = 4.
Phylogenetic relationships among sequences were inferred from a haplotype network based on pairwise differences between haplotypes generated using Arlequin (Excoffier, Laval & Schneider, 2005) and R software (R Core Team, 2013). To test for hierarchical population structure we performed an Analysis of Molecular Variance (AMOVA) in Arlequin. AMOVA significance was estimated using a permutation test of 10,000 replicates. The 11 sites were clustered into three regions: Baja Peninsula, the Midriff Islands and the Sonoran coast (Table 2). Chi-squared analyses were concurrently performed using DnaSP version 5.10 (Librado & Rozas, 2009) to test for patterns of regional subdivision. Pairwise FST and ϕST values were calculated between each location to assess how close populations are to alternate fixation. While FST considers all haplotypes identical and its maximum value decreases as the internal genetic diversity of population samples increases, ϕST effectively groups haplotypes considering their phylogenetic distance and is robust to increased within-populations genetic diversity (reviewed by Bird et al., 2011). When mutation rate is greater than migration rate and there is some restriction to gene flow (i.e., FST/ϕST < 1) the evolutionary relationship among haplotypes can provide additional resolution to explain where an haplotype is found among distinct populations (reviewed by Bird et al., 2011). However, ϕST has the potential to introduce noise in cases where migration rate is much greater than mutation rate and the evolutionary identity of individuals haplotypes is unrelated to the geographic location in which they are located (i.e., FST/ϕST > 1). We also calculated the standardized index (Meirmans & Hedrick, 2011) by dividing the original FST by the maximum value FST can achieve while maintaining the within-population diversity. Maximum values were calculated by recoding the alleles in such a way that every population only contained alleles unique to that population, following Meirmans & Hedrick (2011). In contrast to FST and ϕST which are fixation indexes, measures genetic differentiation and reaches its maximum when no alleles are shared between populations (Bird et al., 2011).
Table 2. Molecular diversity for each site with genetic samples.
For each site, we show their membership to one of three geographic groups, number of samples (n), number of haplotypes (nH), haplotype diversity (h), corrected haplotype diversity (h†) and nucleotide diversity (π).
| Location | Group | n | nH | h | h † | π |
|---|---|---|---|---|---|---|
| 1. La Poma | Baja | 16 | 12 | 0.950 ± 0.040 | 0.920 | 0.0016 ± 0.0010 |
| 2. La Ventana | Baja | 52 | 23 | 0.870 ± 0.035 | 0.885 | 0.0016 ± 0.0010 |
| 4. San Francisquito | Baja | 23 | 17 | 0.949 ± 0.034 | 0.920 | 0.0019 ± 0.0012 |
| 5. Punta Refugio | Midriffs | 25 | 9 | 0.840 ± 0.030? | 0.872 | 0.0015 ± 0.0010 |
| 9. San Lorenzo Island | Midriffs | 11 | 4 | 0.673 ± 0.123 | 0.844 | 0.0011 ± 0.0008 |
| 10. San Pedro Martir Island | Midriffs | 17 | 9 | 0.860 ± 0.068 | 0.881 | 0.0014 ± 0.0010 |
| 11. Datil Island | Midriffs | 20 | 13 | 0.947 ± 0.032 | 0.919 | 0.0020 ± 0.0010 |
| 12. San Esteban Island | Midriffs | 4 | 4 | 1.000 ± 0.177 | 0.943 | 0.0013 ± 0.0011 |
| 14. El Tecomate | Sonora | 26 | 16 | 0.935 ± 0.034 | 0.914 | 0.0019 ± 0.0010 |
| 16. Puerto Libertad | Sonora | 55 | 21 | 0.874 ± 0.033 | 0.887 | 0.0016 ± 0.0010 |
| 17. Puerto Lobos | Sonora | 11 | 7 | 0.909 ± 0.066 | 0.902 | 0.0018 ± 0.0012 |
We constructed a population graph from the distribution of haplotype frequencies with the software GENETICSTUDIO. A population graph is an analysis that allows genetic structure to define a graph-theoretic topology by capturing the high dimensional genetic covariance among all nodes considered simultaneously. An edge is placed between nodes that are conditionally dependent of each other, based on the remaining data in the model. With the empirical network, we calculated the same five node-based network metrics in NODEXL, and the adjacency and graph distance matrices (also known as conditional genetic distance, Dyer, Nason & Garrick, 2010) in GENETICSTUDIO as described above for the modeled networks.
To test if the ocean model could help to explain the empirical genetic data, we performed node-based and matrix-based analyses. Node base analyses were done fitting a linear model in R between empirical corrected haplotype diversity and nucleotide diversity within each site using in-degree, out-degree and betweenness centrality estimates from networks at each PLD as explanatory variables. Theory predicts that genetic diversity levels observed within sites is highly dependent upon the amount of migration from source populations (Gaggiotti, 1996), and that genetic diversity increases in sink sites that accumulate larvae and genetic variants from multiple sources (Kool et al., 2011). We also tested for a correlation between modeled and empirical estimates of betweenness centrality and clustering coefficient for each node with a linear regression. We predicted that if model estimates of node centrality depicted an accurate description of the actual connectivity among sites, modeled and empirical values should be significantly correlated.
Matrix-based analyses were done with Mantel tests implemented in the software IBDWS employing 10,000 randomizations (Jensen, Bohonak & Kelley, 2005). First, we tested whether larval dispersal fit a stepping-stone model where gene flow is limited by geographic distance by comparing the pairwise matrices of ϕST, FST and against geographic distances. Second, we predicted that the presence of a larval dispersal link between two nodes and its intensity (probability), as estimated from an adjacency matrix from the modeled networks, would reduce the level of genetic differentiation observed. We conducted a partial Mantel test between the modeled adjacency matrix and the empirical (log) ϕST, (log) FST and (log) matrices, respectively, while controlling for (log) geographic distance. Third, we predicted that, if oceanographic models represent an accurate description of connectivity patterns among sites, then sites connected by larger graph distances between two sites, considering the topology of the modeled network, would have larger levels of genetic structure. We used a partial Mantel test between modeled graph distances and the empirical (log) ϕST, (log) FST and (log) matrices, respectively, while controlling for (log) geographic distances. Fourth, under the same previous hypothesis, we performed a Mantel test comparing the empirical ϕST, FST and matrices, respectively, against the (log) graph distance from the modeled networks. When log-transforming genetic distances, negative values were excluded from the analyses. We predicted that if the modeled matrices were close to reality, then modeled and empirical matrices should be significantly correlated. We tested for a correlation between the modeled and empirical adjacency and graph distance matrices with linear regression analyses. Given the large number of tests performed in the node-base and matrix-based analyses, we corrected the critical P value 0.05 with the graphically sharpened method (Benjamini & Hochberg, 2000) to account for a false discovery rate in multiple tests within each analysis before assuming statistical significance.
We evaluated three different larval migration models based on the genetic data using Migrate-n 3.2.16 (Beerli & Palczewski, 2010). First, we tested an unrestricted full migration model between all sampling localities. Next, we considered two models with three population sizes comprised of a subsampling (n = 30) from sampling localities in the Baja Peninsula, the Midriff Islands, and the Sonoran coast. One model assessed the hypothesis of unidirectional gene flow from the Baja Peninsula across the Midriff Islands and towards the Sonoran coast, while the other model tested gene flow in the reverse direction. The latter two models reflect seasonal differences in directionality of a cyclonic and anticlyclonic gyres in the northern GC. Using a Bezier approximation, we chose the most appropriate model for our dataset by taking the natural log of the ratio of the marginal likelihoods (Bayes factors) for each model (Beerli & Palczewski, 2010). Running conditions for Migrate-n were as follows: 5,000,000 recorded steps, a burn-in of 2,500,000 steps, a static heating scheme using 20 temperatures, a tree swapping interval of 1, and an upper prior boundary for migration set to 7,500.
To further cross-validate the genetic and modeled connectivity data, we searched for statistical evidence of the presence of modularity or graph structure in the modeled and empirical networks based on the three geographic groups (Baja, Midriffs and Sonora). Analyses were done with GENETICSTUDIO, where we considered each group in a sub-graph one at a time, separated from the other two.
Results
Spawning sites, season and period
We identified 17 sites associated with 17 distinct spatial units for the release of virtual larvae in the simulation model based on direct and indirect evidence of the presence of spawning aggregations for leopard grouper (Table 1). Although some spatial units had evidence of multiple spawning aggregations, we assumed their close proximity along with the spatial resolution of the oceanographic model meant multiple aggregations within the same unit would disperse larvae in similar directions. Individual sites were distributed throughout the region and fulfilled a range of 3–7 criteria used to define a spawning aggregation site (Table 1), with an average of 4.76 criteria ± 1.48 (SD). A marked increase in the abundance of adult groupers during the spawning season relative to the non-spawning season was the most common evidence (recorded at all 17 sites). Other types of indirect evidence such as the observation of gravid females with swollen abdomens, observations of fish exhibiting courtship coloration, the collection of running-ripe males, or elevated catch rates by fishers during the spawning season were also recorded for the majority of sites. Direct evidence of spawning via the collection of hydrated females was recorded for 65% of the sites, whereas spawning was observed at only 35% of the sites.
Table 1. Study sites in the Gulf of California and selection criteria.
Each site was selected based on seven criteria to define where spawning aggregations might act like source of larvae. The order of sites follows the predominant cyclonic (anti-clockwise) circulation in the Midriff Island region during spawning of M. rosacea.
| Site # |
Site name | High abundance of fish |
Elevated catch rates |
Hydrated females collected |
Running-ripe males collected |
Courtship or spawning observed |
Gravid females observed |
Courtship coloration observed |
Criteria observed |
|---|---|---|---|---|---|---|---|---|---|
| 1 | La Poma | ✓ | ✓ | ✓ | 3 | ||||
| 2 | La Ventana | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 |
| 3 | Chorros | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | ||
| 4 | San Francisquito | ✓ | ✓ | ✓ | ✓ | 4 | |||
| 5 | Punta Refugio | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 |
| 6 | Los Machos | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | |
| 7 | Punta Diablo | ✓ | ✓ | ✓ | ✓ | 4 | |||
| 8 | Punta Roja | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | |
| 9 | San Lorenzo Island | ✓ | ✓ | ✓ | ✓ | 4 | |||
| 10 | San Pedro Martir Island | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | |
| 11 | Datil Island | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 |
| 12 | San Esteban Island | ✓ | ✓ | ✓ | 3 | ||||
| 13 | La Tordilla | ✓ | ✓ | ✓ | ✓ | 4 | |||
| 14 | El Tecomate | ✓ | ✓ | ✓ | ✓ | 4 | |||
| 15 | Las Cuevitas | ✓ | ✓ | ✓ | ✓ | 4 | |||
| 16 | Puerto Libertad | ✓ | ✓ | ✓ | ✓ | 4 | |||
| 17 | Puerto Lobos | ✓ | ✓ | ✓ | ✓ | 4 | |||
| # Sites | 17 | 13 | 11 | 12 | 6 | 12 | 11 |
A total of 162 samples of female M. rosacea were collected from commercial fishers, with an average of 14 samples collected each month (range = 8–27). Based on microscopic examinations of gonadal tissue samples, females in the spawning capable phase were collected from March through June, and actively spawning females were collected mainly during May and June. The GSI of adult females showed elevated levels from April to June, with a peak during May (Fig. 2). When the results of the gonadal phases and GSI were combined, they indicate that M. rosacea spawn from April to June in the Midriffs region, with peak spawning activity occurring in May and June. Actively spawning females were collected on three days in 2009 (14 May, 31 May, 25 June) and two days in 2010 (25 April, 7 May).
Figure 2. Spawning season and period.

Monthly proportion of actively spawning and spawning capable females (left y axis) and female gonadosomatic index (GSI, right y axis) for the Midriff Islands collected in 2009.
Modeled connectivity
From all simulated particles released (4,000 particles × 17 sites × eight release dates = 544,000), coastal areas that are suitable for larval recruitment captured 55.13% (PLD 14 days), 48.07% (PLD 21 days) and 42.03% (PLD 28 days). Remaining particles did not reach any coastal habitat by the end of the PLD. Simulations of ocean currents produced the highest concentrations of larvae in: (1) Canal de Ballenas, (2) north end of Angel de la Guarda Island, (3) north end of Tiburon Island, and (4) in coastal areas located to towards the north after PLD 28 days (around Las Cuevitas and Puerto Libertad Fig. 3). Other important areas were located at the south end of Tiburon Island and Puerto Lobos. Simulations at PLD 14 and 21 days indicated similar trends (Fig. 3).
Figure 3. Probability of larval density.

Density of all virtual larvae released in coastal areas after PLD 14 days (A), 21 days (B) and 28 days (C) in each coastal spatial unit of analysis.
The trajectories of particles released from each site at the eight release dates (Video S1) showed sites in the Baja California Peninsula (sites 1–4) generally followed a southward direction in early May that gradually shifted to a northward direction during late May and June, while sites at the north edge of Tiburon Island (Site 14) and on mainland Sonora (sites 15–17) always followed a northward trajectory but the distance traveled by particles increased from May towards June. Most locations around the Midriff islands followed the same pattern described for sites 1–4, with the exception of San Pedro Martir Island and Datil island near the south end of Tiburon Island that had particles dispersing both north and south of the release site. In all cases, the distance traveled by particles was directly proportional to the PLD, which is not always true in such studies (e.g., Selkoe et al., 2010). The graph size of the connectivity networks increased from 57 edges at PLD 14 days, to 94 at PLD 21 days and 132 at PLD 28 days, indicating a longer PLD is associated with more connected and complex networks (Figs. 4A–4C). Based on the directionality of the links in the networks, three main patterns were evident. First, the proportion of southward links decreased from 22.8% at PLD 14, to 9.57% at PLD 21 and 1.51% at PLD 28; the proportion of northward and bi-directional (north–south) links was similar at all PLDs (ranging from 38.59% to 51.51%). Second, at PLD 14, northward links were prevalent along the sites located in the coast of Baja, in the Canal de Ballenas and between the southern end of Angel de la Guarda Island across the GC towards the northern end of Tiburon Island and mainland Sonora; southward links were present between the eastern and southern coasts of Angel de la Guarda Island towards southern locations in San Lorenzo Island and Baja California and across the Midriff Islands to Tiburon Island and San Esteban Island. Third, bi-directional links were evident in a small area located between San Pedro Martir Island, San Esteban and San Lorenzo Islands and the southern end of Tiburon Island at PLD 14; at PLD 21 they were found almost exclusively within sites in the Midriffs, while at PLD28 they were found at sites within Baja, within the Midriffs and between Baja and the Midriffs. In all PLDs, sites in Sonora received larvae from southern sites and were not involved in bi-directional links (Figs. 4A–4C). Overall, the strongest links (i.e., those showing the larger probabilities) were observed between the southern, western and northern coasts of Tiburon Island towards northern localities situated in mainland Sonora (Las Cuevitas and Puerto Libertad). Strong links were also present in the Canal de Ballenas at PLD 14 and PLD 21 (Figs. 5A and 5B).
Figure 4. Modeled and empirical networks of larval connectivity.

Spatial networks of larval dispersal between sites for PLD 14 days (A), 21 days (B) and 28 days (C), and conditional genetic distance from a population graph based on mtDNA sequence data (D), showing dispersal events (links) between sites (nodes). In (A)–(C) line width is proportional to probability, according to the scale to the right; the direction of the larval dispersal events is indicated by different colors: northward (red), southward (blue) or both simultaneously (green). The empirical network is undirected and line width is proportional to conditional genetic distance according to the scale. Black nodes belong to “Baja” geographic group, grey nodes belong to “Midriffs” and blank nodes to “Sonora”.
Figure 5. Performance of 17 sites for different aspects of marine connectivity.

Performance was measured with export probability, import probability, local-retention probability, out-degree, in-degree and clustering coefficient, as estimated by an oceanographic model at PLD 14 and 28 days.
The probability of local retention decreased with increasing PLD. According to the ocean model, local retention was most likely at the northern end of Angel de la Guarda Island (Punta Refugio, range: 0.41–0.28 for PLD 14 and 28 days, respectively), the northern end of Tiburon Island (El Tecomate, range: 0.54–0.21), followed by the southern coast of Tiburon Island (Datil Island, range: 0.23–0.19), San Esteban Island (range: 0.22–0.23) and Puerto Libertad (range: 0.38–0.16).
Our analyses illustrated that several of the connectivity metrics covary (including export, import and local retention probabilities), highlighting a few sites that were important despite variation in PLD (Fig. 5). For example, Puerto Refugio at the north of Angel de la Guarda Island and Tecomate on the north of Tiburon Island had among the largest probabilities of export, import and local retention. The sites with the largest probabilities of exporting larvae where located downstream, from site 11 (San Esteban Island) to 16 (Puerto Libertad), while those sites with the largest probabilities of importing larvae where located even more downstream matching the Sonora group (sites 14–16, with the exception of Puerto Refugio). In the graph theoretic analyses, out-degree peaked at the Baja group (sites 2 and 3) and within the Midriffs group (sites 7–12), while the Sonora group had overall low values. In contrast, in-degree was relatively high over the entire region, but particularly at downstream sites (11–16, Datil Island to Puerto Libertad). Clustering coefficient was highest at the most downstream site (Puerto Libertad) (Fig. 5). Betweenness centrality identified Las Cuevitas on mainland Sonora, the Southern end of Angel de la Guarda Island and La Ventana as key sites for multigenerational larval dispersal through the entire network (results not shown).
Genetic connectivity
We analyzed a 787 bp fragment of cytochrome b and a 726 bp fragment of ATPase for 260 individuals. We identified a total of 79 haplotypes (GenBank accesion numbers KJ004770–KJ004925), with adjacent haplotypes in the haplotype network separated by 1–4 bp (Fig. 6). There was limited evidence of geographic separation of haplotypes, and the three most frequent haplotypes were present in all locations, with the exception of the third most frequent which was absent in San Francisquito on the Baja Peninsula. Corrected estimates of haplotype diversity were high, ranging from 0.844 (San Lorenzo Island) to 0.943 (San Esteban Island) (Table 2). Nucleotide diversity was low, varying from 0.0011 (San Lorenzo Island) to 0.0020 (Datil Island).
Figure 6. Minimum spanning network among haplotypes.

The network shows the relationships among 79 haplotypes found in M. rosacea. Circles are sized proportionally to the number of individuals that possess each haplotype and colors indicate their geographic distribution in 11 sites shown to the left. All haplotypes are separated by one to four mutation steps as denoted by scaling provided.
Statistically significant pairwise estimates of genetic structure were observed (Table 3). Pairwise FST, ϕST, and values suggest Puerto Libertad is genetically divergent from the majority of other sampling localities, while La Poma and San Esteban Island are the most similar to all the other sites (Table 3, Table S1). Global estimates suggest moderate levels of population structure within the northern Gulf (FST = 0.016, P = 0.011; ϕST = 0.0467, P = 0.00059; ). The FST/ϕST ratio based on global values was 0.3426, indicating insufficient gene flow to homogenize populations and that closely related haplotypes show some geographical structure despite occasional long distance dispersal (Pons & Petit, 1996; Bird et al., 2011).
Table 3. Pairwise FST and ϕST statistics between sites.
Pairwise FST values are above diagonal and pairwise ϕST values are below diagonal. For each site, we show their membership to one of three geographic groups. P values below 0.05 are shown in bold.
| Group | Baja | Midriffs | Sonora | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | 1 | 2 | 4 | 5 | 9 | 10 | 11 | 12 | 14 | 16 | 17 |
| 1. La Poma | – | −0.011 | −0.018 | −0.016 | 0.054 | −0.017 | −0.022 | −0.090 | −0.008 | 0.012 | −0.023 |
| 2. La Ventana | −0.0124 | – | −0.004 | 0.004 | 0.030 | 0.011 | 0.009 | −0.054 | 0.025 | 0.025 | 0.004 |
| 4. San Francisquito | −0.0176 | −0.0045 | – | 0.009 | 0.057 | 0.011 | −0.005 | −0.062 | 0.012 | 0.020 | −0.001 |
| 5. Punta Refugio | −0.031 | 0.012 | −0.002 | – | 0.080 | −0.009 | 0.025 | −0.052 | 0.003 | 0.055 | 0.017 |
| 9. San Lorenzo Island | 0.0725 | 0.0381 | 0.0193 | 0.070 | – | 0.098 | 0.043 | 0.042 | 0.118 * | 0.011 | 0.003 |
| 10. San Pedro Martir Island | −0.0196 | −0.0122 | −0.0055 | 0.015 | 0.0869 | – | 0.015 | −0.097 | 0.000 | 0.061 | 0.004 |
| 11. Datil Island | −0.0169 | −0.0001 | −0.0129 | 0.042 | −0.0090 | −0.0018 | – | −0.062 | 0.017 | 0.009 | −0.022 |
| 12. San Esteban Island | −0.0578 | −0.0421 | −0.0455 | 0.017 | 0.0772 | −0.0391 | −0.0605 | – | −0.041 | −0.020 | −0.069 |
| 14. El Tecomate | 0.0038 | 0.0101 | 0.0092 | 0.008 | 0.0621 | −0.0094 | 0.0169 | 0.0203 | – | 0.060 | 0.026 |
| 16. Puerto Libertad | 0.0891 | 0.0912 | 0.0601 | 0.091 | −0.0094 | 0.1189 * | 0.0408 | 0.0386 | 0.1234 * | – | −0.015 |
| 17. Puerto Lobos | 0.0554 | 0.0567 * | 0.0193 | 0.054 | −0.0353 | 0.0555 | −0.0035 | 0.0193 | 0.0433 | 0.0122 | – |
Notes.
Indicate statistical significance after correcting for multiple tests (critical P = 0.0009).
The population graph derived from the genetic covariance among sites included all 11 empirical sites and had a graph size with 17 edges (Fig. 4D). Despite its smaller size and complexity compared to the modeled networks, its topology resembled some of the links suggested by the models, particularly at PLD 28 days (e.g., the links between Puerto Refugio on the north of Angel de la Guarda Island and: La Poma and La Ventana on the Baja Peninsula, and El Tecomate on the north of Tiburon Island; the links between San Esteban Island and: El Tecomate, Datil Island, San Pedro Martir Island and San Francisquito on the Baja Peninsula).
Node-base analyses via fit of a linear model for explaining haplotype and nucleotide diversity using in-degree, out-degree and betweenness centrality at each PLD as explanatory variables revealed that nucleotide diversity was lower at locations showing higher out-degree according to the model at PLD 14 days (P = 0.0318, Table 4, Fig. 7A) and PLD 21 days (P = 0.0305), but patterns were not significant after correcting for multiple tests using FDR (critical P = 0.0029). No other comparisons were significant. The linear regression between modeled and empirical values of betweenness centrality and clustering coefficient at each PLD showed only the clustering coefficient estimates were correlated at PLD 21 days (P = 0.0487, R2 = 0.3655, Table 5, Fig. 7B) and PLD 28 days (P = 0.0019, R2 = 0.6729). After correcting for multiple tests, only patterns at PLD 28 days were significant (critical P = 0.0083).
Table 4. Node-based analyses for explaining genetic diversity.
Results of fitting a linear model for explaining empirical corrected haplotype diversity and nucleotide diversity with modeled in-degree, out-degree and betweenness centrality as explanatory variables at three PLDs. For each variable we show the coefficient estimated in the model, its associated P value, the Incidence Risk Ratio (IRR) coefficient and the 95% lower (L) and upper (U) confidence intervals. For each model tested, we show the multiple R2 value and the associated P value. P values below 0.05 are shown in bold.
| Corrected haplotype diversity (h†) | Nucleotide diversity (π) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Coefficient | P-value | IRR | L | U | Coefficient | P-value | IRR | L | U | |
| PLD 14 | ||||||||||
| In-degree | 0.0048314 | 0.408 | 1.004843 | 0.9984597 | 1.011267 | 6.12E–05 | 0.1775 | 1.0000612 | 1.0000055 | 1.0001169 |
| Out-degree | −0.0033951 | 0.458 | 0.9966107 | 0.9894915 | 1.003781 | −8.61E–05 | 0.0318 | 0.9999139 | 0.99986 | 0.9999678 |
| Betweenness | 0.0002055 | 0.84 | 1.0002055 | 0.9988981 | 1.001515 | 9.74E–06 | 0.2229 | 1.0000097 | 0.9999954 | 1.0000241 |
| Multiple R2 | 0.1445 | 0.7614 | 0.5147 | 0.1459 | ||||||
| PLD 21 | ||||||||||
| In-degree | 0.0007668 | 0.871 | 1.0007671 | 0.9970901 | 1.0044577 | 4.51E–05 | 0.214 | 1.0000451 | 1.0000094 | 1.0000809 |
| Out-degree | −0.0012069 | 0.616 | 0.9987938 | 0.994191 | 1.003418 | −4.53E–05 | 0.0305 | 0.9999547 | 0.9999308 | 0.9999787 |
| Betweenness | −0.0013393 | 0.23 | 0.9986616 | 0.9982158 | 0.9991075 | −1.26E–05 | 0.1319 | 0.9999874 | 0.9999784 | 0.9999964 |
| Multiple R2 | 0.2277 | 0.5875 | 0.5816 | 0.09045 | ||||||
| PLD 28 | ||||||||||
| In-degree | −0.002588 | 0.631 | 0.9974157 | 0.9895889 | 1.005305 | 5.04E–05 | 0.275508 | 1.0000504 | 0.9999974 | 1.0001033 |
| Out-degree | −0.000758 | 0.71 | 0.9992423 | 0.9955642 | 1.002934 | −3.23E–05 | 0.085887 | 0.9999677 | 0.9999465 | 0.9999889 |
| Betweenness | −0.001097 | 0.726 | 0.998904 | 0.9947085 | 1.003117 | −4.08E–05 | 0.144042 | 0.9999592 | 0.9999285 | 0.9999899 |
| Multiple R2 | 0.23 | 0.5829 | 0.4607 | 0.2037 | ||||||
Notes.
Indicate statistical significance after correcting for multiple tests (critical P = 0.0029).
Figure 7. Node-base analyses.

(A) Scatter plot between modeled estimates of out-degree at three PLDs and observed values of nucleotide diversity. (B) Scatter plot between modeled and empirical estimates of clustering coefficient.
Table 5. Node-based analyses correlating modeled and empirical network metrics.
Linear regression analyses between two node-base metrics (betweenness centrality and clustering coefficient) estimated from the modeled and empirical networks. P values below 0.05 are shown in bold.
| PLD | Node-base metric | R 2 | P value |
|---|---|---|---|
| PLD 14 days | Betweenness centrality | 0.0060 | 0.8196 |
| Clustering coefficient | 0.1201 | 0.2963 | |
| PLD 21 days | Betweenness centrality | 0.0699 | 0.4319 |
| Clustering coefficient | 0.3655 | 0.0487 | |
| PLD 28 days | Betweenness centrality | 0.1189 | 0.2990 |
| Clustering coefficient | 0.6729 | 0.0019 * |
Notes.
Indicate statistical significance after correcting for multiple tests (critical P = 0.0083).
Matrix-based analyses showed a lack of correlation between pairwise ϕST, FST and values against geographic distance (Mantel test P = 0.386, R2 = 0.0004, Table 6; P = 0.8080, R2 = 0.0262, Table S2; P = 0.6970, R2 = 0.0056, Table S3, respectively). In contrast, we found significant correlations supporting our predictions at all PLDs between the adjacency and graph distance matrices calculated from the modeled networks and the empirical log ϕST matrix, while controlling for geographic distance (Table 6). First, genetic differentiation decreased between sites with larger adjacency values indicative of high larval dispersal probability, particularly at PLD 21 days (partial Mantel test P < 0.0001, R2 = 0.1213, Fig. 8A). Second, genetic differentiation increased between sites with larger graph distance, especially at PLD 28 days (partial Mantel test P < 0.0001, R2 = 0.0684). The ϕST matrix and the (log) graph distance matrix were also significantly correlated at PLD 28 days (Mantel test P = 0.0060, R2 = 0.2224, Fig. 8B), further corroborating a significant trend of low genetic structure associated with nodes located nearby according to the topology of the modeled network of larval dispersal. Similar analyses employing FST and values showed significant patterns only when compared against adjacency values, particularly at PLD 14 days (FSTP < 0.0001, R2 = 0.1177, Table S2; , R2 = 0.1423, Table S3). Additionally, while modeled and empirical graph distance matrices were not significantly correlated at any PLD according to linear regression analyses (All P values ≥ 0.1402, all R2 ≤ 0.0405, Table 7), the adjacency matrices were significantly correlated at PLD 21 days (Linear regression P = 0.0065, R2 = 0.1315).
Table 6. Matrix-based analyses for explaining patterns of genetic structure.
Mantel and partial Mantel tests between an empirical matrix of genetic structure (ϕST) values and various explanatory variables, including geographic distance and three distinct matrices calculated from the modeled networks, including adjacency, graph distance and log graph distance. P values below 0.05 are shown in bold.
| Matrix 1 | Matrix 2 | Controlling matrix |
P value | R 2 | |
|---|---|---|---|---|---|
| ϕST | GeoD | – | 0.386 | 0.0004 | |
| PLD 14 | log ϕST | Adjacency | log GeoD | <0.00001 * | 0.0660 |
| log ϕST | GraphD | log GeoD | <0.00001 * | 0.0371 | |
| ϕST | log GraphD | – | 0.202 | 0.0306 | |
| PLD 21 | log ϕST | Adjacency | log GeoD | <0.0001 * | 0.1213 |
| log ϕST | GraphD | log GeoD | <0.0001 * | 0.0241 | |
| ϕST | log GraphD | – | 0.29 | 0.0123 | |
| PLD28 | log ϕST | Adjacency | log GeoD | <0.0001 * | 0.0487 |
| log ϕST | GraphD | log GeoD | <0.0001 * | 0.0684 | |
| ϕST | log GraphD | – | 0.0060 * | 0.2224 |
Notes.
Indicate statistical significance after correcting for multiple tests (critical P = 0.0060).
Figure 8. Matrix-based analyses.

(A) Scatter plot between adjacency values from the modeled network at PLD 21 days and empirical log ϕST values. (B) Scatter plot between log graph distances estimated from the modeled network at PLD 28 days and empirical ϕST values.
Table 7. Matrix-based analyses correlating empirical and modeled networks.
Linear regression analyses between two types of matrices (adjacency and graph distance) estimated from the modeled and the empirical networks. P values below 0.05 are shown in bold.
| PLD | Matrix | R 2 | P value |
|---|---|---|---|
| PLD 14 days | Adjacency | 0.0367 | 0.1608 |
| Graph distance | 0.0116 | 0.4334 | |
| PLD 21 days | Adjacency | 0.1315 | 0.0065 * |
| Graph distance | 0.0229 | 0.2693 | |
| PLD 28 days | Adjacency | 0.0187 | 0.3193 |
| Graph distance | 0.0405 | 0.1402 |
Notes.
Indicate statistical significance after correcting for multiple tests (critical P = 0.0083).
Regional genetic subdivision was observed when sampling sites were clustered into the Baja Peninsula, Midriff Islands and Sonoran coast groups, as supported by a chi-squared test (χ2 = 186.876, d.f. = 152, P = 0.0286) and an AMOVA test (FST = 0.0466, P < 0.05, Table S4). Rankings of proposed larval dispersal models between the aforementioned regions are listed in Table 8. The best-supported model of larval migration based on the genetic data was for unidirectional larval dispersal from the Baja Peninsula towards Sonora (File S1–S3). The analysis of modularity revealed the presence of two statistically supported sub-graphs in the modeled networks at all PLDs but with higher support at PLD 21 (Table 9, P = 0.0003). The first segment consisted of Baja + Midriffs and the second segment included Sonora. The empirical network did not show any evidence of modularity.
Table 8. Probability of three larval dispersal models based on the genetic data.
Bayes factors and marginal log likelihoods estimated in Migrate-n version 3.2.16 using Bayesian approximation and thermal integration for three proposed larval dispersal models including: an unrestricted (Full matrix) larval dispersal among three groups (Baja, Midriffs and Sonora), a directional model of larval dispersal from Baja across the Midriffs towards Sonora (Baja to Sonora), and a directional model from Sonora across the Midriffs towards Baja (Sonora to Baja).
| Model | Bezier l mL |
Harmonic l mL |
Choice (Bezier) |
Model probability |
|---|---|---|---|---|
| Full matrix | −2673.20 | −2514.33 | 2 | 0.00001 |
| Baja to Sonora | −2658.64 | −2513.07 | 1 | 0.99999 |
| Sonora to Baja | −2974.33 | −2529.21 | 3 | 0.00000 |
Table 9. Probability of sub-graph structure.
P-values supporting the presence of sub-graph structure among three defined geographic groups (Baja, Midriffs and Sonora) for the modeled connectivity networks at PLD 14 days, PLD 21 days and PLD 28 days, and the empirical network from genetic data. Distinct segments of the proposed sub-graphs are separated by a “/” symbol, while a “+” symbol denotes groups considered as a single segment. P values below 0.05 are shown in bold.
| Sub-graph | PLD 14 days |
PLD 21 days |
PLD 28 days |
Empirical network |
|---|---|---|---|---|
| Baja / Midriffs + Sonora | 0.4724 | 0.1732 | 0.3645 | 0.6064 |
| Baja + Midriffs / Sonora | 0.0006 * | 0.0003 * | 0.0017 * | 0.4086 |
| Midriffs / Baja + Sonora | 0.1074 | 0.0439 | 0.1739 | 0.9940 |
Notes.
Indicate statistical significance after correcting for multiple tests (critical P = 0.0041).
Discussion
Our study contributes to a growing body of literature (Galindo et al., 2010; Selkoe et al., 2010; Alberto et al., 2011; Crandall, Treml & Barber, 2012; Foster et al., 2012; Petitgas et al., 2012; Soria et al., 2012; Feutry et al., 2013) highlighting the inherent value of verifying outputs of biophysical oceanographic models with empirical genetic data to inform larval dispersal patterns and marine connectivity. We corroborated our hypothesis that an oceanographic model describing metapopulation dynamics of larval dispersal in the GC can help to explain empirical genetic patterns. The model helped to explain genetic differences between sites but not genetic diversity within sites, which could result from the combination of using a single relatively slow evolving genetic marker (a protein-coding mitochondrial DNA marker) and the fact that genetic variation between sites evolves more slowly than within sites (Keyghobadi et al., 2005). We predict that the use of multiple fast-evolving and hypervariable markers (e.g., microsatellites), could improve the power of the model to predict genetic diversity within sites. Nevertheless, significant correlations between node-based modeled and empirical clustering coefficient suggest that summarizing genetic data with a population graph increased the power of the model to explain the empirical data. Validation of the passive dispersal model through subsequent studies such as those using parentage analyses and highly polymorphic microsatellite loci are recommended and underway.
Concordance of pairwise genetic differences and matrices describing networks derived from biophysical modeling data for M. rosacea elucidate the role of oceanographic processes in driving patterns of larval dispersal while models helped to explain seemingly chaotic patterns of genetic structure. Our study shows that in addition to contrasting pairwise values of genetic structure to modeled probabilities (Selkoe et al., 2010), or modeled oceanographic distances (White et al., 2010b; Alberto et al., 2011), graph distances over modeled networks or larval dispersal are a simple and promising tool to summarize marine connectivity patterns. The model using graph distances explained a larger variance in ϕST values than the model using adjacency matrices, indicating that although the presence or absence of a link and its probability are relevant, even more important are the topological order and relationships among sites driven by the prevailing oceanographic currents. For instance, Puerto Libertad on mainland Sonora showed the largest genetic differences compared to other sites, which could be explained by its extreme downstream position (i.e., large graph distance values with most sites), along with large levels of local retention according to the model (Fig. 5) that could have contributed to a high proportion of kin than expected by chance and to high levels of genetic differentiation (Iacchei et al., 2013).
Although the Baja, Midfriffs and Sonora groups were significantly differentiated, the best supported gene flow model (based on genetic subdivisions) agreed with the general cyclonic (anti-clockwise) direction of the gyre in the northern GC during the spawning period of M. rosacea that transports larvae from the Baja Peninsula, across the GC and towards the Sonoran coastline. This empirical result is in line with the analysis of graph modularity on the modeled networks that strongly suggested a larger bi-directional exchange of larvae between the Baja and the Midriffs group that were relatively separated from the Sonora group that was a net importer (sink) of larvae from upstream sites.
Our study had several limitations, with the major ones discussed below. While there is no direct evidence that leopard grouper larvae vertically migrate to escape advecting currents, there is a growing body of evidence that suggests that local retention of larvae in groupers can be quite high (Harrison et al., 2012; Almany et al., 2013). Thus our passive model could have overestimated larval exchange rates and underestimated local retention (Cowen et al., 2000; Largier, 2003; Leis, 2007). The effects of vertical migration could sometimes be comparable to those of reducing PLD (Andrello et al., 2013), or substantially increase or decrease dispersal (Woodson & McManus, 2007). Investigations into larval behavior of groupers are warranted and could greatly increase the precision and accuracy of the model. Our models did not include an explicit description of the habitat for larval recruitment, such as beads of the macro algae Sargassum sp. (Aburto-Oropeza et al., 2007), nor did they consider larval mortality after settlement. Thus, we only assessed potential connectivity, as opposed to realized connectivity (sensu Watson et al., 2010).
By coupling modeled and empirical connectivity approaches, we are able to better understand the mechanisms driving dispersal in the GC and inform spatially explicit management efforts for M. rosacea as well as marine organisms with similar life histories. Our multidisciplinary approach provided a mechanistic explanation of why some areas in the Midriff Island region concentrate the fishing effort for leopard grouper in the GC. Most of the largest and most heavily targeted fishing areas, including the north end of Angel de la Guarda Island, the west, north and south edges of Tiburon Island and Las Cuevitas and Puerto Libertad on mainland Sonora (Moreno-Baez et al., 2010; Moreno-Báez et al., 2012), showed the largest values of local retention of larvae, together with a high probability of importing larvae from other spawning sites and for concentrating larvae from all over the region. Notably, some of these sites are known to historically hold huge spawning aggregations of leopard grouper that have been harvested at high levels for decades, like the north end of Angel de la Guarda Island (Cannon, 1966). Thus, the main fishing areas seem to depend simultaneously on both local retention and contributions of larvae from upstream sites, coupled to oceanographic patterns that focus larval density towards these areas that sustain most of the fisheries.
A key result of our study is the observation that marine connectivity for M. rosacea from Baja California Peninsula and the Midriff island region towards Sonora is predominantly asymmetric. Other studies have previously shown the negative effects that asymmetric connectivity has on population persistence (Bode, Burrage & Possingham, 2008; Vuilleumier, Bolker & Leveque, 2010). In the presence of strong asymmetric currents, reserves (no-take zones) can significantly outperform traditional quota based management strategies in terms of fisheries yield, with considerably less risk (Gaines, Gaylord & Largier, 2003). Asymmetry also constrains the notion that benefits of reserves in terms of larval input are proportional to their distance to the reserve (Almany et al., 2009; Buston et al., 2012). For example, one study using DNA parentage analyses found that reserves in the Great Barrier Reef, which accounted for 28% of the local reef area, produced approximately half of all juvenile recruitment of snappers and groupers to both reserve and fished reefs within 30 km of the source spawning site inside the reserve (Harrison et al., 2012). A similar study in Papua New Guinea used parentage to track larval dispersal and predicted that 50% of larvae in a coral grouper settled within 13 km of the spawning aggregation sites (Almany et al., 2013). In contrast, the benefits of reserves in the Midriff Island region are completely biased towards one particular direction towards Sonora, highlighting that the spatial location of no-take zones to ensure connectivity is even more important than the number of zones, compared to other systems.
A network of no-take zones within the Midriff region might very well have a defined zone of influence that does not include the eastern edge of Tiburon Island or any locality towards the south in mainland Sonora. This observation has important practical implications. For example, fishing localities on mainland Sonora South of Tiburon Island are restricted from fishing at no-take areas within MPAs in the Midriffs, yet according to this and another study (Soria et al., 2014) localities south of Tiburon Island receive few benefits by not fishing there. Conversely, fishing communities in mainland Sonora (Puerto Lobos, Puerto Libertad) seem to receive great benefits from San Pedro Martir, San Esteban and Tiburon Islands, even though fishers from Puerto Lobos and Puerto Libertad may not fish there. These observations bring up an important concept in highly advective systems like the GC where there may be a spatial disconnect and strong directionality between the location of no-take zones and the areas that benefit most from them, and highlights that in order for such reserves to be effective they need to be located upstream of targeted fishing sites (Beger et al., 2010). Non-traditional approaches, such as “larval credits” based on regional larval export production (Kough, Paris & Butler, 2014) could help to manage such trade-offs. Our analyses suggest that establishment of smaller no-take zones at the north end of Angel de la Guarda Island within the current MPA will likely boost local fisheries via local retention, while other existing no-take zones within the Canal de Ballenas and San Lorenzo MPA could export larvae to fishing sites across the GC. The establishment of additional no-take zones adjacent to currently heavily fished areas in the western and northern edges of Tiburon Island, and in the coast between Las Cuevitas-Puerto Lobos will likely increase productivity of local fisheries (via local retention) and also fisheries at downstream fished sites on mainland Sonora as north as Puerto Peñasco (located ∼300 km from Tiburon Island) via larval dispersal. Notably, except for San Francisquito on the coast of Baja California, current MPAs do not include downstream sink sites receiving larvae from multiple sources which harbor the largest genetic diversity and evolutionary potential (San Esteban and Tiburon Islands). Our findings highlight that important upstream sites for improving fisheries are not necessarily aligned spatially with other criteria for protection, such as preserving evolutionary potential via genetic variation.
Supplemental Information
The map shows 17 sites where virtual larvae were released (colored polygons) and 11 sites where genetic samples were also collected (blue polygons).
{kind=link}
Tides in the HAMSOM oceanographic model for the northern Gulf of California, showing the four neap and four spring tides during May and June selected for the release of virtual larvae. Red lines separate distinct months.
{kind=link}
Pairwise maximum FST possible while maintaining the within-population diversity above diagonal and pairwise values below diagonal. For each site, we show their membership to one of three geographic groups.
Mantel and partial Mantel tests between an empirical matrix of genetic structure (FST) values and various explanatory variables, including geographic distance and three distinct matrices calculated from the modeled networks, including adjacency, graph distance and log graph distance. P values below 0.05 are shown in bold. * Indicate statistical significance after correcting for multiple tests (critical P = 0.0060).
Mantel and partial Mantel tests between an empirical matrix of genetic structure () values and various explanatory variables, including geographic distance and three distinct matrices calculated from the modeled networks, including adjacency, graph distance and log graph distance. P values below 0.05 are shown in bold. * Indicate statistical significance after correcting for multiple tests (critical P = 0.0060).
Analysis of molecular variance testing for regional genetic subdivision when sampling sites were clustered into the Baja Peninsula, Midriff Islands and Sonoran coast groups.
Maps showing the spatial distribution of larvae released at each of eight release dates corresponding to four neap and four spring tides during May and June at each of 17 sites (“X” in each box) for PLD 14, 21 and 28 days. Scale in kilometers.
Detailed results from three independent runs with Migrate-n software for unrestricted gene flow among Baja, the Midriff Islands and Sonora.
Detailed results from three independent runs with Migrate-n software for a model of gene flow from Baja across the Midriff Islands towards Sonora.
Detailed results from three independent runs with Migrate-n software for a model of gene flow from Sonora across the Midriff Islands towards Baja.
Acknowledgments
We would like to acknowledge Rene Loiaza, Ivan Martinez, Angeles Cruz, Mario Rojo, Kimberly Tenggardjaja, Eva Salas, Gary Longo, Emmett Ziegler and Ana Liedke for their assistance with acquiring samples in the field. This is a scientific contribution of the multidisciplinary PANGAS project (http://cobi.org.mx/en/programas/regions/midriff-islands/pangas). We thank Kimberly A. Selkoe, Robert J. Toonen and an anonymous reviewer for insights that much improved the manuscript.
Funding Statement
This work was funded by the David and Lucile Packard Foundation, Marisla Foundation, Sandler Family Supporting Foundation, National Geographic (Young Explorers Grant #8928-11), the University of California Office of the President (UC-HBCU Initiative grant), Marine Conservation Institute (Mia J. Tegner memorial research grant), Ecology Project International, CenTread, the Dr. Earl H. Myers and Ethel M. Myers Oceanographic and Marine Biology Trust, FCT (SFRH/BPD/26901/2006) and the Friends of Long Marine Lab. Additional funding for this research was provided by Dr. Exequiel Ezcurra through the Pew Fellowship Program on Marine Conservation, and several grants from the Walton Family Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
Adrian Munguia-Vega is an employee of Comunidad y Biodiversidad A.C.
Author Contributions
Adrian Munguia-Vega, Alexis Jackson and Brad Erisman conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Silvio Guido Marinone conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables, reviewed drafts of the paper.
Marcia Moreno-Baez performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables, reviewed drafts of the paper.
Alfredo Girón-Nava performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, reviewed drafts of the paper.
Tad Pfister conceived and designed the experiments, contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Octavio Aburto-Oropeza contributed reagents/materials/analysis tools, wrote the paper, reviewed drafts of the paper.
Jorge Torre conceived and designed the experiments, contributed reagents/materials/analysis tools, wrote the paper, reviewed drafts of the paper.
Animal Ethics
The following information was supplied relating to ethical approvals (i.e., approving body and any reference numbers):
We obtained approval of our sampling protocol from the University of California Santa Cruz IACUC protocol Berng1101.
Field Study Permissions
The following information was supplied relating to field study approvals (i.e., approving body and any reference numbers):
Scientific collection permits were acquired from SAGARPA (No. DGOPA.09151.260809.2885 and DAPA/2/020511/01197).
DNA Deposition
References
- Aburto-Oropeza et al. (2011).Aburto-Oropeza O, Erisman B, Galland GR, Mascarenas-Osorio I, Sala E, Ezcurra E. Large recovery of fish biomass in a no-take marine reserve. PLoS ONE. 2011;6:e511. doi: 10.1371/journal.pone.0023601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aburto-Oropeza et al. (2007).Aburto-Oropeza O, Sala E, Paredes G, Mendoza A, Ballesteros E. Predictability of reef fish recruitment in a highly variable nursery habitat. Ecology. 2007;88:2220–2228. doi: 10.1890/06-0857.1. [DOI] [PubMed] [Google Scholar]
- Alberto et al. (2011).Alberto F, Raimondi PT, Reed DC, Watson JR, Siegel DA, Mitarai S, Coelho N, Serrao EA. Isolation by oceanographic distance explains genetic structure for Macrocystis pyrifera in the Santa Barbara Channel. Molecular Ecology. 2011;20:2543–2554. doi: 10.1111/j.1365-294X.2011.05117.x. [DOI] [PubMed] [Google Scholar]
- Almany et al. (2009).Almany GR, Connolly SR, Heath DD, Hogan JD, Jones GP, McCook LJ, Mills M, Pressey RL, Williamson DH. Connectivity, biodiversity conservation and the design of marine reserve networks for coral reefs. Coral Reefs. 2009;28:339–351. doi: 10.1007/s00338-009-0484-x. [DOI] [Google Scholar]
- Almany et al. (2013).Almany GR, Hamilton RJ, Bode M, Matawai M, Potuku T, Saenz-Agudelo P, Planes S, Berumen ML, Rhodes KL, Thorrold SR, Russ GR, Jones GP. Dispersal of grouper larvae drives local resource sharing in a coral reef fishery. Current Biology. 2013;23:626–630. doi: 10.1016/j.cub.2013.03.006. [DOI] [PubMed] [Google Scholar]
- Andrello et al. (2013).Andrello M, Mouillot D, Beuvier J, Albouy C, Thuiller W, Manel S. Low connectivity between mediterranean marine protected areas: a biophysical modeling approach for the dusky grouper Epinephelus marginatus. PLoS ONE. 2013;8:e511. doi: 10.1371/journal.pone.0068564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anonymous (2012).Anonymous. Diario Oficial de la Federacion Viernes 24 de agosto de 2012. Mexico, D.F.: Secretaria de Agricultura, Ganaderia, Desarrollo Rural, Pesca y Alimentacion; 2012. Carta nacional pesquera; pp. 1–235. Available at http://www.inapesca.gob.mx . [Google Scholar]
- Argote et al. (1995).Argote ML, Amador A, Lavin MF, Hunter JR. Tidal dissipation and stratification in the Gulf of California. Journal of Geophysical Research. 1995;100:16103–16118. doi: 10.1029/95JC01500. [DOI] [Google Scholar]
- Backhaus (1985).Backhaus JO. A three-dimensional model for the simulation of the shelf sea dynamics. Deutsche Hydrographische Zeitschrift. 1985;38:165–187. doi: 10.1007/BF02328975. [DOI] [Google Scholar]
- Baums, Paris & Cherubin (2006).Baums IB, Paris CB, Cherubin LM. A bio-oceanographic filter to larval dispersal in a reef-building coral. Limnology and Oceanography. 2006;51:1969–1981. doi: 10.4319/lo.2006.51.5.1969. [DOI] [Google Scholar]
- Beerli & Palczewski (2010).Beerli P, Palczewski M. Unified framework to evaluate panmixia and migration direction among multiple sampling locations. Genetics. 2010;185:313–326. doi: 10.1534/genetics.109.112532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beger et al. (2010).Beger M, Linke S, Watts M, Game E, Treml E, Ball I, Possingham HP. Incorporating asymmetric connectivity into spatial decision making for conservation. Conservation Letters. 2010;3:359–368. doi: 10.1111/j.1755-263X.2010.00123.x. [DOI] [Google Scholar]
- Beger et al. (2014).Beger M, Selkoe KA, Treml EA, Barber PH, von der Heyden S, Crandall ED, Toonen RJ, Riginos C. Evolving coral reef conservation with genetic information. Bulletin of Marine Science. 2014;90:159–185. doi: 10.5343/bms.2012.1106. [DOI] [Google Scholar]
- Beier & Ripa (1999).Beier E, Ripa P. Seasonal gyres in the Northern Gulf of California. Journal of Physical Oceanography. 1999;29:305–311. doi: 10.1175/1520-0485(1999)029<0305:SGITNG>2.0.CO;2. [DOI] [Google Scholar]
- Benjamini & Hochberg (2000).Benjamini Y, Hochberg Y. On the adaptive control of the false discovery rate in multiple testing with independent statistics. Journal of Educational and Behavioral Statistics. 2000;25:60–83. doi: 10.3102/10769986025001060. [DOI] [Google Scholar]
- Bird et al. (2007).Bird CE, Holland BS, Bowen BW, Toonen RJ. Contrasting phylogeography in three endemic hawaiian limpets (Cellana spp.) with similar life histories. Molecular Ecology. 2007;16:3173–3186. doi: 10.1111/j.1365-294X.2007.03385.x. [DOI] [PubMed] [Google Scholar]
- Bird et al. (2011).Bird CE, Smouse PE, Karl SA, Toonen RJ. Detecting and measuring genetic differentiation. In: Koenemann S, Held C, Schubart C, editors. Crustacean issues: phylogeography and population genetics in crustacea. Boca Raton, FL: CRC Press; 2011. pp. 31–55. [Google Scholar]
- Bode, Burrage & Possingham (2008).Bode M, Burrage K, Possingham HP. Using complex network metrics to predict the persistence of metapopulations with asymmetric connectivity patterns. Ecological Modelling. 2008;214:201–209. doi: 10.1016/j.ecolmodel.2008.02.040. [DOI] [Google Scholar]
- Burgess et al. (2014).Burgess SC, Nickols KJ, Griesemer CD, Barnett LAK, Dedrick AG, Satterthwaite EV, Yamane L, Morgan SG, White JW, Botsford LW. Beyond connectivity: how empirical methods can quantify population persistence to improve marine protected area design. Ecological Applications. 2014;24:257–270. doi: 10.1890/13-0710.1. [DOI] [PubMed] [Google Scholar]
- Buston et al. (2012).Buston PM, Jones GP, Planes S, Thorrold SR. Probability of successful larval dispersal declines fivefold over 1 km in a coral reef fish. Proceedings of the Royal Society B: Biological Sciences. 2012;279:1883–1888. doi: 10.1098/rspb.2011.2041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cannon (1966).Cannon R. The Sea of Cortes. Menlo Park, CA: Lane Magazine and Book Co.; 1966. p. 283. [Google Scholar]
- Carrillo & Palacios-Hernández (2002).Carrillo L, Palacios-Hernández E. Seasonal evolution of the geostrophic circulation in the Northern Gulf of California. Estuarine, Coastal and Shelf Science. 2002;54:157–173. doi: 10.1006/ecss.2001.0845. [DOI] [Google Scholar]
- Colin, Sadovy & Domeier (2003).Colin PL, Sadovy YJ, Domeier ML. Manual for the study and conservation of reef fish spawning aggregations. Society for the Conservation of Reef Fish Aggregations, Special Publication. 2003;1:1–99. [Google Scholar]
- Costello et al. (2012).Costello C, Ovando D, Hilborn R, Gaines SD, Deschenes O, Lester SE. Status and solutions for the world’s unassessed fisheries. Science. 2012;338:517–520. doi: 10.1126/science.1223389. [DOI] [PubMed] [Google Scholar]
- Cowen (2002).Cowen RK. Larval dispersal and retention and consequences for population connectivity. In: Sale PF, editor. Ecology of coral reef fishes: recent advances. San Diego, CA: Academic Press; 2002. pp. 149–170. [Google Scholar]
- Cowen et al. (2000).Cowen RK, Lwiza KMM, Sponaugle S, Paris BC, Olson DB. Connectivity of marine populations: open or closed? Science. 2000;287:857–859. doi: 10.1126/science.287.5454.857. [DOI] [PubMed] [Google Scholar]
- Cowen & Sponaugle (2009).Cowen RK, Sponaugle S. Larval dispersal and marine population connectivity. Annual Review of Marine Science. 2009;1:443–466. doi: 10.1146/annurev.marine.010908.163757. [DOI] [PubMed] [Google Scholar]
- Craig, Sadovy de Mitcheson & Heemstra (2011).Craig MT, Sadovy de Mitcheson YJ, Heemstra PC. Groupers of the world: a field and market guide. Grahamstown: NISC Ltd; 2011. p. 356. [Google Scholar]
- Craig & Sadovy (2008).Craig MT, Sadovy Y. 2008. Mycteroperca rosacea. IUCN 2012 IUCN Red List of Threatened Species Version 20122. Available at www.iucnredlist.org (accessed 16 June 2013)
- Crandall, Treml & Barber (2012).Crandall ED, Treml EA, Barber PH. Coalescent and biophysical models of stepping-stone gene flow in neritid snails. Molecular Ecology. 2012;21:5579–5598. doi: 10.1111/mec.12031. [DOI] [PubMed] [Google Scholar]
- Cudney-Bueno et al. (2009).Cudney-Bueno R, Bourillón L, Sáenz-Arroyo A, Torre-Cosío J, Turk-Boyer P, Shaw WW. Governance and effects of marine reserves in the Gulf of California, Mexico. Ocean & Coastal Management. 2009;52:207–218. doi: 10.1016/j.ocecoaman.2008.12.005. [DOI] [Google Scholar]
- Danell-Jiménez et al. (2009).Danell-Jiménez A, Sánchez-Velasco L, Lavín MF, Marinone SG. Three-dimensional distribution of larval fish assemblages across a surface thermal/chlorophyll front in a semienclosed sea. Estuarine, Coastal and Shelf Science. 2009;85:487–496. doi: 10.1016/j.ecss.2009.09.010. [DOI] [Google Scholar]
- Diaz-Uribe, Elorduy-Garay & Gonzalez-Valdovinos (2001).Diaz-Uribe JG, Elorduy-Garay JF, Gonzalez-Valdovinos MT. Age and growth of the leopard grouper, Mycteroperca rosacea, in the Southern Gulf of California, Mexico. Pacific Science. 2001;55:171–182. doi: 10.1353/psc.2001.0012. [DOI] [Google Scholar]
- Dyer (2009).Dyer RJ. Geneticstudio: a suite of programs for spatial analysis of genetic-marker data. Molecular Ecology Resources. 2009;9:110–113. doi: 10.1111/j.1755-0998.2008.02384.x. [DOI] [PubMed] [Google Scholar]
- Dyer, Nason & Garrick (2010).Dyer RJ, Nason JD, Garrick RC. Landscape modelling of gene flow: improved power using conditional genetic distance derived from the topology of population networks. Molecular Ecology. 2010;19:3746–3759. doi: 10.1111/j.1365-294X.2010.04748.x. [DOI] [PubMed] [Google Scholar]
- Erisman, Buckhorn & Hastings (2007).Erisman BE, Buckhorn ML, Hastings PA. Spawning patterns in the leopard grouper, Mycteroperca rosacea, in comparison with other aggregating groupers. Marine Biology. 2007;151:1849–1861. doi: 10.1007/s00227-007-0623-2. [DOI] [Google Scholar]
- Erisman, Rosales-Casián & Hastings (2007).Erisman BE, Rosales-Casián JA, Hastings PA. Evidence of gonochorism in a grouper, Mycteroperca rosacea, from the Gulf of California, Mexico. Environmental Biology of Fishes. 2007;82:23–33. doi: 10.1007/s10641-007-9246-1. [DOI] [Google Scholar]
- Excoffier, Laval & Schneider (2005).Excoffier L, Laval G, Schneider S. Arlequin (version 3.0): an integrated software package for population genetics data analysis. Evolutionary Bioinformatics. 2005;1:47–50. [PMC free article] [PubMed] [Google Scholar]
- Feutry et al. (2013).Feutry P, Vergnes A, Broderick D, Lambourdiere J, Keith P, Ovenden JR. Stretched to the limit; can a short pelagic larval duration connect adult populations of an indo-pacific diadromous fish (Kuhlia rupestris)? Molecular Ecology. 2013;22:1518–1530. doi: 10.1111/mec.12192. [DOI] [PubMed] [Google Scholar]
- Foster et al. (2012).Foster NL, Paris CB, Kool JT, Baums IB, Stevens JR, Sanchez JA, Bastidas C, Agudelo C, Bush P, Day O, Ferrari R, Gonzalez P, Gore S, Guppy R, McCartney MA, McCoy C, Mendes J, Srinivasan A, Steiner S, Vermeij MJA, Weil E, Mumby PJ. Connectivity of caribbean coral populations: complementary insights from empirical and modelled gene flow. Molecular Ecology. 2012;21:1143–1157. doi: 10.1111/j.1365-294X.2012.05455.x. [DOI] [PubMed] [Google Scholar]
- Gaggiotti (1996).Gaggiotti OE. Population genetic models of source–sink metapopulations. Theoretical Population Biology. 1996;50:178–208. doi: 10.1006/tpbi.1996.0028. [DOI] [PubMed] [Google Scholar]
- Gaines, Gaylord & Largier (2003).Gaines SD, Gaylord B, Largier JL. Avoiding current oversights in marine reserve design. Ecological Applications. 2003;13:S32–S36. doi: 10.1890/1051-0761(2003)013[0032:ACOIMR]2.0.CO;2. [DOI] [Google Scholar]
- Gaines et al. (2010).Gaines SD, White C, Carr MH, Palumbi SR. Designing marine reserve networks for both conservation and fisheries management. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:18286–18293. doi: 10.1073/pnas.0906473107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galindo et al. (2010).Galindo HM, Pfeiffer-Herbert AS, McManus MA, Chao Y, Chai F, Palumbi SR. Seascape genetics along a steep cline: using genetic patterns to test predictions of marine larval dispersal. Molecular Ecology. 2010;19:3692–3707. doi: 10.1111/j.1365-294X.2010.04694.x. [DOI] [PubMed] [Google Scholar]
- Gilg & Hilbish (2003).Gilg MR, Hilbish TJ. The geography of marine larval dispersal: coupling genetics with fine-scale physical oceanography. Ecology. 2003;84:2989–2998. doi: 10.1890/02-0498. [DOI] [Google Scholar]
- Gracia-Lopez et al. (2005).Gracia-Lopez V, Kiewek-Martinez M, Maldonado-Garcia M, Monsalvo-Spencer P, Portillo-Clark G, Civera-Cerecedo R, Linares-Aranda M, Robles-Mungaray M, Mazon-Suastegui JM. Larvae and juvenile production of the leopard grouper, Mycteroperca rosacea (streets, 1877) Aquaculture Research. 2005;36:110–112. doi: 10.1111/j.1365-2109.2004.01186.x. [DOI] [Google Scholar]
- Harrison et al. (2012).Harrison HB, Williamson DH, Evans RD, Almany GR, Thorrold SR, Russ GR, Feldheim KA, van Herwerden L, Planes S, Srinivasan M, Berumen ML, Jones GP. Larval export from marine reserves and the recruitment benefit for fish and fisheries. Current Biology. 2012;22:1023–1028. doi: 10.1016/j.cub.2012.04.008. [DOI] [PubMed] [Google Scholar]
- Hastings & Botsford (2006).Hastings A, Botsford LW. Persistence of spatial populations depends on returning home. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:6067–6072. doi: 10.1073/pnas.0506651103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hastings, Findley & Van der heiden (2010).Hastings PA, Findley LT, Van der heiden AMV. Fishes of the Gulf of California. In: Brusca RC, editor. The Gulf of California biodiversity and conservation. Tucson, AZ: The University of Arizona Press; 2010. pp. 96–118. [Google Scholar]
- Hernandez-Ayon et al. (2013).Hernandez-Ayon JM, Chapa-Balcorta C, Delgadillo-Hinojosa F, Camacho-Ibar VF, Huerta-Diaz MA, Santamaria-del-Angel E, Galindo-Bect MS, Segovia-Zavala JA. Dynamics of disolved inorganic carbon in the Midriff Islands region of the Gulf of California: influence of water masses. Ciencias Marinas. 2013;29:183–201. doi: 10.7773/cm.v39i2.2243. [DOI] [Google Scholar]
- Humason (1972).Humason GL. Animal tissue techniques. 3rd edition. San Francisco: WH Freeman and Company; 1972. [Google Scholar]
- Iacchei et al. (2013).Iacchei M, Ben-Horin T, Selkoe KA, Bird CE, Garcia-Rodriguez FJ, Toonen RJ. Combined analyses of kinship and FST suggest potential drivers of chaotic genetic patchiness in high gene-flow populations. Molecular Ecology. 2013;22:3476–3494. doi: 10.1111/mec.12341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen, Bohonak & Kelley (2005).Jensen JL, Bohonak AJ, Kelley ST. Isolation by distance, web service. BMC Genetics. 2005;6:13. doi: 10.1186/1471-2156-6-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keyghobadi et al. (2005).Keyghobadi N, Roland J, Matter SF, Strobeck C. Among- and within-patch components of genetic diversity respond at different rates to habitat fragmentation: an empirical demonstration. Proceedings of the Royal Society B: Biological Sciences. 2005;272:553–560. doi: 10.1098/rspb.2004.2976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kool et al. (2011).Kool JT, Paris CB, Barber PH, Cowen RK. Connectivity and the development of population genetic structure in Indo-West Pacific coral reef communities. Global Ecology and Biogeography. 2011;20:695–706. doi: 10.1111/j.1466-8238.2010.00637.x. [DOI] [Google Scholar]
- Kough, Paris & Butler (2014).Kough AS, Paris CB, Butler MJ. Larval connectivity and the international management of fisheries. PLoS ONE. 2014;8:e511. doi: 10.1371/journal.pone.0064970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Largier (2003).Largier JL. Considerations in estimating larval dispersal distances from oceanographic data. Ecological Applications. 2003;13:S71–S89. doi: 10.1890/1051-0761(2003)013[0071:CIELDD]2.0.CO;2. [DOI] [Google Scholar]
- Lavin et al. (1997).Lavin MF, Durazo R, Palacios E, Argote ML, Carrillo L. Lagrangian observations of the circulation in the Northern Gulf of California. Journal of Physical Oceanography. 1997;27:2298–2305. doi: 10.1175/1520-0485(1997)027<2298:LOOTCI>2.0.CO;2. [DOI] [Google Scholar]
- Lavin & Marinone (2003).Lavin M, Marinone SG. An overview of the physical oceanography of the Gulf of California. In: Velasco-Fuentes OU, Sheinbaum J, Ochoa de la Torre JL, editors. Nonlinear processes in geophysical fluid dynamics. Dordrecht: Kluwer Academic Publishers; 2003. pp. 173–204. [Google Scholar]
- Leis (2007).Leis JM. Behaviour as input for modelling dispersal of fish larvae: behaviour, biogeography, hydrodynamics, ontogeny, physiology and phylogeny meet hydrography. Marine Ecology Progress Series. 2007;347:185–193. doi: 10.3354/meps06977. [DOI] [Google Scholar]
- Librado & Rozas (2009).Librado P, Rozas J. Dnasp v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics. 2009;25:1451–1452. doi: 10.1093/bioinformatics/btp187. [DOI] [PubMed] [Google Scholar]
- Marinone (2003).Marinone SG. A three-dimensional model of the mean and seasonal circulation of the Gulf of California. Journal of Geophysical Research. 2003;108:1–27. doi: 10.1029/2002JC001720. [DOI] [Google Scholar]
- Marinone (2008).Marinone SG. On the three-dimensional numerical modeling of the deep circulation around Angel de la Guarda island in the Gulf of California. Estuarine, Coastal and Shelf Science. 2008;80:430–434. doi: 10.1016/j.ecss.2008.09.002. [DOI] [Google Scholar]
- Marinone (2012).Marinone SG. Seasonal surface connectivity in the Gulf of California. Estuarine, Coastal and Shelf Science. 2012;100:133–141. doi: 10.1016/j.ecss.2012.01.003. [DOI] [Google Scholar]
- Meirmans & Hedrick (2011).Meirmans PG, Hedrick PW. Assessing population structure: FST and related measures. Molecular Ecology Resources. 2011;11:5–18. doi: 10.1111/j.1755-0998.2010.02927.x. [DOI] [PubMed] [Google Scholar]
- Moreno-Báez et al. (2012).Moreno-Báez M, Cudney-Bueno R, Orr BJ, Shaw WW, Pfister T, Torre-Cosio J, Loaiza R, Rojo M. Integrating the spatial and temporal dimensions of fishing activities for management in the Northern Gulf of California, Mexico. Ocean & Coastal Management. 2012;55:111–127. doi: 10.1016/j.ocecoaman.2011.10.001. [DOI] [Google Scholar]
- Moreno-Baez et al. (2010).Moreno-Baez M, Orr BJ, Cudney-Bueno R, Shaw WW. Using fishers local knowledge to aid management at regional scales: spatial distribution of small-scale fisheries in the Northern Gulf of California, Mexico. Bulletin of Marine Science. 2010;86:339–353. [Google Scholar]
- Newman (2003).Newman MEJ. The structure and function of complex networks. SIAM Review. 2003;45:167–256. doi: 10.1137/S003614450342480. [DOI] [Google Scholar]
- Paden & Abbott (1991).Paden CA, Abbott MR. Tidal and atmospheric forcing of the upper ocean in the Gulf of California. 1. Sea surface temperature variability. Journal of Geophysical Research. 1991;96:18337–18359. doi: 10.1029/91JC01597. [DOI] [Google Scholar]
- Palacios-Hernandez et al. (2002).Palacios-Hernandez E, Beier E, Lavin M, Ripa P. The effect of the seasonal variation of stratification on the circulation of the Northern Gulf of California. Journal of Physical Oceanography. 2002;32:705–728. doi: 10.1175/1520-0485(2002)032<0705:TEOTSV>2.0.CO;2. [DOI] [Google Scholar]
- Palumbi et al. (1991).Palumbi SR, Martin A, Romano S, McMillan WO, Stice L, Grabowski G. The simple fool’s guide to PCR. Version 2.0. Honolulu, HI: 1991. Privately published document. [Google Scholar]
- Petit, El Mousadik & Pons (1998).Petit RJ, El Mousadik A, Pons O. Identifying populations for conservation on the basis of genetic markers. Conservation Biology. 1998;12:844–855. doi: 10.1046/j.1523-1739.1998.96489.x. [DOI] [Google Scholar]
- Petitgas et al. (2012).Petitgas P, Alheit J, Peck MA, Raab K, Irigoien X, Huret M, van der Kooij J, Pohlmann T, Wagner C, Zarraonaindia I, Dickey-Collas M. Anchovy population expansion in the North sea. Marine Ecology Progress Series. 2012;444:1–13. doi: 10.3354/meps09451. [DOI] [Google Scholar]
- Pons & Petit (1996).Pons O, Petit RJ. Measuring and testing genetic differentiation with ordered versus unordered alleles. Genetics. 1996;144:1237–1245. doi: 10.1093/genetics/144.3.1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Proehl et al. (2005).Proehl JA, Lynch DR, McGillicuddy DJ, Ledwell JR. Modeling turbulent dispersion on the North flank of Georges Bank using lagrangian particle methods. Continental Shelf Research. 2005;25:875–900. doi: 10.1016/j.csr.2004.09.022. [DOI] [Google Scholar]
- R Core Team (2013).R Core Team . R: a language and environment for statistical computing. Vienna: R foundation for statistical computing; 2013. Available at http://www.R-project.Org/ [Google Scholar]
- Robertson & Cramer (2009).Robertson DR, Cramer KL. Shore fishes and biogeographic subdivisions of the tropical eastern pacific. Marine Ecology Progress Series. 2009;380:1–17. doi: 10.3354/meps07925. [DOI] [Google Scholar]
- Sala et al. (2003).Sala E, Aburto-Oropeza O, Paredes G, Thompson G. Spawning aggregations and reproductive behavior of reef fishes in the Gulf of California. Bulletin of Marine Science. 2003;72:103–121. [Google Scholar]
- Sala et al. (2004).Sala E, Aburto-Oropeza O, Reza M, Paredes G, López-Lemus LG. Fishing down coastal food webs in the Gulf of California. Fisheries. 2004;29:19–25. doi: 10.1577/1548-8446(2004)29[19:FDCFWI]2.0.CO;2. [DOI] [Google Scholar]
- Sale et al. (2005).Sale P, Cowen R, Danilowicz B, Jones G, Kritzer J, Lindeman K, Planes S, Polunin N, Russ G, Sadovy Y, Steneck R. Critical science gaps impede use of no-take fishery reserves. Trends in Ecology & Evolution. 2005;20:74–80. doi: 10.1016/j.tree.2004.11.007. [DOI] [PubMed] [Google Scholar]
- Sambrook, Fritsch & Maniatis (1989).Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. Second edition. Vols. 1, 2, and 3. New York: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Selkoe et al. (2010).Selkoe KA, Watson JR, White C, Horin TB, Iacchei M, Mitarai S, Siegel DA, Gaines SD, Toonen RJ. Taking the chaos out of genetic patchiness: seascape genetics reveals ecological and oceanographic drivers of genetic patterns in three temperate reef species. Molecular Ecology. 2010;19:3708–3726. doi: 10.1111/j.1365-294X.2010.04658.x. [DOI] [PubMed] [Google Scholar]
- Smith et al. (2010).Smith M, Milic-Frayling N, Shneiderman B, Mendes-Rodriguez E, Leskovec J, Dunne C. 2010. Nodexl: a free and open network overview, discovery and exploration add-in for Excel 2007/2010. Available at http://nodexlcodeplex.com .
- Soria et al. (2012).Soria G, Munguía-Vega A, Marinone SG, Moreno-Báez M, Martínez-Tovar I, Cudney-Bueno R. Linking bio-oceanography and population genetics to assess larval connectivity. Marine Ecology Progress Series. 2012;463:159–175. doi: 10.3354/meps09866. [DOI] [Google Scholar]
- Soria et al. (2014).Soria G, Torre-Cosio J, Munguia-Vega A, Marinone SG, Lavín MF, Cinti A, Moreno-Báez M. Dynamic connectivity patterns from an insular marine protected area in the Gulf of California. Journal of Marine Systems. 2014;129:248–258. doi: 10.1016/j.jmarsys.2013.06.012. [DOI] [Google Scholar]
- Thomson, Findley & Kerstitch (2000).Thomson DA, Findley LT, Kerstitch AN. Reef fishes of the sea of Cortez. Austin, TX: University of Texas Press; 2000. [Google Scholar]
- Toonen et al. (2011).Toonen RJ, Andrews KR, Baums IB, Bird CE, Concepcion GT, Daly-Engel TS, Eble JA, Faucci A, Gaither MR, Iacchei M, Puritz JB, Schultz JK, Skillings DJ, Timmers MA, Bowen BW. Defining boundaries for ecosystem-based management: a multispecies case study of marine connectivity across the Hawaiian archipelago. Journal of Marine Biology. 2011;2011:1–13. doi: 10.1155/2011/460173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toonen et al. (2013).Toonen RJ, Wilhelm TA, Maxwell SM, Wagner D, Bowen BW, Sheppard CRC, Taei SM, Teroroko T, Moffitt R, Gaymer CF, Morgan L, Lewis N, Sheppard ALS, Parks J, Friedlander AM. One size does not fit all: the emerging frontier in large-scale marine conservation. Marine Pollution Bulletin. 2013;77:7–10. doi: 10.1016/j.marpolbul.2013.10.039. [DOI] [PubMed] [Google Scholar]
- Treml et al. (2008).Treml EA, Halpin PN, Urban DL, Pratson LF. Modeling population connectivity by ocean currents, a graph-theoretic approach for marine conservation. Landscape Ecology. 2008;23:19–36. doi: 10.1007/s10980-007-9138-y. [DOI] [Google Scholar]
- Treml et al. (2012).Treml EA, Roberts JJ, Chao Y, Halpin PN, Possingham HP, Riginos C. Reproductive output and duration of the pelagic larval stage determine seascape-wide connectivity of marine populations. Integrative and Comparative Biology. 2012;52:525–537. doi: 10.1093/icb/ics101. [DOI] [PubMed] [Google Scholar]
- Visser (1997).Visser AW. Using random walk models to simulate the vertical distribution of particles in a turbulent water column. Marine Ecology Progress Series. 1997;158:275–281. doi: 10.3354/meps158275. [DOI] [Google Scholar]
- Vuilleumier, Bolker & Leveque (2010).Vuilleumier S, Bolker BM, Leveque O. Effects of colonization asymmetries on metapopulation persistence. Theoretical Population Biology. 2010;78:225–238. doi: 10.1016/j.tpb.2010.06.007. [DOI] [PubMed] [Google Scholar]
- Wallace & Selman (1981).Wallace RA, Selman K. Cellular and dynamic aspects of oocyte growth in teleosts. American Zoology. 1981;21:325–343. [Google Scholar]
- Watson et al. (2010).Watson JR, Mitarai S, Siegel DA, Caselle JE, Dong C, McWilliams JC. Realized and potential larval connectivity in the Southern California Bight. Marine Ecology Progress Series. 2010;401:31–48. doi: 10.3354/meps08376. [DOI] [Google Scholar]
- White et al. (2010a).White JW, Botsford LW, Hastings A, Largier JL. Population persistence in marine reserve networks: incorporating spatial heterogeneities in larval dispersal. Marine Ecology Progress Series. 2010a;398:49–67. doi: 10.3354/meps08327. [DOI] [Google Scholar]
- White et al. (2010b).White C, Selkoe KA, Watson J, Siegel DA, Zacherl DC, Toonen RJ. Ocean currents help explain population genetic structure. Proceedings of the Royal Society B: Biological Sciences. 2010b;277:1685–1694. doi: 10.1098/rspb.2009.2214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodson & McManus (2007).Woodson CB, McManus MA. Foraging behavior can influence dispersal of marine organisms. Limnology and Oceanography. 2007;52:2701–2709. doi: 10.4319/lo.2007.52.6.2701. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The map shows 17 sites where virtual larvae were released (colored polygons) and 11 sites where genetic samples were also collected (blue polygons).
Tides in the HAMSOM oceanographic model for the northern Gulf of California, showing the four neap and four spring tides during May and June selected for the release of virtual larvae. Red lines separate distinct months.
Pairwise maximum FST possible while maintaining the within-population diversity above diagonal and pairwise values below diagonal. For each site, we show their membership to one of three geographic groups.
Mantel and partial Mantel tests between an empirical matrix of genetic structure (FST) values and various explanatory variables, including geographic distance and three distinct matrices calculated from the modeled networks, including adjacency, graph distance and log graph distance. P values below 0.05 are shown in bold. * Indicate statistical significance after correcting for multiple tests (critical P = 0.0060).
Mantel and partial Mantel tests between an empirical matrix of genetic structure () values and various explanatory variables, including geographic distance and three distinct matrices calculated from the modeled networks, including adjacency, graph distance and log graph distance. P values below 0.05 are shown in bold. * Indicate statistical significance after correcting for multiple tests (critical P = 0.0060).
Analysis of molecular variance testing for regional genetic subdivision when sampling sites were clustered into the Baja Peninsula, Midriff Islands and Sonoran coast groups.
Maps showing the spatial distribution of larvae released at each of eight release dates corresponding to four neap and four spring tides during May and June at each of 17 sites (“X” in each box) for PLD 14, 21 and 28 days. Scale in kilometers.
Detailed results from three independent runs with Migrate-n software for unrestricted gene flow among Baja, the Midriff Islands and Sonora.
Detailed results from three independent runs with Migrate-n software for a model of gene flow from Baja across the Midriff Islands towards Sonora.
Detailed results from three independent runs with Migrate-n software for a model of gene flow from Sonora across the Midriff Islands towards Baja.