

Abstract
That similar words can prime one another isn’t news. However, recently this phenomenon has been exploited to make inferences about conceptual representations. What types of similarity matter? While there is evidence that similarity in function, shape, and even manipulation figure in the organization of semantic memory, evidence for color similarity is sparse. This is surprising: intuition suggests that color is a prominent feature of many concepts. We report an experiment that clarifies this puzzle and illustrates the dynamic nature of conceptual representations. We report color priming (e.g., the word emerald primes the word cucumber) in subjects who have previously completed a color-Stroop task. Notably, the size of the Stroop effect predicts the size of the priming effect across subjects. When the order of tasks is reversed, priming is eliminated. By highlighting that extrinsic and intrinsic factors can both influence conceptual activation, these findings have implications for theories of semantic memory.
Keywords: semantic representations, semantic features, semantic attributes, context, color, individual differences, attention, embodied cognition
Introduction
If you are searching your refrigerator to find a cucumber, its color (green) is a useful feature. In contrast, color is less relevant while slicing the cucumber. Does the salience of color in the search context make cucumbers, in a sense, more similar to other green objects than they are in a slicing context? That is, does the set of “similar” objects change as a function of which properties are contextually relevant? How about after you’ve found the cucumber—does color “linger”, remaining salient even though it’s no longer important to the task? Here we explore whether conceptual activation (i.e., which object properties are activated when) is influenced by task relevance. We also explore individual differences in this conceptual activation.
The meaning of an object is often characterized as a pattern that is distributed across semantic features (e.g., Masson, 1995). One consequence of this distributed architecture is that relationships among concepts can be captured via overlapping patterns. Another is that attention can be focused on specific (e.g., contextually relevant) properties of a representation (e.g., attention can be focused on greenness when searching for a cucumber). Thus the extent to which an object concept activates a related concept may depend on both attention to relevant properties, and on the extent to which those properties are shared between the concepts.
Behavioral evidence in support of representational overlap comes primarily from semantic priming studies demonstrating that the word cucumber, for example, primes conceptually related things like mushrooms (e.g., Fischler, 1977). Such studies have also demonstrated priming for perceptual relationships. For example, cherry primes ball, due to similarity in shape (Schreuder et al., 1986; Taylor 2005), and piano primes typewriter, due to similarity in the way the fingers move over each (Myung et al., 2005). Interestingly, however, there has been little, if any, evidence of an analogous effect for color (Huettig & Altmann, 2010; cf. Taylor & Heindel, 2004).
Why might color be different? One possibility is that color is less essential to object knowledge than other features, and is only encoded when it is “diagnostic” – i.e., has high information value (Tanaka & Presnell, 1999; but cf. Rossion & Pourtois, 2004), or only for objects that need to be recognized in the context of similarly shaped objects (Price & Humphreys, 1989). And even these objects can usually be recognized in black and white, suggesting that color may still be less important than other features (e.g., Biederman, 1987). If true, color may constitute only a small portion of an object’s representation. This may explain why evidence of conceptual overlap based on color has been scarce (cf. Joseph, 1997). It may also make color a good case for exploring the extent to which conceptual representations can be dynamically affected by context: Color may be more sensitive to context than other, more important features (e.g., function) which, because of their importance, may be less variable in the degree to which they become active.
In the current work, we used semantic priming to assess whether objects that share color partially activate each other, and if so, whether this activation is context-dependent. We manipulated the extent to which attention is focused on color by having participants complete a standard color Stroop task before or after the priming task. This allowed us to test whether the experience of having recently focused attention on color (in the Stroop task) influences the degree of color activation in a subsequent priming task. We also tested whether individual differences in susceptibility to Stroop interference predicted individual differences in color priming. We show that: 1) Rather than having a fixed activation profile, conceptual representations can be dynamically affected by context, 2) Contextually-modulated attention to color varies across individuals, and importantly, 3) Context’s influence can “linger” beyond when it is relevant.
Methods
Participants
120 participants (66 female) from the University of Pennsylvania community received $10/hour or course credit for participating. All were native English speakers and had normal or corrected-to-normal color vision. 60 participants performed the Stroop task prior to the priming task (“Stroop first” order). The remainder performed the priming task first (“priming first” order).
Materials & Procedure
Stroop task
Participants saw written color words (red, yellow, green, blue, black) and were instructed to press the key (labeled in black ink) that corresponded to the font color. The word remained on the screen until the response. The inter-trial interval was 500ms. There were 100 trials. In half, the word and font color were congruent. Trial order was randomized.
Animal judgment task
We created 60 color-related prime-target pairs (e.g., emerald-cucumber), and 60 control pairs using the same target but a prime unrelated in color (e.g., pendant-cucumber). The task was a semantic judgment (animal or not?). No critical targets were animals. Related and control primes were matched for animacy, frequency, number of letters, number of syllables, and similarity as computed via Latent Semantic Analysis. Two separate groups of 30 participants (who did not participate in the priming study) confirmed via 1–7 ratings that color-related pairs were more likely to be the same color (M=5.9) than control pairs (M=2.3), and that color was more important for recognizing objects constituting color-related pairs (M=4.9) than for recognizing control primes (M=2.8).
Critical stimuli were divided into two counterbalanced lists containing 30 color-related, 30 control, and 60 filler trials (all fillers had animal targets, e.g., pants-worm). Each subject saw one list, in a fixed random order. There were 2 practice and 6 filler trials at the beginning of the experiment.
Participants were presented with the prime (300ms), a mask (50ms) and then the target until they responded, pressing “A” or “I” respectively if the target was/was not an animal (Figure 1). To ensure that participants attended to the prime, in 12% of trials, after their response, a prompt appeared asking if the prime was an animal. A post-test questionnaire asked what participants thought the experiment was about. Two noticed that the 1st and 2nd words occasionally referred to things of the same color. The pattern was unchanged with these participants removed. See supplementary materials for additional details and a full list of stimuli.
Figure 1.
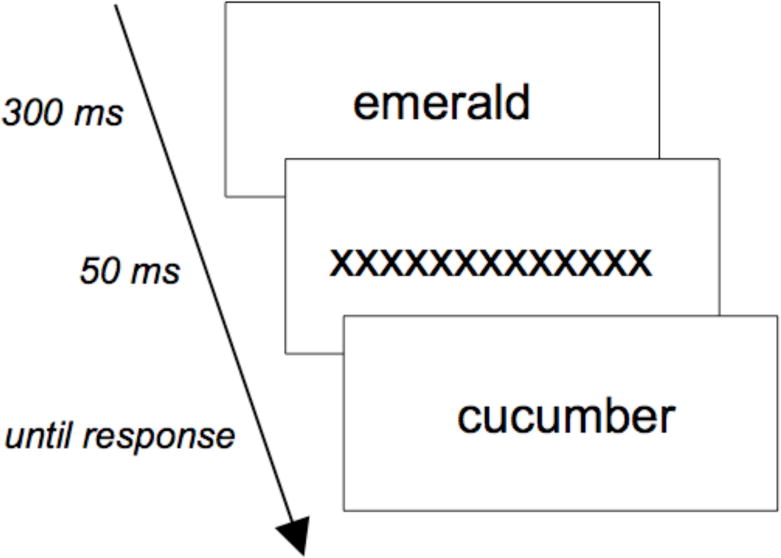
Structure of priming task. Participants judged whether the 2nd (target) word was an animal.
Results
For the Stroop task (Figure 2), ANOVA on reaction times (correct only) revealed a significant main effect of congruency F(1,118) = 189.8, p < .001, no effect of task order, and no interaction between congruency and order (both Fs < 1). The pattern for accuracy was the same.
Figure 2.
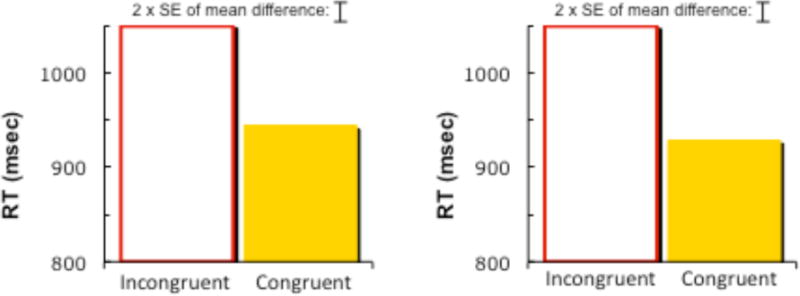
Response times (correct only) in Stroop task when performed before animal judgment (left panel), and when performed after animal judgment (right panel).
For the priming task (Figure 3), ANOVA on reaction times (correct only) revealed that across both orders there was a non-significant main effect of relatedness F1(1,118) = 3.6, p = .06, F2(1,581) = 1.2, p = .28. The effect of task order was not significant by subjects (F1<1), but was by items F2(1,58) = 15.7, p < .01. Critically, the interaction between relatedness and task order was significant by both subjects F1(1,118) = 3.9, p = .05, and items F2(1,58) = 3.9, p = .05. Planned comparisons showed that in the Stroop-first order, responses were faster in color-related than control trials, t1(59) = 2.6 p = .01, although the difference was only marginally significant by items t2(58) = 1.7, p = .09. In the priming-first order, there was no difference between color-related and control trials t1(59) = −0.1, p = .96, t2(58) = 0.0, p = .98.
Figure 3.
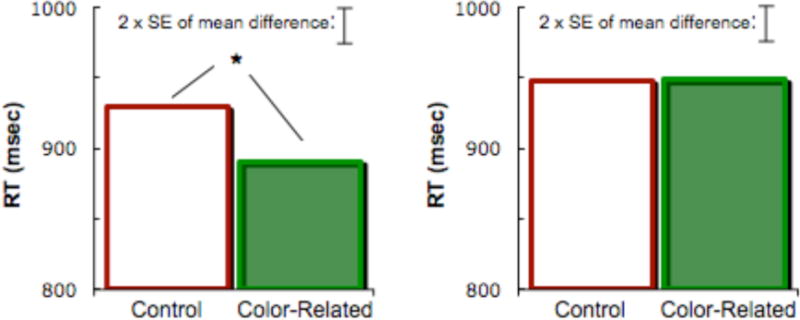
Response times (correct only) in animal judgment task when performed after Stroop task (left panel), and when performed before Stroop task (right panel).
Accuracy in the priming task followed the same pattern: No main effect of relatedness F1(1,118) = 2.4, p = .12, F2(1,58) = 1.7, p = .20, or task order, F1<1, F2(1,58) = 2.0, p = .17. However, the critical interaction between relatedness and task order was again present, F1(1,118) = 4.8, p = .03, F2(1,58) = 5.3, p = .02. Planned comparisons showed that color-related trials were responded to more accurately in the Stroop-first order, t1(59) = 2.7 p = .01, t2(58) = 2.2, p = .03, but not in the priming-first order, t1(59) = −0.4, p = .67, t2(58) = −0.3, p = .73.
We also tested whether the size of the Stroop interference effect for a given participant (RT incongruent – RT congruent / RT incongruent) was predictive of the size of the color priming effect (RT unrelated – RT related / RT unrelated). We found that in the Stroop-first order, Stroop interference was positively correlated with color priming r = .38, p = .003. However, when the order was reversed, there was no correlation r = .04, p = .76 (Figure 4). The difference between the correlations was significant, z = 1.9, p = .05.
Figure 4.
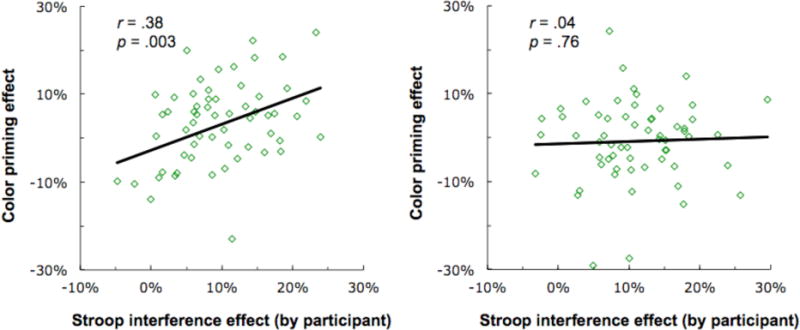
Correlation between congruency effect and priming effect when animal judgment task performed after Stroop (left panel), and when animal judgment task performed before Stroop (right panel).
Discussion
When attention has first been drawn to color via a standard Stroop task, reading the name of an object primes other objects that share the same diagnostic color – an effect that has not been previously reported. Further, an individual’s susceptibility to Stroop interference predicted the amount of priming. Importantly, priming occurred despite the fact that color was irrelevant to the immediate task—animal judgment of written words that were presented in black type. In contrast, when attention was not previously drawn to color, no priming was observed. Hence, the extent to which color information about an object is activated appears to depend not only on long-term, object-related factors (i.e., is color relevant in general for the identification of this object?) as shown by Tanaka & Presnell (1999), but also on shorter-term, context-related factors (i.e., has attention been recently focused on color?). There are parallels to context-dependent activation of color at the cellular level: Recordings from inferotemporal neurons in monkeys showed that in some cells, firing rate to color stimuli depended upon whether color was task-relevant (Fuster & Jervey, 1981). We find further that the influence of these context-related factors appears to be dependent on individual factors (which we return to below).
Previous work has demonstrated that altering the immediate context in which a word is presented can influence which aspects of its representation are active. For example, if a cucumber is mentioned in a sentence about eating it, you might access how crunchy it is (cf. Barclay et al., 1974); and sentence context can influence the activation of its color (Connell, 2007; Connell & Lynott, 2009). Similarly, immediate context can influence the kind of motor information that is activated by a word (van Dam, Rueschemeyer, Lindemann & Bekkering, 2010). However, to our knowledge, only one prior study has demonstrated that non-immediate, but recent experience can “linger” enough to influence the activation of semantic features in a subsequent task: Semantic priming based on shape similarity is enhanced if, prior to the priming task, participants first make perceptual judgments about the objects to which the to-be-primed words refer (Pecher et al., 1998). The present work extends this finding to color but importantly, also demonstrates that the effect is general: it is neither object-specific (only color names were used in the Stroop task), nor isolated to the colors presented in the Stroop task—the correlation between Stroop interference and priming was the same for items whose colors and were or were not included in the Stroop task (r=.31 in both cases). Thus, general attention to color is sufficient to alter subsequent processing. An open question is whether the influence of recent experience can generalize across objects for features besides color. For example, after chopping cucumbers (a task that focuses attention on manual action), would piano activate typewriter (as shown by Myung et al., 2005) more than usual? Or is color uniquely sensitive to context because it is rarely essential for object identification, and thus benefits more than other features from contextual enhancement (for discussion, see Barsalou, 1982)?
In addition to demonstrating color priming and its dependence on context, we also observed that when the Stroop task was performed prior to the priming task there was a positive correlation between participants’ Stroop incongruency and color priming effects. What might drive this correlation? One possibility concerns individual differences in selective attention: If the ability to selectively attend to a word’s ink color (while ignoring its meaning) in the Stroop task is due to a generally high capacity for selective attention, this could manifest, in the priming task, as selective attention to features most relevant for judging animal status. This would result in less activation of, and hence less priming from, the object’s other features (like color). Interestingly, this kind of account would suggest that Stroop performance (or other measures of cognitive control) may correlate with semantic priming effects in general, rather than color priming specifically (cf. Kiefer, Ahlegian & Spitzer, 2005).
Another possibility is that the correlation reflects individual differences in the content of representations: Information processing preferences can produce differences in the degree to which people typically attend to or perceive color (Hsu et al., 2011). This should lead to differences in how strongly color is encoded, which would influence how strongly associated color becomes both with names of color words (e.g., green) and with names of objects (e.g., cucumber). Strong associations would cause more interference from color words in the Stroop task, and more activation of color from object names in the priming task.
Future work can explore whether the observed correlation reflects individual variability in a general capacity to selectively attend, or in attention to color specifically. In either case though, context must moderate these individual differences—for when the Stroop task was performed after the priming task, no correlation (and also no priming) was observed.
With respect to models of semantic memory, by showing that objects that share color have overlapping representations and that attention can be focused on specific parts of an object’s representation, our findings provide support for distributed models (e.g., Masson, 1995; McRae de Sa & Seidenberg, 1997; and see also Schyns, Goldstone & Thibaut, 1998). Further, because color is a perceptual feature, the finding that color-related objects activate one another provides support for a critical prediction of sensorimotor-based distributed models in particular (e.g., Warrington & McCarthy, 1987; Barsalou, 1999). These models claim that semantic information about an object is represented in the neural substrate that is invoked when it is perceived and/or interacted with, and recent fMRI studies suggest that this is true for color (Simmons et al., 2007; Hsu et al., 2011). Thus, sensorimotor-based models require that objects that share perceptual features such as color have overlapping representations, but until now, priming based on shared color has not been reported.
Traditionally, when studying semantic representations (and in fact, in cognitive psychology more broadly), it has been assumed that only effects that can be demonstrated across a variety of contexts should be considered informative with regards to the architecture of the system being investigated. Yet our findings show that rather than being fixed, the prominence of a concept’s features can change from one context to another, and within a given context, from one person to another. We contend that instead of being “nuisance noise”, this variability is an important consequence of semantic memory’s distributed architecture—wherein because attention can be focused on specific (e.g., contextually relevant) properties of a representation, conceptual representations are dynamic (Hoenig et al., 2008; see Humphreys & Forde, 2001; Spivey, 2007 and Kiefer & Martens, 2010 for discussion).
For decades, semantic priming has been a major source of evidence in the way we think about semantic knowledge. The current work highlights that in both the way we interpret semantic priming, and the way we think of concepts, we must take the influence of context, and individual susceptibility to context, into account; critically, this context does not have to be immediate—it can also include “lingering” traces of recent experience: Colorless green ideas can prime—in the right context, for the right people.
Supplementary Material
Acknowledgments
Supported by NIH grant R01MH70850 to S.L.T-S. and Kirschstein NRSA (F32HD051364) to E.Y. We thank Katherine White, Jesse Hochstadt, Gerry Altmann and Thompson-Schill lab members for helpful comments. Portions of this research were presented at the Annual Meeting of the Psychonomic Society, November 2010.
Footnotes
One item elicited response times and errors more than 3SD higher than the rest and was excluded from analysis.
References
- Barclay JR, Bransford JD, Franks JJ, McCarrell NS, Nitsch KE. Comprehension and semantic flexibility. Journal of Verbal Learning and Verbal Behavior. 1974;13:471–481. [Google Scholar]
- Barsalou LW. Perceptual symbol systems. Behavioral and Brain Sciences. 1999;22:577–660. doi: 10.1017/s0140525x99002149. [DOI] [PubMed] [Google Scholar]
- Barsalou LW. Context-independent and context-dependent information in concepts. Memory & Cognition. 1982;10:82–93. doi: 10.3758/bf03197629. [DOI] [PubMed] [Google Scholar]
- Biederman I. Recognition-by-Components: A Theory of Human Image Understanding. Psychological Review. 1987;94:115–147. doi: 10.1037/0033-295X.94.2.115. [DOI] [PubMed] [Google Scholar]
- Connell L. Representing object colour in language comprehension. Cognition. 2007;102:476–485. doi: 10.1016/j.cognition.2006.02.009. [DOI] [PubMed] [Google Scholar]
- Connell L, Lynott D. Is a bear white in the woods? Parallel representation of implied object color during language comprehension. Psychonomic Bulletin & Review. 2009;16:573–577. doi: 10.3758/PBR.16.3.573. [DOI] [PubMed] [Google Scholar]
- Fischler I. Semantic facilitation without association in a lexical decision task. Memory and Cognition. 1977;5:699–716. doi: 10.3758/BF03197580. [DOI] [PubMed] [Google Scholar]
- Flores d’Arcais G, Schreuder R, Glazenborg G. Semantic activation during recognition or referential words. Psychological Research. 1985;47:39–49. [Google Scholar]
- Fuster JM, Jervey JP. Inferotemporal neurons distinguish and retain behaviorally relevant features of visual stimuli. Science. 1981;212(4497):952–955. doi: 10.1126/science.7233192. [DOI] [PubMed] [Google Scholar]
- Hoenig K, Sim E-J, Bochev V, Herrnberger B, Kiefer M. Conceptual flexibility in the human brain: dynamic recruitment of semantic maps from visual, motion and motor-related areas. Journal of Cognitive Neuroscience. 2008;20:1799–1814. doi: 10.1162/jocn.2008.20123. [DOI] [PubMed] [Google Scholar]
- Hsu NS, Kraemer DJM, Oliver RT, Schlichting ML, Thompson-Schill SL. Color, Context, and Cognitive Style: Variations in Color Knowledge Retrieval as a Function of Task and Subject Variables. Journal of Cognitive Neuroscience. 2011 Jan 25; doi: 10.1162/jocn.2011.21619. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- Huettig F, Altmann GTM. Looking at anything that is green when hearing ‘frog’ – How object surface colour and stored object colour knowledge influence language-mediated overt attention. Quarterly Journal of Experimental Psychology. 2010 doi: 10.1080/17470218.2010.481474. [DOI] [PubMed] [Google Scholar]
- Humphreys GW, Forde EME. Hierarchies, similarity, and interactivity in object recognition: “Category-specific” neuropsychological deficits. Behavioral and Brain Sciences. 2001;24:453–509. [PubMed] [Google Scholar]
- Joseph J. Color Processing in Object Verification. Acta Psychologica. 1997;97:95–127. doi: 10.1016/s0001-6918(97)00026-7. [DOI] [PubMed] [Google Scholar]
- Kiefer M, Ahlegian M, Spitzer M. Working Memory Capacity, Indirect Semantic Priming, and Stroop Interference: Pattern of Interindividual Prefrontal Performance Differences in Healthy Volunteers. Neuropsychology. 2005;19(3):332–344. doi: 10.1037/0894-4105.19.3.332. [DOI] [PubMed] [Google Scholar]
- Kiefer M, Martens U. Attentional Sensitization of Unconscious Cognition: Task Sets Modulate Subsequent Masked Semantic Priming. Journal of Experimental Psychology: General. 2010;139(3):464–489. doi: 10.1037/a0019561. [DOI] [PubMed] [Google Scholar]
- Masson MEJ. A distributed memory model of semantic priming. Journal of Experimental Psychology: Learning, Memory and Cognition. 1995;21:3–23. [Google Scholar]
- McRae K, de Sa VR, Seidenberg MS. On the nature and scope of featural representations of word meaning. Journal of Experimental Psychology: General. 1997;126:99–130. doi: 10.1037//0096-3445.126.2.99. [DOI] [PubMed] [Google Scholar]
- McRae K, Cree GS, Seidenberg MS, McNorgan C. Semantic feature production norms for a large set of living and nonliving things. Behavior Research Methods. 2005;37:547–559. doi: 10.3758/bf03192726. [DOI] [PubMed] [Google Scholar]
- Pecher D, Zeelenberg R, Raaijmakers JGW. Does pizza prime coin? Perceptual priming in lexical decision and pronunciation. Journal of Memory and Language. 1998;38:401–418. [Google Scholar]
- Price CJ, Humphreys GW. The effects of surface detail on object categorization and naming. Quarterly Journal of Experimental Psychology A. 1989;41(4):797–827. doi: 10.1080/14640748908402394. [DOI] [PubMed] [Google Scholar]
- Schreuder R, Flores D’Arcais GB, Glazenborg G. Effects of perceptual and conceptual similarity in semantic priming. Psychological Research. 1984;45(4):339–354. [Google Scholar]
- Schyns PG, Goldstone RL, Thibaut JP. The development of features in object concepts. Behavioral and Brain Sciences. 1998;21:1–54. doi: 10.1017/s0140525x98000107. [DOI] [PubMed] [Google Scholar]
- Simmons W, Ramjee V, Beauchamp M, McRae K, Martin A, Barsalou L. A common neural substrate for perceiving and knowing about color. Neuropsychologia. 2007;45:2802–2810. doi: 10.1016/j.neuropsychologia.2007.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spivey MJ. The continuity of mind. New York: Oxford University Press; 2007. [Google Scholar]
- Tanaka JM, Presnell LM. Perception & Psychophysics. 1999;61:1140–1153. doi: 10.3758/bf03207619. [DOI] [PubMed] [Google Scholar]
- Taylor JR, Heindel W. Electrophysiological evidence for a fundamental role of perceptual features in concept representation: N400 priming by shared color and shape information. Journal of Vision. 2004;4(8):516a. [Google Scholar]
- Taylor JR. Doctoral Dissertation. 2005. On The Perceptual Basis of Semantic Memory: Representation, process and attentional control revealed by behavior and event-related brain potentials. [Google Scholar]
- van Dam WO, Rueschemeyer SA, Lindemann O, Bekkering H. Context effects in embodied lexical-semantic processing. Frontiers in Psychology. 2010;1:150. doi: 10.3389/fpsyg.2010.00150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warrington EK, McCarthy RA. Categories of knowledge. Further fractionations and an attempted integration. Brain. 1987;110(Pt 5):1273–1296. doi: 10.1093/brain/110.5.1273. [DOI] [PubMed] [Google Scholar]
- Yee E, Huffstetler, Thompson-Schill SL. Function follows form: Activation of shape and function features during object identification. Journal of Experimental Psychology: General. 2011;140(3):348–363. doi: 10.1037/a0022840. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.