

Abstract
Klebsiella pneumoniae (KP) with the hypermucoviscosity (HV) phenotype has abundant capsular polysaccharides (CPS) and usually causes an invasive syndrome. Sialic acid (Sia), a component of CPS in KP strains with the HV phenotype, may be anti-phagocytic. Sia-binding immunoglobulin-like lectin-9 (Siglec-9) act as an MHC class-I receptor on neutrophils that recognizes Sia and sends a signal to dampen inflammatory response. Three clinical KP strains with KP-M1 (HV-positive; capsular serotype K1), KP-14 (HV-negative; capsular serotype non-K1/K2), and DT-X (HV-negative; capsular serotype K1) were studied. We assessed total Sia in CPS extracts using enzymatic methods and phagocytosis by neutrophils of neuraminidase-treated bacteria using flow cytometry. Neutrophil killing was evaluated in the presence and absence of antibodies against Siglec-9. The concentration of Sia was significantly higher in the CPS extract of KP-M1 (56.75 ± 6.75 μmole/109 cfu) than in the CPS extract of KP-14 (0.02 ± 0.01 μmole/109 cfu) and DT-X (a negligible value). The KP-M1 (compared with the KP-14 and DT-X) was more resistant to neutrophil phagocytosis. Both the HV phenotype and resistance to phagocytosis of KP-M1 were significantly decreased after Sia removal with neuraminidase treatment. Fluorescence microscopy with an antibody against human Siglec-9 showed attachment of KP-M1 (but were absent of KP-14 and DT-X) to the surface of neutrophils and colocalization with human Siglec-9. Engagement of Siglec-9 via Sia enhanced neutrophils killing of KP-M1 by ex vivo human neutrophils bactericidal activity assay. The result showed that Sia might be a constituent of KP-M1 CPS responsible for HV, thereby contributing to anti-phagocytic activity of this pathogen.
Keywords: sialic acid, K. pneumoniae, hypermucoviscosity, neutrophil, phagocytosis
Introduction
Klebsiella pneumoniae (KP) is the leading cause of primary liver abscesses (PLA) in Taiwan,1 and worldwide.2 Several bacterial virulence factors, including capsular polysaccharide (CPS), have been identified as determinants of KP infection.3 This capsule consists of high molecularweight polysaccharides and contributes to the mucoid phenotype. CPS may help the bacteria evade phagocytosis and impede bacterial clearance from the host.4 Several CPS-associated characteristics have been shown to correlate with the occurrence of PLA, including serotype K1 or K22,3 and a muco-polysaccharide web outside the capsule, also known as the hypermucoviscosity (HV) phenotype.5 The composition of CPS from KP can be determined by enzyme treatment and lectin binding assay,6 and the presence of terminal sialic acid (Sia) and fucose in CPS is accessible by fucosidase after removal of Sia by neuraminidase (NANase).6
In microbial infections, neutrophils are major elements of the innate immune defense against invasive bacterial pathogen.7 The ability of the HV phenotype to resist phagocytosis by neutrophils in humans has been recognized,8,9 but the mechanism underlying this ability is unknown. We hypothesized that the presence of Sia in KP CPS dampens the innate immune response of the infected host. Cell surfaces in the immune system are richly equipped with glycan-binding protein, which could recognize glycans. Sia-binding immunoglobulin-like lectin (Siglec), which are structurally related type I trans-membrane proteins binding to Sia-containing carbohydrate structure (sialoglycans), are broadly expressed on immune cells.10 Siglec-9 is predominantly expressed on the surface of neutrophils that recognizes Sia and transduces inhibitory signals to modulate the inflammatory responses.11 Sialylation of glycoconjugate in microbes seems to be crucial for their survival in the phagocytes, possibly reducing in the neutrophil oxidative brust, diminishing formation of neutrophil extracellular DNA traps, then increasing pathogens survival through Siglecs.12
To elucidate the pathogenesis of infection by KP with the HV phenotype, we investigated whether and how CPS Sia affects phagocytosis by neutrophils.
Results
Quantification of capsular polysaccharide (CPS) and sialic acid (Sia)
The quantity of CPS (in μg/109 cfu) was 24.8 ± 1.8, 0.13 ± 0.03, and negligible on KP-M1, KP-14, and DT-X, respectively. Total Sia (μmole/109 cfu) in CPS extracts from KP-M1 and KP-14 was 56.75 ± 6.75 and 0.02 ± 0.01, respectively. The Sia level was significantly higher in the CPS of KP-M1 than that of KP-14 (P < 0.01).
Differential effects of heat and UV on the mucoviscosity of K. pneumoniae
The capsule of KP-14 was thin without obvious mucoviscosity. It was easily pelleted by centrifugation. In contrast, the capsular ring of KP-M1 was thick with high mucoviscosity and could not be pelleted by centrifugation (Fig. 1). Heat treatment (70 °C for 60 min or 95 °C for 30 min) killed the KP organisms and reduced their mucoviscosity (string length), while UV treatment for 5 min killed the KP organisms but had little effect on the mucoviscosity of KP-M1, suggesting that UV treatment causes lethal damage without affecting CPS structure. This suggestion was also supported by the observation that the capsule of KP was disrupted by heat but not by UV treatment (Fig. 1). We therefore use a 5 min UV treatment for bacterial inactivation in subsequent experiments.

Figure 1. The hypermucoviscosity phenotype and capsular ring of K. pneumoniae susceptible to heat but not UV.
NANase effect on the hypermucoviscosity of KP-M1
As shown in Figure 2A and B, treatment with 20 or 100 mU/mL NANase significantly decreased the HV of KP-M1 (P < 0.01) but had no effect on the growth of the three KP strains 24 h after treatment (data not shown).

Figure 2. Positive string test for hypermucoviscosity in K. pneumoniae. (A) Each closed bar indicatesthe string-test mean length ± standard deviation (mm) of neuraminidase (NANase)-treated KP-M1. (B) Representative photographs of stretched, mucoviscous strings from a single colony of untreated KP-M1 (medium) or NANase (100 mU/mL)-treated KP-M1.
Phagocytosis of K. pneumoniae with and without HV phenotype by human neutrophils
The HV-positive KP-M1 strain (compared with the HV-negative KP-14 strain and HV-negative DT-X strain) was more resistant to neutrophil phagocytosis (P < 0.01; Fig. 3A). The percentage of phagocytosis against KP-M1 increased significantly after treatment with 20 or 100 mU/mL NANase (P < 0.05; Fig. 3B). However, the difference of the percentage of KP-14 phagocytosed between before and after treatment with 20 or 100 mU/mL NANase was not significant (Fig. 3B). No significant difference in the level of phagocytosis of the KP-M1 and KP-14 strain remained after Sia removal with 100 mU/mL NANase treatment, which compared with the level of phagocytosis of the DT-X strain (Fig. 3B).
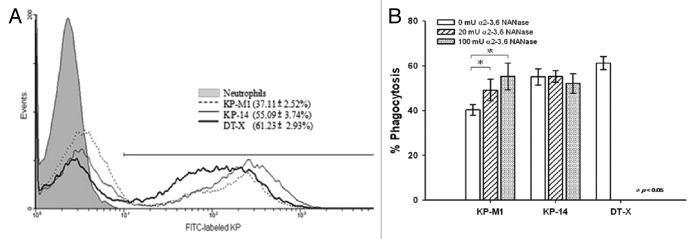
Figure 3. Flow cytometric measurement of neutrophil phagocytosis. Trypan blue quenching of the extracellular fluorescence of bound bacteria, and uptake of bacteria by neutrophils after incubation at 37 °C for 60 min was evaluated by flow cytometry. (A) KP-M1 was significantly more resistant to phagocytosis by neutrophils than either KP-14 or DT-X. (B) The amount of bacterial uptake of KP-M1 was significantly different between before and after treatment with neuraminidase (NANase 20 or 100 mU/mL; P < 0.05), but not for KP-14. There was no significant difference in the level of phagocytosis of KP-M1 and KP-14 strain after 100 mU/mL NANase treatment, which compared with the level of phagocytosis of DT-X strain.
Interaction of K. pneumoniae with hSiglec-9 present on the surface of neutrophils
Neutrophil–KP interactions were detected with a hSiglec-9 specific Fab antibody fragment (a fluorescein-tagged secondary antibody) under a fluorescence microscope. On the surface of human neutrophils, punctate staining of hSiglec-9 was observed in the presence or absence of bacteria. When interacted with the neutrophil surface, KP-M1 (yellow) (Fig. 4A) but neither KP-14 (Fig. 4B) nor DT-X (Fig. 4C) exhibited clear foci of colocalized hSiglec-9 staining (red) and KP staining (green).
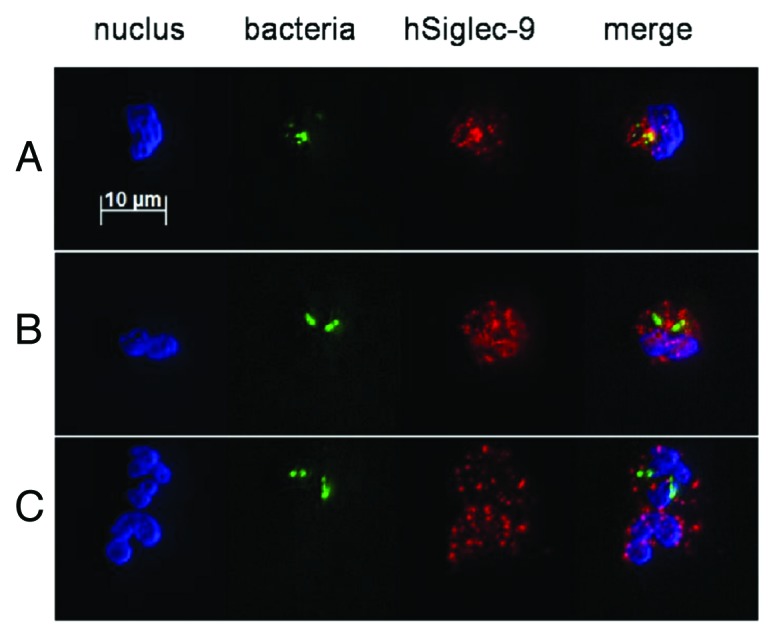
Figure 4.K. pneumoniae attachment to neutrophils after induction of hSiglec-9 colocalization. Human neutrophils were incubated (5 min) with FITC labeled K. pneumoniae, fixed, labeled with fluorescein-tagged antibodies to hSiglec-9, and mounted on slides. Three images show the nucleus (blue Hoechst stain), bacteria (green FITC), and hSiglec-9 (red phycoerythrin) separately, and one last image merges the previous three. The areas of colocalization between FITC labeled K. pneumoniae (green) and hSiglec-9 (red) appear yellow for (A) KP-M1 but are absent for (B) KP-14 and (C) DT-X. The experiment was repeated three times, and this image is representative.
Engagement of Siglec-9 inhibits human neutrophils bactericidal activity against KP-M1
Siglec-9 attachment to KP CPS sialoglycans dampens neutrophil activation in response to bacterial recognition. The bactericidal activity of neutrophils unable to engage sialylated KP via Siglec-9 (following incubation with Siglec-9 BSAb) was enhanced toward KP-M1 (P < 0.01; Fig. 5A) but not toward KP-14 (P = 0.29) or DT-X (P = 0.11) (Fig. 5B). Thus, a novel functional consequence of KP-M1 sialoglycan molecular mimicry interacts with neutrophil Siglec-9, preventing these cells from phagocytic killing, and thereby contributing to the pathogenicity of KP-M1. Indeed, neutrophils killed more KP-M1 following co-incubation with Siglec-9 BSAb than KP-M1 following co-incubation with Siglec-9 NBSAb (P < 0.01; Fig. 5A). Binding to KP-M1 was Sia-dependent and selectively blocked by the Siglec-9 BSAb, confirming the Siglec-9 dependency of KP-M1 anti-phagocytotic activity, as it did not occur with either KP-14 or DT-X.

Figure 5. Ex vivo human neutrophils bactericidal activity assay. Human neutrophils collected from 5 healthy male were incubated with anti-hSiglec neutralizing Ab. KP (1 × 107 cfu/mL) was incubated with these neutrophils (multiplicity of infection, 10; 37 °C; 60 min), and the surviving bacteria were counted the next day. Siglec-9 BSAb enhanced human neutrophils killing toward KP-M1 (A; P < 0.01) but not toward KP-14 (B; P = 0.29) or DT-X (B; P = 0.11).
Discussion
Sia is prominently found at the terminal position of vertebrate cell surface glycoconjugates and is involved in cell–cell recognition.13 Many bacteria incorporate Sia into cell surface features like CPS, which help them evade the innate immune response of the host. Pathogenic bacteria such as Escherichia coli K1, Hemophilus influenzae, and Streptococcus agalactiae have evolved ways to bypass host defenses including the use of this molecule as a shield.14 Although there were not any reference to the presence of Sia on capsule of serotype K1 KP, Di Martino et al. described the carbohydrate part receptor involved in a protein expressed by KP contained N-acetylneuraminic acid residue.15 The presence of Sia in the cell envelope of serotype K2 KP was reported under the Sia specific topo-optical analysis in a more recent study.16 A detailed structural analysis of the surface polysaccharide could confirm these results but this was outside the purview of this experiment. However, this is something worthy of further research.
The incorporation of Sia into pathogen cell-surface features (Sia-containing capsules) may allow pathogens to circumvent and/or counteract the host’s innate immune responses through a strategy of “molecular mimicry”.12,17 Previous findings of studies examining the phagocytosis of bacterial isolates suggest that the copious capsule (mucoidy phenotype) of KP is a virulence factor for infection.8,18 In the present study, the HV phenotype of KP (KP-M1) had significantly more resistance to neutrophil phagocytosis and killing than KP with the non-HV phenotype (KP-14 or DT-X). A prominent feature of KP-M1 CPS is Sia, suggesting its possible use in a molecular mimicry strategy to survive inside the host and cause invasive syndrome. The amount of Sia on the microbial cell surface is regulated by NANase, which cleaves sialyl residues from cell-surface glycoconjugates.19 According to our results, treatment of the HV phenotype with increasing doses of NANase decreased HV and increased the percentage of KP cells phagocytosed by neutrophils. However, string tests for HV is a semi-quantitative method and NANase treatment affects other surface structure; hence, the HV may not be the only thing effected after NANase treatment in these cells. These data still emphasize the role of Sia in HV and resistance to neutrophil phagocytosis.
Most surface Sia is linked to oligosaccharide chains by α-glycosidic bonds.20 The two most important saccharides, glucose and galactose, are frequently bound to Sia by α (2→3) and α (2→6) linkages.20 The Sia linked by α (2→3) and α (2→6) could be cleaved by NANase from Clostridium perfringens. The cleavage of galactose- and glucose-linked Sia from the surface of CPS may impair the functions or alter the structure of CPS. The depletion of Sia might decrease HV phenotype and enhance phagocytosis by neutrophils. However, NANase treatment did not completely reverse HV or neutrophil phagocytosis even at high doses (up to 100 mU/mL). Another approach might be to target bacterial sialyltransferase, measuring activity by binding competition or enzyme inhibition assays.
The present study demonstrated that KP-M1 could engage hSiglec-9, which expressed on the surface of human neutrophils. Siglec-9 recognizes α2→3 linked Sia and can participate in binding. BSAb, but not NBSAb, was recently reported to inhibit Siglec-9-Fc binding to Sia α2→3 galβ1–4GlcNAcβ1.12 In vitro studies have also disclosed Siglec to act as inhibitors, including to downregulate T-cell signaling21and inhibit NK cell toxicity.22 Therefore, we suppose that KP might has evolved the HV phenotype to suppress the innate immune response through engagementof Siglec-9 on neutrophils, and that evolutionary forces modulating hSiglec–KP interactions may constantly alter the host–pathogen relationship. Thus, the consequence of KP sialoglycan molecular mimicry is phagocytosis inhibition through interaction of KP sialoglycan with neutrophil Siglec-9, and thereby evasion of neutrophil killing of HV-KP. This can contribute to KP pathogenicity. However, our data are not sufficient to conclude that direct interaction between Siglec-9 and KP-M1. Further study to demonstrate the direct binding of HV-KP CPS to Siglec-9, by using recombinant Siglec-9 protein or a cell line that overexpresses Siglec-9 is warranted. Finally, only three clinical KP strains were assessed in this study. Epidemiological reports have indicated that capsular serotype K1 is the most prevalent in KP invasive syndrome.2,3,23 Therefore, our results derived from the representative serotype K1 strain might be extrapolated for the majority of KP strains that cause invasive syndrome.
Sia has another important function in many bacterial CPS to provide a virulence factor. High levels of Sia in CPS might promote the binding of Factor H of the alternative complement pathway thereby limiting the interaction between C3b and Factor B to allow completion of the terminal membrane attack complex.24 Two major mechanisms of resistance to the complement-mediated bactericidal activity in KP had been identified. The most frequently identified mechanism is expression of a lipo-polysaccharides O side chain, which prevents access of complement components to activators. In a minority of strains, resistance is due to coverage of the activators by the CPS. While some particular K serotypes (K1, K10, and K16) in KP have been involved in complement resistance had been reported.25 Comparing in the presence of unheated serum, our pilot study also showed poor neutrophils phagocytosis of KP-M1 (serotype K1) in the presence of heated serum. It might be explained by serum exerts its effect by opsonic function for phagocytosis on serotype K1 KP strain. The relationship between Sia expression in KP-M1 organisms and reduction of susceptibility to complement activity deserve further investigation.
In the present study, we used a depletion of Sia by sialidase and found Sia might contribute to the HV phenotype of serotype K1 KP, and thereby is associated with anti-phagocytic activity of this pathogen. The interaction of HV-positive KP with the host immune system by interrupting Sia-hSiglec-9 may provide valuable information of management of these infections by HV phenotype of KP.
Materials and Methods
Bacteria strains
HV-positive KP-M1 (capsular serotype K1) was isolated from a patient with PLA; HV-negative KP-14 (capsular serotype non-K1/K2) was isolated from a patient with urinary tract infection, and HV-negative DT-X was isolated by subculture of strain DT-S (biotype edwardsii, capsular serotype K1).26 The protocols for all experiments were approved by the Institutional Review Board, Chang Gung Memorial Hospital (document no. 97-0998B) and the need for informed consent was waived.
Quantification of capsular polysaccharides and sialic acid
Purified CPS was quantified by determining uronic acid levels in samples using a modified carbazole assay.27 The enzymatic method witha Sialic Acid Quantitation Kit (Sigma-Aldrich) was used to determine the total Sia in the CPS extracts.28 The CPSs extracted from these three KP strains were heated briefly (90 °C for 5 min) to prevent metabolism of the free Sia. The samples were then incubated with 1 μL of NANase (100 mU/mL Clostridium perfringensα-[2–3,6] sialidase) (Sialidase C/NANase II GK80030; ProZyme) overnight at 37 °C to release free Sia from the glycoconjugates. The free Sia (N-acetylneuraminic acid) was converted to pyruvic acid by N-acetylneuraminic acid aldolase and then treated with lactic dehydrogenase to form lactic acid with the oxidation of β-NADH to β-NAD. β-NADH oxidation can be accurately measured spectrophotometrically. Bovine fetuin (Sigma-Aldrich) was used as a reference glycoprotein. All samples were assayed at least three times. The quantification CPS and total Sia in the CPS extracts detected were expressed as μg and μmole per 109 colony-forming units (cfu), separately.
String test for hypermucoviscosity
The HV phenotype of the KP isolates was determined using a modified string test.29 The formation of mucoviscous string was assessed by measuring string length (mm). For string-test assays after enzymatic removal of Sia, bacteria were cultivated on trypticase soy broth (TSB) agar (Becton, Dickinson and Company) containing 20 or 100 mU/mL NANase for 2 h at 37 °C, and then the string length was measured.
Differential effects of heat and UV on the capsular hypermucosity of K. pneumoniae
Before incubation with neutrophils, the KP bacteria were inactivated by heat (95 °C for 30 min or 70 °C for 60 min) or UV (20 J/cm2 for 5 min or 15 min). The effects of heat and UV on the capsular hypermucosity were determined by the string test and negative staining for the capsular ring.
FITC labeling of K. pneumoniae
KP cells were grown to the early exponential phase, and then treated first with 20 or 100 mU/mL NANase or PBS for 2 h at 37 °C and then with UV for 5 min. The cells were washed in 50 mM carbonate buffer (pH 8.0), resuspended in a 5 mL carbonate buffer (pH 8.0) plus 0.1% fluorescein isothiocyanate (FITC), and incubated for 1 h at 37 °C. FITC-labeled K. pneumoniae (FITC-KP) (1 × 106 colony-forming units [cfu]/mL) were analyzed by flow cytometry (FACSCalibur; Becton, Dickinson and Company) to verify that bacterial staining was uniform throughout each sample.
Flow cytometric analysis of phagocytosis
Phagocytosis was measured using a standard assay.30 Briefly, 10 μL of FITC-KP (representing 5 × 108 cfu/mL) was added to each 990 μL volume containing a mixture of 100 μL of a neutrophil suspension (representing 5 × 106 cells/mL), 100 μL of pooled normal human serum (10% v/v for opsonization), and 790 μL of PBS. The final volume was 1.0 mL and the multiplicity of infection was 10:1. Each tube was incubated in a shaking water bath at 37 °C and transferred to an ice bath at a designated time point. A FACScan emitting an argon laser beam at 488 nm was used to detect FITC fluorescence. A total of 20 000 cells were processed using Cellquest version 1.0 software (Becton, Dickinson and Company). Green fluorescence (FL1-H) intensity data (collected using a logarithmic amplifier) were displayed as single histograms. By processing phagocytosis mixtures of unstained and FITC-stained bacteria, the boundary of positive and negative fluorescence was determined and the percentage of ingested bacteria was assessed.
Assessment of K. pneumoniae attachment to neutrophils by fluorescence microscopy
FITC-KP was added to human neutrophils in RPMI with 10% pooled serum at a ratio of 10:1. The neutrophils were fixed with 2% paraformaldehyde in PBS. Human neutrophils were labeled using mouse anti-human Siglec-9 (hSiglec-9) antibody Fab fragments. Anti-hSiglec-9 Fab fragments were prepared using the BD PharMingen mouse anti-hSiglec-9 clone E10-286 and the Pierce ImmunoPure IgG1 Fab and F(ab)2 preparation kit (Thermo Fisher Scientific). Neutrophils were washed and treated with goat anti-mousephycoerythrin antibody (4 °C for 30 min). Images were captured with an AxioVision digital imaging system (Carl Zeiss Micro Imaging).
Assay of bactericidal activity in the presence of anti-Siglec-9 antibody
Neutrophils collected from 5 healthy male volunteers were incubated for 5 min in the presence or absence of mouse anti-human Siglec-9 IgG monoclonal antibody directed against the Sia-binding site (BSAb; murine IgG2a clone 191240; R&D Systems) or not directed against the Sia-binding site (NBSAb; murine IgG1 clone E10–286; Becton, Dickinson and Company) at 10 μg/mL before use in the bactericidal assays. Bactericidal activity was measured using a standard assay method.31 The bacteria (1 × 107 cfu/mL) were opsonized with pooled human 10% serum collected from the same 5 healthy male and then added to human neutrophils either in the presence or absence of anti-Siglec-9 antibody at a multiplicity of infection of 10:1. Samples (50 μL) were withdrawn at 60 min and diluted with 2.45 mL of H2O (pH 11.0) to lyse the neutrophils and disperse the bacteria sufficiently to permit evaluation by a colony assay. All tests were performed in triplicate to assure their reproducibility.
Statistical analysis
Data are expressed as means ± standard deviation. Differences in string length (mm) and phagocytosis (%) were analyzed using the Mann–Whitney U-test. The counts of surviving bacteria were analyzed using the log rank test. Data were considered statistically significant when two-sided P values were < 0.05.
Disclosure of Potential Conflicts of Interest
There are no potential conflicts of interest for the authors.
Acknowledgments
This study was supported by grants from Ministry of Science and Technology of Taiwan (MOST 100-2314-B-182A-026, 100-2314-B-758-001, and 101-2314-B-182A-061-MY3). We are grateful to Leung Kei Siu, PhD, National Health Research Institutes, for kindly providing the K. pneumoniae strain DT-X.
References
- 1.Lee CH, Leu HS, Wu TS, Su LH, Liu JW. Risk factors for spontaneous rupture of liver abscess caused by Klebsiella pneumoniae. Diagn Microbiol Infect Dis. 2005;52:79–84. doi: 10.1016/j.diagmicrobio.2004.12.016. [DOI] [PubMed] [Google Scholar]
- 2.Ko WC, Paterson DL, Sagnimeni AJ, Hansen DS, Von Gottberg A, Mohapatra S, Casellas JM, Goossens H, Mulazimoglu L, Trenholme G, et al. Community-acquired Klebsiella pneumoniae bacteremia: global differences in clinical patterns. Emerg Infect Dis. 2002;8:160–6. doi: 10.3201/eid0802.010025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Fang CT, Lai SY, Yi WC, Hsueh PR, Liu KL, Chang SC. Klebsiella pneumoniae genotype K1: an emerging pathogen that causes septic ocular or central nervous system complications from pyogenic liver abscess. Clin Infect Dis. 2007;45:284–93. doi: 10.1086/519262. [DOI] [PubMed] [Google Scholar]
- 4.Cortés G, Borrell N, de Astorza B, Gómez C, Sauleda J, Albertí S. Molecular analysis of the contribution of the capsular polysaccharide and the lipopolysaccharide O side chain to the virulence of Klebsiella pneumoniae in a murine model of pneumonia. Infect Immun. 2002;70:2583–90. doi: 10.1128/IAI.70.5.2583-2590.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lee CH, Liu JW, Su LH, Chien CC, Li CC, Yang KD. Hypermucoviscosity associated with Klebsiella pneumoniae-mediated invasive syndrome: a prospective cross-sectional study in Taiwan. Int J Infect Dis. 2010;14:e688–92. doi: 10.1016/j.ijid.2010.01.007. [DOI] [PubMed] [Google Scholar]
- 6.Wu JH, Wu AM, Tsai CG, Chang XY, Tsai SF, Wu TS. Contribution of fucose-containing capsules in Klebsiella pneumoniae to bacterial virulence in mice. Exp Biol Med (Maywood) 2008;233:64–70. doi: 10.3181/0706-RM-170. [DOI] [PubMed] [Google Scholar]
- 7.Sawyer DW, Donowitz GR, Mandell GL. Polymorphonuclear neutrophils: an effective antimicrobial force. Rev Infect Dis. 1989;11(Suppl 7):S1532–44. doi: 10.1093/clinids/11.Supplement_7.S1532. [DOI] [PubMed] [Google Scholar]
- 8.Lin JC, Chang FY, Fung CP, Xu JZ, Cheng HP, Wang JJ, Huang LY, Siu LK. High prevalence of phagocytic-resistant capsular serotypes of Klebsiella pneumoniae in liver abscess. Microbes Infect. 2004;6:1191–8. doi: 10.1016/j.micinf.2004.06.003. [DOI] [PubMed] [Google Scholar]
- 9.Lin JC, Siu LK, Fung CP, Tsou HH, Wang JJ, Chen CT, Wang SC, Chang FY. Impaired phagocytosis of capsular serotypes K1 or K2 Klebsiella pneumoniae in type 2 diabetes mellitus patients with poor glycemic control. J Clin Endocrinol Metab. 2006;91:3084–7. doi: 10.1210/jc.2005-2749. [DOI] [PubMed] [Google Scholar]
- 10.Varki A, Angata T. Siglecs--the major subfamily of I-type lectins. Glycobiology. 2006;16:1R–27R. doi: 10.1093/glycob/cwj008. [DOI] [PubMed] [Google Scholar]
- 11.Carlin AF, Lewis AL, Varki A, Nizet V, Group B. Group B streptococcal capsular sialic acids interact with siglecs (immunoglobulin-like lectins) on human leukocytes. J Bacteriol. 2007;189:1231–7. doi: 10.1128/JB.01155-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Carlin AF, Uchiyama S, Chang YC, Lewis AL, Nizet V, Varki A. Molecular mimicry of host sialylated glycans allows a bacterial pathogen to engage neutrophil Siglec-9 and dampen the innate immune response. Blood. 2009;113:3333–6. doi: 10.1182/blood-2008-11-187302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Varki A. Sialic acids in human health and disease. Trends Mol Med. 2008;14:351–60. doi: 10.1016/j.molmed.2008.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Severi E, Hood DW, Thomas GH. Sialic acid utilization by bacterial pathogens. Microbiology. 2007;153:2817–22. doi: 10.1099/mic.0.2007/009480-0. [DOI] [PubMed] [Google Scholar]
- 15.Di Martino P, Bertin Y, Girardeau JP, Livrelli V, Joly B, Darfeuille-Michaud A. Molecular characterization and adhesive properties of CF29K, an adhesin of Klebsiella pneumoniae strains involved in nosocomial infections. Infect Immun. 1995;63:4336–44. doi: 10.1128/iai.63.11.4336-4344.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tigyi Z, Gährs W, Emody L, Makovitzky J. Topo-optical investigations on the surface of bacterial cells during the phagocytosis of Klebsiella pneumoniae in mouse. Acta Histochem. 2009;111:300–7. doi: 10.1016/j.acthis.2008.11.009. [DOI] [PubMed] [Google Scholar]
- 17.Harvey HA, Swords WE, Apicella MA. The mimicry of human glycolipids and glycosphingolipids by the lipooligosaccharides of pathogenic neisseria and haemophilus. J Autoimmun. 2001;16:257–62. doi: 10.1006/jaut.2000.0477. [DOI] [PubMed] [Google Scholar]
- 18.Nassif X, Fournier JM, Arondel J, Sansonetti PJ. Mucoid phenotype of Klebsiella pneumoniae is a plasmid-encoded virulence factor. Infect Immun. 1989;57:546–52. doi: 10.1128/iai.57.2.546-552.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ferrari J, Harris R, Warner TG. Cloning and expression of a soluble sialidase from Chinese hamster ovary cells: sequence alignment similarities to bacterial sialidases. Glycobiology. 1994;4:367–73. doi: 10.1093/glycob/4.3.367. [DOI] [PubMed] [Google Scholar]
- 20.Schauer R. Chemistry, metabolism, and biological functions of sialic acids. Adv Carbohydr Chem Biochem. 1982;40:131–234. doi: 10.1016/S0065-2318(08)60109-2. [DOI] [PubMed] [Google Scholar]
- 21.Ikehara Y, Ikehara SK, Paulson JC. Negative regulation of T cell receptor signaling by Siglec-7 (p70/AIRM) and Siglec-9. J Biol Chem. 2004;279:43117–25. doi: 10.1074/jbc.M403538200. [DOI] [PubMed] [Google Scholar]
- 22.Nicoll G, Avril T, Lock K, Furukawa K, Bovin N, Crocker PR. Ganglioside GD3 expression on target cells can modulate NK cell cytotoxicity via siglec-7-dependent and -independent mechanisms. Eur J Immunol. 2003;33:1642–8. doi: 10.1002/eji.200323693. [DOI] [PubMed] [Google Scholar]
- 23.Lin YT, Siu LK, Lin JC, Chen TL, Tseng CP, Yeh KM, Chang FY, Fung CP. Seroepidemiology of Klebsiella pneumoniae colonizing the intestinal tract of healthy Chinese and overseas Chinese adults in Asian countries. BMC Microbiol. 2012;12:13. doi: 10.1186/1471-2180-12-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vogel U, Weinberger A, Frank R, Müller A, Köhl J, Atkinson JP, Frosch M. Complement factor C3 deposition and serum resistance in isogenic capsule and lipooligosaccharide sialic acid mutants of serogroup B Neisseria meningitidis. Infect Immun. 1997;65:4022–9. doi: 10.1128/iai.65.10.4022-4029.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Merino S, Camprubí S, Albertí S, Benedí VJ, Tomás JM. Mechanisms of Klebsiella pneumoniae resistance to complement-mediated killing. Infect Immun. 1992;60:2529–35. doi: 10.1128/iai.60.6.2529-2535.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yoshida K, Matsumoto T, Tateda K, Uchida K, Tsujimoto S, Yamaguchi K. Induction of interleukin-10 and down-regulation of cytokine production by Klebsiella pneumoniae capsule in mice with pulmonary infection. J Med Microbiol. 2001;50:456–61. doi: 10.1099/0022-1317-50-5-456. [DOI] [PubMed] [Google Scholar]
- 27.Bitter T, Muir HM. A modified uronic acid carbazole reaction. Anal Biochem. 1962;4:330–4. doi: 10.1016/0003-2697(62)90095-7. [DOI] [PubMed] [Google Scholar]
- 28.Kolisis FN. An immobilized bienzyme system for assay of sialic acid. Biotechnol Appl Biochem. 1986;8:148–52. [PubMed] [Google Scholar]
- 29.Lee HC, Chuang YC, Yu WL, Lee NY, Chang CM, Ko NY, Wang LR, Ko WC. Clinical implications of hypermucoviscosity phenotype in Klebsiella pneumoniae isolates: association with invasive syndrome in patients with community-acquired bacteraemia. J Intern Med. 2006;259:606–14. doi: 10.1111/j.1365-2796.2006.01641.x. [DOI] [PubMed] [Google Scholar]
- 30.Heinzelmann M, Gardner SA, Mercer-Jones M, Roll AJ, Polk HC., Jr. Quantification of phagocytosis in human neutrophils by flow cytometry. Microbiol Immunol. 1999;43:505–12. doi: 10.1111/j.1348-0421.1999.tb02435.x. [DOI] [PubMed] [Google Scholar]
- 31.Hampton MB, Vissers MC, Winterbourn CC. A single assay for measuring the rates of phagocytosis and bacterial killing by neutrophils. J Leukoc Biol. 1994;55:147–52. doi: 10.1002/jlb.55.2.147. [DOI] [PubMed] [Google Scholar]