

Abstract
In this short report, the genome-wide homologous recombination events were re-evaluated for classical swine fever virus (CSFV) strain AF407339. We challenged a previous study which suggested only one recombination event in AF407339 based on 25 CSFV genomes. Through our re-analysis on the 25 genomes in the previous study and the 41 genomes used in the present study, we argued that there should be possibly at least two clear recombination events happening in AF407339 through genome-wide scanning. The reasons for identifying only one recombination event in the previous study might be due to the limited number of available CSFV genome sequences at that time and the limited usage of detection methods. In contrast, as identified by most detection methods using all available CSFV genome sequences, two major recombination events were found at the starting and ending zones of the genome AF407339, respectively. The first one has two parents AF333000 (minor) and AY554397 (major) with beginning and ending breakpoints located at 19 and 607 nt of the genome respectively. The second one has two parents AF531433 (minor) and GQ902941 (major) with beginning and ending breakpoints at 8397 and 11,078 nt of the genome respectively. Phylogenetic incongruence analysis using neighbor-joining algorithm with 1000 bootstrapping replicates further supported the existence of these two recombination events. In addition, we also identified additional 18 recombination events on the available CSFV strains. Some of them may be trivial and can be ignored. In conclusion, CSFV might have relatively high frequency of homologous recombination events. Genome-wide scanning of identifying recombination events should utilize multiple detection methods so as to reduce the risk of misidentification.
Keywords: Sequence analysis, Homologous recombination, Virus evolution, PCR contamination
1. Introduction
Recombination, as the exchange of nucleotide segments among sequences, is believed to be ubiquitous in many viruses (Posada and Crandall, 2001; Worobey, 2000). However, in some viruses, the recombination events based on available genomic sequences were rarely detected, for example, influenza A, B and C viruses (Han and Worobey, 2011; He et al., 2009; Liu et al., 2010); Canine distemper virus (Han et al., 2008; McCarthy et al., 2007).
Classical swine fever virus (CSFV) is an ssRNA positive-strand virus that causes the contagious disease of swine. Gene evolution of the virus has been widely explored, from synonymous codon usage pattern (Cao et al., 2012; Tao et al., 2009) to selection pressure analysis (Perez et al., 2012; Shen et al., 2011; Tang et al., 2008). The putative homologous recombination of CSFV has been evaluated as well in an early paper (He et al., 2007), which only identified one recombination evidence. This seminal report would give us a first impression that the frequency of homologous recombination in CSFV might be low. However, given the increasing genome data of CSFV published in NCBI and the common recombination of many viruses (Posada and Crandall, 2001; Van der Walt et al., 2009; Worobey, 2000; Wu et al., 2009), we may expect to see some new recombination evidence happening on CSFV genomes when new sequences data are included. A comprehensive recombination analysis of CSFV has been performed in a previous study (He et al., 2007). The central objective of the present study is to re-evaluate and quantify the genome-wide homologous recombination evidence of CSFV. We would perform extensive recombination detection analyses using all available CSFV genome sequences in order to detect all possible recombination events and compare them to the previous study (He et al., 2007).
2. Materials and methods
2.1. Sequences
42 non-redundant complete genomes of CSFV were obtained from GenBank (http://www.ncbi.nlm.nih.gov/). The coding regions of the genomes were extracted and retained for subsequent analysis. The accession numbers for the genomes and other information were available in Appendix I. The sequences were aligned using MUSCLE software (Edgar, 2004) and available from the authors upon request.
Besides, since our study is to compare the results with the previous study (He et al., 2007), we also downloaded the 25 genomes listed in He et al. (2007)’s paper. Some genomes have been included in the above 42 genome dataset. To avoid confusions, we defined He et al. (2007)’s dataset as ‘He25.’ And, a full genome dataset with 54 sequences of CSFV, which is made by combining the genomes from the above 42 ones and ‘He25’ together after removing duplicated genomes, is defined as ‘Tot54’ dataset for comparative study.
2.2. Recombination analysis
The aligned sequences were then subjected to recombination analysis. The RDP software was used to detect homologous recombination events (Martin and Rybicki, 2010; Martin et al., 2010). The software contained a series of recombination detection algorithms, including GENECOV (Padidam et al., 1999), Bootscan/Rescan (Martin et al., 2005), Chimaera (Posada and Crandall, 2001), MaxChi (Maynard Smith, 1992), SiScan (Gibbs et al., 2000), 3Seq (Boni et al., 2007), and RDP (Martin and Rybicki, 2010). All these methods are utilized and compared so as to obtain consensus results. A putative recombination event was retained to subsequent analysis only when it was consistently identified by at least three of the above-mentioned seven algorithms (Liu et al., 2010). Here we defined the minor parent as the one contributing the smaller fraction of the recombinant, while the major parent as the one contributing the larger fraction of the recombinant (Martin et al., 2010).
To avoid false-positive results, phylogenetic analysis of recombination was performed (Boni et al., 2008; Liu et al., 2010). For each putative recombinant, the entire dataset alignment was divided at the breakpoint positions. If two recombination breakpoints were found in a single sequence, the sequence region between the breakpoints was denoted the “minor” region, generated by the minor parent, and the remainder is the “major” region, generated by the major parent. Neighbor-joining phylogenetic trees were constructed to show topological shifts of specific sequences. Phylogenetic incongruence is reflected by a putative recombinant whose distance in the phylogeny is clearly close to one parent and far from another one for each sequence segment. Also, the recombination signal “weak” if one of the components received only low bootstrap support (Boni et al., 2008; Liu et al., 2010).
3. Results and discussion
3.1. Recombination events in CSFV using ‘He25’ dataset
When redoing the 25 genomes used in He et al. (2007), six unique recombination events were supported by at least 4 out of 7 detection methods and we did find that AF407339 was a recombinant which had two most significant recombination events (the other 4 ones are listed in Table 1). The first one has corresponding parents Z46258 (minor) and AY072924 (major), which is supported by all the 7 recombination methods with starting and ending breakpoints at 8421 and 12,260 nt of the genome (Table 2). The second one has corresponding parents AF333000 (minor) and AY554397 (major) with starting and ending breakpoints at 19 and 639 nt of the genome respectively. This event was only supported by 4 methods (Table 1). Also, it was found that the location of breakpoints for the event which took AF333000 as one parent was different between our re-analysis and He et al. (2007)’s report.
Table 1.
Recombinant events for CSFV genomes found in He25 dataset.
| Recombinant | Major | Minor | Starting | Ending | Supporting methods |
|---|---|---|---|---|---|
| D49532 | AF326963 | X87939 | 1386 | 2375 | RDP, GENECONV, Bootscan, SiScan, 3Seq |
| D49532 | AF326963 | U90951 | 2754 | 3571 | RDP, GENECONV, Bootscan, SiScan |
| DQ127910 | Unknown (U90951) | U45478 | 9787 | 10,739 | RDP, GENECONV, Bootscan, SiScan, 3Seq |
| AF333000 | AY072924 | Z46258 | 88 | 12,260 | All methods |
| AF407339 | AY554397 | AF333000 | 19 | 639 | RDP, GENECONV, Bootscan, SiScan |
| AF407339 | AY072924 | Z46258 | 8421 | 12,260 | All methods |
Table 2.
Recombinant events for CSFV genomes found in Tot54 combined dataset.
| Recombinant | Major | Minor | Starting | Ending | Supporting methods |
|---|---|---|---|---|---|
| JQ268754 | FJ529205 | GU592790 | 10,678 | 11,514 | RDP, GENECONV, Bootscan, MaxChi, Chimaera, SiScan |
| AF407339 | AY554397 | AF333000 | 19 | 607 | RDP, GENECONV, Bootscan, SiScan |
| AF407339 | GQ122383 | AF333000 | Undetermined (11,079) | 12,268 | RDP, GENECONV, Bootscan, Chimaera, SiScan, 3Seq |
| AF407339 | GQ902941 | AF531433 | 8397 | 11,078 | All methods |
| AF407339 | HQ380231 | Unknown (DQ127910) | Undetermined (8397) | 8633 | RDP, GENECONV, MaxChi, SiScan, 3Seq |
| DQ127910 | GQ902941 | AF531433 | 57 | 12,280 | All methods |
| EU497410 | GQ902941 | AF531433 | 57 | 12,281 | All methods |
| AY775178 | GQ902941 | AF531433 | 57 | 12,280 | All methods |
| D49533 | GQ902941 | AF531433 | 172 | 12,281 | All methods |
| HM237795 | GQ902941 | AF531433 | 172 | 12,281 | All methods |
| AF326963 | GQ902941 | AF531433 | 172 | 12,284 | All methods |
| D49532 | AF326963 | U90951 | 2754 | 3473 | GENECONV, Bootscan, SiScan, 3Seq |
| D49532 | HM237795 | U90951 | 1111 | 2397 | RDP, GENECONV, Bootscan, SiScan, 3Seq |
| D49532 | GQ902941 | AF531433 | 172 | 12,281 | All methods |
| X87939 | GQ902941 | AF531433 | 172 | 12,281 | All methods |
| U90951 | GQ902941 | AF531433 | 172 | 12,281 | All methods |
| EU789580 | HM237795 | U90951 | 12,244 | 2369 | RDP, GENECONV, Bootscan, SiScan, 3Seq |
| EU789580 | GQ902941 | AF531433 | 172 | 12,281 | All methods |
| EU915211 | HM237795 | U90951 | 163 | 2369 | RDP, GENECONV, Bootscan, SiScan, 3Seq |
| EU915211 | GQ902941 | AF531433 | 172 | 12,281 | All methods |
3.2. Recombination events in CSFV using ‘Tot54’ dataset
When analyzing all the available CSFV genomes (Table 2), it was found that 20 recombination events were identified with the support from at least four out of seven detection methods. As such, it was concluded that the recombination frequency in CSFV genomes is quite high.
Four recombination events have been identified on the recombinant AF407339 (Table 2). The first one had two parents AF333000 (minor) and AY554397 (major) with beginning and ending breakpoints located at 19 and 607 nt of the genome. The second one had two parents AF531433 (minor) and GQ902941 (major) with beginning and ending breakpoints at 8397 and 11,078 nt of the genome respectively. The phylogenetic analyses confirmed the existence of these two recombination events with strong bootstrapping support (Figs. 1 and 2). Finally, the last two ones had very short recombination segments and one/or both of the parents are unknown. Thus, the last two ones were ignored for subsequent discussion.
Figure 1.
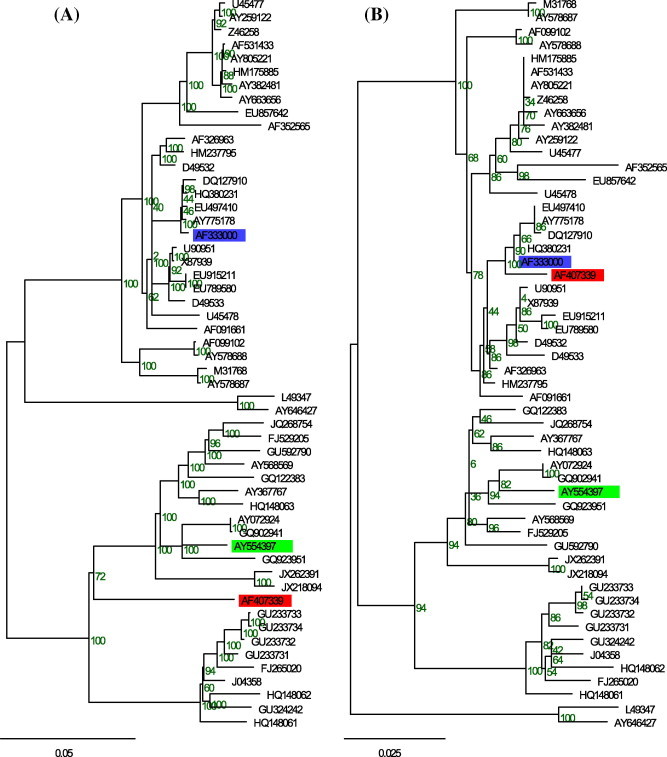
Neighbor-joining trees for the CSFV genomes, showing the evidence for the first major recombination event on the recombinant AF407339. The tree on subplot A was inferred from the major region, while the one on the subplot B was inferred from the minor region. All branch lengths are drawn to a scale of nucleotide substitutions per site. Numbers on the nodes indicate the bootstrapping support scores. The sequences marked with red, blue and green colors indicate the recombinant, the minor parent and major parent, respectively.
Figure 2.
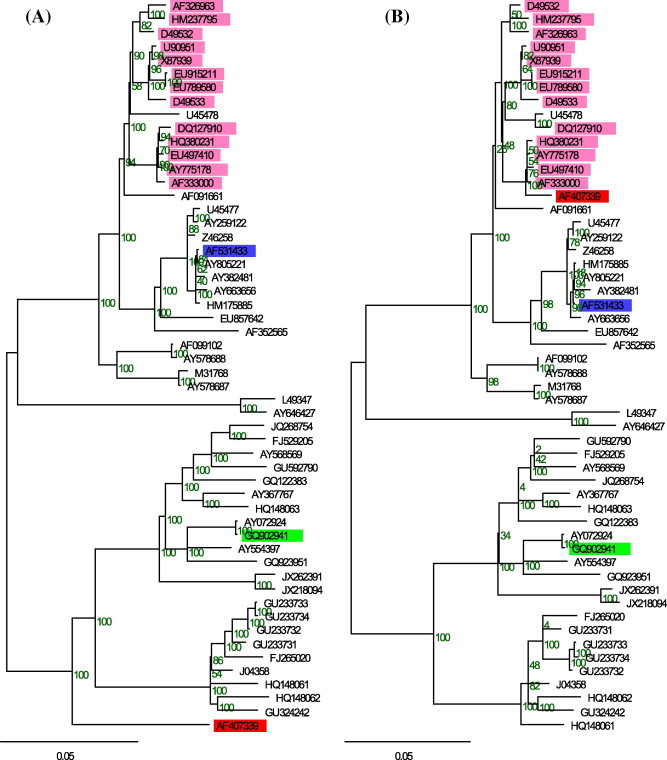
Neighbor-joining trees for the CSFV genomes, showing the evidence for the second major recombination event on the recombinant AF407339. The tree on subplot A was inferred from the major region, while the one on the subplot B was inferred from the minor region. All branch lengths are drawn to a scale of nucleotide substitutions per site. Numbers on the nodes indicate the bootstrapping support scores. The sequences marked with red, blue and green colors indicate the recombinant, the minor parent and major parent respectively. Sequences marked in pink indicate the ones with partial evidence of the same recombination event.
3.3. A systematic comparison and analysis of the recombination events on AF407339
We explored the three recombination-related sequences AF333000, AY367767 and AF407339 identified by He et al. (2007) by checking possible signals of recombination in RDP program. However, no recombination signals were found for these three sequences. However, when we added another three sequences AY554397, AF531433 and GQ902941 into the analysis, the two recombination events discovered above by using ‘He25’ and ‘Tot54’ datasets were clearly present. As such, it is concluded that the recombination event identified by He et al. (2007) might not be robust enough.
We argued that the limitation of He et al. (2007)’s work may be due to the limited sampling of the genome sequences at the year 2007 and the limited usage of recombination methods (He et al., 2007). At that time, other CSFV sequences have become available online, but not been considered in the analyses in the previous study. These sequences included AF531433 (available since the year of 2002), AY663656 (available since the year of 2004), and AY805221 (available since the year of 2004). In particular, the exclusion of the significantly putative recombination parent AF531433 in the analysis can largely limit the identification sensitivity of recombination events in CSFV genomes. As seen in the present results (Table 2), AF531433 was found to be one of the parents for many recombinants. Moreover, in their study, only one method was utilized, SimPlot (Lole et al., 1999), therefore largely reducing the reliability and accuracy of the identification of recombination events. SimPlot only employed one method Bootscan for detecting recombination events. In contrast, in our study, we utilized all 7 statistical methods available in RDP program, plus the phylogenetic analysis to identify the recombination parents. As such, we find no support of AF333000 as the minor parent of the recombination event.
At last, although the current recombination detection methods yielded high significance levels for the recombination events for CSFV, we have to acknowledge that they have the risk of false-positive detection of recombination (Leal et al., 2012). Further lab experiments might be required to verify these recombination events proposed in our study.
As the implications, the present study suggested that multiple statistical methods should be utilized so as to accurately identify the recombination events (Attoui et al., 2007; Shin et al., 2013). Finally, as seen, CSFV possesses relatively high frequency of homologous recombination events.
Footnotes
Peer review under responsibility of King Saud University.
References
- Attoui H., Sailleau C., Jaafar F., Belhouchet M., Biagini P., Cantaloube J., Micco P., Mertens P., Zientara S. Complete nucleotide sequence of Middelburg virus, isolated from the spleen of a horse with severe clinical disease in Zimbabwe. J. Gen. Virol. 2007;88:3078–3088. doi: 10.1099/vir.0.83076-0. [DOI] [PubMed] [Google Scholar]
- Boni M., Posada D., Feldman M. An exact nonparametric method for inferring mosaic structure in sequence triplets. Genetics. 2007;176:1035–1047. doi: 10.1534/genetics.106.068874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boni M., Zhou Y., Taubenberger J., Holmes E. Homologous recombination is very rare or absent in human influenza A virus. J. Virol. 2008;82:4807–4811. doi: 10.1128/JVI.02683-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao H., Zhang H., Cui Y. Synonymous codon usage bias of E2 genes of classical swine fever virus. Israel J. Vet. Med. 2012;67:253–258. [Google Scholar]
- Edgar R. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–1797. doi: 10.1093/nar/gkh340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibbs M., Armstrong J., Gibbs A. Sister-Scanning: a Monte Carlo procedure for assessing signals in recombinant sequences. Bioinformatics. 2000;16:573–582. doi: 10.1093/bioinformatics/16.7.573. [DOI] [PubMed] [Google Scholar]
- Han G., Liu X., Li S. Cross-species recombination in the haemagglutinin gene of canine distemper virus. Virus Res. 2008;136:198–201. doi: 10.1016/j.virusres.2008.04.022. [DOI] [PubMed] [Google Scholar]
- Han G., Worobey M. Homologous recombination in negative sense RNA viruses. Viruses. 2011;3:1358–1373. doi: 10.3390/v3081358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He C., Ding N., Chen J., Li Y. Evidence of natural recombination in classical swine fever virus. Virus Res. 2007;126:179–185. doi: 10.1016/j.virusres.2007.02.019. [DOI] [PubMed] [Google Scholar]
- He C., Xie Z., Han G., Dong J., Wang D., Liu J., Ma L., Tang X., Liu X., Pang Y., Li G. Homologous recombination as an evolutionary force in the avian influenza A virus. Mol. Biol. Evol. 2009;26:177–187. doi: 10.1093/molbev/msn238. [DOI] [PubMed] [Google Scholar]
- Leal E., Villanova F., Lin W., Hu F., Liu Q., Liu Y., Cui S. Interclade recombination in porcine parvovirus strains. J. Gen. Virol. 2012;93:2692–2704. doi: 10.1099/vir.0.045765-0. [DOI] [PubMed] [Google Scholar]
- Liu X., Wu C., Chen A. Codon usage bias and recombination events for neuraminidase and hemagglutinin genes in Chinese isolates of influenza A virus subtype H9N2. Arch. Virol. 2010;155:685–693. doi: 10.1007/s00705-010-0631-2. [DOI] [PubMed] [Google Scholar]
- Lole K., Bollinger R., Paranjape R., Gadkari D., Kulkarni S., Novak N., Ingersoll R., Sheppard H., Ray S. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999;73:152–160. doi: 10.1128/jvi.73.1.152-160.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin D., Lemey P., Lott M., Moulton V., Posada D., Lefeuvre P. RDP3: a flexible and fast computer program for analyzing recombination. Bioinformatics. 2010;26:2462–2463. doi: 10.1093/bioinformatics/btq467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin D., Posada D., Crandall K., Williamson C. A modified bootscan algorithm for automated identification of recombinant sequences and recombination breakpoints. AIDS Res. Hum. Retroviruses. 2005;21:98–102. doi: 10.1089/aid.2005.21.98. [DOI] [PubMed] [Google Scholar]
- Martin D., Rybicki E. RDP: detection of recombination amongst aligned sequences. Bioinformatics. 2010;26:2462–2463. doi: 10.1093/bioinformatics/16.6.562. [DOI] [PubMed] [Google Scholar]
- Maynard Smith J. Analyzing the mosaic structure of genes. J. Mol. Evol. 1992;34:126–129. doi: 10.1007/BF00182389. [DOI] [PubMed] [Google Scholar]
- McCarthy A., Shaw M., Goodman S. Pathogen evolution and disease emergence in carnivores. Proc. R. Soc. B Biol. Sci. 2007;274:3165–3174. doi: 10.1098/rspb.2007.0884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Padidam M., Sawyer S., Fauquet C. Possible emergence of new geminiviruses by frequent recombination. Virology. 1999;265:218–225. doi: 10.1006/viro.1999.0056. [DOI] [PubMed] [Google Scholar]
- Perez L., Arce H., Perera C., Rosell R., Frias M., Percedo M., Tarradas J., Doinguez P., Nuenez J., Ganges L. Positive selection pressure on the B/C domains of the E2-gene of classical swine fever virus in endemic areas under C-strain vaccination. Infect. Genet. Evol. 2012;12:1405–1412. doi: 10.1016/j.meegid.2012.04.030. [DOI] [PubMed] [Google Scholar]
- Posada D., Crandall K. Evaluation of methods for detecting recombination from DNA sequences: computer simulations. Proc. Natl. Acad. Sci. 2001;98:13757–13762. doi: 10.1073/pnas.241370698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen H., Pei J., Bai J., Zhao M., Ju C., Yi L., Kang Y. Genetic diversity and positive selection analysis of classical swine fever virus isolates in south China. Virus Genes. 2011;43:234–242. doi: 10.1007/s11262-011-0625-5. [DOI] [PubMed] [Google Scholar]
- Shin D., Richards S., Alto B., Bettinardi D., Smartt C. Genome sequence analysis of Dengue virus 1 isolated in Key West, Florida. PLoS One. 2013;8:e74582. doi: 10.1371/journal.pone.0074582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang F., Pan Z., Zhang C. The selection pressure analysis of classical swine fever virus envelope protein genes Erns and E2. Virus Res. 2008;131:132–135. doi: 10.1016/j.virusres.2007.08.015. [DOI] [PubMed] [Google Scholar]
- Tao P., Dai L., Luo M., Tang F., Tien P., Pan Z. Analysis of synonymous codon usage in classical swine fever virus. Virus Genes. 2009;38:104–112. doi: 10.1007/s11262-008-0296-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Walt E., Rybicki E., Varsani A., Polston J., Billharz R., Donaldson L., Monjane A., Martin D. Rapid host adaptation by extensive recombination. J. Gen. Virol. 2009;90:734–746. doi: 10.1099/vir.0.007724-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worobey M. Extensive homologous recombination among widely divergent TT viruses. J. Virol. 2000;74:7666–7670. doi: 10.1128/jvi.74.16.7666-7670.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J., Yu T., Bao Q., Zhao F. Evidence of extensive homologous recombination in the core genome of Rickettsia. Comp. Funct. Genomics. 2009;2009 doi: 10.1155/2009/510270. [DOI] [PMC free article] [PubMed] [Google Scholar]