

Abstract
Anthracimycin is a recently discovered novel marine-derived compound with activity against Bacillus anthracis. We tested anthracimycin against an expanded panel of Staphylococcus aureus strains in vitro and in vivo. All strains of S. aureus tested, including methicillin-sensitive (MSSA), methicillin-resistant (MRSA), and vancomycin-resistant strains of S. aureus were sensitive to anthracimycin at minimum inhibitory concentrations (MIC) of < 0.25 mg/L. Although its post-antibiotic effects were minimal, anthracimycin exhibited potent and rapid bactericidal activity, with a > 4-log kill of USA300 MRSA within 3 hours at 5 times its MIC. At concentrations significantly below the MIC, anthracimycin slowed MRSA growth and potentiated the bactericidal activity of the human cathelicidin, LL-37. The bactericidal activity of anthracimycin was somewhat mitigated in the presence of 20% human serum, and the compound was minimally toxic to human cells, with an IC50 = 70 mg/L against human carcinoma cells. At concentrations near the MIC anthracimycin inhibited S. aureus nucleic acid synthesis as determined by optimized macromolecular synthesis methodology, with inhibition of DNA and RNA synthesis occurring in the absence of DNA intercalation. Anthracimycin at a single dose of 1 or 10 mg/kg was able to protect mice from MRSA-induced mortality in a murine peritonitis model of infection. Anthracimycin provides an interesting new scaffold for future development of a novel MRSA antibiotic.
Keywords: Anthracimycin, Methicillin-resistant Staphylococcus aureus, novel antibiotic
INTRODUCTION
The continued prevalence of methicillin-resistant Staphylococcus aureus (MRSA) infections represents a major clinical challenge in the US.1,2 Complicating this fact, the rate of new antibiotic discovery over the last decade has not nearly kept pace with the rapid development of antibiotic resistance.3 Discovery of novel chemical entities with potent antibiotic activities is critical to continue the pipeline of therapeutic development. Marine-derived bacteria represent an important source of new chemical scaffolds with anti-MRSA activity.4-8 A novel tricyclic dione, termed anthracimycin, was recently purified from a previously uncharacterized marine-derived species of Streptomycetes and found to have potent activity against Bacillus anthracis and other Gram-positive bacteria.9 However, its activity against clinically-relevant drug-resistant pathogens including MRSA is unknown. Therefore, we undertook a more comprehensive analysis of anthracimycin activity in vitro and in vivo.
MATERIALS AND METHODS
Isolation of Anthracimycin
Anthracimycin was purified exactly as described from the marine-derived Streptomyces strain CNH365,9 and the structure of the purified compound was determined by combined spectroscopic methods and confirmed by a single crystal X-ray experiment (Figure 1).9
Figure 1. Chemical structure of anthracimycin.
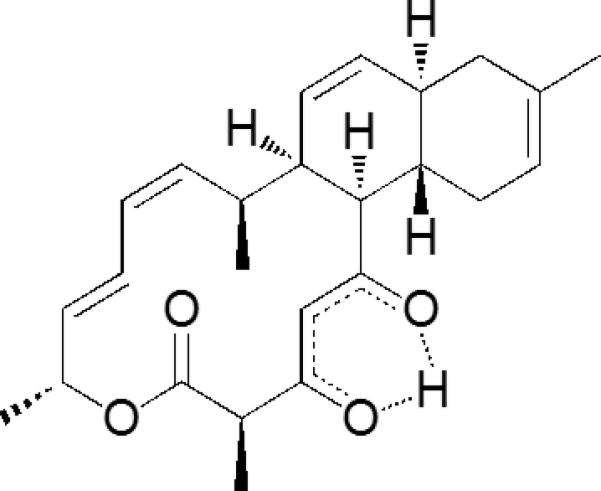
A single crystal X-ray experiment was used to confirm the structure of anthracimycin as determined by combined spectroscopic methods.
Minimum Inhibitory Concentration (MIC) Assays
Antimicrobial activity was evaluated by broth microdilution according to CLSI guidelines (Clinical and Laboratory Standards Institute. 2009. M100-S19 Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard, 9th ed. CLSI, Wayne, PA). MIC analysis in serum was done as previously described10 by addition of 20% pooled human serum (collected from > 5 donors according to an approved UCSD Institutional Review Board protocol). Bacterial viability at the end of the assay was determined by addition of resazurin to the wells. The MIC in serum was then determined to be the lowest compound concentration that did not produce a visible conversion of the resazurin (blue color) to resorufin (pink color).10
Time-Kill Kinetics
Anthracimycin time-kill kinetics and post-antibiotic effects were performed in duplicate by broth macrodilution. For the time-kill kinetics anthracimycin at 0x, 1x, 5x, 10x, or 20x the MIC (MIC = 0.125 mg/L for USA300 MRSA strain TCH1516) was added to CA-MHB in duplicate sterile polystyrene tubes (Falcon, Bedford MA). The media was then inoculated with ~ 5 x 105 colony-forming units (CFU)/mL in a final volume of 5 mL, and the tubes were incubated in a 37°C shaking incubator (New Brunswick). Viable bacteria over time were quantitated by removal of 25 μL aliquots for serial dilution in phosphate-buffered saline and plating on Todd-Hewitt agar (Hardy Diagnostics, Santa Maria, CA). Time-kill kinetic studies were performed in triplicate.
Post-Antibiotic Effect
MRSA strain TCH1516 was inoculated into 5 mL of CA-MHB containing anthracimycin (MIC = 0.125 mg/L) or vancomycin (MIC = 0.78 mg/L) at 1x or 10x MIC and incubated in a shaking incubator at 37°C. At one hour the bacteria were pelleted and then washed twice in 10 mL of phosphate-buffered saline and then resuspended in 5 mL CA-MHB. The tubes were placed back in the 37°C shaking incubator, and bacterial re-growth was measured at selected timepoints up to 24 hours by plating 25 μL samples of serial dilutions on Todd-Hewitt agar plates and counting colonies.
MRSA Growth in sub-MIC Anthracimycin
Growth curves at sub-MIC concentrations of anthracimycin, vancomycin, or vehicle control were performed by broth macrodilution format. Duplicate glass tubes containing CA-MHB (5 mL per tube) with either anthracimycin or vancomycin at predetermined concentrations were inoculated with ~ 5 x 105 CFU/mL MRSA strain TCH1516 and incubated in a 37°C shaking incubator. Growth was monitored at various timepoints up to 24 hours by measuring absorbance at 600 nm in a spectrophotometer (Spectronic 20D+, Thermo Scientific, Waltham, MA).
Growth studies at sub-MIC anthracimycin and LL-37 were done in broth microdilution format in 96 well plates using RPMI supplemented with 5% Luria broth (RPMI/LB). RPMI/LB (0.2 mL) containing ½x MIC of either: LL-37 (LL-37 MIC in RPMI/LB = 16 μM), anthracimycin, LL-37 in combination with anthracimycin, or vehicle was inoculated with ~ 5 – 10 × 105 CFU/mL MRSA strain TCH1516. The plates were incubated at 37°C, and bacterial survival over time was monitored by plating 25 μL serial dilutions on Todd-Hewitt agar for quantitative cultures.
Mammalian Cell Cytotoxicity
We assessed mammalian cytotoxicity using the human cervical carcinoma line HeLa (ATCC CCL-2). HeLa cells were plated at a density of 2 × 104 cells per well of sterile flat-bottom 96 well tissue-culture-treated plates (Corning), and anthracimycin was added at increasing concentrations. The plates were incubated in 5% CO2, 37°C, and cell viability (proliferation) was analyzed at 72 hours using the Promega Cell Proliferation kit (Promega, Madison, WI) according to manufacturer’s instructions, and IC50 was determined.
Mechanism of Action Studies
Anthracimycin mechanism of action studies were carried out using an optimized macromolecular synthesis assay11. Briefly, S. aureus strain ATCC 29213 was grown to mid-logarithmic phase in CA-MHB. Macromolecular synthesis in the presence of increasing doses of anthracimycin was monitored by measuring the incorporation of radiolabeled precursors of DNA, RNA, protein and cell wall synthesis ([3H]-Thymidine, [3H]-Uridine, [3H]-Leucine, and [3H]-N-Acetylglucosamine, respectively) in the presence of increasing anthracimycin. The extent by which synthesis of each pathway was inhibited was determined by calculating the difference in label incorporation relative to untreated controls. DNA intercalation was examined using a commercially available kit (Topogen) and observation of the products by agarose gel electrophoresis.
Murine Infection Studies
For the in vivo infection studies, eight week old female CD1 mice (Charles River Laboratories, Wilmington, MA) were inoculated intraperitoneally with ~ 1 × 109 CFU of MRSA strain Sanger 252 (a sequenced hospital-associated MRSA strain) and then treated one hour later with a single intraperitoneal dose of anthracimycin (1 or 10 mg/kg) or equivalent vehicle control (n = 10 mice per group). Survival was monitored over seven days, and mice that appeared moribund were humanely euthanized. This in vivo infection study was performed in duplicate, and data from one representative experiment are shown. All studies involving animals were reviewed and approved by the University of California San Diego Animal Care and Use Committee.
RESULTS
We found that anthracimycin was potent against all strains of S. aureus tested, including methicillin-susceptible and –resistant strains and vancomycin-resistant S. aureus, with MIC < 0.25 mg/L (Table 1). Anthracimycin was also active against clinical isolates of vancomycin-resistant Enterococcus faecalis. The potent Gram-positive activity was, however, mitigated in the presence of 20% human serum, which resulted in significant increases in the MIC against MRSA and methicillin-sensitive S. aureus (Table 1). Although this compound was active against the Gram-negative pathogen, Moraxella catarrhalis (MIC = 4 mg/L), other clinically-significant Gram-negatives including Klebsiella pneumoniae and Acinetobacter baumannii lacked anthracimycin susceptibility, with MIC >64 mg/L for these strains.
Table 1.
Minimum Inhibitory Concentration (MIC) Values
| Strain | MIC (mg/L) |
|---|---|
| MRSA | |
| Sanger 252 (USA200) | 0.063 |
| TCH1516 (USA300) | 0.125 (16 in 20% serum) |
| UAMS1182 (USA300) | 0.125 |
| ATCC33591 | 0.125 |
| NRS70 (ST5) | 0.08 |
| NRS100 (ST250) | 0.08 |
| NRS192 (ST1) | 0.16 |
| VRSA-PA | 0.125 |
| VRSA-MI | 0.25 |
| Other S. aureus | |
| UAMS1 | 0.125 (8 in 20% serum) |
| NRS77 (sequenced ST8) | 0.16 |
| NRS135 (ST8) | 0.16 |
| RN4220 | 0.125 (32 in 20% serum) |
| Newman | 0.16 |
| Non-S. aureus | |
| Enterococcus faecalis (ATCC 51299) | 0.25 |
| E. faecalis isolate 6981 | 0.125 |
| B. anthracis (Sterne) | 0.03 |
| Moraxella catarrhalis (ATCC 25238) | 4 |
| Pseudomonas aeruginosa (ATCC 27853) | > 64 |
| Klebsiella pneumoniae (ATCC 700603) | > 64 |
| Acinetobacter baumannii (ATCC 17978) | > 64 |
Anthracimycin exhibited rapid killing kinetics, with a > 4-log kill of USA300 MRSA within 3 hours at > 5x MIC (Figure 2a). Despite these rapid killing kinetics, anthracimycin showed minimal post-antibiotic effects against USA300 MRSA, with re-growth occurring rapidly after removal of the compound (Figure 2b). Mammalian cell cytotoxicity over 72 hours yielded an IC50 = 70 mg/L, which is several-fold higher than the serum MIC of 16 mg/L for USA300 MRSA. Given the rapid killing kinetics, we investigated the effects of sub-inhibitory concentrations of anthracimycin on USA300 MRSA. Even at 1/16x MIC (0.0078mg/L), anthracimycin substantially reduced the growth rate of MRSA during the first 8 hours of its logarithmic growth phase (Figure 3a). Comparatively, the cell wall antibiotic vancomycin had no effect on MRSA growth at 1/2x MIC (0.78 mg/L, Figure 3a). Although no interactions with other commonly prescribed MRSA antibiotics including vancomycin and daptomycin were detected in traditional checkerboard assays, we tested anthracimycin for interactions with a key constituent of the host immune defense, cathelicidin antimicrobial peptides. For these studies MRSA was co-incubated with sub-MIC concentrations of both anthracimycin and the human cathelicidin LL-37, and bacterial survival was monitored at selected time points by plating on agar. The addition of 1/4x MIC anthracimycin reduced viable MRSA counts in the presence of 1/2x MIC of LL-37 compared to either anthracimycin or LL-37 alone (Figure 3b). Additional studies indicated that this potentiation of activity was significantly mitigated in the presence of 20% human serum (data not shown).
Figure 2. Kinetics of anthracimycin activity against USA300 MRSA (strain TCH1516).
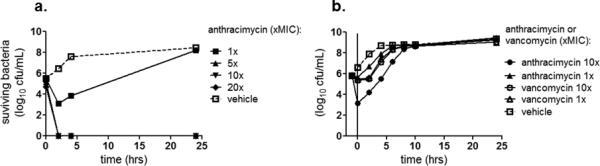
(a) Time-kill kinetics of anthracimycin at 1x, 5x, 10x, or 20x its MIC (MIC = 0.125 mg/L) during 24 hour incubation. (Note that the data points for the 5x, 10x, and 20x MIC anthracimycin are superimposed on the graph.) (b) Post-antibiotic effect of anthracimycin against USA300 MRSA (strain TCH1516) compared to vancomycin. The post-antibiotic effect was measured by incubating MRSA with anthracimycin or vancomycin for one hour (vertical line at 0 hr on the graph) and assessing 24 hour regrowth (viable bacteria) after washing the bacteria to remove the antibiotics. (Note that the data points for the 1x and 10x MIC vancomycin are nearly superimposed on the graph.)
Figure 3. Effects of sub-minimal inhibitory concentrations of anthracimycin on USA300 MRSA growth and antimicrobial peptide sensitivity.
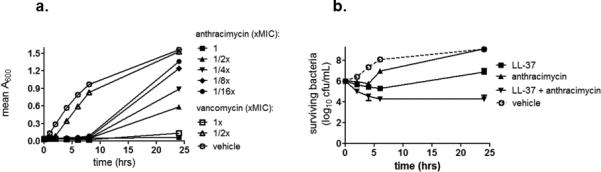
(a) Growth curves of MRSA (strain TCH1516) in the presence of vehicle or increasing sub-minimal inhibitory concentrations of anthracimycin (MIC = 0.125 mg/L) compared to 1/2x or 1x MIC vancomycin (vancomycin MIC = 0.78 mg/L). Turbidity as a measure of bacterial growth was monitored by optical density (A600) over time in duplicate tubes. (b) Killing of MRSA strain TCH1516 during incubation with: vehicle (MRSA alone), 1/2x MIC LL-37 alone, 1/4x anthracimycin alone, or 1/2x MIC LL-37 + 1/4x MIC anthracimycin. Surviving bacteria were enumerated by plating on agar, and the data represent mean +/- SD of duplicate samples.
Given the potent in vitro activity of anthracimycin, we investigated its mechanism of action in S. aureus using an optimized macromolecular synthesis assay to quantitate incorporation of radiolabeled precursors11. The primary effect on metabolic labeling was on [3H]-Thymidine and [3H]-Uridine incorporation, indicating the disruption of DNA and RNA synthesis, respectively (Figure 4). Disruption of these pathways occurs around the MIC concentration, suggesting that this effect is likely related to the antibacterial mechanism of action. Additional secondary effects on protein synthesis were also observed at much higher concentrations, at least 10-fold higher than the MIC. Primary disruption of DNA and RNA synthesis in metabolic labeling is often associated with DNA intercalators. However, when anthracimycin was examined for evidence of DNA intercalation over the same concentration range as used in metabolic labeling, no effect on DNA migration was observed relative to the untreated control (relaxed plasmid). Concentrations of 128 mg/L, 1000-fold higher than the MIC, show no evidence of DNA intercalation (Figure 4). These data suggest that the disruption of DNA and RNA synthesis by anthracimycin is not due to DNA intercalation.
Figure 4. Macromolecular synthesis in the presence of increasing anthracimycin.
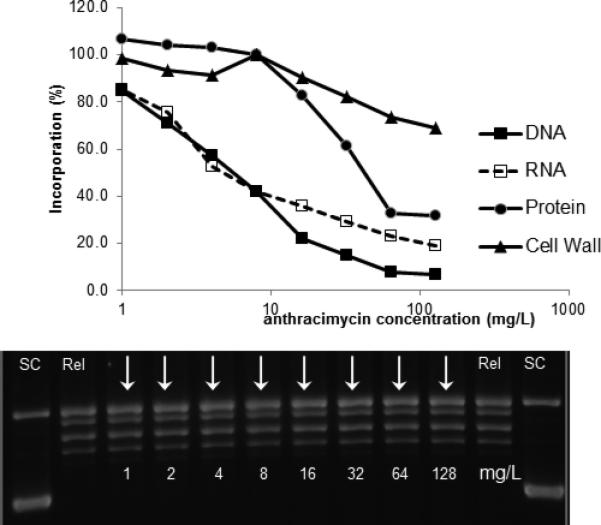
Incorporation of radiolabeled precursors of DNA, RNA, protein and cell wall synthesis ([3H]-Thymidine, [3H]-Uridine, [3H]-Leucine, and [3H]-N-Acetylglucosamine, respectively) was measured using S. aureus ATCC29213 (MSSA). DNA intercalation was examined using a commercially available kit. In the absence of DNA intercalation, relaxed plasmid migrates as a series of discrete topomers, with a characteristic ladder pattern. In contrast, intercalation results in a change in linking number, resulting in a change in the migration pattern. “SC” = supercoiled; “Rel” = relaxed.
To ascertain potential in vivo efficacy of anthracimycin, we tested its ability to protect mice from MRSA-induced mortality in a peritonitis model. Female CD1 mice were first infected intraperitoneally with ~109 CFU of MRSA strain Sanger 252, a route and inoculum producing rapid bacteremia and subsequent lethality. One hour after infection, the mice were divided into three groups (n = 10 per group) and received a single intraperitoneal injection of either anthracimycin (1 or 10 mg/kg) or equivalent vehicle control; anthracimycin at these doses appeared to be well-tolerated by the mice. Anthracimycin at either dose injected post-infection was sufficient to provide significant protection against mortality over seven days compared to infected mice treated with vehicle control alone (Figure 5). These results suggest that anthracimycin retained anti-MRSA activity and was also well-tolerated when introduced in vivo.
Figure 5. Anthracimycin protection of mice from MRSA mortality.
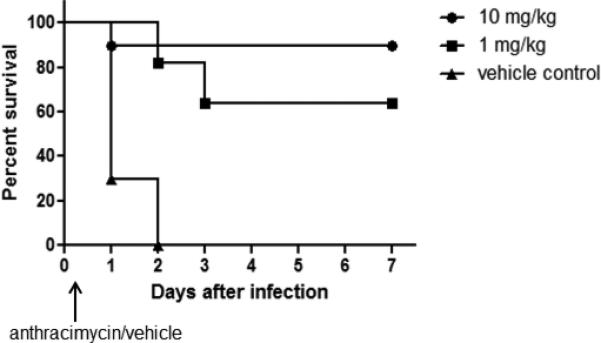
Female CD1 mice were infected intraperitoneally (~ 109 colony-forming units per mouse) with MRSA strain Sanger 252 and treated intraperitoneally one hour postinfection with either anthracimycin (1 or 10 mg/kg) or vehicle control (n = 10 mice per group). Survival was monitored over 7 days. These survival data are from one representative study that was repeated a total of two times.
DISCUSSION
The dearth of new chemical entities targeting problematic pathogens such as S. aureus has created a vacuum in the availability of novel antibiotics to treat these multidrug-resistant pathogens, and new antibiotics are urgently needed. The marine environment has been probed for decades as a source of novel scaffolds targeting cancer; however, similar studies targeting infectious diseases are much less common. The purification of anthracimycin from a marine-derived species of Streptomycetes and initial identification of its Gram-positive activity prompted us to explore further the potential of this scaffold as an anti-MRSA compound. Our results show that anthracimycin activity extends across 14 strains of S. aureus tested, including USA300 strains of MRSA, vancomycin-resistant S. aureus, and methicillin-sensitive S. aureus. No significant Gram-negative activity was observed. Human serum significantly mitigated but did not completely eliminate anthracimycin activity. Despite this serum-induced loss of activity, a single dose of anthracimycin was still protective in the murine peritonitis model even when the compound was administered post-infection. Given this initial demonstration of in vivo activity, investigation of additional dosing regimens for this compound and infection models (cutaneous, pulmonary, systemic) is now ongoing.
Anthracimycin demonstrated rapid killing kinetics against MRSA, although the post-antibiotic effect was quite negligible. It is possible that very little of the compound remains associated with the bacteria once it is removed from the media or that anthracimycin is highly susceptible to degradation, and future studies will address anthracimycin stability. Interestingly, our in vitro data suggest that anthracimycin was still able to exert effects on MRSA at concentrations below its MIC. We found a marked increase in lag phase when anthracimycin was present at 1/16x MIC; no similar effects were observed with the cell wall agent vancomycin up to 1/2x its MIC against MRSA. Notably, sub-MIC anthracimycin was able to potentiate the effects of the human cathelicidin LL-37 on MRSA growth. In these assays MRSA was able to grow during 24 hours in the presence of 1/2x MIC LL-37 or 1/4x MIC anthracimycin; however, no growth over the same time period was observed when the two were combined at those respective concentrations. It is possible that anthracimycin may act in synergy with LL-37 to hinder MRSA growth. This sensitization is not without precedent; for example, at well below its MIC, ampicillin has previously been shown to sensitize ampicillin-resistant Enterococcus to LL-37-mediated killing.12 Additional studies will continue to investigate anthracimycin interactions with the host immune system, including with other host-derived antimicrobial peptides.
In summary, our initial in vitro and in vivo studies suggest that the anthracimycin scaffold may serve as a viable lead for a medicinal chemistry effort to achieve a useful new MRSA therapeutic.
ACKNOWLEDGEMENTS
This work was supported by the National Institutes of Health (NIH) Training Program in Marine Biotechnology (T32 GM067550), a Ruth L Kirschstein National Research Service Award (NRSA) from National Institutes of Health grants (5 F31 GM090658-02), and by National Institutes of Health grant GM084350 to WF and VN. Part of this work was funded by the Transformational Medical Technologies program contract HDTRA1-10-C-004 from the Department of Defense Chemical and Biological Defense program through the Defense Threat Reduction Agency. The authors gratefully acknowledge the in vivo technical expertise of Joshua Olson.
REFERENCES
- 1.David MZ, Medvedev S, Hohmann SF, Ewigman B, Daum RS. Increasing burden of methicillin-resistant Staphylococcus aureus hospitalizations at US academic medical centers, 2003-2008. Infect. Control. Hosp. Epidemiol. 2012;33:782–789. doi: 10.1086/666640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Klein EY, Sun L, Smith DL, Laxminarayan R. The changing epidemiology of methicillin-resistant Staphylococcus aureus in the United States: A national observational study. Am. J. Epidemiol. 2013;177:666–674. doi: 10.1093/aje/kws273. [DOI] [PubMed] [Google Scholar]
- 3.Uhlemann AC, Otto M, Lowy FD, Deleo FR. Evolution of community- and healthcare-associated methicillin-resistant Staphylococcus aureus. Infect. Genet. Evol. 2013 doi: 10.1016/j.meegid.2013.04.030. doi:10.1016/j.meegid.2013.04.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Oh DC, Kauffman CA, Jensen PR, Fenical W. Induced production of emericellamides A and B from the marine-derived fungus Emericella sp. in competing co-culture. J. Nat. Prod. 2007;70:515–520. doi: 10.1021/np060381f. [DOI] [PubMed] [Google Scholar]
- 5.Hughes CC, Prieto-Davo A, Jensen PR, Fenical W. The marinopyrroles, antibiotics of an unprecedented structure class from a marine Streptomyces sp. Org. Lett. 2008;10:629–631. doi: 10.1021/ol702952n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Asolkar RN, Kirkland TN, Jensen PR, Fenical W. Arenimycin, an antibiotic effective against rifampin- and methicillin-resistant Staphylococcus aureus from the marine actinomycete Salinispora arenicola. J. Antibiot. 2010;63:37–39. doi: 10.1038/ja.2009.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sun P, et al. Fijimycins A-C, three antibacterial etamycin-class depsipeptides from a marine-derived Streptomyces sp. Bioorg. Med. Chem. 2011;19:6557–6562. doi: 10.1016/j.bmc.2011.06.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sakoulas G, et al. Novel bacterial metabolite merochlorin A demonstrates in vitro activity against multi-drug resistant methicillin-resistant Staphylococcus aureus. PloS One. 2012;7:e29439. doi: 10.1371/journal.pone.0029439. doi:10.1371/journal.pone.0029439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jang KH, et al. Anthracimycin, a Potent Anthrax Antibiotic from a Marine-Derived Actinomycete. Angew. Chem. Int. Ed. Engl. 2013;52(30):7822–7824. doi: 10.1002/anie.201302749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Haste NM, et al. Pharmacological properties of the marine natural product marinopyrrole A against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011;55:3305–3312. doi: 10.1128/AAC.01211-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cunningham ML, Kwan BP, Nelson KJ, Bensen DC, Shaw KJ. Distinguishing On-Target versus Off-Target Activity in Early Antibacterial Drug Discovery Using a Macromolecular Synthesis Assay. Journal Biomol. Screen. 2013;18(9):1018–1026. doi: 10.1177/1087057113487208. [DOI] [PubMed] [Google Scholar]
- 12.Sakoulas G, et al. Ampicillin enhances daptomycin- and cationic host defense peptide-mediated killing of ampicillin- and vancomycin-resistant Enterococcus faecium. Antimicrob. Agents Chemother. 2012;56:838–844. doi: 10.1128/AAC.05551-11. [DOI] [PMC free article] [PubMed] [Google Scholar]