

Abstract
As a kind of medicine which can also be used as food, Alpinia oxyphylla Miq. has a long clinical history in China. A variety of studies demonstrated the significant neuroprotective activity effects of chloroform (CF) extract from the fruits of Alpinia oxyphylla. In order to further elucidate the possible mechanisms of CF extract which mainly contains sesquiterpenes with neuroprotection on the cognitive ability, mice were injected with Aβ 1−42 and later with CF in this study. The results showed that the long-term treatment of CF enhanced the cognitive performances in behavior tests, increased activities of glutathione peroxidase (GSH-px) and decreased the level of malondialdehyde (MDA), acetylcholinesterase (AChE), and amyloid-β (Aβ), and reversed the activation of microglia, degeneration of neuronal acidophilia, and nuclear condensation in the cortex and hippocampus. These results demonstrate that CF ameliorates learning and memory deficits by attenuating oxidative stress and regulating the activation of microglia and degeneration of neuronal acidophilia to reinforce cholinergic functions.
1. Introduction
As the global population ages, Alzheimer's disease (AD) is rapidly becoming an urgent public health challenge. It has been estimated that the number of people with dementia in the worldwide was 35.6 million in 2010, and this number will almost double every 20 years, to 65.7 million in 2030 and 115.4 million in 2050 [1]. Therefore, searching for safe, better tolerated, and effective drugs is necessary.
AD is a progressive degenerative disease of the brain which is characterized by deterioration of memory and cognitive functions with formation of senile plaques and neurofibrillary tangles and the loss of synapses in the selected regions of the brain. Several hypotheses of AD, including the β-amyloid peptide (Aβ) cascade hypothesis and the cholinergic hypothesis, have been applied to investigate the etiology of AD [2]. Scientists have proposed the cholinergic hypothesis; namely, the alternations of cholinergic system are closely related to the damage of cognitive function and AD. Based on this theory, a number of studies have been launched on acetyl cholinesterase (AChE) inhibitor, and then AChE inhibitors have been successfully developed for clinical treatment of AD [3–5]. To eliminate the deposition of Aβ, the main research strategy includes the following two pathways: one is to search Aβ secretase inhibitors, mainly including β-secretase inhibitors; the other one is to prepare antibodies of Aβ by using the immunological method in order to reduce the deposition of Aβ.
Aβ is a 40–42 amino acid proteolytic fragment of amyloid precursor protein (APP). The cascade begins with the cleavage of the APP sequentially by β- and γ-secretase. The 42 amino acid Aβ fragment self-assembles into oligomers. Its oligomers have been considered as the principal toxic substances which induce oxidative stress, neuronal apoptosis, and increase of neuronal loss [6]. Aβ 1–42 also has direct pharmacologic effects on synaptic function, impairing memory, and long-term potentiation in animal models [7]. Neurofibrillary tangle formation, oxidation, excitotoxicity, inflammation, synaptic compromise, demyelination, mitochondrial dysfunction, and neurodegeneration follow the interaction with oligomeric Aβ [8]. Aβ may also fibrillize to form insoluble aggregates that compose the neuritic plaques characteristic of AD. In rats treated with Aβ, the release of acetylcholine (ACh) and dopamine stimulated by nicotine decreased in the brain, which demonstrated the learning deficits observed in the Aβ protein-infused rats, is partly due to the impairment of neurotransmitter [9]. On the basis of the accumulating evidences on pathological roles of Aβ in the progress of AD, Aβ-injected animals have become a useful model for understanding the pathogeneses and progression of AD.
Alpinia oxyphylla Miq. is regarded as a precious drug and also a kind of condiment in Hainan district in southern China. Sesquiterpenes, diterpenes, flavonoids, and diarylheptanoids have been found in Alpinia oxyphylla Miq. previously and some of which showed inhibitory effect on nitric oxide (NO) production in lipopolysaccharide- (LPS-) activated mouse peritoneal macrophages [10–13]. There have been growing evidences showing that chloroform extract from the fruits of Alpinia oxyphylla possesses significant neuroprotective activity [14]. There are a few studies showed that Alpinia oxyphylla had therapeutic efficacy for senile dementia by reducing the apoptosis and free radical. However, the exact mechanism or components are not explicit [15]. On the basis of the findings above mentioned, we hypothesized that chloroform extract of Alpinia oxyphylla could ameliorate aging through inhibiting oxidative stress, improving the cholinergic system, and reducing Aβ levels in the brain. Therefore, in the present study, we investigated the actions of chloroform extract of Alpinia oxyphylla on cognitive ability, oxidative stress biomarkers, and Aβ deposition in the hippocampus and cortex of aging mice induced by Aβ 1–42 to elucidate the underlying molecular mechanisms. Furthermore, compounds of the active fraction of the extract have been identified.
2. Materials and Methods
2.1. Material
Alpinia oxyphylla Miq. was purchased from Shenyang Tongrentang Drug Co., Ltd. (Shenyang, China). The crude drugs were of high quality and authenticated by Professor Ying Jia of Pharmacognosy Department, Shenyang Pharmaceutical University. Donepezil was supplied by Wanbang Pharmaceutical Company (Zhejiang, China). Aβ 1–42 was obtained from Sigma-Aldrich (St Louis, MO, USA) and dissolved in sterile physiological saline (1.0 μg/μL) in the tube, which was then sealed and incubated for 120 h at 37°C to cause the peptide to aggregate. Commercial kits used for determination of AChE, GSH-px, MDA, β-secretase, and Aβ 1–42 were purchased from Jiancheng Institute of Biotechnology (Nanjing, China) and Qiming Biotechnology Company (Shanghai, China).
2.2. Sample Extraction and Fractionation
The air-dried fruits of Alpinia oxyphylla Miq. (10.0 kg) were extracted three times for 2 h each time by refluxing in 95% ethanol (1 : 10, w/v). The filtrates were concentrated and dried in vacuum at 60°C. The crude extract was dissolved in distilled water and then partitioned sequentially in different solvents, namely, petroleum ether, chloroform (CF), ethyl acetate, and n-butanol, to fractionate the polar and nonpolar compounds in the crude extract. The resulting solvent fractions were concentrated by rotary evaporator and dried by a vacuum oven at 45°C. The doses of chloroform extract were expressed as gram of the original dry materials per kilogram body weight.
2.3. UPLC-ESI/MS Analysis for CF
The chemical composition of CF was analysed by using a Waters-UPLC-Q-TOF/MS with an ultraviolet/visible detector (UV/Vis) coupled to an ion trap mass spectrometer with an ESI interface. The chromatogram was recorded at 255 nm. An HSS T3 Column (100 mm × 2.1 mm, 1.8 μm) with the column temperature set at 25°C was used for separation. The injection volume was 5 μL, and elution was performed at a flow rate of 0.6 mL/min using a mixture consisting of acetonitrile (A) and 0.1% (v/v) formic acid (B). A gradient program was used as follows: 0–3 min, 20% A; 3–20 min, 20%–50% A; 20–25 min, 50%–90% A; 25-26 min, 90%–20% A; 26–28 min, 20% A.
Mass analyses were performed using an ESI interface in the positive ion mode. The data were acquired in the full scan and MS/MS2 scanning mode. The optimized instrumental parameters were set as follows: positive mode: desolvation temperature, 250°C; source temperature, 120°C; capillary, 3.0 kV; sampling cone 30.0; extraction cone 4.0; source temperature 130; desolvation temperature 450; desolvation gas flow (L/Hr) 800.0; collision energy 6.0 ev; and scan range, m/z 100–1000 amu.
2.4. Animals and Administration
Seventy-two male ICR mice weighing 18–22 g were provided by the Experimental Animal Center of Shenyang Pharmaceutical University (Shenyang, China). They were maintained on standard laboratory conditions of temperature 25 ± 1°C and a 12 h light/12 h dark cycle with food and water available ad libitum for the duration of the study. After 1 week of acclimatization, all mice were randomly divided into 6 groups (n = 12/group): vehicle control group, sham-operated group, model group, donepezil group, chloroform group 1 (CF1), and chloroform group 2 (CF2). All the mice were anesthetized with 3.5% chloral hydrate (0.1 mL/10 g). Then, model group, donepezil group, CF1 group (180 mg/kg), and CF2 (360 mg/kg) were injected with aggregated Aβ 1–42 peptide (3 μL) into the right lateral ventricle within 3 min by means of a stereotaxic apparatus (AP, −0.5 mm, ML, ±1.1 mm, DV, −3.0 mm). The needle was removed with 1 min delay to allow diffusion. Mice in the sham-operated group were injected in an identical manner with the same amount of physiological saline (3 μL). From the next day, mice in the CF1 and CF2 were administered with CF of 180 mg/kg and 360 mg/kg in distilled water containing 1.0% DMSO (20 mL/kg, i.g.) daily for 20 consecutive days by intragastric infusion (i.g.). Mice in donepezil group were administered with DPZ (0.65 mg/kg, i.g.). Mice in vehicle control group, sham-operated group, and model group were treated with distilled water containing 1.0% DMSO (20 mL/kg, i.g.) in the same period. The experiment schedule is shown in Figure 1. Animal care was in accordance with the Guidelines for Animal Experimentation of Shenyang Pharmaceutical University and the protocol was approved by the Animal Ethics Committee of the institution.
Figure 1.
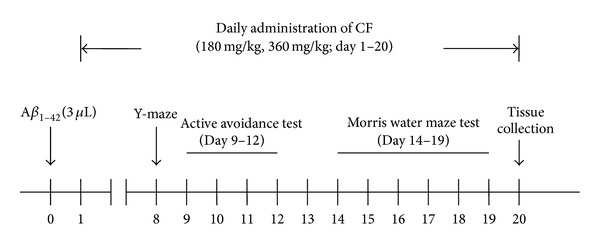
The experiment schedule.
2.5. Behavioral Experiments
2.5.1. Y-Maze Test
Y-maze test was used as a measure of immediate spatial working memory which was a form of short-term memory [16]. Y-maze is a three-arm maze with equal angles between all arms. Mice were initially placed within one arm, and the sequence and number of arm entries were recorded manually for each mouse over an 8 min period. The alternation score (%) for each mouse was defined as the ratio of the actual number of alternations to the possible number (defined as the total number of arm entries minus two) multiplied by 100 as shown by the following equation: Alternation% = [(Number of alternations)/(Total arm entries −2)] × 100%. The number of arm entries was used as an indicator of locomotor activity.
2.5.2. Active Avoidance Test
Learning and memory ability were detected by the active avoidance as described previously [17]. During the training session each trial began when the animal was introduced into any of the compartments with its head oriented toward the wall opposite to the mouse hole. After a variable period (±60 s), a conditioned stimulus was delivered. If the mouse crossed to the opposite compartment during the presence of the conditioned stimulus, an avoidance response was scored. If the mouse did not cross during the presence of the conditioned stimulus the unconditioned stimulus was delivered and remained on for 10 s or until the animal escaped to the opposite compartment. If the animals crossed to the opposite compartment within 10 s, an escape response was scored and a new trial began. The session consisted of 30 trials and it ended when 30 trials finished. The percent of conditioned avoidance response and total time of test were recorded.
2.5.3. Morris Water Maze Test
Learning and memory ability were detected by Morris water maze test as described previously [18]. The experimental apparatus consisted of a circular water basin (150 cm in diameter, 60 cm in height), containing water (25 ± 2°C) to a depth of 40 cm, which was rendered opaque by adding black nontoxic carbon ink. A platform (9 cm in diameter, 38.5 cm in height) was submerged below the water surface and placed at the midpoint of one quadrant. It was given 90 s to find the platform and was allowed to rest on it for 15 s. The animals which failed to find the location within the given time were gently guided to the platform and were allowed to stay on it for 15 s; each mouse was given two trial sessions each day for five consecutive days, with an intertrial interval of about 15 min. To determine whether the animal would take a spatial learning strategy to locate the platform, a single spatial probe trial was assessed on day six, the platform was removed from the water basin, and the mice were allowed to swim freely for 60 s. All data were recorded and analyzed by a computerized video imaging analysis system (Huaibei Zhenghua biology apparatus Co., Ltd, Anhui, China).
2.6. Sample Preparation
After probe trial sessions of Morris water maze test, 72 mice were sacrificed by cervical dislocation and the brain was immediately removed. The cerebral cortex and hippocampus of 6 mice in each group were each dissected out [19]. The brain (except for the cerebellum) of the rest of the 6 mice in each group was also dissected out. Each part of the brain tissue was stored at −80°C until the biochemical studies [20]. Before detection, each part of the brain tissue was rapidly homogenized in ice-cold saline and the homogenates were centrifuged at 3500 rpm at 4°C for 15 min. The supernatant was collected for assay.
2.7. Assay of GSH-px and Lipid Peroxidation within the Brain of Mice
The left cerebral cortex tissue was homogenized in ice-cold saline and centrifuged at 2000 rpm for 10 min, and the supernatant was collected. The activities of GSH-px and the amount of MDA in the supernatant were measured using commercial assay kits.
2.8. Determination of AChE Activity in the Cerebral Cortex and Hippocampus of Mice
The activities of AChE were measured using colorimetric methods [21]. Briefly, the reaction mixture containing samples, DTNB, and sodium phosphate (1 mmol/L, pH 8.0) was preincubated for 10 min at 37°C, and then acetylthiocholine iodide was added to the reaction mixture to incubate for 5 min at 37°C. The absorbance was measured at 412 nm at room temperature. AChE activity was expressed as nmol/mg of protein.
2.9. Assay of β-Secretase in the Cerebral Cortex of Mice
The activities of β-secretase in cortical of mouse brain were measured using a specific β-secretase ELISA kit according to the manufacturers' protocols. This formation of fluorescence was read using a fluorescence plate reader with excitation at 335–355 nm and emission at 450 nm.
2.10. Determination of Aβ 1–42 in Hippocampus of Mice
Each brain was homogenized in 8 vol. (w/v) of cold 5 mol/L guanidine HCl/50 mmol/L Tris-HCl and mixed at room temperature for 4 h. Dilutions of the extracts were made in Dulbecco's phosphate-buffered saline containing 5% BSA and 0.03% Tween 20 (pH 7.4) supplemented with 1x protease inhibitor cocktail (Roche, Germany). Following centrifugation at 16,000 ×g for 20 min at 4°C, aliquots were diluted with sample buffer provided by the manufacturer and used for the measurement of Aβ 1–42 levels by enzyme-linked immunosorbent assay (ELISA) (IBL, Germany).
2.11. Histology
For the histological examination of tissue sections, hematoxylin-eosin and Congo red were used as described previously [22]. The brain was removed and kept overnight in the last fixative solution for dehydration. Then cut it into transparent slices and imbedded it in paraffin. Brain samples were cut into coronal sections. Serial sections were selected around the needle trace. Sections were stained with hematoxylin-eosin reagent and then dehydrated with graded alcohol and mounted with neutral balsam medium to observe changes in the cortical and hippocampal neurons.
2.12. Statistical Analysis
All values were expressed as the mean ± SD. Statistical differences in all groups were analyzed using one-way ANOVA. Student's t-test was used to determine significant differences between groups. Differences were considered statistically significant at a value of P < 0.05.
3. Results and Analysis
3.1. UPLC-ESI/MS Analysis for Chloroform Extract of Alpinia oxyphylla
The chemical compositions in CF were analysed via UPLC-ESI/MS. The chromatogram of CF was obtained at 255 nm (Figure 2). The results of UPLC-ESI/MS and tentative identification are shown in Table 1. Among these compounds, some showed typical fragmentation patterns as previously reported, while the others were identified based on the retention times and the UV spectra of the reference standards [23, 24].
Figure 2.
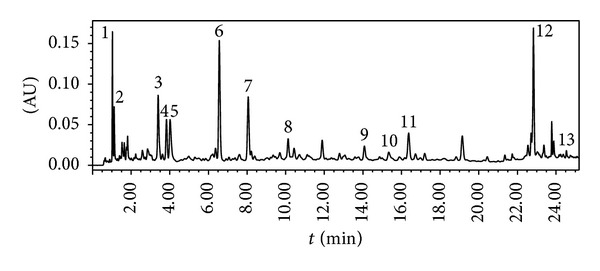
UPLC chromatogram of chloroform extract of Alpinia oxyphylla at 255 nm. 1, oxyphyllanene A; 2, protocatechuic acid; 4, 11S-nootkatone-11,12-diol; 5, 11R-nootkatone-11,12-diol; 6, teuhetenone A; 7, teuhetenone B; 11, oxyphyllol B; 12, Nootkatone; 13, dibutyl phthalate.
Table 1.
Retention times, MS, and MS/MS2 fragmentation patterns of the substances in chloroform extract of Alpinia oxyphylla.
| Peak | T R (min) | MS+ (m/z) | Major fragments (positive ion mode) (m/z) | Tentative identification |
|---|---|---|---|---|
| 1 | 1.08 | 192.1234 | 175.1[M+H-H2O]+→ 151.1[M+H-CH2CO]+
→ 133.1[M+H-CH2CO-H2O]+ |
Oxyphyllanene A |
|
| ||||
| 2 | 1.22 | 153.8901 | 110.8[M+H-CO2]+ | Protocatechuic acid |
|
| ||||
| 3 | 3.47 | 220.1544 | 177.1 → 133.1 | Unknown |
|
| ||||
| 4 | 3.96 | 252.1821 | 235.1[M+H-H2O]+→ 206.1[M-3CH3]+
→ 189.1[M+H-3CH3-H2O]+→ 177.1 [M-C3H5-2OH]+→ 107.1 |
11S-nootkatone-11,12-diol |
|
| ||||
| 5 | 4.06 | 252.1797 | 235.1[M+H-H2O]+→ 206.1[M-3CH3]+
→ 189.1[M+H-3CH3-H2O]+→ 177.1 [M-C3H5-2OH]+→ 107.1 |
11R-nootkatone-11,12-diol |
|
| ||||
| 6 | 6.62 | 194.1307 | 177.1[M+H-H2O]+→ 149.1[M+H-H2O-CO]+→ 135.1 [M+H-H2O-CO-CH3]+→ 120.1[M+H-H2O-CO-2CH3]+ |
Teuhetenone A |
|
| ||||
| 7 | 8.10 | 194.1389 | 177.1[M+H-H2O]+→ 161.1[M+H-H2O-CH2]+→ 137.1[M+H-H2O-CO-CH2]+ |
Teuhetenone B |
|
| ||||
| 8 | 10.18 | 220.1547 | 203.1 → 159.1 | Unknown |
|
| ||||
| 9 | 14.21 | 222.1705 | 180.1 → 138.1 | Unknown |
|
| ||||
| 10 | 15.82 | 216.1595 | 179.1 → 137.1 | Unknown |
|
| ||||
| 11 | 16.48 | 234.1704 | 217.2[M+H-H2O]+→ 207.2[M+H-CO]+→ 203.2[M+H-H2O-CH2]+
→ 189.2[M+H-H2O-2CH2]+ |
Oxyphyllol B |
|
| ||||
| 12 | 20.47 | 218.1671 | 176.1[M+H-C3H6]+ → 163.1[M+H-C3H5-CH]+ → 149.1[M+H-C3H5-CH-CH2]+ → 135.1[M+H-C3H5-CH-CO]+ |
Nootkatone |
|
| ||||
| 13 | 24.53 | Dibutyl phthalate | ||
3.2. Y-Maze Test
The effect of CF on short-term or working memory was investigated in the spontaneous alternation behavior Y-maze test. As presented in Figure 3(a), spontaneous alternation was significantly different between groups (F (5, 54) = 3.338, P < 0.05). The spontaneous alternation of model mice was significantly lower than that of sham-operated mice by 18% (P < 0.05), and the lowered spontaneous alternation induced by Aβ 1–42 was significantly reversed by CF2 (360 mg/kg) by 17% (P < 0.01). Moreover, the effect of CF (360 mg/kg) on the spontaneous alternation behavior was similar to that of donepezil (P < 0.05, Figure 3(a)). However, numbers of arm entries were similar in all experimental groups, demonstrating that general locomotor activity was not affected by CF (Figure 3(b)).
Figure 3.
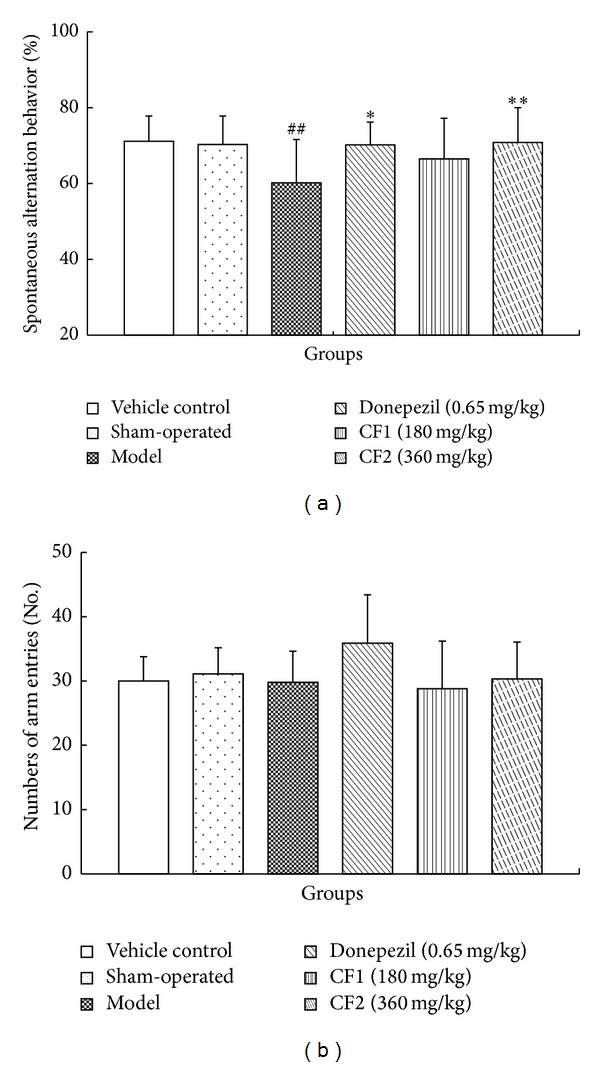
Effect of CF on Aβ 1–42-induced memory deficits in the Y-maze test. Spontaneous alternation behavior (a) and the number of arm entries (b) during an 8 min session were measured. Data represent means ± SD (n = 10). # P < 0.05, compared with sham-operated group. *P < 0.05, **P < 0.01 compared with model group.
3.3. Active Avoidance Test
We assessed the effects of CF on learning and memory ability in mice exposed to Aβ 1–42 (3.0 μg/mouse) using active avoidance test described above. Table 2 showed that condition avoidance response percent every day and total time in four days but had no marked difference between the control and sham-operated groups. Compared with control group, the condition avoidance response (CR) was significantly decreased (day 2–4) in model group (d2, F (5, 54) = 4.289, P < 0.05; d3, F (5, 54) = 3.617, P < 0.01; d4, F (5, 54) = 4.471, P < 0.001) and total time was significantly increased (F (5, 54) = 4.490, P < 0.01). These results revealed that the Aβ 1–42-treated mice had obvious cognitive impairment. Moreover, the decrease of CR was reversed, respectively, by CF (180 mg/kg and 360 mg/kg) from second day to fourth day (P < 0.001 and P < 0.01; P < 0.05; P < 0.01 and P < 0.05 versus the model) and by donepezil treatment from the third to fourth day (P < 0.05, P < 0.05 versus the model). Besides, the increase of total time was shortened, respectively, by CF (180 mg/kg and 360 mg/kg) (P < 0.01 and P < 0.05 versus the model) and by donepezil treatment (P < 0.05 versus the model). In addition, the memory enhancing activity of CF was shown to be more potent than donepezil-treated group (0.65 mg/kg body weight, p.o.).
Table 2.
Effect of CF on condition avoidance response and total time induced by Aβ 1–42 in mice.
| Group | Condition avoidance response/% | Total time/s | |||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 (d) | ||
| Control | 46.0 ± 29.3 | 59.3 ± 20.0 | 86.0 ± 9.7 | 86.0 ± 9.7 | 1542 ± 102 |
| Sham-operated | 46.7 ± 30.5 | 74.0 ± 21.4 | 85.3 ± 14.3 | 85.3 ± 15.6 | 1482 ± 104 |
| Model | 34.0 ± 18.9 | 37.3 ± 20.4# | 49.3 ± 28.5### | 51.4 ± 23.3### | 1683 ± 70## |
| Donepezil 0.65 mg/kg | 42.7 ± 34.6 | 48.0 ± 26.9 | 70.0 ± 29.2* | 68.7 ± 23.5* | 1586 ± 115* |
| CF1 180 mg/kg | 48.7 ± 32.9 | 71.3 ± 18.6*** | 70.0 ± 24.0* | 74.7 ± 16.9** | 1544 ± 121** |
| CF2 360 mg/kg | 42.6 ± 27.5 | 67.3 ± 23.6** | 68.0 ± 22.0 | 68.7 ± 22.4* | 1585 ± 77* |
Results are expressed as mean ± SD, n = 60 (10 mice each group), # P < 0.05, ## P < 0.01, and ### P < 0.001 versus the control mice; *P < 0.05, **P < 0.01, and ***P < 0.001 versus the model group mice.
3.4. Morris Water Maze Test
Morris water maze test was used to assess the spatial learning and memory ability of animals. As shown in Figure 4(a), the mean latency to find the platform declined progressively during the five training days. The model group mice markedly spent longer time in finding the platform than the vehicle control mice in all training days (d3, F (5, 234) = 2.707, P < 0.05; d4, F (5, 234) = 4.259, P < 0.01; d5, F (5, 234) = 3.232, P < 0.05). These results revealed that the model group mice had significant cognitive impairment. Moreover, CF1 and CF2 group mice significantly shortened the escape latency compared with the model group mice from the third to fifth day (42.92 ± 17.31 s versus 60.66 ± 15.75 s, P < 0.05; 29.32 ± 11.25 s versus 46.29 ± 13.48 s, P < 0.01; and 28.87 ± 14.56 s versus 44.36 ± 13.52 s, P < 0.01, resp.) and from the third day backwards (39.01 ± 19.92 versus 60.66 ± 15.75 s, P < 0.01; 27.78 ± 9.70 s versus 46.29 ± 13.48 s, P < 0.01; 27.55 ± 11.42 s versus 44.36 ± 13.52 s, P < 0.01), respectively. Meanwhile, donepezil (0.65 mg/kg) treatment reached the similar effect from the fourth to fifth day of training trials (28.77 ± 10.75 s versus 46.29 ± 13.48 s, P < 0.01; 29.83 ± 14.33 s versus 44.36 ± 13.52, P < 0.05, resp.). Figure 4(b) illustrated the swim paths of mice in the second trial of the second and fifth day in this test. Mice tended to explore all four quadrants of the pool in the second day. Thereafter, they changed this search strategy. On the fifth day, the vehicle control mice swam in the direction of the platform; however, the model group mice took longer swimming paths. In the probe test (Figure 4(c)), the vehicle control mice spent longer time in the target quadrant than the model group mice (F (5, 54) = 3.376, P < 0.01). CF (180 and 360 mg/kg) and donepezil (0.65 mg/kg) treatment mice also took longer time in the target quadrant than the model group mice (P < 0.05, P < 0.001 and P < 0.05, resp.). It is more accurate to evaluate the effects of durg on impairment of spatial learning and memory through comparison of the number of crossing, as shown in Figure 4(d). The model group made fewer platform crossings than the vehicle control group (F (5, 54) = 2.628, P < 0.05) and the CF (180 and 360 mg/kg) and donepezil (0.65 mg/kg) treatments could increase the number of times of crossing over the platform site than the model group (P < 0.05, P < 0.01 and P < 0.05); however, CF2 had greater effect to reverse the deficit of spatial learning and memory induced by Aβ 1–42 than donepezil treatments.
Figure 4.
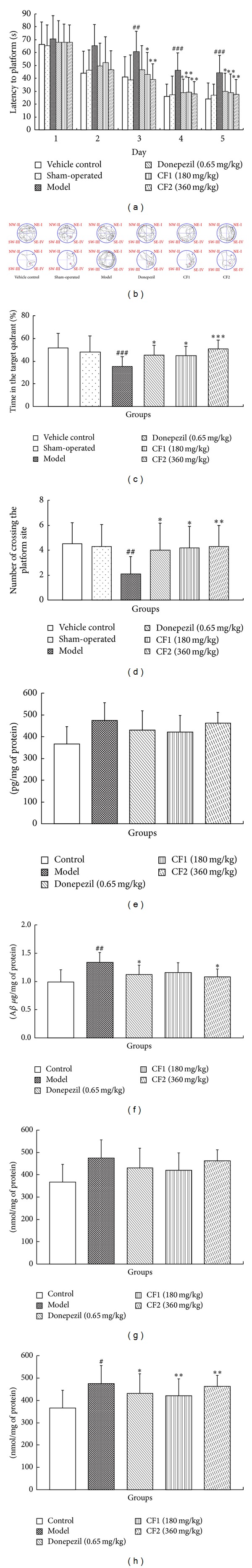
The Morris water maze test of mice treated with vehicle, Aβ 1–42, Aβ 1–42 + donepezil (0.65 mg/kg), Aβ 1–42 + CF (180 mg/kg), and Aβ 1–42 + CF (360 mg/kg), respectively. (a) Latencies to find a hidden platform in the water maze during five consecutive training days. (b) Search strategy of mice in the second trial on the second and fifth day. Traces show the swim path of all groups of mice. (c) The time spent in the quadrant where the platform was once placed within 60 s. (d) Number of crossings over the former platform location in the probe trial. (e) and (f) The effect of CF on β-secretase activity in the frontal cortex and accumulation of Aβ 1–42 in hippocampus. (g) and (h) The effect of CF on acetylcholinesterase activity in the cortex and hippocampus of Aβ 1–42-injected mice. Data are expressed as mean ± SD. # P < 0.05, ## P < 0.01, and ### P < 0.001 versus the control mice; *P < 0.05, **P < 0.01, and ***P < 0.001 versus the model group mice.
3.5. Effect of CF on Brain GSH-px Activities and MDA Contents
Table 3 summarized the antioxidant effect of CF in the Aβ 1–42-injected mice. GSH-px is an important antioxidant enzyme involved in cellular protection against damage caused by oxygen-derived free radicals, by means of removing harmful peroxide metabolites and blocking lipid peroxidation chain reaction. Aβ 1–42 suppressed GSH-px activity in the brain (F (4, 25) = 3.178, P < 0.05). However, CF2 (360 mg/kg) and donepezil groups displayed a significant elevation of GSH-px activity (P < 0.05 and P < 0.05 versus the model group, resp.). The MDA level in the brain of Aβ 1–42-injected mice was higher than that of the vehicle control group (F (4, 25) = 4.439, P < 0.01). The increase was ameliorated by treatment of CF at doses of 180 or 360 mg/kg or donepezil (P < 0.01, P < 0.01 and P < 0.05 versus the Aβ 1–42-treated group, resp.).
Table 3.
Effect of CF on brain tissue GSH-px activities and MDA contents.
| Group | GSH-px (U/mg protein) |
MDA (nmol/mg protein) |
|---|---|---|
| Control | 4.33 ± 0.49 | 1.42 ± 0.30 |
| Model | 3.44 ± 0.39# | 1.93 ± 0.47## |
| Donepezil 0.65 mg/kg | 4.34 ± 0.94∗ | 1.56 ± 0.40∗ |
| CF 180 mg/kg | 3.77 ± 0.77 | 1.31 ± 0.23∗∗ |
| CF 360 mg/kg | 4.54 ± 0.98∗ | 1.25 ± 0.21∗∗ |
Results are expressed as mean ± SD, n = 6, # P < 0.05, ## P < 0.01 versus the control mice; *P < 0.05, **P < 0.01 versus the model group mice.
3.6. Inhibitory Effect of CF on β-Secretase Activity in the Frontal Cortex and Accumulation of Aβ 1–42 in Hippocampus
The activity of β-secretase in the cortex of the model group was increased compared with vehicle control (Figure 4(e)). The administration of donepezil and CF lowered the increased activity of β-secretase induced by Aβ 1–42 injection in cortex of mouse brain. However, it did not show any significant differences (F (4, 25) = 1.690, P > 0.05).
We further measured the Aβ 1–42-levels in brain homogenate by ELISA. The ELISA results revealed that the Aβ 1–42 levels were increased in Aβ 1–42-treated group (F (4, 25) = 3.151, P < 0.05). Compared with the model group, the Aβ 1–42 levels were significantly decreased in donepezil and CF2 treatment groups (P < 0.05, Figure 4(f)).
3.7. Inhibitory Effect of CF on AChE Activity in the Frontal Cortex and Hippocampus
As shown in Figure 4(g), the administration of CF or donepezil to Aβ 1–42-injected mice produced no significant change on the activity of AchE in cortex (F (4, 25) = 1.533, P > 0.05). The activity of AchE in hippocampus was increased in Aβ 1–42-injected mice and greater than those in vehicle control (F (5, 54) = 3.151, P < 0.05). However, the increased activity of AchE in hippocampus was significantly inhibited by the treatment with CF1 and CF2 (P < 0.01 and P < 0.01) (Figure 4(h)).
3.8. Effects of CF on Neurodegenerative Changes in the Frontal Cortex and Hippocampus
As shown in Figure 5, (a) and (b) showed normal cells which are intact and there are less degeneration in normal cortex and hippocampus of mice. (c) Aβ 1–42-induced neuron death was observed consistently in the cortex and hippocampus, as indicated by the appearance of pyknotic black neurons, karyorrhexis, and karyolysis with condensed nucleus. Figure 5(d) showed moderate damage to the cortex and hippocampus area pretreated with donepezil 0.65 mg/kg, indicated by presence of less number of degenerative cells compared to the model group. Figures 5(e) and 5(f) showed mild damage to the cortex and hippocampus area pretreated with CF (180 mg/kg, 360 mg/kg) indicated by presence of medium number of degenerative cells compared to the model group.
Figure 5.
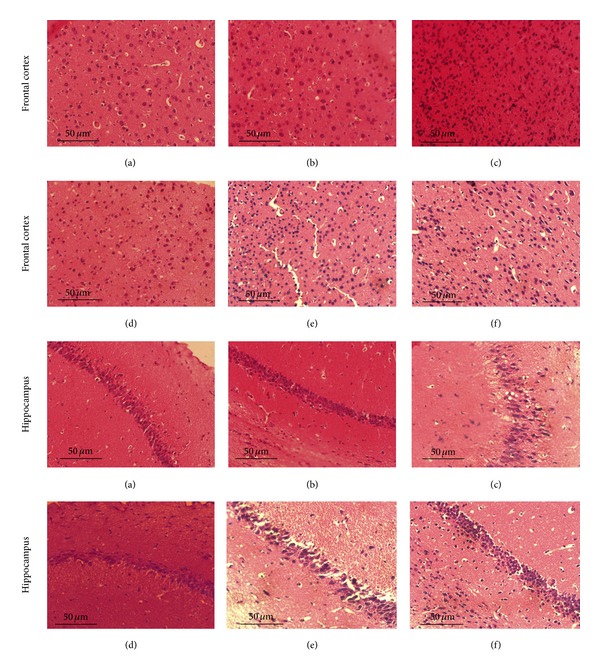
Effect of CF on neuronal degeneration in frontal cortex and hippocampus induced by Aβ 1–42 treatment in mice (H&E staining, 400x). (a) Vehicle control group; (b) sham-operated group; (c) model group; (d) donepezil group (0.65 mg/kg); (e) CF1 group (180 mg/kg); (f) CF2 group (360 mg/kg).
4. Discussion and Conclusion
In common parlance, Alpinia oxyphylla Miq. has been often touted as a drug hid in kitchen; therefore, it is not necessary to worry about the side effect of taking it. In terms of neuroprotection, it has been reported that ethanol extract of Alpinia oxyphylla fructus shows inhibition of tau protein phosphorylation in cell culture and the mechanism of sharp leaf galangal fruit extract can improve the spatial learning ability having business with its inhibiting ability on serum levels of cytokines [25]. For the preparation of the bioactive fraction of Alpinia oxyphylla which includes major sesquiterpenes with neuroprotective, antioxidant, or cognitive-enhancing activities, we chose chloroform fraction of ethanol extract of Alpinia oxyphylla Miq. However, screening of activities in vitro always lacks details of facts. To verify the potential of Alpinia oxyphylla, which was rich with neuroprotective or cognitive enhancing, we examined the protective effects of CF on cognitive impairment and neurotoxicity within the cortex and hippocampus in Aβ 1–42-injected mice.
However, Aβ is transported bidirectionally across the blood-brain barrier (BBB), that is, both in brain-to-blood (efflux) and the blood-to-brain (influx) directions. Therefore, how to remove the toxic Aβ42 peptide through BBB safely and effectively is considered to be an effective method of the prevention and treatment of AD. Previous studies have shown the influence of traditional Chinese aromatic medicine between the structure and function of BBB. Modern pharmacological studies indicate that the permeability of BBB could be changed by traditional Chinese aromatic medicine in three ways. Firstly, it can develop potency by permeating the BBB reinlessly, secondly, it can protect the brain tissue by reducing the permeability of BBB, and thirdly, it can promote the opening of BBB, while its mechanism of action may be concerned with restraining the expression and function of P-glycoprotein. We supposed that sesquiterpenoids contained in CF may play the same role.
At present, there is no animal model can mimic all the cognitive, behavioral, biochemical, and histopathological abnormalities observed in patients with AD. Mice intracerebroventricularly injected with Aβ 1–42 can mimic some cognitive deficits in AD, which is an economic and reliable model. Therefore, we first tested the memory enhancing effect of CF in mice treated with Aβ 1–42. The cognitive-enhancing activity of CF was evaluated by Y-maze test, active avoidance test, and Morris water maze test. CF mitigated the impairment of recognition induced by Aβ in mice. The treatment of CF more effectively improved the deficit in long term memory than in short term memory. Oral treatment with CF also exhibited a mitigation of memory deficit in mice induced by i.c.v. injection of Aβ on escape latency, time spent in target quadrant, and cross-platform times in Morris water maze test. The treatment with CF decreased the latency to a shorter level than that of nontreated normal control. In addition, cognitive-enhancing effect of CF was found to be much more effective than that of donepezil. Besides, the same effect was observed in active avoidance test; the data showed that CF exhibited significant influence on condition avoidance response (CR) and total time (TT). The results indicate that CF has the potential to ameliorate cognitive deficits induced by Aβ 1–42.
In spite of conflicting hypothesis on the pathological progress in AD patients, it is now generally recognized that the accumulated amyloid plaque in the brain is characteristic and diagnostic features of AD. β-secretase initiates the cleavage of β-amyloid precursor protein (APP) leading to the production of amyloid-β (Aβ); the gathering of Aβ is the main cause of senile plaque in the brain of AD patient, causes neuron damage and the hypofunction of cognition, and works as the center and cogallery in the pathogenesis of AD. The suppressant's partial inhibition of β-secretase can decrease the production of Aβ and get a manifesting therapeutic effect. Further study on the way BACE inhibitors work has favourable perspective in the treatment of AD. Our data showed that the activities of BACE1 increased by Aβ injection in the cortex of mouse brain were significantly inhibited by CF. Accumulation of deposits of Aβ is one of the classical neuropathological hallmarks in AD and may be the primary event in the pathogenesis of AD. In accordance with the reduction in BACE1 activity by CF, the aggregation of Aβ in the hippocampus of Aβ-injected mouse brain was found to be markedly reduced by CF by enzyme-linked immunosorbent assay (ELISA).
Growing data from experimental models and human brain studies suggest that oxidative stress, inflammation, and apoptotic cell death induced by Aβ play important roles in neuronal degeneration in AD [26]. Each of these factors can act independently or collectively damage neurons and disturb cognitive processes.
The oxidative stress hypothesis of AD suggests that the aggregation of Aβ on the membrane generates reactive oxygen species (ROS), resulting in protein oxidation, lipid peroxidation, cellular dysfunction, and subsequent neuronal death [27]. It may be one of the first pathogenic events during disease progression. In the Aβ 1–42-induced model mice, the level of MDA (a well-known indicator of lipid peroxidation) increased, and activities of GSH-px (the line of the antioxidant defense systems) decreased. GSH-px scavenges ROS by directly reacting with it and prevents H2O2-induced hydroxyl radical formation. GSH-px level parallels the antioxidant defense capacity and is a first indicator for oxidative stress in the brain [28]. Thus, the increase in activities of antioxidant defense systems is considered to be beneficial in the event of oxidative stress. The results indicated that an increase of oxidative stress occurred in the brain of Aβ 1–42-treated mice. After the administration of CF, the decreased activities of GSH-px, as well as the increased level of MDA, were restored in the brain. The possible mechanisms of CF are largely related to its antioxidant and other scavenging properties, for example, over nitric oxide (NO), possibly, through downregulation of NOS activity in brain. Previous phytochemical investigation on this plant revealed oxyphyllanene A, teuhetenone A, oxyphyllol B, and nootkatone with inhibitory activity against NO production in LPS and IFN-c-induced RAW 264.7 macrophages, which showed that these compounds exhibited potent activities of NO inhibitory and neuroprotection [12]. Besides, it was reported that Alpinia protocatechuic acid protects against oxidative damage in vitro and reduces oxidative stress in vivo [14]. Therefore, it is undoubtedly that CF could effectively attenuate oxidative stress induced by Aβ.
Microglia, the resident macrophages in the central nervous system, can be activated by Aβ. Oligo-Aβ 1–42 is always likely to damage neurons through the microglia-inflammation response. Activated microglia not only can eliminate Aβ but also can release inflammatory factors, which play a critical role in AD pathology. Recently reports strongly suggest that regulating microglia function may be a promising therapeutic approach to AD. From histological evidence of tissue sections, we observed that the model group mice showed significant microglial cell hyperplasia, neuronal disorder, and degeneration in the frontal cortex and hippocampus, while in CF (180 mg/kg and 360 mg/kg) treated group the microglia activation, neuronal acidophilia degeneration, and nuclear condensation decreased, resulting in homogeneous morphology of neuronal cellular.
It has been demonstrated that impairments in learning, memory, and behavior observed in AD patients are associated with the cholinergic hypofunction. Neuronal loss in the basal forebrain particularly within the septohippocampal acetylcholinergic systems involved in learning and memory processes constitutes a pathological hallmark of AD. The mechanism of aggregated Aβ 1–42 peptide-induced increase of AChE activity was related with two pathways: (1) indirect action-increase intracellular Ca2+ and free radical, which are symbols of cellular damage and (2) direct action-change conformation of AChE. Recently, it has been reported that ethanolic extracts of Alpinia oxyphylla Miq. showed most potent AChE inhibitory activities at the concentration of 0.1 mg/mL, with 44.49% inhibition among 48 traditional Chinese medicinal herbs [29]. In our research, administration of CF significantly showed greater inhibition activity of AChE than donepezil known as the most common prescribed AChE inhibitor in hippocampus of Aβ-injected mouse. It may be related to acetylcholinesterase inhibition of nootkatone [30].
From the above results, it could be deduced that cognitive-enhancing activity of CF might result in part from the inhibition on the accumulation of Aβ and the BACE1 and AChE and from the reduction in ROS by recovering the antioxidative defense system. These results suggested that chloroform extract of Alpinia oxyphylla containing bioactive sesquiterpenes, such as oxyphyllanene A, teuhetenone A, oxyphyllol B, and nootkatone, might offer a useful therapeutic choice in either the prevention or the treatment of Alzheimer's disease.
Acknowledgment
This research was supported by Shenyang Scientific Project no. F13-287-1-00 and Shenyang Scientific Project no. F12-153-9-00.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Prince M. The global prevalence of dementia. In: Prince M, Jackson J, editors. World Alzheimer Report 2009. Alzheimer’s Disease International; 2009. pp. 25–45. [Google Scholar]
- 2.Li XL, Wang DS, Zhao BQ, et al. Effects of Chinese herbal medicine fuzhisan on aged rats. Experimental Gerontology. 2008;43(9):853–858. doi: 10.1016/j.exger.2008.05.018. [DOI] [PubMed] [Google Scholar]
- 3.Krall WJ, Sramek JJ, Cutler NR. Cholinesterase inhibitors: a therapeutic strategy for Alzheimer disease. Annals of Pharmacotherapy. 1999;33(4):441–450. doi: 10.1345/aph.18211. [DOI] [PubMed] [Google Scholar]
- 4.Giacobini E. Cholinesterase inhibitors: new roles and therapeutic alternatives. Pharmacological Research. 2004;50(4):433–440. doi: 10.1016/j.phrs.2003.11.017. [DOI] [PubMed] [Google Scholar]
- 5.Ballard CG, Greig NH, Guillozet-Bongaarts AL, Enz A, Darvesh S. Cholinesterases: roles in the brain during health and disease. Current Alzheimer Research. 2005;2(3):307–318. doi: 10.2174/1567205054367838. [DOI] [PubMed] [Google Scholar]
- 6.Bulbarelli A, Lonati E, Cazzaniga E, et al. TrkA pathway activation induced by amyloid-beta (Abeta) Molecular and Cellular Neuroscience. 2009;40(3):365–373. doi: 10.1016/j.mcn.2008.12.006. [DOI] [PubMed] [Google Scholar]
- 7.Lesné S, Ming TK, Kotilinek L, et al. A specific amyloid-β protein assembly in the brain impairs memory. Nature. 2006;440(7082):352–357. doi: 10.1038/nature04533. [DOI] [PubMed] [Google Scholar]
- 8.Bates KA, Verdile G, Li Q-X, et al. Clearance mechanisms of Alzheimer's amyloid-Β peptide: implications for therapeutic design and diagnostic tests. Molecular Psychiatry. 2009;14(5):469–486. doi: 10.1038/mp.2008.96. [DOI] [PubMed] [Google Scholar]
- 9.Itoh A, Nitta A, Nadai M, et al. Dysfunction of cholinergic and dopaminergic neuronal systems in β- amyloid protein-infused rats. Journal of Neurochemistry. 1996;66(3):1113–1117. doi: 10.1046/j.1471-4159.1996.66031113.x. [DOI] [PubMed] [Google Scholar]
- 10.Luo JG, Lv XQ, Wang XB, Kong L. Sesquiterpenoids from the fruits of Alpinia oxyphylla and inhibition of nitric oxide production in lipopolysaccaride-activated macrophages. Phytochemistry Letters. 2012;5(1):134–138. [Google Scholar]
- 11.Muraoka O, Fujimoto M, Tanabe G, et al. Absolute stereostructures of novel norcadinane-and trinoreudesmane-type sesquiterpenes with nitric oxide production inhibitory activity from Alpinia oxyphylla. Bioorganic and Medicinal Chemistry Letters. 2001;11(16):2217–2220. doi: 10.1016/s0960-894x(01)00413-9. [DOI] [PubMed] [Google Scholar]
- 12.Morikawa T, Matsuda H, Toguchida I, Ueda K, Yoshikawa M. Absolute stereostructures of three new sesquiterpenes from the fruit of Alpinia oxyphylla with inhibitory effects on nitric oxide production and degranulation in RBL-2H3 cells. Journal of Natural Products. 2002;65(10):1468–1474. doi: 10.1021/np020078o. [DOI] [PubMed] [Google Scholar]
- 13.Xu J, Tan N, Zeng G, et al. Studies on chemical constituents in fruit of Alpinia oxyphylla . China Journal of Chinese Materia Medica. 2009;34(8):990–993. [PubMed] [Google Scholar]
- 14.Shi GF, An LJ, Jiang B, Guan S, Bao Y. Alpinia protocatechuic acid protects against oxidative damage in vitro and reduces oxidative stress in vivo. Neuroscience Letters. 2006;403(3):206–210. doi: 10.1016/j.neulet.2006.02.057. [DOI] [PubMed] [Google Scholar]
- 15.Yu X, An L, Wang Y, Zhao H, Gao C. Neuroprotective effect of Alpinia oxyphylla Miq. fruits against glutamate-induced apoptosis in cortical neurons. Toxicology Letters. 2003;144(2):205–212. doi: 10.1016/s0378-4274(03)00219-4. [DOI] [PubMed] [Google Scholar]
- 16.Sarter M, Bodewitz G, Stephens DN. Attenuation of scopolamine-induced impairment of spontaneous alternation behaviour by antagonist but not inverse agonist and agonist β-carbolines. Psychopharmacology. 1988;94(4):491–495. doi: 10.1007/BF00212843. [DOI] [PubMed] [Google Scholar]
- 17.León LA, Landeira-Fernandez J, Cardenas FP. Effects of chronic intracerebroventricular 3,4-methylenedioxy-N-methamphetamine (MDMA) or fluoxetine on the active avoidance test in rats with or without exposure to mild chronic stress. Behavioural Brain Research. 2009;205(1):259–264. doi: 10.1016/j.bbr.2009.06.039. [DOI] [PubMed] [Google Scholar]
- 18.Lu J, Zheng Y, Wu D, Luo L, Sun D, Shan Q. Ursolic acid ameliorates cognition deficits and attenuates oxidative damage in the brain of senescent mice induced by d-galactose. Biochemical Pharmacology. 2007;74(7):1078–1090. doi: 10.1016/j.bcp.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 19.Ahmed T, Gilani A. Inhibitory effect of curcuminoids on acetylcholinesterase activity and attenuation of scopolamine-induced amnesia may explain medicinal use of turmeric in Alzheimer's disease. Pharmacology Biochemistry and Behavior. 2009;91(4):554–559. doi: 10.1016/j.pbb.2008.09.010. [DOI] [PubMed] [Google Scholar]
- 20.Kang SY, Lee KY, Kim YC, Park MJ, Markelonis GJ, Oh TH. Decursin from Angelica gigas mitigates amnesia induced by scopolamine in mice. Neurobiology of Learning and Memory. 2003;79(1):11–18. doi: 10.1016/s1074-7427(02)00007-2. [DOI] [PubMed] [Google Scholar]
- 21.Ellman GL, Courtney KD, Andres V, Jr., Featherstone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochemical Pharmacology. 1961;7(2):88–IN2. doi: 10.1016/0006-2952(61)90145-9. [DOI] [PubMed] [Google Scholar]
- 22.Hyman BT, Van Hoesen GW, Damasio AR, Barnes CL. Alzheimer's disease: cell-specific pathology isolates the hippocampal formation. Science. 1984;225(4667):1168–1170. doi: 10.1126/science.6474172. [DOI] [PubMed] [Google Scholar]
- 23.Fraga BM, Hernandez MG, Mestres T, Terrero D, Arteaga JM. Nor-sesquiterpenes from Teucrium heterophyllum. Phytochemistry. 1995;39(3):617–619. [Google Scholar]
- 24.Xu J, Ji C, Zhang Y, Su J, Li Y, Tan N. Inhibitory activity of eudesmane sesquiterpenes from Alpinia oxyphylla on production of nitric oxide. Bioorganic and Medicinal Chemistry Letters. 2012;22(4):1660–1663. doi: 10.1016/j.bmcl.2011.12.114. [DOI] [PubMed] [Google Scholar]
- 25.Wong KK, Wan CC, Shaw PC. Ethanol extract of Alpinia oxyphylla fructus shows inhibition of tau protein phosphorylation in cell culture. Neurobiology of Aging. 2004;25(2):595–597. [Google Scholar]
- 26.Pimplikar SW, Nixon RA, Robakis NK, Shen J, Tsai L. Amyloid-independent mechanisms in Alzheimer's disease pathogenesis. Journal of Neuroscience. 2010;30(45):14946–14954. doi: 10.1523/JNEUROSCI.4305-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lynch T, Cherny RA, Bush AI. Oxidative processes in Alzheimer’s disease: the role of Aβ-metal interactions. Experimental Gerontology. 2000;35(4):445–451. doi: 10.1016/s0531-5565(00)00112-1. [DOI] [PubMed] [Google Scholar]
- 28.Lan Z, Liu J, Chen L, et al. Danggui-Shaoyao-San ameliorates cognition deficits and attenuates oxidative stress-related neuronal apoptosis in d-galactose-induced senescent mice. Journal of Ethnopharmacology. 2012;141(1):386–395. doi: 10.1016/j.jep.2012.02.050. [DOI] [PubMed] [Google Scholar]
- 29.Chen P, Wang P, Shao G, Xiang L. Acetylcholinesterase inhibitory activities of 48 traditional Chinese medicinal herbs. Journal of Chinese Pharmaceutical Sciences. 2013;22(1):106–109. [Google Scholar]
- 30.Anderson JA, Coats JR. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pesticide Biochemistry and Physiology. 2012;102(2):124–128. [Google Scholar]