

Abstract
Cyclic stretch is known to alter a number of cellular and subcellular processes, including those involved in nonviral gene delivery. We have previously shown that moderate equibiaxial cyclic stretch (10% change in basement membrane area, 0.5 Hz, 50% duty cycle) of human pulmonary A549 cells enhances gene transfer and expression of reporter plasmid DNA in vitro, and that this phenomena may be due to alterations in cytoplasmic trafficking [Taylor et al, 2003, Mol Therapy 7:542–549]. Although the path by which plasmid DNA travels through the cytoplasm toward the nucleus is not well understood, the cytoskeleton and the constituents of the cytoplasm are known to significantly hinder macromolecular diffusion. Using biochemical techniques and immunofluorescence microscopy, we show that both the microfilament and microtubule networks are significantly reorganized by equibiaxial cyclic stretch. Prevention of this reorganization through the use of cytoskeletal stabilizing compounds mitigates the stretch-induced increase in gene expression, however depolymerization in the absence of stretch is not sufficient to increase gene expression. These results suggest that cytoskeletal reorganization plays an important role in stretch-induced gene transfer and expression.
Keywords: electroporation, microtubule, microfilament, trafficking, transfection, actin, plasmid
INTRODUCTION
Although the cell biology of viral and nonviral gene transfer has been studied over the past several years, all studies have focused on the mechanisms employed in static cultures in the absence of any external stresses applied to the cells. This is very different from the in vivo setting, a very dynamic environment where cells experience a number of external stresses. These stresses include, but are not limited to shear, stretch and compression.1 Consequently, if we are to optimize gene delivery in vivo, we must understand what effects such stresses have on the gene transfer process.
It has been well documented that a variety of cells behave very differently under stress than under static conditions. Signaling cascades and transcription factors are upregulated and downregulated, cytoskeletal elements are reoriented, reactive oxygen species are produced, and, under extreme stresses, cells enter apoptosis.2 A whole field of mechanotransduction has emerged, and models such as tensegrity have been developed in an effort to describe how cells behave under various forms of stress, and how these stresses are translated into biochemical cascades and other cellular responses.3–5 This is particularly true in the lung, a dynamic organ that undergoes continuously modulating exogenous forces due to cyclical respiratory patterns. In the injured lung, these forces can be even more prevalent, due to mechanical ventilation or decreased airway space resulting from edema or compromised lung tissue.6 Understanding how these forces affect gene transfer is critical to developing rational delivery approaches.
Using plasmid DNA, our lab has previously shown that biaxial cyclic stretch using physiologic deformations (10% change in mean membrane surface area)7 causes an increase in reporter gene expression following gene transfer.8 When the barriers that plasmid DNA must cross in order to successfully transfect a cell are examined, it becomes apparent that these same barriers are altered by exogenous mechanical forces; namely alterations in the cytoskeleton and extracellular matrix,9–11 activation of cell signaling pathways12 and changes in transcription factor activation.13,14 We concluded that due to the time course of cyclic stretch necessary to induce increased expression, as well as the ability for this enhancement to occur in cells transfected by either electroporation or liposomal transfection, the mechanisms responsible for the stretch-enhanced expression were at the level of either nuclear entry or cytoplasmic trafficking. In the present study, we have closely examined how cyclic stretch affects the cytoskeleton of our in vitro model. By using biochemical and fluorescence microscopy techniques to examine the effects of cyclic stretch on cytoskeletal organization, we looked at the response of the microtubule and microfilament networks to cyclic stretch, and how their reorganization affected gene transfer. Our results suggest that cytoskeletal reorganization of both the microtubule and microfilament network due to biaxial cyclic stretch is a necessary component in stretch-enhanced gene transfer and expression.
RESULTS
Cyclic Stretch Causes Significant Reorganization and Depolymerization of the Microtubule Network
Previous work in the response of microtubule networks to exogenous stress has looked at the effects of either static stretch or static compression on the microtubules.10,15 We found that when microtubules undergo a dynamic deformation in the form of biaxial cyclic stretch, significant remodeling of the microtubule network occurs. Analysis via Western Blot indicated that tubulin in the TX-100 insoluble fraction (i.e. polymerized tubulin) decreased significantly over time with stretch (Figure 1A and 1B), indicating a depolymerization of the microtubule network. Stretching for as little as 30 minutes caused nearly a 50% reduction in the amount of polymerized tubulin while by 24 hours, the amount of microtubules was only 10–20% of unstretched control cells. This result was confirmed by immunofluorescence microscopy (Figure 1C), where after 24 hours of equibiaxial cyclic stretch, fewer microtubules were seen (right panel) compared to unstretched control (left panel). These fibers also appeared to be slightly thickened following stretch. Although the stretched cells were more rounded than their unstretched counterparts, we found that stretch out to 24 hours did not cause significant cell death as determined by the ability to further passage these cells upon cessation of cyclic stretch, nor did it cause detachment from the fibronectin-coated silastic membrane (not shown).
Figure 1. Microtubule reorganization due to equibiaxial cyclic stretch.
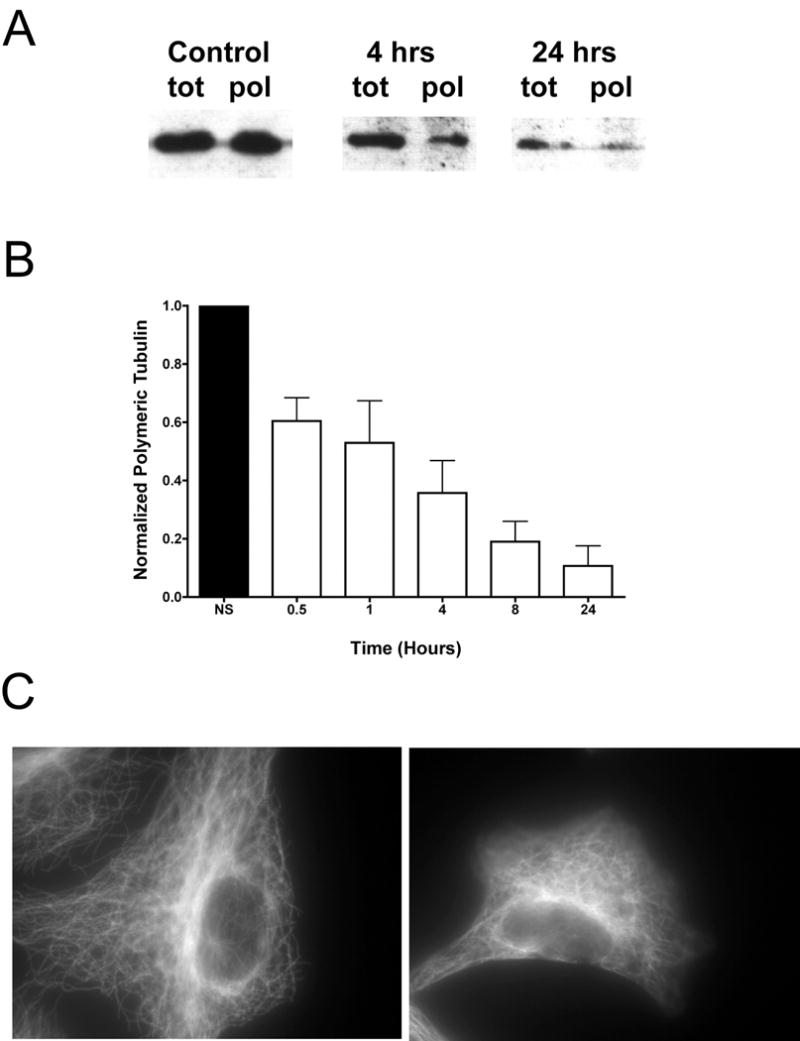
Cells were grown on silastic membranes and stretched at 80–90% confluency for up to 24 hours at 0.5 Hz, with a 10% change in basement membrane area. Cells were removed from the stretching environment at t = 0.5, 1, 4, 8 or 24 hours and allowed to grow under static conditions out to 24 hours, at which time the protein containing either total tubulin or polymerized tubulin was extracted. A) A typical western blot with cells stretched for 0, 4 and 24 hours showing both polymeric and total tubulin. B) Densitometry measurements for polymeric tubulin relative to unstretched controls were averaged from at least 6 samples in 3 separate experiments (± st. dev.). Cells were also immunostained for tubulin (C) after stretch for 24 hours (right panel) and compared to unstretched controls (left panel).
Cyclic Stretch Reorganizes, But Does Not Depolymerize the Actin Network
We next examined the effects of cyclic stretch on the organization of the actin cytoskeleton. The NP-40 soluble and insoluble fractions of the microfilament network, representing G- and F-actin, respectively, were not significantly altered by the addition of cyclic stretch (Figure 2A and 2B). On average, we never saw more than a 10% change in the filamentous actin pool when cells were stretched out to 24 hours. Harvesting these cells immediately after stretching seemed to demonstrate the greatest change in filamentous actin, but again these changes were typically in the 7–10% range. Surprisingly, when the stretched cells were viewed by immunofluorescence microscopy (Figure 2C), significant reorganization of the actin cytoskeleton was observed (right panel) when compared to unstretched control (left panel). The once cell-spanning fibers were reduced to much shorter actin fragments following stretch. Additionally, there was an accumulation of actin at the periphery of the cell that was not seen in unstretched cells.
Figure 2. Microfilament reorganization due to equibiaxial cyclic stretch.
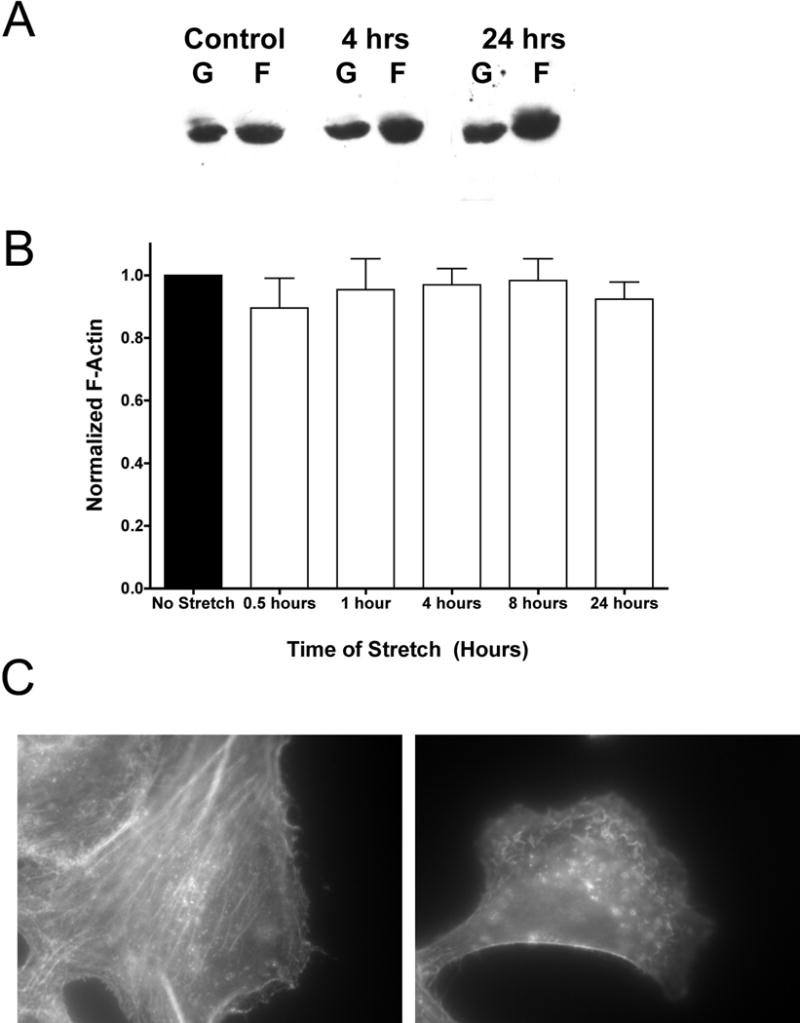
Cells were grown for up to 24 hours and stretched as in Figure 1. Cells were removed from the stretching environment at t = 0.5, 1, 4, 8 or 24 hours and allowed to grow under static conditions out to 24 hours, at which time the protein containing either G-actin or F-actin was extracted. A) A typical western blot with cells stretched for 0, 4 and 24 hours showing both G- and F-actin. B) Densitometry measurements for F-actin relative to unstretched controls were averaged from at least 8 samples in 4 separate experiments (± st. dev.). Cells were also immunostained for actin (C) using Alexa 546 conjugated phalloidin after 24 hours of stretch (right panel) and compared to unstretched controls (left panel).
Thirty Minutes of Cyclic Stretch Causes Depolymerization of the Microtubule Network
While we do see significant reorganization of the microtubule network after 24 hours of cyclic stretch, we have previously shown that as little as 30 minutes of equibiaxial cyclic stretch following electroporation is sufficient to increase gene expression in A549 cells.8 As shown in Figure 3A, as little as 30 minutes of cyclic stretch was also sufficient to decrease the amount of polymeric tubulin as determined by Western Blot, but had little to no effect on microfilaments (Figure 3B). The depolymerization of the microtubules following 30 minutes of stretch caused a 50% reduction in microtubules that did not recover when the cells were then grown under static conditions for up to 24 hours. This suggested that the stretch-induced changes in the microtubule network due to 30 minutes of cyclic stretch were retained by the cells throughout the course of our experiments. When stretched for longer periods of time, the amount of monomeric tubulin increased out to 24 hours, where greater than 80% of the tubulin was found to be in the monomeric form as assessed by western blot (Figure 1A). This result again indicates that cyclic stretch can cause significant reorganization of the cytoskeletal network, and it is possible that this reorganization may aid in the cytoplasmic trafficking of plasmid DNA.
Figure 3. Thirty minutes of equibiaxial cyclic stretch is sufficient for cytoskeletal reorganization.
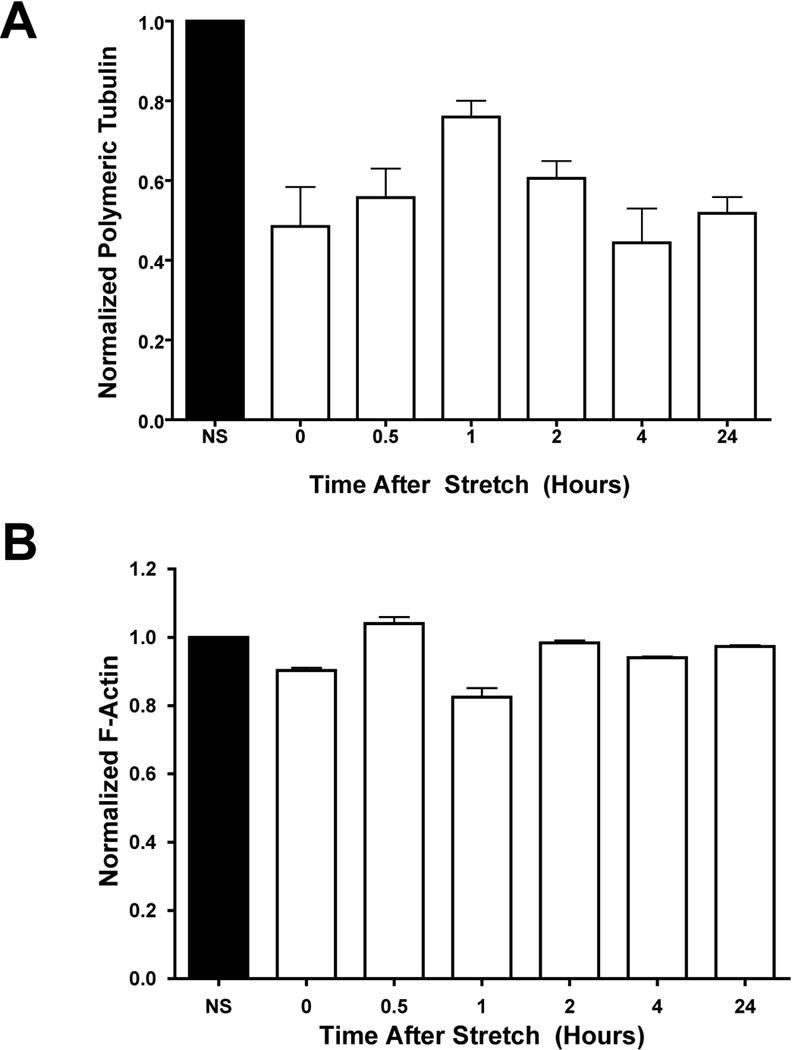
Densitometry measurements for polymeric tubulin (A) or F-actin (B) relative to unstretched control after 30 minutes of stretch followed by static growth for up to 24 hours. Cells were grown on silastic membranes and stretched for 30 minutes at 0.5 Hz, with a 10% change in basement membrane area and a duty cycle of 50% upon reaching 80–90% confluency. After stretching, cells were harvested at t = 0, 0.5, 1, 2, 4 or 24 hours later. Mean densitometry measurements (± st. dev.) were from at least 8 samples in 4 independent experiments.
Reorganization of the Cytoskeleton is Necessary, but not Sufficient, for Stretch Induced Increases in Gene Expression
To further assess the role of the cytoskeletal networks in stretch-enhanced gene transfer, we probed the networks using a variety of cytoskeleton stabilizing and destabilizing drugs. In this set of experiments, drugs affecting either the microfilament network (the microfilament stabilizing drug jasplakinolide or destabilizing drug latrunculin B) or the microtubule network (the microtubule stabilizing drug taxol or destabilizing drug nocodazole) were added to the cell cultures immediately following electroporation of a luciferase expressing reporter plasmid. Because electroporation does not utilize the endocytic pathway to deliver DNA into the cytoplasm, drugs that alter the cytoskeleton can be used to study intracellular movement of the DNA following electroporation independent of their confounding effects on endocytosis. The cultures were then subjected to stretch or non-stretch conditions similar to previous experiments. When reorganization of either the microtubule or microfilament network was prevented through the addition of cytoskeletal stabilizing drugs, the stretch-induced increase in gene expression was mitigated (Figure 4). Furthermore, while depolymerization of either the microfilament or microtubule networks by cytoskeletal destabilizing drugs did not prevent the stretch-induced transfection enhancement, it also was not sufficient to significantly increase gene expression in the static cells (Figure 4). This indicates that stretch-induced reorganization of either the microtubule or actin cytoskeleton alone is not sufficient to account for the stretch-enhanced transfection efficiency. At the very least, it would seem that reorganization of both the microfilament and the microtubule networks is required to drive enhanced gene expression.
Figure 4. Stretch-enhanced gene transfer and expression is mitigated by cytoskeletal stabilization.
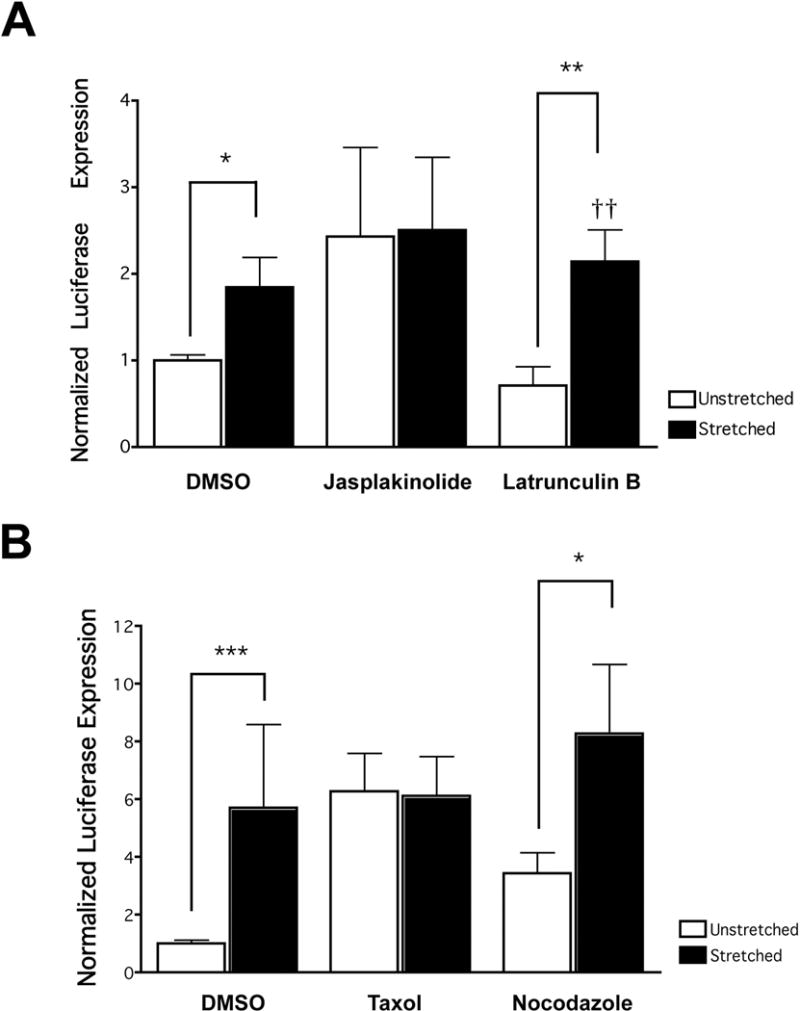
Cells were electroporated with pCMV-Lux-DTS and media containing serum and either one of the actin drugs jasplakinolide or latrunculin B (A) or one of the microtubule drugs taxol or nocodazole (B) were administered to the cells. The cells were then stretched (0.5 Hz, with a 10% change in basement membrane area and a duty cycle of 50%) or grown statically for 24 hours. After 24 hours, luciferase expression was determined (n = 8–12 wells of cells from at least 3 separate experiments for each data set). * p < 0.05, ** p < 0.01, *** p < 0.001 between stretched and unstretched pairs. †† p < 0.01 when compared with unstretched vehicle only (DMSO).
Cyclic Stretch Causes Increased Gene Expression without Increased Proliferation or Decreased Survival
As previously published, our results showed that there was increased expression of the luciferase reporter gene in cells that underwent cyclic stretch for a period of 24 hours following transfection.8 Since stretch and many cytoskeletal drugs have been shown to increase cellular proliferation, we wanted to make sure that the enhanced expression we measured was not due to enhanced proliferation. When cells plated under similar conditions were either stretched or unstretched, we found no statistical difference in the number of cells between these groups 24 hours later (Figure 5). Intermediate time points of stretch were also examined, and again no increase in cell proliferation was seen (data not shown). Further, as shown in Table 1, the amount of stretch used in our experiments was not sufficient to increase cell death as measured by apoptosis. Similar studies in ATII cells have had similar findings, and have also shown that these levels of stretch do not induce necrosis.16 Taken together, these results suggest that cell proliferation is not required for stretch-increased transfection efficiency.
Figure 5.
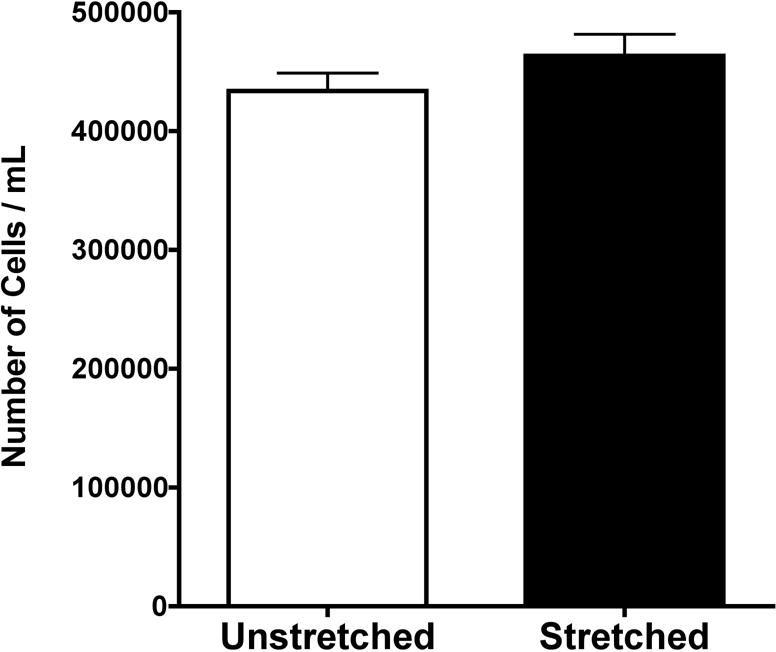
Equibiaxial cyclic stretch does not induce cellular proliferation. Cells were plated at equal concentrations from the same cell stock, and allowed to adhere to the silastic membrane plates for 48–72 hours. The cells were then grown under conditions of stretch (0.5 Hz, with a 10% change in basement membrane area and a duty cycle of 50%) or no stretch for 24 hours, at which point the cells were counted in duplicate (n = 8 for each data set from two separate experiments).
Table 1. Cyclic stretch does not induce apoptosis.
Cells were grown and stretched as described in Figure 1. After 24 hours under the appropriate growth condition, the cells were fixed and assessed for apoptosis by TUNNEL assay and verified by examination of condensed nuclear chromatin with DAPI. Cells were treated for 4 hours with single bolus administrations of hydrogen peroxide at 0.25 mM or 5 mM and served as positive controls.39 Between 700 and1000 cells were counted per condition, in duplicate. The percent of apoptotic cells (± st. dev.) is shown.
| % Apoptosis Detected | ||
|---|---|---|
| Hours of Stretch | ApopTag | DAPI |
| 0 | 0.58 ± 0.06 | 0.55 |
| 0.5 | 0.71 ± 0.03 | 0.66 |
| 1 | 0.37 ± 0.03 | 0.66 |
| 2 | 0.74 ± 0.14 | 0.9 |
| 4 | 0.64 ± 0.09 | 0.81 |
| 24 | 0.61 ± 0.12 | 1.05 |
| H2O2 | ||
| 0.25 mM | 1.21 ± 0.28 | 1.87 |
| 5 mM | 81.36 ± 0.84 | 81.77 |
DISCUSSION
These studies demonstrate that physiologic levels of cyclic stretch in vitro can lead to significant reorganization of the cytoskeletal network and enhanced transfection efficiency. This enhancement appears to be due in part to cytoskeletal reorganization, since stabilization of the cytoskeleton mitigated the stretch-enhanced transgene transfer and expression. Furthermore, this reorganization is more complex than simple depolymerization, as demonstrated by the inability of cytoskeletal depolymerization drugs to enhance gene delivery in the absence of cyclic stretch.
While viral movement through the cytoplasm has been well studied (see Campbell and Hope17 for a comprehensive review), less is known about cytoplasmic movement of naked plasmid DNA toward the nucleus.18 Since plasmid DNA, like any large macromolecule, is hindered by the surrounding dense cytoskeleton from freely diffusing through the cytoplasm,19–21 it would appear that some form of cytoplasmic active transport is necessary for eventual nuclear localization and exogenous gene expression to occur. In the case of viral particles, many use the cytoskeleton and its associated motor proteins for transport through the cytoplasm. Both microtubules and actin filaments exhibit directionality and possess molecular motors that selectively operate in either one direction or the other. For example, HIV, adenovirus, canine parvovirus and HSV-1 all use the microtubule network and the minus-end directed dynein motor complex to transport their genomes toward the microtubule organizing center located adjacent to the nucleus.22–26 In these studies, significant accumulation of viral particles near the nucleus was seen within 1–2 hours. The actin network can also act as a transport conduit, as demonstrated by bacculovirus, which uses the actin network exclusively for transport,27 and HIV, which uses both actin and microtubule networks for cytoplasmic trafficking.22
Non-viral vectors, depending on their mode of transfection, can also use cytoskeletal machinery to travel toward the nucleus. When non-viral vectors are introduced to the cell through liposomal transfections, they are endocytosed and carried via microtubules through the endosomal-lysosomal pathway and are found to accumulate around the nucleus prior to release into the cytoplasm.28,29 With the method of transfection we primarily use in our lab, either electroporation or microinjection, it is unclear as to what pathways plasmids use to traverse the cytoplasm and enter the nucleus. Regardless of the process involved, it is clear that once in the cytoplasm, the plasmid must be quickly shuttled to the nucleus, since nucleases found in the cytoplasm can begin to degrade DNA in a matter of minutes.30,31
In our studies, we found that cyclic equibiaxial stretch of pulmonary epithelial cells caused a reorganization of the actin filaments leading to a network of much smaller fibers that appear to increase their prevalence at the periphery of the cell. To our knowledge, this is the first such report of this type of cytoskeletal reorganization. However, this is also to our knowledge the first report examining cytoskeletal reorganization of epithelial cells undergoing cyclic stretch. When the cytoskeleton was stabilized through the addition of jasplakinolide, we were able to prevent the stretch-induced increase in gene expression. By contrast, destabilization of the actin cytoskeleton through the use of latrunculin B caused a greater difference in gene expression between the stretched and unstretched groups, indicating that actin cytoskeletal rearrangement plays an important role in stretch-induced increases in gene transfer and expression. Verkman and colleagues recently reported that the actin cytoskeleton is the principle determinant for passive diffusion through the cytoplasm, and that disruption of the actin cytoskeleton through the addition of either cytochalasin D or latrunculin B was sufficient in vitro to dissipate the retarding sieving effect seen in the diffusion of large macromolecules or linear DNA fragments.32 This result echoes the earlier findings of Popov and Poo,33 suggesting that disruption or dissipation of the actin cytoskeleton could improve cytoplasmic trafficking of macromolecules. Viruses such as SV40 take a similar approach in disrupting the actin cytoskeleton through the activation of tyrosine kinases to aid in their movement through the cytoplasm.34 Thus, it would seem likely that actin depolymerization plays some role in the stretch-induced increase in gene expression. However, it would seem that simple depolymerization alone is not sufficient for enhanced expression as shown by the fact that the addition of latrunculin B does not increase gene transfer and expression in static cells to the same level that stretching the cells does.
Microtubules have been described in the literature as the compressive load bearing structures of a cell, behaving much like beams whereas actin bears a significant portion of the tensile forces in the cell, behaving more like ropes.4 Previous literature has suggested that microtubule polymerization occurs more freely in stretched cells, possibly due to the decreased compressional load placed on the microtubule.10 Likewise, depolymerization is increased in cells that undergo compressive stresses, with a twofold decrease in polymerized tubulin seen in cells undergoing active compression.15 Unlike the microfilaments, which showed structural reorganization by fluorescence microscopy but did not decrease in number as measured biochemically, microtubules decreased in abundance after as little as 30 minutes of stretch. By 24 hours, polymerized tubulin in stretched cells was less than 20% than in their unstretched counterparts, suggesting that tubulin depolymerization also plays a role in enhanced gene delivery. Furthermore, as seen for actin, stabilization of the microtubule network eliminated any stretch-induced increase in gene expression whereas destabilization of the microtubule network did not statistically increase gene transfer and expression above control levels in unstretched cells, nor did it increase expression to similar levels as those seen with stretch. Taken together, these results indicate that the cytoskeletal networks play an integral role in the trafficking of plasmid DNA through the cytoplasm, and that alterations in the networks through the application of equibiaxial cyclic stretch can lead to enhanced gene transfer and expression.
MATERIALS AND METHODS
Cell Culture and Gene Transfer
All experiments, except where noted, were conducted on A549 cells, (ATCC, Manassas, VA) a human lung adenocarcinoma cell line. Cells were grown in high glucose DMEM supplemented with 10% fetal bovine serum, kanamyacin, and antibiotic/antimycotic solution (Invitrogen, Carlsbad, CA). Cells were passaged every three to five days and maintained at 37°C with 5% CO2.
For transfection experiments, cells were grown for 48–72 hours on Pronectin treated BioFlex culture plates (Flexcell International, Hillsborough, NC) or on regular tissue-culture treated 6-well dishes. Cells were electroporated at 80–90% confluency using a BTX ECM 830 Square Wave Electroporator coupled with a Petri-Pulser PP35-2P electrode (Harvard Apparatus, Holliston, MA) using a single 10 ms 160V square wave to introduce plasmid DNA into the adherent cells. The reporter plasmid used was a pCMV-LUX-DTS construct that uses the CMV immediate early promoter/enhancer to drive luciferase expression.35 Cells were lysed using Promega Lysis Buffer and luciferase expression was measured using the Promega Luciferase Assay System (Promega, Madison, WI) as previously described.35
All cytoskeletal drugs were added immediately after electroporation and prior to stretching. Jasplakinolide (Calbiochem, La Jolla, CA) and Latrunculin B (Sigma-Aldrich, St. Louis, MO) were dissolved in DMSO and added at a final concentration of 0.125 μM and 2.5 μM respectively. Nocodazole and Taxol (Calbiochem) were dissolved in DMSO and added at a final concentration of 6.6 μM and 10 μM respectively. DMSO vehicle was added at equal volumes for all controls.
Equibiaxial Cyclic Stretch
For all stretch related experiments, A549 cells were plated on Pronectin treated BioFlex culture plates. After allowing for attachment for 48–72 hours, the cells were stretched using 25mm BioFlex loading stations with a 10% membrane surface area change at 30 cycles per minute and a 50% duty cycle. These parameters are the same as previously reported,8 and the 25mm loading stations ensure an equibiaxial stretch in the radial and circumferential directions over the width of the loading post.
Protein extraction and quantitation
Protein extracts for either actin or tubulin were prepared as previously described.36,37 All extractions, washes and fixations were conducted with buffers warmed to 37°C unless otherwise noted. For actin extraction, the cells were washed once in PBS and removed from the membrane in PBS by scraping. The cells were centrifuged for 1 minute at 400×g and resuspended in 20 pellet volumes of extraction buffer (0.1% Nonidet P-40, 5 mM KH2PO4, 27mM Na2HPO4 (pH 7.2), 2 mM MgSO4, 2 mM ethylene glycol-bis(β-aminoethyl ether)N, N, N′, N′-tetraacetic acid (EGTA), 0.2 mM ATP, 0.5 mM dithiotheitol, 2 M glycerol, and protease inhibitors (Complete protease inhibitor cocktail tablet; Roche Applied Science, Indianapolis, IN)). After 15 minutes of agitation at room temperature, the insoluble fraction was centrifuged for one minute at 400×g. The supernatant, containing the G-actin fraction, was removed and the pellet was resuspended in the volume of the initial extraction buffer volume.
For tubulin extractions, the cells were washed once in PBS, followed by two 15-minute incubations with microtubule stabilization buffer (MTSB; 0. l M PIPES, pH 6.75, 1 mM EGTA, l mM MgSO4, 2 M glycerol, and protease inhibitors) with or without 0.1% Triton X-100. Cells incubated in the MTSB lacking Triton X-100 contained both polymerized and depolymerized tubulin, while those cells incubated in MTSB with Triton X-100 contained only polymerized tubulin. The cells were then incubated in lysis buffer (25 mM Tris-HCI, pH 7.4, 0.4 M NaC1, 0.5% SDS) for 5 minutes prior to scraping. Cell lysates were boiled for 3 minutes, centrifuged at 12,000×g for 2 minutes, and the resulting supernatant was transferred to a new tube and ß-mercaptoethanol was added to 0.1%. Lysates were boiled for an additional 3 minutes and stored at −70°C prior to separation on SDS-PAGE gels.
All extracted samples were mixed with SDS-PAGE sample buffer (112.5 mM Tris-HCl, pH 6.8, 10% β-mercaptoethanol, 3.6% SDS, 1.8% glycerol, 0.001% bromophenol blue) such that equal amounts of total protein were loaded for each fraction. The samples were boiled for 3 minutes and separated by 12% SDS-PAGE gels according to the methods previously described by Lugtenberg.38 Proteins were transferred to nitrocellulose and probed using anti-actin (1:1000, Sigma, Catalog #A-2066) or anti-α-tubulin (1:1000, Sigma, Catalog #T-9026) in PBS with 5% fat-free powdered milk. Blots were developed with a chemiluminescence detection kit, and autoradiograms were digitized prior to densitometry using ImageJ (NIH, Bethesda, MD). All samples were run in duplicate and were from at least 3 independent experiments.
Immunofluorescence Imaging
After all treatment regimes, cells were washed twice in 37°C PBS followed by a 10 minute incubation in a 1× fixation/permeabilization buffer (60 mM PIPES, 25 mM HEPES, 10 mM EGTA, 3 mM MgCl2, 0.2% Triton X-100, and 3.7% paraformaldehyde) at 37°C. Cells were washed 3 times in PBS and blocked for 1 hour with PBS containing 1 mg/mL BSA. After blocking, the cells were incubated for 1 hour at 37°C with an anti-α-tubulin primary antibody (1:1000, Sigma) in PBS with 0.1 mg/mL BSA, washed 3 times in PBS, and reacted for 30 minutes at 37°C with Alexa 488 conjugated secondary antibody (1:200, Molecular Probes, Eugene, OR) and Alexa 546 conjugated phalloidin (1:200, Molecular Probes) in PBS with 0.1% BSA. Following a second set of washes in PBS, the silastic membranes were excised from the culture plates, placed face up on a microscope slide, and covered with a coverslip. All images were acquired in OpenLab (Improvision, Lexington, MA) with a Hamamatsu Orca II-ER camera attached to Leica DRMX-2 inverted fluorescent microscope using a 100x oil-immersion objective.
Apoptosis Measurements
After all treatment regimes, cells were either prepared for TUNNEL assessment per the manufacturers instructions (ApopTag Red In situ Apoptosis Detection Kit, Chemicon, Temecula, CA) or immediately fixed for DAPI staining and visualization. For those cells that were prepared for DAPI staining, a 1:4 dilution of 37% formalin was added to the growth media and the cells were incubated overnight at 4°C. The next day, the cells were gently washed once with dH2O, and 0.5 ng/mL DAPI in PBS was added to the cells for 10 minutes. The cells were then washed with dH2O and the membranes were placed on a glass coverslip to allow for microscopic visualization. With both methods, 700–1000 cells were counted per condition using a 40× objective.
Statistical Analysis
All statistical analysis was performed with Prism software (GraphPad Software, San Diego, CA), using the Mann-Whitey Wilcoxon test for paired, non-parametric samples. Statistically significant results were denoted at a p value of less than or equal to 0.05.
Acknowledgments
We would like to thank Chris Capaccio, Joshua Gasiorowski, Erin Vaughan, and Jennifer Young for their helpful discussions and advice. This work was supported in part by grants HL71643 (DAD and MRG) and HL076139 (RCG) from the NIH, and funds from the Institute for BioNanotechnology in Medicine of Northwestern University.
References
- 1.Fung YC. Biomechanics: mechanical properties of living tissues. Springer-Verlag; New York: 1993. [Google Scholar]
- 2.Ingber DE. Tensegrity: the architectural basis of cellular mechanotransduction. Annu Rev Physiol. 1997;59:575–599. doi: 10.1146/annurev.physiol.59.1.575. [DOI] [PubMed] [Google Scholar]
- 3.Ingber DE. Opposing views on tensegrity as a structural framework for understanding cell mechanics. J Appl Physiol. 2000;89:1663–1670. doi: 10.1152/jappl.2000.89.4.1663. [DOI] [PubMed] [Google Scholar]
- 4.Wang N, et al. Mechanical behavior in living cells consistent with the tensegrity model. Proc Natl Acad Sci U S A. 2001;98:7765–7770. doi: 10.1073/pnas.141199598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang N, Butler JP, Ingber DE. Mechanotransduction across the cell surface and through the cytoskeleton. Science. 1993;260:1124–1127. doi: 10.1126/science.7684161. [DOI] [PubMed] [Google Scholar]
- 6.Tschumperlin DJ, Oswari J, Margulies AS. Deformation-induced injury of alveolar epithelial cells. Effect of frequency, duration, and amplitude. Am J Respir Crit Care Med. 2000;162:357–362. doi: 10.1164/ajrccm.162.2.9807003. [DOI] [PubMed] [Google Scholar]
- 7.Tschumperlin DJ, Margulies AS. Alveolar epithelial surface area-volume relationship in isolated rat lungs. J Appl Physiol. 1999;86:2026–2033. doi: 10.1152/jappl.1999.86.6.2026. [DOI] [PubMed] [Google Scholar]
- 8.Taylor W, et al. The effects of cyclic stretch on gene transfer in alveolar epithelial cells. Mol Ther. 2003;7:542–549. doi: 10.1016/s1525-0016(03)00041-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Putnam AJ, Schultz K, Mooney DJ. Control of microtubule assembly by extracellular matrix and externally applied strain. Am J Physiol Cell Physiol. 2001;280:C556–564. doi: 10.1152/ajpcell.2001.280.3.C556. [DOI] [PubMed] [Google Scholar]
- 10.Putnam AJ, et al. Microtubule assembly is regulated by externally applied strain in cultured smooth muscle cells. J Cell Sci. 1998;111(Pt 22):3379–3387. doi: 10.1242/jcs.111.22.3379. [DOI] [PubMed] [Google Scholar]
- 11.Hayakawa K, Sato N, Obinata T. Dynamic reorientation of cultured cells and stress fibers under mechanical stress from periodic stretching. Exp Cell Res. 2001;268:104–114. doi: 10.1006/excr.2001.5270. [DOI] [PubMed] [Google Scholar]
- 12.Li C, Xu Q. Mechanical stress-initiated signal transductions in vascular smooth muscle cells. Cell Signal. 2000;12:435–445. doi: 10.1016/s0898-6568(00)00096-6. [DOI] [PubMed] [Google Scholar]
- 13.Chaqour B, Howard PS, Richards CF, Macarak EJ. Mechanical stretch induces platelet-activating factor receptor gene expression through the NF-kappaB transcription factor. J Mol Cell Cardiol. 1999;31:1345–1355. doi: 10.1006/jmcc.1999.0967. [DOI] [PubMed] [Google Scholar]
- 14.Park JM, et al. AP-1 mediates stretch-induced expression of HB-EGF in bladder smooth muscle cells. Am J Physiol. 1999;277:C294–301. doi: 10.1152/ajpcell.1999.277.2.C294. [DOI] [PubMed] [Google Scholar]
- 15.Dennerll TJ, et al. Tension and compression in the cytoskeleton of PC-12 neurites. II: Quantitative measurements. J Cell Biol. 1988;107:665–674. doi: 10.1083/jcb.107.2.665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hammerschmidt S, et al. Apoptosis and necrosis induced by cyclic mechanical stretching in alveolar type II cells. Am J Respir Cell Mol Biol. 2004;30:396–402. doi: 10.1165/rcmb.2003-0136OC. [DOI] [PubMed] [Google Scholar]
- 17.Campbell EM, Hope TJ. Role of the cytoskeleton in nuclear import. Adv Drug Deliv Rev. 2003;55:761–771. doi: 10.1016/s0169-409x(03)00049-8. [DOI] [PubMed] [Google Scholar]
- 18.Zhou R, Geiger RC, Dean DA. Intracellular trafficking of nucleic acids. Expert Opinion in Drug Delivery. 2004;1:127–140. doi: 10.1517/17425247.1.1.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Luby-Phelps K. Cytoarchitecture and physical properties of cytoplasm: volume, viscosity, diffusion, intracellular surface area. Int Rev Cytol. 2000;192:189–221. doi: 10.1016/s0074-7696(08)60527-6. [DOI] [PubMed] [Google Scholar]
- 20.Seksek O, Biwersi J, Verkman AS. Translational diffusion of macromolecule-sized solutes in cytoplasm and nucleus. J Cell Biol. 1997;138:131–142. doi: 10.1083/jcb.138.1.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lukacs GL, et al. Size-dependent DNA mobility in cytoplasm and nucleus. J Biol Chem. 2000;275:1625–1629. doi: 10.1074/jbc.275.3.1625. [DOI] [PubMed] [Google Scholar]
- 22.McDonald D, et al. Visualization of the intracellular behavior of HIV in living cells. J Cell Biol. 2002;159:441–452. doi: 10.1083/jcb.200203150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Suomalainen M, et al. Microtubule-dependent plus- and minus end-directed motilities are competing processes for nuclear targeting of adenovirus. J Cell Biol. 1999;144:657–672. doi: 10.1083/jcb.144.4.657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Suikkanen S, et al. Exploitation of microtubule cytoskeleton and dynein during parvoviral traffic toward the nucleus. J Virol. 2003;77:10270–10279. doi: 10.1128/JVI.77.19.10270-10279.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Leopold PL, et al. Dynein- and microtubule-mediated translocation of adenovirus serotype 5 occurs after endosomal lysis. Hum Gene Ther. 2000;11:151–165. doi: 10.1089/10430340050016238. [DOI] [PubMed] [Google Scholar]
- 26.Sodeik B, Ebersold MW, Helenius A. Microtubule-mediated transport of incoming herpes simplex virus 1 capsids to the nucleus. J Cell Biol. 1997;136:1007–1021. doi: 10.1083/jcb.136.5.1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.van Loo ND, et al. Baculovirus infection of nondividing mammalian cells: mechanisms of entry and nuclear transport of capsids. J Virol. 2001;75:961–970. doi: 10.1128/JVI.75.2.961-970.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Coonrod A, Li FQ, Horwitz M. On the mechanism of DNA transfection: efficient gene transfer without viruses. Gene Ther. 1997;4:1313–1321. doi: 10.1038/sj.gt.3300536. [DOI] [PubMed] [Google Scholar]
- 29.Zabner J, et al. Cellular and molecular barriers to gene transfer by a cationic lipid. J Biol Chem. 1995;270:18997–19007. doi: 10.1074/jbc.270.32.18997. [DOI] [PubMed] [Google Scholar]
- 30.Lechardeur D, et al. Metabolic instability of plasmid DNA in the cytosol: a potential barrier to gene transfer. Gene Ther. 1999;6:482–497. doi: 10.1038/sj.gt.3300867. [DOI] [PubMed] [Google Scholar]
- 31.Pollard H, et al. Ca2+-sensitive cytosolic nucleases prevent efficient delivery to the nucleus of injected plasmids. J Gene Med. 2001;3:153–164. doi: 10.1002/jgm.160. [DOI] [PubMed] [Google Scholar]
- 32.Dauty E, Verkman AS. Actin Cytoskeleton as the Principal Determinant of Size-dependent DNA Mobility in Cytoplasm: A NEW BARRIER FOR NON-VIRAL GENE DELIVERY. J Biol Chem. 2005;280:7823–7828. doi: 10.1074/jbc.M412374200. [DOI] [PubMed] [Google Scholar]
- 33.Popov S, Poo MM. Diffusional transport of macromolecules in developing nerve processes. J Neurosci. 1992;12:77–85. doi: 10.1523/JNEUROSCI.12-01-00077.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pelkmans L, Puntener D, Helenius A. Local actin polymerization and dynamin recruitment in SV40-induced internalization of caveolae. Science. 2002;296:535–539. doi: 10.1126/science.1069784. [DOI] [PubMed] [Google Scholar]
- 35.Vacik J, Dean BS, Zimmer WE, Dean DA. Cell-specific nuclear import of plasmid DNA. Gene Ther. 1999;6:1006–1014. doi: 10.1038/sj.gt.3300924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Caron JM, Jones AL, Kirschner MW. Autoregulation of tubulin synthesis in hepatocytes and fibroblasts. J Cell Biol. 1985;101:1763–1772. doi: 10.1083/jcb.101.5.1763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Forte JG, et al. State of actin in gastric parietal cells. Am J Physiol. 1998;274:C97–104. doi: 10.1152/ajpcell.1998.274.1.C97. [DOI] [PubMed] [Google Scholar]
- 38.Lugtenberg B, et al. Electrophoretic resolution of the “major outer membrane protein” of Escherichia coli K12 into four bands. FEBS Lett. 1975;58:254–258. doi: 10.1016/0014-5793(75)80272-9. [DOI] [PubMed] [Google Scholar]
- 39.Kazzaz JA, et al. Cellular oxygen toxicity. Oxidant injury without apoptosis. J Biol Chem. 1996;271:15182–15186. doi: 10.1074/jbc.271.25.15182. [DOI] [PubMed] [Google Scholar]