

Abstract
Neuropeptide Y has been shown to inhibit the immunological activity of reactive microglia in the rat cerebral cortex, to reduce N-methyl-D-aspartate current (INMDA) in cortical neurons, and protect neurons. In this study, after primary cultured microglia from the cerebral cortex of rats were treated with lipopolysaccharide, interleukin-1β and tumor necrosis factor-α levels in the cell culture medium increased, and mRNA expression of these cytokines also increased. After primary cultured cortical neurons were incubated with the lipopolysaccharide-treated microglial conditioned medium, peak INMDA in neurons increased. These effects of lipopolysaccharide were suppressed by neuropeptide Y. After addition of the neuropeptide Y Y1 receptor antagonist BIBP3226, the effects of neuropeptide Y completely disappeared. These results suggest that neuropeptide Y prevents excessive production of interleukin-1β and tumor necrosis factor-α by inhibiting microglial reactivity. This reduces INMDA in rat cortical neurons, preventing excitotoxicity, thereby protecting neurons.
Keywords: nerve regeneration, microglia, immunological activity, neuropeptide Y, interleukin-1β, tumor necrosis factor-α, INMDA, neural regeneration
Introduction
Microglia are a type of macrophage extensively distributed in the central nervous system (Ginhoux et al., 2010), and strongly associated with immunity. Microglia play major roles in nervous system disease and immune responses (Graeber and Streit, 1990; Aloisi, 1999; Wirenfeldt et al., 2011). Moreover, microglia can produce inflammatory factors and mediate inflammation in the nervous system (Rivest, 2009; Kraft and Harry, 2011). Under normal conditions, microglia are in a resting state and play an immune surveillance role in the central nervous system. When stimulated, microglia are activated, and can release interleukin-1β and tumor necrosis factor-α, as well as a large number of bioactive substances, including reactive oxygen, nitrogen and lipid species (Giulian et al., 1994; Liu et al., 2002; Rivest, 2009; Wu et al., 2009; Mayer et al., 2011). Excessive release of these bioactive substances can induce neuronal injury (Auvin et al., 2010; Friedman and Dingledine, 2011; Vezzani et al., 2011b). Microglial activation is a key step in the inflammatory process in the central nervous system (Hanisch, 2002; Kaur et al., 2010; Kraft and Harry, 2011). Animal experiments and clinical studies show that an inflammatory reaction occurs in the focal zone after epileptic seizures (Auvin et al., 2010; Vezzani and Friedman, 2011; Vezzani et al., 2013). Moreover, microglia are noticeably activated in the focal zone, and interleukin-1β and tumor necrosis factor-α expression are increased. Simultaneously, neuronal excitability is elevated, and the number of apoptotic neurons increases (Ravizza et al., 2005; Vezzani and Granata, 2005). In addition, inflammatory factors are strongly associated with epilepsy. Thus, reducing the release of inflammatory factors or blocking their effects could lessen the severity of epilepsy (Maroso et al., 2011; Vezzani et al., 2011a, b; Gao et al., 2012).
Previous studies have shown that some medicines protect neurons by inhibiting microglial reactivity (Tikka et al., 2001; Dheen et al., 2007; Park et al., 2007). For example, one study showed that after microglial activation was suppressed by the cyclooxygenase 2 inhibitor celecoxib in the hippocampus of epileptic rats, spontaneous seizures and neuronal injury were significantly reduced (Zhang et al., 2008), suggesting that inhibiting microglial activation and bioactive substance release is an effective method of treating epilepsy.
It remains poorly understood whether there was a kind of medicine that directly acted on neurons, regulated the immunological activity of microglia and finally protected neurons against injury through nerve-immunity pathway.
Neuropeptide Y is widely expressed in the central and peripheral nervous systems, and is strongly associated with epilepsy, learning and memory (Wettstein et al., 1995; Vezzani et al., 1999). Neuropeptide Y exerts protective effects against neuronal injury and can alleviate epilepsy (Smialowska et al., 2009; Baptista et al., 2012; Goncalves et al., 2012; Gotzsche et al., 2012; Malva et al., 2012). Neuropeptide Y exerts protective effects on neurons mainly through the Y2 receptor (Greber et al., 1994; Woldbye et al., 2010; Decressac et al., 2012; Goncalves et al., 2012). In a recent study, neuropeptide Y suppressed the activation of the mouse microglial cell line N9 induced by lipopolysaccharide, and it reduced the generation of interleukin-1β and nitric oxide (Ferreira et al., 2010). Neuropeptide Y was also found to suppress microglial migration and phagocytosis (Ferreira et al., 2011, 2012).
It is unclear whether neuropeptide Y can reduce the release of bioactive substances and protect neurons by regulating microglial immunological activity. Cytokines can activate glutamic acid receptors and induce changes in ion currents, thereby contributing to neuronal injury (Yang et al., 2005). Glutamic acid receptors include ionotropic and metabotropic receptors. The N-methyl-D-aspartate (NMDA) receptor, an ionotropic glutamic acid receptor, is extensively distributed in the central nervous system. The NMDA receptor can be controlled by membrane potential, glutamic acid and NMDA. In the resting state, the NMDA receptor binds to Mg2+, which blocks the opening of the ion channel. When bound to excitatory neurotransmitters such as glutamate and NMDA, the receptor is activated, opening the ion channel and allowing cations such as Ca2+, Na+ and K+ to enter, resulting in the NMDA current (INMDA). Excessive cation flux, especially Ca2+ influx, results in excitotoxic neuronal injury (Norris et al., 2006; Deshpande et al., 2008; Zhu et al., 2010).
Previous studies suggested that interleukin-1β and tumor necrosis factor-α elevate NMDA receptor expression, and increase NMDA activity (Balosso et al., 2008; Zheng et al., 2010; Zhu et al., 2010). Therefore, neuropeptide Y may diminish the release of interleukin-1β and tumor necrosis factor-α derived from microglia by inhibiting microglial activation, thereby suppressing NMDA receptor expression, decreasing NMDA activity, reducing INMDA, and ultimately protecting neurons.
In the present study, lipopolysaccharide was used to activate primary cultured microglia derived from the rat cerebral cortex. We observed the effects of neuropeptide Y on interleukin-1β and tumor necrosis factor-α in activated microglia. Microglial conditioned medium treated or untreated with neuropeptide Y was used to incubate primary neurons from the rat cerebral cortex. The patch-clamp technique was used to detect changes in INMDA induced by the different microglial conditioned media.
Materials and methods
Animals
A total of 64 clean Sprague-Dawley rats born within 24 hours were provided by the Experimental Animal Center of Hebei Medical University in China (animal license No. SCXK (Ji) 2013-1-03). The investigation conformed to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication No. 85-23, revised 1996), and the protocol was approved by the Institutional Animal Care Committee of Hebei Medical University.
Neuronal and microglial cultures
Mixed microglial cultures, microglial isolation and purification were conducted in accordance with a previous method (Nakajima et al., 1992). The brain was obtained aseptically from neonatal rats after craniotomy. After removal of the meninges and blood vessels, a part of the cerebral cortex was cut into pieces and digested in 0.125% trypsin at 37°C for 15 minutes. Samples were agitated with a pipette, centrifuged at 1,000 r/min for 5 minutes, and filtered. The supernatant was discarded. The pellet was resuspended with microglia medium (Dulbecco's modified Eagle's medium/F12 medium, DMEM) (Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (Gibco), 1 U/mL penicillin and 100 μg/mL streptomycin. The cell suspension was incubated in a 250 mL culture flask at 37°C, 5% CO2. On the second day, the medium was replaced once with new medium. From then on, half of the medium was replaced every 3 days. At 14 days, when cells were confluent, they were shaken on a rocker at 37°C, 180 r/min for 2 hours. The cell suspension was centrifuged at 1,000 r/min for 5 minutes. After removal of the supernatant, the samples were agitated into a cell suspension using microglia medium. Cells were adjusted to about 1 × 105/mL, and seeded in 6-well plates precoated with polylysine (Sigma, St. Louis, MO, USA), 3 mL in each well, in a constant-temperature incubator (Heraeus, Hanau, Germany) for 30 minutes without stirring. The medium was replaced once by new medium. After removal of oligodendrocytes, the samples were incubated with microglia medium for 3–5 days, resulting in a purified microglial culture.
Isolation and culture of rat cerebral cortical neurons were conducted in accordance with a previous method (Yang et al., 2006). The brain was obtained aseptically from neonatal rats by craniotomy. Brain tissue was placed in a petri dish with precooled DMEM/F12 medium. The petri dish was placed on ice. A median sagittal incision was made. After removal of the cerebellum and subcortical tissue, meninges and blood vessels were dissociated with a dissecting microscope. The cortex was placed in an additional petri dish with precooled DMEM/F12 medium. The petri dish was placed on ice. The cerebral cortex was cut into about 1 mm3 blocks, digested in 0.125% trypsin at 37°C for 5 minutes, and agitated into a cell suspension. Cell aggregates were removed by filtration to obtain a single-cell suspension. Cells were adjusted to about 1 × 105/mL using microglia medium. Cell suspensions were seeded in a 6-well plate with a polylysine-treated coverslip, 3 mL in each well, in a 5% CO2 incubator at 37°C for co-culture. Twenty-four hours later, the medium was replaced by neuronal medium (Neurobasal medium) (Gibco), supplemented with 2% B27 (Gibco), 100 U/mL penicillin and 100 μg/mL streptomycin. The medium was replaced once every 3 days. After 7–9 days of culture, purified neurons were obtained.
Identification of microglia and neurons
Purity of microglia was assessed using immunofluorescence staining (Schluesener et al., 1999; Yamasaki et al., 2010; Azemi et al., 2011; Savard et al., 2013; Yao et al., 2013). Isolated microglia were incubated in a culture plate with polylysine-treated slides for 3 days. The samples were washed with PBS, fixed in 4% paraformaldehyde for 30 minutes, treated with 0.3% Triton X-100 for 10 minutes, blocked with 3% goat serum for 30 minutes, incubated with mouse anti-ionized calcium-binding adapter molecule 1 monoclonal antibody (1:200; Sigma) at 4°C overnight, and then with FITC-labeled goat anti-mouse IgG (1:100; Proteintech, Chicago, IL, USA) at 37°C for 1 hour. Except for goat serum blocking, three PBS washes were performed between each step. After mounting, a fluorescence microscope (DMI3000B and DFC450C, Leica, Wetzlar, Germany) was used to observe the percentage of positive cells at 400 × magnification. Microglial morphology was observed after treatment with lipopolysaccharide (100 ng/mL; Sigma) for 6 hours.
Immunofluorescence staining was used to identify whether the cultured cells were neurons (Andreyeva et al., 2012; Soetedjo et al., 2013). The procedures were identical to that for microglia. The antibody used was rabbit anti-microtubule-associated protein-2 polyclonal antibody (1:100; Proteintech) and TRITC-labeled goat anti-rabbit IgG (1:100; Proteintech). Under a fluorescence microscope, five high-power fields (400 ×) were randomly selected. The percentage of positive cells was calculated with an Image-Pro Plus image analysis system (Media Cybernetics, Silver Spring, ML, USA).
Enzyme linked immunosorbent assay (ELISA) for interleukin-1β and tumor necrosis factor-α protein levels in lipopolysaccharide-treated microglial conditioned medium
Purified microglia were seeded in a 6-well polylysine-treated plate at 1 × 105/mL, 3 mL in each well. Three days later, microglia were cultured with fresh serum-free DMEM/F12 medium for 12 hours, and then incubated with serum-free DMEM/F12 medium containing lipopolysaccharide, 100 ng/mL (Sigma), for 1, 3, 6, 12 and 24 hours. Cell culture medium was collected from each group and centrifuged. Interleukin-1β and tumor necrosis factor-α levels in the medium were assessed with an ELISA kit (Bio-Swamp; Wuhan, China). Microtiter plates were assigned to the positive control well, blank control well and sample well. Standard preparations at different concentrations were added in the positive control well, 50 μL in each well. Blank control well did not contain any sample, reagent, biotinylated anti-tumor necrosis factor-α antibody or anti-interleukin-1β antibody. In the sample well, 40 μL of sample was first added, and then biotinylated anti-tumor necrosis factor-α antibody or anti-interleukin-1β antibody, 10 μL, was added. The plate was sealed with membrane, and then placed at 37°C for 30 minutes. The plate was washed five times, and patted dry. ELISA reagents, 50 μL, were added in each well, except the blank well. After incubation for 30 minutes, the plate was washed. The samples were visualized with developer at 37°C in the dark for 15 minutes. Reaction was terminated by the addition of stop buffer, 50 μL, in each well. Absorbance valus of each well were measured at 450 nm using a microplate reader (Thermo Labsystems, Helsinki, Finland). Blank control well was adjusted to zero. The measurement was performed within 15 mintues after adding stop buffer. Protein levels were calculated according to a standard curve of known concentrations.
Microglial treatment with neuropeptide Y and lipopolysaccharide
Microglia at 1 × 105/mL were seeded in a 6-well polylysine-treated plate, 3 mL in each well, for 3 days. Microglia were incubated with fresh serum-free DMEM/F12 medium for 12 hours to synchronize cells. Subsequently, cells were divided into control, lipopolysaccharide, neuropeptide Y + lipopolysaccharide, neuropeptide Y and BIBP3226 + neuropeptide Y + lipopolysaccharide groups. In the control group, microglia were incubated with serum-free DMEM/F12 medium for 6 hours. In the lipopolysaccharide group, microglia were incubated with serum-free medium containing 100 ng/mL lipopolysaccharide for 6 hours. In the neuropeptide Y + lipopolysaccharide group, microglia were incubated with serum-free DMEM/F12 medium containing neuropeptide Y (final concentration: 1 μmol/L; ENZO, New York, NY, USA) for 0.5 hours, and then treated with lipopolysaccharide (final concentration: 100 ng/mL) for 6 hours. In the neuropeptide Y group, microglia were incubated with serum-free medium containing neuropeptide Y (1 μmol/L) for 6 hours. In the BIBP3226 + neuropeptide Y + lipopolysaccharide group, microglia were first incubated with serum-free DMEM/F12 medium containing the neuropeptide Y1 receptor antagonist BIBP3226 (N-[(1R)]-4-[(Aminoiminomethyl)amino-1-[[[(4-hydroxyphenyl)methyl]amino]carbonyl]butyl-α-henylbenzeneacetamide trifluoroacetate; molecular weight: 587.59; molecular formula: C27 H31 N5 O3•C F3 CO2 H; purity > 98%; final concentration: 1 μmol/L; Tocris Bioscience, Ellisville, MO, USA) for 0.5 hours, then with neuropeptide Y (final concentration: 1 μmol/L) for 0.5 hours, and finally with lipopolysaccharide (final concentration: 100 ng/mL) for 6 hours.
Effects of neuropeptide Y on interleukin-1β and tumor necrosis factor-α mRNA levels in lipopolysaccharide-treated microglia, as detected by real-time PCR
Culture medium was discarded and Trizol (1 mL/well) was added in each well. Samples were agitated and placed in an RNAse-free centrifuge tube for 5 minutes. Into each tube, 0.2 mL chloroform was added. After shaking for 15 seconds and incubating for 5 minutes, samples were centrifuged at 12,000 r/min for 15 mintues. The colorless liquid in the upper layer was removed to a new centrifuge tube. An equal volume of isopropanol was added, and mixed by inversion, followed by centrifugation at 12,000 r/min for 10 minutes. A white precipitate was visible at the bottom of the tube. Supernatant was completely discarded. The precipitate, following addition of 1 mL 75% alcohol (in DEPC-treated water), was washed. After centrifugation at 7,500 r/min at 4°C for 5 minutes, the supernatant was discarded. The samples were air-dried for 3–5 minutes. 20–30 μL of DEPC-treated water was added to thoroughly dissolve RNA. To verify RNA integrity and purity, 1 % agarose gel electrophoresis was used. A UV spectrophotometer was used to determine RNA concentration. Primers were synthesized by Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China.
Primer sequences were shown in Table 1. Reverse transcription reaction: Total RNA 8 μL, random primer 1 μL, 2 × ES reaction mix 10 μL, RT/RI enzyme mix 1 μL; total volume: 20 μL. Amplification system: 2 × UltraSYBR mixture (with ROX) 10 μL, forward primer (10 μmol/L) 1 μL, reverse primer (10 μmol/L) 1 μL, cDNA 8 μL, total volume: 20 μL. RT-PCR parameters: predenaturation at 95°C for 10 minutes; 40 cycles of denaturation at 95°C for 15 seconds, annealing at 58°C for 20 seconds, extension at 72°C for 27 seconds. An ABI 7300 Real-Time PCR System (Applied Biosystems, Foster, CA, USA) was used to detect and calculate the relative quantity of the target gene to the internal reference (GAPDH).
Table 1.
Primer sequences for interleukin-β and tumor necrosis factor-α transcript amplification
INMDA in neurons as determined by patch-clamp technique
Microglial conditioned media obtained from the various groups were individually mixed with neuronal medium at a ratio of 1:1. Control medium after mixing with neuronal medium at 1:1 was divided into three groups: control (CON) group, CON + lipopolysaccharide group and CON + neuropeptide Y group. Lipopolysaccharide (final concentration of 50 ng/mL) and neuropeptide Y (final concentration of 0.5 μmol/L) were added in the CON + lipopolysaccharide group and CON + neuropeptide Y group, respectively. Purified neuronal cells (3 × 105/well) in each group were incubated with their corresponding treatment for 6 hours. INMDA in neurons was recorded using the patch-clamp technique (Yang et al., 2005; Zhang et al., 2011). Glass slides with neuronal cells were placed in a 0.3 mL-bath. Cells were continuously perfused with extracellular fluid at 2 mL/min to ensure that fluid exchange was finished within 2 minutes. An inverted microscope (TE2000-S; Nikon, Tokyo, Japan) was used to observe cells. Neurons with a clear outline, a defined three-dimensional structure and a smooth surface were selected for recording. Using a three-dimensional manipulator (Sutter Instruments Company, Novato, CA), a glass microelectrode at 1–3 MΩ impedance and filled with pipette solution (including KCl 10 mmol/L, K-D-gluconate 120 mmol/L, HEPES 10 mmol/L, EGTA 11 mmol/L, NaCl 5 mmol/L, MgCl2 2 mmol/L, CaCl2 1 mmol/L, Mg-ATP 2 mmol/L and Li-GTP 1 mmol/L; pH 7.2 adjusted with NaOH) was sealed onto the cell surface. Negative pressure was applied until the patch was formed. Capacitive current and series resistance of the electrode were compensated, and whole-cell recording was commenced. Clamping voltage was set at –60 mV. After whole-cell recording for 5 minutes, the intracellular fluid was thoroughly replaced with electrode solution. At this time, 100 μmol/L NMDA (Sigma) and 10 μmol/L glycine (Sigma) were added extracellularly to induce inward current changes. INMDA was recorded.
Statistical analysis
The data were analyzed using SPSS 10.0 software (SPSS, Chicago, IL, USA) and expressed as mean ± SD. Test of normality and homogeneity test for variance showed that all data obeyed a normal distribution and had homogeneous variance. Completely random design was employed in analysis of variance. Paired comparison was done using least significant difference test. A value of P < 0.05 was considered statistically significant.
Results
Isolated culture of microglia and neurons
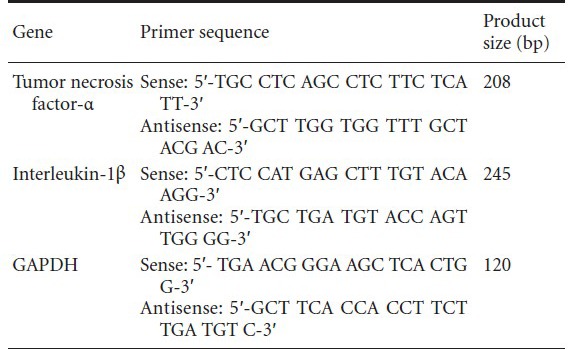
Ionized calcium-binding adapter molecule and microtubule-associated protein 2 were used as markers of microglia and neurons, respectively (Izant and McIntosh, 1980; Dehmelt and Halpain, 2005; Nakamura et al., 2013). The purities of microglia and neurons were > 95%, which met the requirements of the experiments (Figure 1).
Figure 1.
Primary cultured microglia and neurons (immunofluorescence, × 400).
(A) Microglia in the control (lipopolysaccharide-untreated) group had a ramified appearance, a small body, thin long processes, and expressed ion-ized calcium-binding adapter molecule 1 (IBA-1) (arrow); (B) after treatment with lipopolysaccharide, microglia were in an activated state, with a round amoeboid body, retracted processes, and darkly stained with IBA-1 (arrow); (C) neurons extended axons and long dendrites and expressed microtubule-associated protein 2 (arrow).
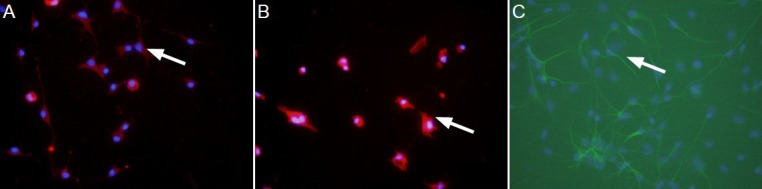
Lipopolysaccharide promoted the secretion of interleukin-1β and tumor necrosis factor-α by microglia
ELISA results revealed that interleukin-1β and tumor necrosis factor-α levels in the medium of each group were significantly greater after incubation with lipopolysaccharide compared with the control group (P < 0.01 or P < 0.05). As incubation proceded, interleukin-1β content gradually increased (P < 0.05), peaked at 3 hours, and then gradually decreased (P < 0.05; Figure 2A). Moreover, tumor necrosis factor-α content gradually increased over the incubation period (P < 0.05), peaked at 6 hours, and then gradually diminished (P < 0.05; Figure 2B).
Figure 2.
Effects of lipopolysaccharide on IL-1β and TNF-α secretion by microglia (enzyme linked immunosorbent assay).
(A) IL-1β levels in medium after incubation with 100 ng/mL lipopolysaccharide. (B) TNF-α levels in medium after incubation with 100 ng/mL lipopolysaccharide. Data are expressed as mean ± SD. The experiment was performed in triplicate. Completely random design was employed in analysis of variance. Paired comparison was done using least significant difference test. *P < 0.05, **P < 0.01, vs. control group; #P < 0.05, vs. the previous time point. 1, 3, 6, 12, 24 h: 1, 3, 6, 12, 24 hours after incubation with lipopolysaccharide. IL-1β: Interleukin-1β; TNF-α: tumor necrosis factor-α.
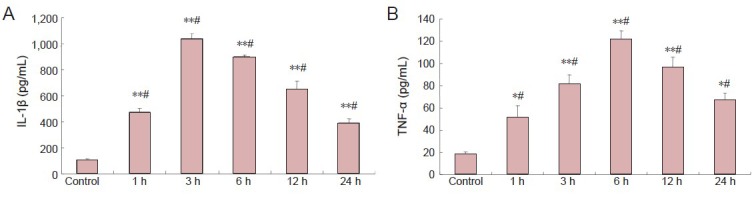
Neuropeptide Y reduced lipopolysaccharide-induced secretion of interleukin-1β and tumor necrosis factor-α by microglia
ELISA results revealed that interleukin-1β and tumor necrosis factor-α levels were significantly lower in the neuropeptide Y + lipopolysaccharide group than in the lipopolysaccharide group (P < 0.01). Interleukin-1β and tumor necrosis factor-α levels were significantly increased in the BIBP3226 + neuropeptide Y + lipopolysaccharide group (P < 0.01). Interleukin-1β and tumor necrosis factor-α levels did not significantly change in the medium after normal cells were treated with neuropeptide Y (P > 0.05; Figure 3).
Figure 3.
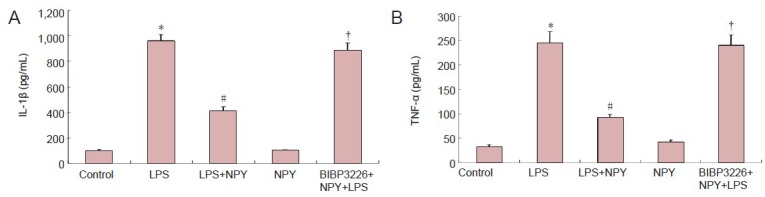
Inhibitory effects of NPY on LPS-induced production of IL-1β and TNF-α by microglia (enzyme linked immunosorbent assay).
IL-1β (A) and TNF-α (B) levels in microglial conditioned medium after culture in 1 μmol/L BIBP3226, 1 μmol/L NPY and 100 ng/mL LPS. Data are expressed as mean ± SD. The experiment was performed in triplicate. Completely random design was employed in analysis of variance. Paired comparison was done using least significant difference test. *P < 0.01, vs. control group; #P < 0.01, vs. LPS group; †P < 0.01, vs. LPS + NPY group. NPY: Neuropeptide Y; LPS: lipopolysaccharide; IL-1β: interleukin-1β; TNF-α: tumor necrosis factor-α.
Neuropeptide Y suppressed interleukin-1β and tumor necrosis factor-α mRNA expression in microglia treated with lipopolysaccharide
At 6 hours after incubation, interleukin-1β and tumor necrosis factor-α mRNA levels were significantly higher in the lipopolysaccharide group and in the BIBP3226 + neuropeptide Y + lipopolysaccharide group than in the control group (P < 0.01). No significant difference in interleukin-1β or tumor necrosis factor-α mRNA levels was detected between the lipopolysaccharide and BIBP3226 + neuropeptide Y + lipopolysaccharide groups. Interleukin-1β and tumor necrosis factor-α mRNA levels were significantly lower in the lipopolysaccharide + neuropeptide Y group compared with the lipopolysaccharide group (P < 0.01). Interleukin-1β and tumor necrosis factor-α mRNA levels were significantly higher in the BIBP3226 + neuropeptide Y + lipopolysaccharide group than in the lipopolysaccharide + neuropeptide Y group (P < 0.01). Interleukin-1β and tumor necrosis factor-α mRNA levels were similar in the neuropeptide Y and control groups (P > 0.05). These results suggest that lipopolysaccharide elevates interleukin-1β and tumor necrosis factor-α mRNA levels in microglia and that neuropeptide Y inhibits the effects of lipopolysaccharide. When BIBP3226 was used to block the neuropeptide Y1 receptor, neuropeptide Y had no effect. Neuropeptide Y did not impact interleukin-1β or tumor necrosis factor-α mRNA levels in microglia (Figure 4).
Figure 4.
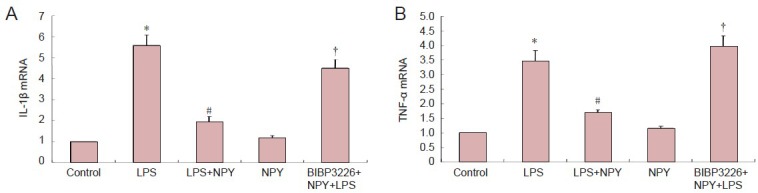
Inhibitory effects of NPY on LPS-induced expression of IL-1β and TNF-α by microglia (real-time PCR).
(A) IL-1β mRNA levels in microglia cultured in 1 μmol/L BIBP3226, 1 μmol/L NPY and 100 ng/mL LPS. (B) TNF-α mRNA levels in microglia cul-tured in 1 μmol/L BIBP3226, 1 μmol/L NPY and 100 ng/mL LPS. The ratio of target gene expression in the control group to the internal reference GAPDH was 1. Data are expressed as mean ± SD. The experiment was performed in triplicate. Completely random design was employed in analysis of variance. Paired comparison was done using least significant difference test. *P < 0.01, vs. control group; #P < 0.01, vs. LPS group; †P < 0.01, vs. LPS + NPY group. NPY: Neuropeptide Y; LPS: lipopolysaccharide; IL-1β: interleukin-1β; TNF-α: tumor necrosis factor-α.
Neuropeptide Y inhibits INMDA in neurons after incubation with lipopolysaccharide-treated microglial conditioned medium
Patch-clamp recording showed that, compared with the CON group, INMDA was higher in the lipopolysaccharide group (P < 0.01). INMDA was significantly lower in the neuropeptide Y + lipopolysaccharide group compared with the lipopolysaccharide group. INMDA was significantly higher in the BIBP3226 + neuropeptide Y + lipopolysaccharide group compared with the neuropeptide Y + lipopolysaccharide group (P < 0.01). INMDA was not substantially altered in neurons cultured in normal microglia medium (Figure 5).
Figure 5.
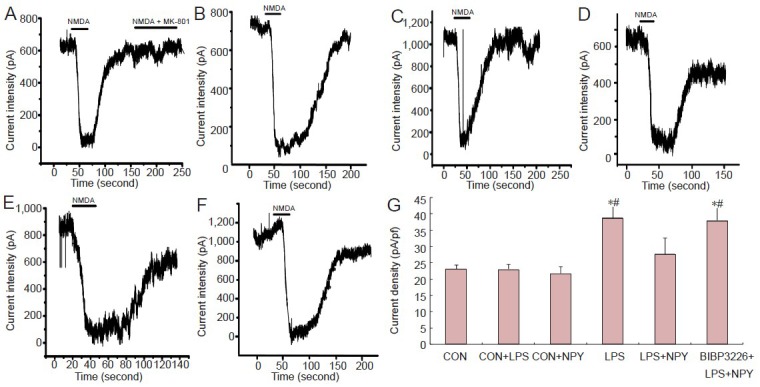
Effects of NPY on INMDA in neurons incubated with LPS-treated microglial conditioned medium.
(A) CON group; (B) CON + LPS group; (C) LPS group; (D) CON + NPY group; (E) LPS + NPY group; (F) BIBP3226 + NPY + LPS group. NMDA and glycine were simultaneously added to the perfusate to activate the NMDA receptor. After whole-cell recording for 5 minutes, intracel-lular fluid was thoroughly replaced by interelectrode solution. At this time, 100 μmol/L NMDA and 10 μmol/L glycine were added extracellularly to induce inward current changes. INMDA was measured. After eluting NMDA and glycine, the membrane current was restored to normal. After treatment with 100 μmol/L NMDA, 10 μmol/L glycine and 10 μmol/L NMDA receptor antagonist MK-801, this inward current disappeared. Therefore, the recorded current was INMDA. (G) Peak INMDA after incubation with microglial conditioned medium. Data are expressed as mean ± SD. The experiment was performed in triplicate. Completely random design was employed in analysis of variance. Paired comparison was done using least significant difference test. *P < 0.01, vs. CON group; #P < 0.01, vs. LPS + NPY group. NPY: Neuropeptide Y; LPS: lipopolysaccharide; INMDA: N-methyl-D-aspartate current; NMDA: N-methyl-D-aspartate; CON: control.
Discussion
Microglia play an important role in regulating immune function and are strongly associated with the occurrence and development of central nervous system diseases (Harry and Kraft, 2012; Zhao et al., 2013). In central nervous system pathologies, microglia rapidly respond and switch from a resting to an activated state, from a ramified to an amoeboid morphology, with an enlarged body and retracted processes (Harry and Kraft, 2012). Microglia undergo a series of changes, and function as sensors in the central nervous system (Kreutzberg, 1996). Microglias are the macrophages of the central nervous system, and are strongly associated with immune functions in brain tissue. The accumulation of numerous cytokines and bioactive substances after microglial activation is a key event in neuronal injury (Vezzani et al., 2011b). Among these cytokines, interleukin-1β and tumor necrosis factor-α play important roles in physiological and pathological processes in the central nervous system (Lee et al., 2004; Maroso et al., 2011; Savard et al., 2013). The increased concentrations of interleukin-1β and tumor necrosis factor-α in brain tissues exert toxic effects on surrounding neurons (Zheng et al., 2010; Zhu et al., 2010). Lipopolysaccharide-activated microglia produce various factors implicated in pathologies (Lee et al., 2004).
Lipopolysaccharide was used as an activator of microglia in the present study. After 6 hours of intervention with lipopolysaccharide, the immunological activity of microglia increased, and interleukin-1β and tumor necrosis factor-α levels in microglial conditioned medium substantially increased. Neuropeptide Y suppressed the lipopolysaccharide-induced activation of microglia, and it reduced interleukin-1β and tumor necrosis factor-α protein and mRNA levels in these cells. These inhibitory effects of neuropeptide Y were completely suppressed by BIBP3226, indicating that neuropeptide Y diminishes the immunological activity of microglia through the Y1 receptor, thereby reducing interleukin-1β and tumor necrosis factor-α generation, consistent with a previous study (Ferreira et al., 2010). Many studies on the effects of cytokines on neurons suggest that these factors modulate the glutamic acid receptor (Vezzani et al., 2008). For example, interleukin-1β and tumor necrosis factor-α elevate NMDA receptor expression and activity. A previous study demonstrated that interleukin-1β can phosphorylate the NMDA NR2B subunit and activate the receptor (Balosso et al., 2008). Tumor necrosis factor-α increases phosphorylation of the NR1 subunit, and increases its expression as well (Wheeler et al., 2009). In a previous study, after rat hippocampal neurons were incubated with microglial conditioned medium containing tumor necrosis factor-α, NMDARI expression rose in neurons, and this effect was inhibited by addition of an anti-tumor necrosis factor-α antibody (Zhu et al., 2010). Interleukin-1β and tumor necrosis factor-α increase the probability and duration of channel opening, thereby increasing INMDA. The resulting Ca2+ influx into neurons causes calcium overload, resulting in cytotoxicity (Viviani et al., 2003). INMDA is a direct index of changes in the NMDA receptor. An increase in INMDA indicates increased activity of the NMDA receptor and an increased number of cations (mainly Ca2+) passing through the ion channel (Malenka and Nicoll, 1999; Gardoni et al., 2001; Yang et al., 2005).
Patch-clamping showed that INMDA was enhanced after treatment with lipopolysaccharide compared with the control group. Neuropeptide Y suppressed microglial activation and this lipopolysaccharide-mediated increase in INMDA. When BIBP3226 was added to block the neuropeptide Y Y1 receptor, this effect of neuropeptide Y disappeared. INMDA did not remarkably change after incubation with lipopolysaccharide and neuropeptide Y conditioned medium, suggesting that changes in INMDA in neurons were not caused by lipopolysaccharide or neuropeptide Y, similar to a previous study (Bronstein et al., 1995), but were induced by factors in the conditioned medium of activated microglia.
In summary, neuropeptide Y suppresses microglial reactivity, reduces the release of microglial bioactive factors, diminishes INMDA, and prevents excitotoxicity induced by excessive Ca2+ entry into neurons. These effects of neuropeptide Y were fully blocked by the Y1 receptor antagonist BIBP3226. We demonstrated that neuropeptide Y can reduce INMDA in neurons by regulating the immune function of microglia. Thus, neuropeptide Y plays a major role in regulating immune function in the central nervous system. Neuropeptide Y reduces the generation of interleukin-1β and tumor necrosis factor-α, preventing intracellular Ca2+ overload induced by NMDA receptor hyperactivation, thereby protecting neurons.
Footnotes
Conflicts of interest: None declared.
Copyedited by Patel B, Rave W, Yu J, Qiu Y, Li CH, Song LP, Zhao M
References
- 1.Aloisi F. The role of microglia and astrocytes in CNS immune surveillance and immunopathology. Adv Exp Med Biol. 1999;468:123–133. doi: 10.1007/978-1-4615-4685-6_10. [DOI] [PubMed] [Google Scholar]
- 2.Andreyeva A, Nieweg K, Horstmann K, Klapper S, Muller-Schiffmann A, Korth C, Gottmann K. C-terminal fragment of N-cadherin accelerates synapse destabilization by amyloid-beta . Brain. 2012;135:2140–2154. doi: 10.1093/brain/aws120. [DOI] [PubMed] [Google Scholar]
- 3.Auvin S, Mazarati A, Shin D, Sankar R. Inflammation enhances epileptogenesis in the developing rat brain. Neurobiol Dis. 2010;40:303–310. doi: 10.1016/j.nbd.2010.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Azemi E, Lagenaur CF, Cui XT. The surface immobilization of the neural adhesion molecule L1 on neural probes and its effect on neuronal density and gliosis at the probe/tissue interface. Biomaterials. 2011;32:681–692. doi: 10.1016/j.biomaterials.2010.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Balosso S, Maroso M, Sanchez-Alavez M, Ravizza T, Frasca A, Bartfai T, Vezzani A. A novel non-transcriptional pathway mediates the proconvulsive effects of interleukin-1beta. Brain. 2008;131:3256–3265. doi: 10.1093/brain/awn271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Baptista S, Bento AR, Goncalves J, Bernardino L, Summavielle T, Lobo A, Fontes-Ribeiro C, Malva JO, Agasse F, Silva AP. Neuropeptide Y promotes neurogenesis and protection against methamphetamine-induced toxicity in mouse dentate gyrus-derived neurosphere cultures. Neuropharmacology. 2012;62:2413–2423. doi: 10.1016/j.neuropharm.2012.02.015. [DOI] [PubMed] [Google Scholar]
- 7.Bronstein DM, Perez-Otano I, Sun V, Mullis Sawin SB, Chan J, Wu GC, Hudson PM, Kong LY, Hong JS, McMillian MK. Glia-dependent neurotoxicity and neuroprotection in mesencephalic cultures. Brain Res. 1995;704:112–116. doi: 10.1016/0006-8993(95)01189-7. [DOI] [PubMed] [Google Scholar]
- 8.Decressac M, Pain S, Chabeauti PY, Frangeul L, Thiriet N, Herzog H, Vergote J, Chalon S, Jaber M, Gaillard A. Neuroprotection by neuropeptide Y in cell and animal models of Parkinson's disease. Neurobiol Aging. 2012;33:2125–2137. doi: 10.1016/j.neurobiolaging.2011.06.018. [DOI] [PubMed] [Google Scholar]
- 9.Dehmelt L, Halpain S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2005;6:204. doi: 10.1186/gb-2004-6-1-204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Deshpande LS, Lou JK, Mian A, Blair RE, Sombati S, Attkisson E, De-Lorenzo RJ. Time course and mechanism of hippocampal neuronal death in an in vitro model of status epilepticus: role of NMDA receptor activation and NMDA dependent calcium entry. Eur J Pharmacol. 2008;583:73–83. doi: 10.1016/j.ejphar.2008.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dheen ST, Kaur C, Ling EA. Microglial activation and its implications in the brain diseases. Curr Med Chem. 2007;14:1189–1197. doi: 10.2174/092986707780597961. [DOI] [PubMed] [Google Scholar]
- 12.Ferreira R, Xapelli S, Santos T, Silva AP, Cristovao A, Cortes L, Malva JO. Neuropeptide Y modulation of interleukin-1{beta} (IL-1{beta})-induced nitric oxide production in microglia. J Biol Chem. 2010;285:41921–41934. doi: 10.1074/jbc.M110.164020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ferreira R, Santos T, Viegas M, Cortes L, Bernardino L, Vieira OV, Malva JO. Neuropeptide Y inhibits interleukin-1beta-induced phagocytosis by microglial cells. J Neuroinflammation. 2011;8:169. doi: 10.1186/1742-2094-8-169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ferreira R, Santos T, Cortes L, Cochaud S, Agasse F, Silva AP, Xapelli S, Malva JO. Neuropeptide Y inhibits interleukin-1 beta-induced microglia motility. J Neurochem. 2012;120:93–105. doi: 10.1111/j.1471-4159.2011.07541.x. [DOI] [PubMed] [Google Scholar]
- 15.Friedman A, Dingledine R. Molecular cascades that mediate the influence of inflammation on epilepsy. Epilepsia. 2011;52(Suppl 3):33–39. doi: 10.1111/j.1528-1167.2011.03034.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gao F, Liu Y, Li X, Wang Y, Wei D, Jiang W. Fingolimod (FTY720) inhibits neuroinflammation and attenuates spontaneous convulsions in lithium-pilocarpine induced status epilepticus in rat model. Pharmacol Biochem Behav. 2012;103:187–196. doi: 10.1016/j.pbb.2012.08.025. [DOI] [PubMed] [Google Scholar]
- 17.Gardoni F, Schrama LH, Kamal A, Gispen WH, Cattabeni F, Di Luca M. Hippocampal synaptic plasticity involves competition between Ca2+/calmodulin-dependent protein kinase II and postsynaptic density 95 for binding to the NR2A subunit of the NMDA receptor. J Neurosci. 2001;21:1501–1509. doi: 10.1523/JNEUROSCI.21-05-01501.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ginhoux F, Greter M, Leboeuf M, Nandi S, See P, Gokhan S, Mehler MF, Conway SJ, Ng LG, Stanley ER, Samokhvalov IM, Merad M. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science. 2010;330:841–845. doi: 10.1126/science.1194637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Giulian D, Li J, Leara B, Keenen C. Phagocytic microglia release cytokines and cytotoxins that regulate the survival of astrocytes and neurons in culture. Neurochem Int. 1994;25:227–233. doi: 10.1016/0197-0186(94)90066-3. [DOI] [PubMed] [Google Scholar]
- 20.Goncalves J, Ribeiro CF, Malva JO, Silva AP. Protective role of neuropeptide Y Y(2) receptors in cell death and microglial response following methamphetamine injury. Eur J Neurosci. 2012;36:3173–3183. doi: 10.1111/j.1460-9568.2012.08232.x. [DOI] [PubMed] [Google Scholar]
- 21.Gotzsche CR, Nikitidou L, Sorensen AT, Olesen MV, Sorensen G, Christiansen SH, Angehagen M, Woldbye DP, Kokaia M. Combined gene overexpression of neuropeptide Y and its receptor Y5 in the hippocampus suppresses seizures. Neurobiol Dis. 2012;45:288–296. doi: 10.1016/j.nbd.2011.08.012. [DOI] [PubMed] [Google Scholar]
- 22.Graeber MB, Streit WJ. Microglia: immune network in the CNS. Brain Pathol. 1990;1:2–5. doi: 10.1111/j.1750-3639.1990.tb00630.x. [DOI] [PubMed] [Google Scholar]
- 23.Greber S, Schwarzer C, Sperk G. Neuropeptide Y inhibits potassium-stimulated glutamate release through Y2 receptors in rat hippocampal slices in vitro. Br J Pharmacol. 1994;113:737–740. doi: 10.1111/j.1476-5381.1994.tb17055.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hanisch UK. Microglia as a source and target of cytokines. Glia. 2002;40:140–155. doi: 10.1002/glia.10161. [DOI] [PubMed] [Google Scholar]
- 25.Harry GJ, Kraft AD. Microglia in the developing brain: a potential target with lifetime effects. Neurotoxicology. 2012;33:191–206. doi: 10.1016/j.neuro.2012.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Izant JG, McIntosh JR. Microtubule-associated proteins: a monoclonal antibody to MAP2 binds to differentiated neurons. Proc Natl Acad Sci U S A. 1980;77:4741–4745. doi: 10.1073/pnas.77.8.4741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kaur G, Han SJ, Yang I, Crane C. Microglia and central nervous system immunity. Neurosurg Clin N Am. 2010;21:43–51. doi: 10.1016/j.nec.2009.08.009. [DOI] [PubMed] [Google Scholar]
- 28.Kraft AD, Harry GJ. Features of microglia and neuroinflammation relevant to environmental exposure and neurotoxicity. Int J Environ Res Public Health. 2011;8:2980–3018. doi: 10.3390/ijerph8072980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kreutzberg GW. Microglia: a sensor for pathological events in the CNS. Trends Neurosci. 1996;19:312–318. doi: 10.1016/0166-2236(96)10049-7. [DOI] [PubMed] [Google Scholar]
- 30.Lee SM, Yune TY, Kim SJ, Kim YC, Oh YJ, Markelonis GJ, Oh TH. Minocycline inhibits apoptotic cell death via attenuation of TNF-alpha expression following iNOS/NO induction by lipopolysaccharide in neuron/glia co-cultures. J Neurochem. 2004;91:568–578. doi: 10.1111/j.1471-4159.2004.02780.x. [DOI] [PubMed] [Google Scholar]
- 31.Liu B, Gao HM, Wang JY, Jeohn GH, Cooper CL, Hong JS. Role of nitric oxide in inflammation-mediated neurodegeneration. Ann N Y Acad Sci. 2002;962:318–331. doi: 10.1111/j.1749-6632.2002.tb04077.x. [DOI] [PubMed] [Google Scholar]
- 32.Malenka RC, Nicoll RA. Long-term potentiation--a decade of progress. Science. 1999;285:1870–1874. doi: 10.1126/science.285.5435.1870. [DOI] [PubMed] [Google Scholar]
- 33.Malva JO, Xapelli S, Baptista S, Valero J, Agasse F, Ferreira R, Silva AP. Multifaces of neuropeptide Y in the brain--neuroprotection, neurogenesis and neuroinflammation. Neuropeptides. 2012;46:299–308. doi: 10.1016/j.npep.2012.09.001. [DOI] [PubMed] [Google Scholar]
- 34.Maroso M, Balosso S, Ravizza T, Iori V, Wright CI, French J, Vezzani A. Interleukin-1beta biosynthesis inhibition reduces acute seizures and drug resistant chronic epileptic activity in mice. Neurotherapeutics. 2011;8:304–315. doi: 10.1007/s13311-011-0039-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mayer AM, Clifford JA, Aldulescu M, Frenkel JA, Holland MA, Hall ML, Glaser KB, Berry J. Cyanobacterial Microcystis aeruginosa lipopolysaccharide elicits release of superoxide anion, thromboxane B(2), cytokines, chemokines, and matrix metalloproteinase-9 by rat microglia. Toxicol Sci. 2011;121:63–72. doi: 10.1093/toxsci/kfr045. [DOI] [PubMed] [Google Scholar]
- 36.Nakajima K, Takemoto N, Kohsaka S. Retinoic acid enhances the secretion of plasminogen from cultured rat microglia. FEBS Lett. 1992;314:167–170. doi: 10.1016/0014-5793(92)80966-k. [DOI] [PubMed] [Google Scholar]
- 37.Nakamura R, Nishimura T, Ochiai T, Nakada S, Nagatani M, Ogasawara H. Availability of a microglia and macrophage marker, Iba-1, for differential diagnosis of spontaneous malignant reticuloses from astrocytomas in rats. J Toxicol Pathol. 2013;26:55. doi: 10.1293/tox.26.55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Norris CM, Blalock EM, Thibault O, Brewer LD, Clodfelter GV, Porter NM, Landfield PW. Electrophysiological mechanisms of delayed excitotoxicity: positive feedback loop between NMDA receptor current and depolarization-mediated glutamate release. J Neurophysiol. 2006;96:2488–2500. doi: 10.1152/jn.00593.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Park YK, Chung YS, Kim YS, Kwon OY, Joh TH. Inhibition of gene expression and production of iNOS and TNF-alpha in LPS-stimulated microglia by methanol extract of Phellodendri cortex. Int Immunopharmacol. 2007;7:955–962. doi: 10.1016/j.intimp.2006.03.018. [DOI] [PubMed] [Google Scholar]
- 40.Ravizza T, Rizzi M, Perego C, Richichi C, Veliskova J, Moshe SL, De Simoni MG, Vezzani A. Inflammatory response and glia activation in developing rat hippocampus after status epilepticus. Epilepsia. 2005;46(Suppl 5):113–117. doi: 10.1111/j.1528-1167.2005.01006.x. [DOI] [PubMed] [Google Scholar]
- 41.Rivest S. Regulation of innate immune responses in the brain. Nat Rev Immunol. 2009;9:429–439. doi: 10.1038/nri2565. [DOI] [PubMed] [Google Scholar]
- 42.Savard A, Lavoie K, Brochu ME, Grbic D, Lepage M, Gris D, Sebire G. Involvement of neuronal IL-1beta in acquired brain lesions in a rat model of neonatal encephalopathy. J Neuroinflammation. 2013;10:110. doi: 10.1186/1742-2094-10-110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Schluesener HJ, Seid K, Meyermann R. Effects of autoantigen and dexamethasone treatment on expression of endothelial-monocyte activating polypeptide II and allograft-inflammatory factor-1 by activated macrophages and microglial cells in lesions of experimental autoimmune encephalomyelitis, neuritis and uveitis. Acta Neuropathol. 1999;97:119–126. doi: 10.1007/s004010050964. [DOI] [PubMed] [Google Scholar]
- 44.Smialowska M, Domin H, Zieba B, Kozniewska E, Michalik R, Piotrowski P, Kajta M. Neuroprotective effects of neuropeptide Y-Y2 and Y5 receptor agonists in vitro and in vivo. Neuropeptides. 2009;43:235–249. doi: 10.1016/j.npep.2009.02.002. [DOI] [PubMed] [Google Scholar]
- 45.Soetedjo L, Glover DA, Jin H. Targeting of vasoactive intestinal peptide receptor 2, VPAC2, a secretin family G-protein coupled receptor, to primary cilia. Biol Open. 2013;2:686–694. doi: 10.1242/bio.20134747. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 46.Tikka T, Fiebich BL, Goldsteins G, Keinanen R, Koistinaho J. Minocycline a tetracycline derivative, is neuroprotective against excitotoxicity by inhibiting activation and proliferation of microglia. J Neurosci. 2001;21:2580–2588. doi: 10.1523/JNEUROSCI.21-08-02580.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Vezzani A, Granata T. Brain inflammation in epilepsy: experimental and clinical evidence. Epilepsia. 2005;46:1724–1743. doi: 10.1111/j.1528-1167.2005.00298.x. [DOI] [PubMed] [Google Scholar]
- 48.Vezzani A, Friedman A. Brain inflammation as a biomarker in epilepsy. Biomark Med. 2011;5:607–614. doi: 10.2217/bmm.11.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Vezzani A, Sperk G, Colmers WF. Neuropeptide Y: emerging evidence for a functional role in seizure modulation. Trends Neurosci. 1999;22:25–30. doi: 10.1016/s0166-2236(98)01284-3. [DOI] [PubMed] [Google Scholar]
- 50.Vezzani A, Balosso S, Ravizza T. The role of cytokines in the pathophysiology of epilepsy. Brain Behav Immun. 2008;22:797–803. doi: 10.1016/j.bbi.2008.03.009. [DOI] [PubMed] [Google Scholar]
- 51.Vezzani A, French J, Bartfai T, Baram TZ. The role of inflammation in epilepsy. Nat Rev Neurol. 2011a;7:31–40. doi: 10.1038/nrneurol.2010.178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Vezzani A, Aronica E, Mazarati A, Pittman QJ. Epilepsy and brain inflammation. Exp Neurol. 2013;244:11–21. doi: 10.1016/j.expneurol.2011.09.033. [DOI] [PubMed] [Google Scholar]
- 53.Vezzani A, Maroso M, Balosso S, Sanchez MA, Bartfai T. IL-1 receptor/Toll-like receptor signaling in infection, inflammation, stress and neurodegeneration couples hyperexcitability and seizures. Brain Behav Immun. 2011b;25:1281–1289. doi: 10.1016/j.bbi.2011.03.018. [DOI] [PubMed] [Google Scholar]
- 54.Viviani B, Bartesaghi S, Gardoni F, Vezzani A, Behrens MM, Bartfai T, Binaglia M, Corsini E, Di Luca M, Galli CL, Marinovich M. Interleukin-1beta enhances NMDA receptor-mediated intracellular calcium increase through activation of the Src family of kinases. J Neurosci. 2003;23:8692–8700. doi: 10.1523/JNEUROSCI.23-25-08692.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wettstein JG, Earley B, Junien JL. Central nervous system pharmacology of neuropeptide Y. Pharmacol Ther. 1995;65:397–414. doi: 10.1016/0163-7258(95)98598-k. [DOI] [PubMed] [Google Scholar]
- 56.Wheeler D, Knapp E, Bandaru VV, Wang Y, Knorr D, Poirier C, Mattson MP, Geiger JD, Haughey NJ. Tumor necrosis factor-alpha-induced neutral sphingomyelinase-2 modulates synaptic plasticity by controlling the membrane insertion of NMDA receptors. J Neurochem. 2009;109:1237–1249. doi: 10.1111/j.1471-4159.2009.06038.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wirenfeldt M, Babcock AA, Vinters HV. Microglia - insights into immune system structure, function, and reactivity in the central nervous system. Histol Histopathol. 2011;26:519–530. doi: 10.14670/HH-26.519. [DOI] [PubMed] [Google Scholar]
- 58.Woldbye DP, Angehagen M, Gotzsche CR, Elbrond-Bek H, Sorensen AT, Christiansen SH, Olesen MV, Nikitidou L, Hansen TV, Kanter-Schlifke I, Kokaia M. Adeno-associated viral vector-induced overexpression of neuropeptide Y Y2 receptors in the hippocampus suppresses seizures. Brain. 2010;133:2778–2788. doi: 10.1093/brain/awq219. [DOI] [PubMed] [Google Scholar]
- 59.Wu CY, Kaur C, Sivakumar V, Lu J, Ling EA. Kv1. 1 expression in microglia regulates production and release of proinflammatory cytokines, endothelins and nitric oxide. Neuroscience. 2009;158:1500–1508. doi: 10.1016/j.neuroscience.2008.11.043. [DOI] [PubMed] [Google Scholar]
- 60.Yamasaki R, Tanaka M, Fukunaga M, Tateishi T, Kikuchi H, Motomura K, Matsushita T, Ohyagi Y, Kira J. Restoration of microglial function by granulocyte-colony stimulating factor in ALS model mice. J Neuroimmunol. 2010;229:51–62. doi: 10.1016/j.jneuroim.2010.07.002. [DOI] [PubMed] [Google Scholar]
- 61.Yang S, Zhou W, Zhang Y, Yan C, Zhao Y. Effects of Liuwei Dihuang decoction on ion channels and synaptic transmission in cultured hippocampal neuron of rat. J Ethnopharmacol. 2006;106:166–172. doi: 10.1016/j.jep.2005.12.017. [DOI] [PubMed] [Google Scholar]
- 62.Yang S, Liu ZW, Wen L, Qiao HF, Zhou WX, Zhang YX. Interleukin-1beta enhances NMDA receptor-mediated current but inhibits excitatory synaptic transmission. Brain Res. 2005;1034:172–179. doi: 10.1016/j.brainres.2004.11.018. [DOI] [PubMed] [Google Scholar]
- 63.Yao ZG, Zhang L, Huang L, Zhu H, Liu Y, Ma CM, Sheng SL, Qin C. Regional and cell-type specific distribution of HDAC2 in the adult mouse brain. Brain Struct Funct. 2013;218:563–573. doi: 10.1007/s00429-012-0416-3. [DOI] [PubMed] [Google Scholar]
- 64.Zhang HJ, Sun RP, Lei GF, Yang L, Liu CX. Cyclooxygenase-2 inhibitor inhibits hippocampal synaptic reorganization in pilocarpine-induced status epilepticus rats. J Zhejiang Univ Sci B. 2008;9:903–915. doi: 10.1631/jzus.B0820018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhang XJ, Liu LL, Wu Y, Jiang SX, Zhong YM, Yang XL. receptor 1 is preferentially involved in modulation of N-methyl-D-aspartate receptor-mediated light-evoked excitatory postsynaptic currents in rat retinal ganglion cells. Neurosignals. 2011;19:110–116. doi: 10.1159/000326784. [DOI] [PubMed] [Google Scholar]
- 66.Zhao YN, Wang F, Fan YX, Ping GF, Yang JY, Wu CF. Activated microglia are implicated in cognitive deficits, neuronal death, and successful recovery following intermittent ethanol exposure. Behav Brain Res. 2013;236:270–282. doi: 10.1016/j.bbr.2012.08.052. [DOI] [PubMed] [Google Scholar]
- 67.Zheng H, Zhu W, Zhao H, Wang X, Wang W, Li Z. Kainic acid-activated microglia mediate increased excitability of rat hippocampal neurons in vitro and in vivo: crucial role of interleukin-1beta. Neuroimmunomodulation. 2010;17:31–38. doi: 10.1159/000243083. [DOI] [PubMed] [Google Scholar]
- 68.Zhu W, Zheng H, Shao X, Wang W, Yao Q, Li Z. Excitotoxicity of TNFalpha derived from KA activated microglia on hippocampal neurons in vitro and in vivo. J Neurochem. 2010;114:386–396. doi: 10.1111/j.1471-4159.2010.06763.x. [DOI] [PubMed] [Google Scholar]