

Abstract
The gastrointestinal tract is responsible for a multitude of digestive and immune functions which depend upon the balanced interaction of the intestinal microbiota, diet, gut barrier function, and mucosal immune response. Disruptions in one or more of these factors can lead to intestinal disorders or enteropathies which are characterized by intestinal inflammation, increased gut permeability, and reduced capacity to absorb nutrients. Enteropathy is frequently associated with human immunodeficiency virus (HIV) infection, inflammatory bowel disease, autoimmune enteropathy, radiation enteritis, and irritable bowel syndrome (IBS), where pathologic changes in the intestinal tract lead to abdominal discomfort, bloating, abnormal bowel function (e.g., diarrhea, urgency, constipation and malabsorption). Unfortunately, effective therapies for the management of enteropathy and restoring intestinal health are still not available. An accumulating body of preclinical studies has demonstrated that oral administration of plasma- or serum-derived protein concentrates containing high levels of immunoglobulins can improve weight, normalize gut barrier function, and reduce the severity of enteropathy in animal models. Recent studies in humans, using serum-derived bovine immunoglobulin/protein isolate, demonstrate that such protein preparations are safe and improve symptoms, nutritional status, and various biomarkers associated with enteropathy. Benefits have been shown in patients with HIV infection or diarrhea-predominant IBS. This review summarizes preclinical and clinical studies with plasma/serum protein concentrates and describes the effects on host nutrition, intestinal function, and markers of intestinal inflammation. It supports the concept that immunoglobulin-containing protein preparations may offer a new strategy for restoring functional homeostasis in the intestinal tract of patients with enteropathy.
Keywords: Immunoglobulins, Plasma proteins, Inflammation, Gut barrier, Diarrhea, Malabsorption, Treatment, Nutrition
Core tip: This review article summarizes previous preclinical and clinical studies with serum- or plasma-derived protein preparations with an emphasis on potential benefits for intestinal health and recovery from intestinal disorders. Specifically, how serum-derived bovine immunoglobulin/protein preparations may be useful in restoring intestinal homeostasis (e.g., gut barrier function, immune regulation) following episodes of enteropathy associated with various human disease conditions, such as human immunodeficiency virus infection, inflammatory bowel disease, or irritable bowel syndrome.
INTRODUCTION
The intestinal epithelium is contiguous with the external environment and adaptively organized to both amplify surface area for nutrient absorption and provide a barrier against harmful microorganisms and toxins[1]. The lumen of the gastrointestinal (GI) tract is occupied by a complex assortment of microbial species, the gut microbiota, which changes in response to diet, age, disease, and medical or pharmaceutical intervention[2,3]. This complex array of bacteria and other microbes exist in symbiosis with the intestinal mucosa and play an important role in nutrient absorption, immune regulation, and gut barrier function. At the same time, a variety of immune and physiological adaptations exist within the GI tract to maintain constant vigilance against potentially harmful pathogens and luminal antigens, while preventing the development of uncontrolled inflammation[4].
A variety of factors, both host-related and environmental, can disrupt intestinal homeostasis and lead to development of intestinal disorders or enteropathies. Such enteropathies are characterized by inflammation in the epithelium and lamina propria of the intestine and occur in association with a variety of human conditions or disease states, including: gluten-sensitivity[5], protein-losing enteropathy[6], environmental enteropathy[7,8], radiation-induced enteropathy[9], drug-associated enteropathy[10], irritable bowel syndrome (IBS)[11,12], inflammatory bowel disease (IBD)[13], and human immunodeficiency virus (HIV) infection[14]. In the small intestine, pathologic mucosal changes include blunting of villi and deepening of the crypts, both associated with inflammation that reduces absorptive capacity and tends to increase gut permeability. In the colon, inflammatory changes may be associated with overt epithelial damage or loss. Clinical signs associated with inflammatory mucosal changes may include abdominal pain or discomfort, nausea, bloating, and abnormal bowel function (e.g., urgency, diarrhea, constipation). The pathophysiologic mechanisms leading to enteropathy are not well understood but may involve the effects of exposure to luminal antigens, toxins, or alterations in intestinal microbiota, as well as host diet, genetics, or dysregulated immune responses.
Certain genetic mutations may also predispose to conditions that are associated with enteropathy. For example, more than 200 polymorphisms have been linked to Crohn’s disease. These polymorphisms may predict disease manifestations (stenosis, fistulization, or inflammation), location, and need for surgery[15]. The mutations fall broadly into categories reflecting toll-like receptor-mediated bacterial recognition, autophagy, organic cation transport, lymphocyte differentiation, and barrier function[16,17]. There is some evidence that genetic polymorphisms also play a role in ulcerative colitis[18,19] and IBS[20]. Metabolomic studies have found that malabsorption leads to depleted levels of alanine, glutamine, glutamic acid, isoleucine, leucine, valine, choline and select dietary organic acids, including formate, lactate and succinate, in patients with ulcerative colitis[19,21]. Interestingly, altered expression of genes involved in the production of metabolites from tryptophan through the kynurenine pathway has also been associated with IBS. Clarke et al[22] found significantly higher kynurenine: tryptophan ratios in IBS subjects compared to controls, and acute depletion of tryptophan has been associated with higher levels of abdominal pain and lower levels of serotonin production in IBS[23,24]. Altered tryptophan metabolism through the kynurenine pathway has also been implicated in the events leading to intestinal inflammation in HIV-infected individuals[25] and in IBD[26]. In summary, a variety of enteropathies occur with various human health conditions which are governed by a harmful and continuing cycle of gut barrier dysfunction, immune activation, altered gut microbiota, and impaired nutrient absorption[1,13](Figure 1).
Figure 1.
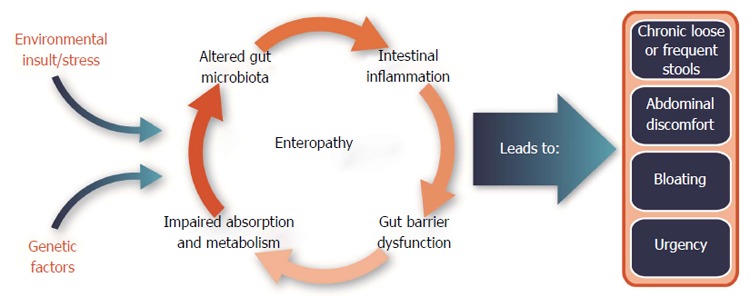
Factors involved in the pathogenesis of enteropathy associated with certain human disease states or conditions (e.g., diarrhea-predominant irritable bowel syndrome or human immunodeficiency virus infections).
Unfortunately, available therapies are often directed at symptoms and not causative factors and include such things as dietary modifications or restrictions, steroids, broad spectrum antibiotics, antidiarrheals, or supportive IV fluids. Due to a broad range of potential causes, multidimensional approaches may be needed, including nutritional interventions alongside current drug treatments to manage these complex disorders. It is well established that plasma-derived protein concentrates (PPC) from bovine, porcine and other sources, when added to the diets of several species of animals, leads to improvements in appetite, weight gain, intestinal growth, and gut barrier function in a number of intestinal disorders[27-30]. Serum-derived bovine immunoglobulin/protein isolate (SBI), specially-formulated to increase IgG and other proteins, has been extensively studied in animal models and recently has been found to be safe and effective in the management of the enteropathy associated with diarrhea-predominant IBS (IBS-D) and HIV infection[31,32]. The purpose of this review is to summarize the scientific evidence supporting the benefits of orally-administered, immunoglobulin-containing protein preparations for host nutrition and protection of gut barrier integrity, particularly as it relates to conditions associated with enteropathy. Studies on the impact of these protein preparations [PPC, bovine serum concentrate (BSC), serum-derived bovine immunoglobulin (SBI)] on host nutrition, gut barrier function, tight junctions, and immune regulation will be summarized.
COMPOSITION OF BOVINE PLASMA- AND SERUM-DERIVED PROTEIN ISOLATES
Plasma-derived protein concentrates are commonly used in animal husbandry to promote growth and modulate intestinal inflammation in immunocompromised young animals[27-30]. Agricultural PPC products are prepared from blood obtained from abattoirs using hygienic collection and processing procedures to ensure high quality plasma products. Preparation first involves the addition of an anticoagulant with subsequent centrifugation to separate the cellular fraction. The plasma is then concentrated by filtration, using inverse osmotic membranes or ultrafiltration and then spray-dried to create a plasma protein powder. During spray drying, plasma proteins are exposed to high temperatures for a very short period of time to avoid denaturation of proteins and to preserve their biological activity[33,34]. In contrast, SBI is produced through a series of shifts in pH and specific salt additions to chilled, edible grade plasma [United States Department of Agriculture (USDA) approved] to reduce the albumin and fibrinogen content and increase the concentration of immunoglobulins and other proteins[35].
Plasma protein concentrates used for animals typically contain over 80% protein on a weight basis, with over 15% of the protein consisting of immunoglobulins (Ig), mainly IgG. In contrast, SBI preparations are specially formulated to increase protein content and reduce levels of albumin and fibrinogen, which results in proportionally higher levels of immunoglobulins. EnteraGam™ is a specially formulated commercial SBI preparation, manufactured according to FDA good manufacturing practice guidelines and intended for human use as a prescription medical food product. It contains about 92% protein (> 50% IgG) with high levels of essential amino acids, including lysine, threonine, tryptophan, and leucine, and is indicated for the clinical dietary management of enteropathy for patients under physician supervision[36]. Arginine and glutamic acid are also found in relatively high levels compared to common dietary sources of protein and the total caloric content of EnteraGam™ is 372 Kcal per 100 g. Although the exact mechanism of how plasma proteins, and SBI in particular, work in patients with chronic loose and frequent loose stools is unknown, findings from both preclinical and clinical studies demonstrate that plasma proteins or SBI has nutritive benefits, binds bacterial endotoxins, supports immune homeostasis, preserves gut barrier function, and promotes a stable microbiota (Figure 2).
Figure 2.
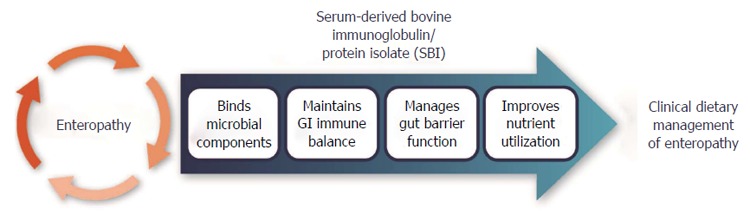
Proposed mode of action for serum-derived bovine immunoglobulin protein isolates to aid management of enteropathy.
NUTRITIONAL BENEFIT
While the purpose of this review is not to provide an extensive summary of all studies conducted on this topic, several representative studies are summarized in Table 1 and the reader is referred to review articles on this topic[27,30]. Torrallardona reviewed the results from 75 trials in 43 publications, to evaluate the feeding and nutritive benefits of PPC from a variety of sources for weaning piglets[30]. Most studies evaluating PPC showed improvements in caloric intake, growth and metabolism, as well as utilization of feed nutrients. Replacement of several high quality protein sources (e.g., meat extracts, soy, pea, potato, skimmed milk, whey, and fishmeal) with PPC at comparable levels led to improved weight gain and feed intake in piglets. Jiang et al[37] evaluated growth performance in piglets after pair-feeding a diet containing soy protein or PPC for 24 d. Protein intake was similar among groups while the rate of weight gain and protein conversion efficiency was significantly higher in the PPC group, especially during the early weaning period. Pigs fed PPC also had improved body weight and absolute mass of protein with no difference in fat mass, suggesting a higher efficiency of dietary protein utilization for lean tissue growth. Feeding PPC reduced the circulating concentrations of urea, arginine, citrulline and ornithine, suggesting a reduction in the catabolism of amino acids to urea and increased availability of dietary amino acids for lean tissue mass. In addition, there were also significant increases in bone mineral content and bone mineral density in the PPC-fed compared to the soy protein-fed group.
Table 1.
Weight gain and growth following dietary supplementation with plasma protein concentrates
| Animal Model (age) | Impact of dietary supplementation with SBI | Ref. |
| Piglets: 14-21 d | Superior growth and feed intakes during the first week in 4 of 5 experiments | Pierce et al[28] |
| Growth performance improved by the IgG-rich fraction | ||
| Piglets: Varying age groups | Consistent improvement in growth, feed intake and sometimes feed conversion; similar results with spray dried plasma from porcine, bovine, and mixed origin | Torrallardona et al[30] |
| Piglets: Weaned at 14 d | Significantly increased mean daily body weight gains and food conversion efficiencies; no difference in protein intake | Jiang et al[37] |
| Significantly greater lean body mass and total carcass mass (P < 0.05) | ||
| Significantly lower circulating urea concentrations (P < 0.05), indicating greater retention of nitrogen and reduced amino acid catabolism | ||
| Piglets: Weaned at 21 d, infected with ETEC K88 | Increased average daily weight gain and food intake | Bosi et al[44] |
| Protected against E. coli-induced inflammation |
SBI: Serum-derived immunoglobulin/protein isolates; E. coli: Escherichia coli; ETEC K88: Enterotoxigenic E. coli, K88 strain.
Pierce et al[28] conducted several experiments to evaluate the growth and feed intake of weaned piglets fed porcine PPC, bovine PPC, or different molecular weight fractions of PPC. Collectively, the results demonstrated that both porcine and bovine PPC enhanced growth rate and feed intake of weaned piglets, while the IgG fraction of porcine or bovine plasma appeared to stimulate growth performance that was comparable to intact PPC and superior to the albumin or low MW fractions of PPC. These data suggests that a distinct nutritional role may exist for the IgG-rich fraction of PPC to support growth performance.
SAFETY AND DIGESTIBILITY
Plasma-derived protein concentrates (e.g., PPC, SBI) are composed of > 50% IgG and other proteins and peptides that reflect the composition of plasma and are similar to other serum proteins present in colostrum and milk. Such products typically do not contain milk ingredients such as lactose, casein, or whey, so adverse reaction rates would be expected to be minimal. However, patients who have an allergy to beef should not take SBI or PPC products. The rigorous process used to prepare commercial forms of SBI meets strict industry standards to ensure that finished products do not become contaminated with infectious agents, including the bovine spongiform encephalitis (BSE) agent. In addition, SBI has been self-affirmed as Generally Recognized as Safe (GRAS) with no safety-related questions by the US Food and Drug Administration (FDA) for doses up to 50 g/d. SBI has not yet been tested in pregnant or nursing mothers or immunocompromised individuals, so use in such patients should be at the discretion of the patients’ physician.
The safety of SBI has been evaluated in both pediatric and adult subjects. Tolerance and digestibility of SBI was evaluated in 12 healthy adult volunteers by Hanning et al[38]. Volunteers were administered 10 g of SBI orally and blood samples were obtained at various time points, which showed elevated levels of plasma total amino acids and leucine at 1-2 h following SBI administration. Bovine IgG was not detected in serum samples from study subjects, suggesting that bovine IgG remains in the intestinal tract and does not pass the luminal barrier into the blood stream. Subjects then consumed 5 g of SBI daily for 2 wk and completed daily diaries for general health and adverse events (AEs). No serious AEs were reported by test subjects. The following AEs were reported: increased urination (3); stomach cramps (3); fatigue (2); headache (2); sore throat, softened stools, nausea, constipation, and irritability (1 each). Bovine IgG was detected by enzyme-linked immunosorbent assay (ELISA) in stool samples from test subjects on day 14 but not at baseline (day 0), suggesting survival of some IgG following GI transit, which is similar to previous reports[39,40].
A standard diet with graded amounts of PPC about 0.9 to 2.5 mg/kg BW/d was also fed to infants 9 to 25 mo of age at entry (n = 10) recovering from severe protein-energy malnutrition to evaluate acceptability, safety, and digestibility[41]. Study diets were well accepted by study subjects with no evidence of intolerance and no AEs were reported. In another study, malnourished infants (age 6-7 mo of age at entry; n = 107) fed a diet containing PPC (about 3.5 g/d) for up to 8 months showed no side effects or adverse impact on growth or morbidity rates when compared to infants fed supplemented with whey protein concentrate[42]. Studies in HIV+ patients (n = 8)[31], a longer term open-label exposure in HIV+ patients (n = 35) (data on file), and subjects with IBS-D (n = 66)[32] also showed only minor or non-medication related adverse events, as well as no clinically relevant changes in blood chemistries or hepatic or renal markers in any studies. Collectively, the results from available clinical studies suggest that SBI is safe and well-tolerated when consumed up to 8 mo in doses ranging from 0.18 to 10 g per day in infants, children and adults.
In order for PPC supplementation to provide benefits to dysfunctional intestinal mucosa, the immunoglobulin and other active protein components must resist digestion and remain active in the lumen of the intestine. Morel et al[39] used radial immunodiffusion to evaluate survival of IgG at various points along the intestine in weaned piglets fed PPC. They found 50% undigested IgG located in the proximal small intestine, 17% in mid-small intestine and 10% in the distal small intestine, but none in the cecum and colon. Rodriguez et al[40] found IgG survival through the intestinal tract at 8% and 5%, in adult dogs and cats fed PPC or purified IgG, respectively, which suggests partial resistance to digestion. The authors found that the immunoglobulin fraction present in the feces of these animals was the Fab fraction.
IMPACT ON GUT BARRIER AND INTESTINAL RECOVERY
The ability of PPC and SBI to modulate intestinal barrier function, permeability, and malabsorption has been evaluated in a number of preclinical and clinical studies.
Preclinical studies
Studies on the effects of bovine immunoglobulin isolates (PPC or SBI) on inflammation in the GI tract have primarily come from preclinical models in which animals were challenged by infection or exposure to bacterial toxins (Table 2). In one study of piglets infected with rotavirus, PPC was effective at reducing diarrhea, improving intestinal recovery and maintaining growth[43]. Infected soy-fed pigs had significantly greater diarrhea scores (P < 0.001) from day 1 to 7 post-infection, while diarrhea scores of infected pigs fed PPC ranked the same as scores from uninfected controls. Administration of PPC was not able to attenuate the reductions in intestinal villus height and the villus height/crypt depth ratio caused by rotavirus infection. Nevertheless, oral feeding of PPC maintained greater intestinal mucosa protein and estimated total lactase activity than infected, soy protein-fed piglets. In a second study, weaned pigs were challenged with enterotoxigenic Escherichia coli K88 (ETEC K88), used as a model of in vivo pig IBD, to investigate whether PPC could improve growth, immune defense and reduce intestinal inflammation[44]. Compared to a diet based on fish protein, ETEC K88 infected pigs fed PPC showed higher calorie intake and daily weight gain, less intestinal mucosal damage and inflammatory cell infiltration, and reduced expression of pro-inflammatory cytokines.
Table 2.
Effects of plasma-derived protein concentrates on intestinal function in animal models
| Species | Model/indication | Impact of dietary supplementation with SBI | Ref. |
| Pig | Postweaning | Reduced colonic paracellular permeability | Peace et al[46] |
| Reduced ileal permeability | |||
| Fewer lamina propria cells in ileum and colon | |||
| Reduced transepithelial electrical resistance in the colon - improved tight junction | |||
| Significantly improved fecal scores | |||
| Rotavirus infection | Significantly reduced clinical signs of diarrhea | Corl et al[43] | |
| Significantly greater intestinal mucosal protein and lactase activity | |||
| Infection by ETEC K88 | Decreased inflammatory cell infiltration and mucosal damage | Bosi et al[44] | |
| Increased crypt depth, reduced intestinal expression of proinflammatory TNF-α and IL-8 | |||
| Rat | Exposure to SEB | Improved ion transport function, as measured by reductions in the potential difference across the jejunum and Na-K-ATPase activity | Pérez-Bosque et al[47] |
| Improved mucosal permeability (dextran flux and HRP paracellular flux) |
SBI: Serum-derived immunoglobulin/protein isolates; ETEC K88: Enterotoxigenic Escherichia coli, K88 strain; TNF-α: Tumor necrosis factor alpha; IL-8: Interleukin 8; SEB: Staphylococcus aureus enterotoxin B; Na-K-ATPase: Sodium-potassium adenosine triphosphatase; HRP: Horse radish peroxidase.
In a third study of infectious enteritis - Cryptosporidium parvum infection in neonatal calves, a disease which produces moderate intestinal inflammation, watery diarrhea, and increased intestinal permeability - Hunt et al[45] showed that the daily addition of a bovine serum product (compared with a soy protein control) reduced diarrheal volume, oocyte shedding, and intestinal permeability, while facilitating villus re-growth and increasing mucosal surface area. Lactase activity was significantly improved in response to bovine serum concentrate.
Other data in preclinical models have specifically evaluated tight junction protein expression in response to early weaning and toxin challenge. Peace et al[46] evaluated the effects of PPC in piglets undergoing early weaning, a condition known to induce impairment in intestinal epithelial barrier function. Piglets were fed a control diet containing PPC for 7 or 14 d to evaluate impact on ileal and colonic barrier function. Co-administration of PPC with radiolabeled nutrients reduced paracellular permeability as indicated by significant reductions in colonic 14C-inulin permeability on day 7 post-weaning and reduced ileal 3H-mannitol and 14C-inulin permeability on day 14. Protein plasma concentrate also reduced the predominantly lymphocytic cellular infiltration in the lamina propria in both ileum and colon, concomitantly reducing levels of pro-inflammatory cytokines in colon (see below). As shown by immunofluorescence staining, claudin-1, a tight junction protein was more highly expressed and localized to tight junctions in animals fed PPC.
The protective effects of spray-dried porcine PPC in a rat model of intestinal inflammation were also evaluated[47]. Weaned rats were fed a diet with or without PPC for 14 d then exposed to intraperitoneal challenge with Staphylococcus aureus enterotoxin B (SEB) known to disturb barrier function and ion transport. Addition of PPC to diets significantly ameliorated SEB-induced increases in intestinal permeability as measured by dextran flux (P < 0.05) and horseradish peroxidase (HRP) paracellular flux (P < 0.05) across the intestinal epithelium. Plasma protein concentrate was also shown to increase β-catenin expression, part of the adherens complex positioned adjacent to the tight junction. These data suggest that PPC beneficially promoted endogenous repair of the tight junctions, modulated inflammation, reduced permeability, and improved diarrhea in pigs challenged with enterotoxin B.
Collectively, the results of these experimental studies suggest that dietary plasma protein preparations strengthen intestinal barrier function and prevent alterations in intestinal epithelium during inflammation. Two reviews have been published on the effects of PPC and the proteins in SBI on intestinal barrier function in animal models of human disease[48,49].
Clinical studies
Two clinical trials evaluated the efficacy of dietary SBI for improving intestinal absorption, GI symptom scores, and quality of life measures in patients with HIV-associated enteropathy or IBS-D (Table 3). An open-label study was conducted by Asmuth et al[31] to evaluate the impact of oral SBI on GI symptoms and systemic markers of immune activation in patients with a diagnosis of HIV-associated enteropathy. To qualify, patients with enteropathy were given an extensive evaluation to exclude other GI disease. Eight patients were enrolled in the study and received 5 g of SBI/d for 8 wk followed by a 4 wk washout period. Administration of SBI led to consistent improvement in GI symptoms associated with HIV enteropathy. After 8 wk of SBI administration, bowel movements per day decreased from 5.8 to 2.0 (P = 0.008) and stool consistency scores (1-formed to 6-watery) improved from 5.3 to 3.0 (P = 0.013). A GI symptom questionnaire showed a marked decrease in score from 6 to 0.5 (P =0.008). After a 4 wk washout period, 5 patients continued on SBI to week 48 maintaining similar bowel movements and stool consistency and GI symptom questionnaire responses. An additional open-label, in-market analysis of 31 patients taking various nutritional formulas which contained 2.5 to 5.0 g SBI showed improved management of loose stools, thus providing further evidence for the management of HIV-associated enteropathy (data on file).
Table 3.
Human studies with serum-derived immunoglobulin/protein isolates to evaluate intestinal benefits and quality of life
| Species | Model/indication | Impact of dietary supplementation with SBI | Ref. |
| Human n = 8, HIV positive adults | HIV-associated enteropathy | Significant reduction in mean bowel movements/day and improvement in stool consistency scores after 8 wk (P = 0.008) | Asmuth et al[31] |
| Significant reduction in GI questionnaire scores from 17 at baseline to 8.0 at 8 wk (P =0.008) | |||
| No change in gut permeability (disaccharide absorption); increase in D-xylose absorption in 7/8 subjects | |||
| Maintained stool frequency and consistency for an additional 9 mo (n = 5) | |||
| Human n = 66 adults | IBS-D | 10 g/d showed significant decrease in # symptom days with abdominal pain, flatulence, bloating, loose stools, urgency or any symptom over 6 wk (P < 0.05) | Wilson et al[32] |
| 5 g/d showed significant improvements in loose stools, hard stools, flatulence and incomplete evacuation (P < 0.05) | |||
| Human n = 10 infants or children (9-25 mo) | Malnutrition | Significant reductions in fecal wet and dry weights, and lower fecal fat and energy losses compared with the control diet (P < 0.05) in relation to the amount of SBI in the diet during three randomly ordered 7-d periods | Lembcke et al[41] |
| Human n = 259 infants (6-7 mo) | Malnutrition | Trends toward weight gain and upper arm circumference (a measure of lean body mass) increases were found in the SBI + micronutrient group vs SBI alone | Bégin et al[42] |
HIV: Human immunodeficiency virus; IBS-D: Irritable bowel syndrome, diarrhea predominant.
A randomized, double-blind, placebo-controlled study was conducted in individuals with IBS-D to investigate the efficacy of SBI on decreasing gastrointestinal symptom scores and improving the quality of life[32]. Study subjects (n = 66) with a diagnosis of IBS-D for at least 6 mo prior to enrollment met the Rome II diagnostic criteria for IBS. Test groups received placebo (10g /d soy protein isolate), SBI 5 g/d + 5.0 g/d placebo or SBI 10 g/d for 6 wk and completed an IBS-36 questionnaire at baseline (day 0) and at the end of the study (week 6). The daily symptom diary assessed the presence and severity of the following symptoms: nausea, abdominal pain, flatulence, bloating, hard stools, loose stools, urgency, straining, incomplete evacuation and mucus. Forty-five subjects completed the study per protocol and were included in the analysis: 10 g/d SBI (n = 15), 5 g/d SBI (n = 15), and placebo group (n = 13). Results showed that subjects receiving 10 g/d of SBI experienced significant within-group reductions in the number of days with abdominal pain (P < 0.01), loose stools (P < 0.01), bloating (P < 0.05), flatulence (P < 0.01), urgency (P < 0.05) and any symptom (P < 0.01) at EOT vs baseline (Table 3). Subjects receiving 5 g/d of SBI (n = 15) reported statistically significant within-group reductions in days with flatulence (P < 0.035), incomplete evacuation (P < 0.05), and ‘‘any symptom’’ (P < 0.01). No significant within group improvements were seen in the placebo group. There were no significant changes in quality of life (QoL) scores or in hematology or clinical chemistry values among the therapy groups.
Studies have also been performed in infants and children recovering from malnutrition. A standard diet with graded amounts of SBI was also administered to infants or children 9 to 25 mo of age at entry (n = 10) recovering from severe protein-energy malnutrition to evaluate acceptability, safety, and digestibility during three randomly ordered 7-d dietary periods[41]. Replacing 50% of the protein in the standard diet with SBI led to significant reductions in fecal wet and dry weights, and lower fecal fat and energy losses, suggesting greater absorption of fat and energy compared with the control diet (P < 0.05) (Table 3). Investigators suggested that SBI enhanced intestinal recovery from severe malnutrition.
Another randomized, controlled, community-based intervention study evaluated the effects of SBI and/or multiple micronutrients on children’s growth, morbidity, and micronutrient status[42]. A total of 259 children who were initially 6 to 7 mo of age received 1 of 4 maize-based dietary products daily for 8 mo with or without protein supplementation. Groups studied: SBI, whey protein concentrate (WPC, control group), SBI plus multiple micronutrients, or WPC plus multiple micronutrients. Two hundred and 25 (225; 86%) children completed ≥ 60 d of observation, 184 (71%) completed ≥ 180 d of observation, and 132 (51%) distributed among the 4 treatment groups finished the full 8 mo of observation. There were no significant differences in growth or morbidity by treatment group for those children who completed 8 mo of observation. Although not statistically significant, there were trends toward weight gain and mid arm circumference (a measure of lean body mass) increases in the SBI+ micronutrient group suggesting better utilization of these nutritional substances (Table 3).
EFFECTS ON GUT MICROBIOTA
Changes in gut microbiota has been identified as one potential factor in causing inflammation that leads to alterations in gut barrier function with associated increases in mucosal permeability. An increase in firmicutes over bacteroidetes bacteria has been reported in IBS patients[50-52]. Another study in IBS-D patients found an increase in bacteroides and clostridia with an associated reduction in bifidobacteria[53]. Pediatric IBS-D patients were reported to show significant differences compared to healthy controls having statistically greater numbers of gammaproteobacteria[54]. There is also a well-recognized dysbiosis that occurs in IBD, although the colonic bacterial imbalances are less well-characterized for ulcerative colitis compared to Crohn’s disease[55-58]. Recently it was also reported that the microbiota in both ulcerative colitis and Crohn’s disease was relatively unaltered, but metabolism by bacteria in the microbiota was significantly changed[59]. There were notable shifts in fecal metabolome showing reduced carbohydrate processing and alterations in various amino acid biosynthesis pathways. This alteration in gut microbiota may contribute to increased tight junction permeability with associated decreases in barrier function, and changes in bacterial metabolic products and host nutrient malabsorption. Diet may play a role in the causality and/or in the progression of both IBS and IBD[60,61]. Therefore, it is reasonable to assume that diet may play a role in restoring a natural balance to the gut microbiota and metabolome.
In the HIV-associated enteropathy population treated with SBI, the firmicutes and bacteroidales were the dominant phyla in all 8 patients[62]. When SBI was administered to these patients, proinflammatory gammaproteobacteria decreased from 0.70% to 0.12%. Clostridium (genus) decreased from 6.5% to 3.4% in the stool and correlated with duodenal CD3+/CD4+ density (r = -0.63; P < 0.01). Ruminococcus and the bacteroidetes/firmicutes ratio, which increased in 6/8 SBI-treated subjects in the study, have been shown to contribute to better calorie utilization from the diet[63,64]. Changes in gut microbiota in the study also correlated with local lymphocyte populations that increased significantly with short-term SBI administration over 8 wk. These results suggest that some component in the formulation may be normalizing gut bacteria, perhaps the IgG fraction. Work is underway to further characterize these interactions.
EFFECTS ON INTESTINAL INFLAMMATION
The release of inflammatory mediators, such as reactive oxygen species, prostaglandins, leukotrienes, and cytokines, from mucosal leukocytes is associated with the altered barrier function and increased permeability caused by intestinal inflammation. Cytokines such as interleukin (IL)-1β and tumor necrosis factor (TNF)-α play an important governing role in such inflammatory responses, while other cytokines such as IFN-γ, IL-12 and IL-18 affect the production and cellular response to IL-1β and TNF-α[65]. In models of inflammation where several cytokines are produced, specific blockade of IL-1β and/or TNF-α results in a reduction in the severity of the inflammation[65]. Dietary PPC has been shown to reduce the expression of proinflammatory cytokines and alter the lymphocyte response during immune activation in weaned piglets[46] as well as experimental models of intestinal inflammation in mice[66,67], rats[68-70] and pigs[44] (Table 4). For example, a study by Pérez-Bosque et al[71] investigated the effects of dietary SBI on immune responses of mucosal-associated lymphoid tissue in mice with a genetic predisposition to IBD. Wild type (WT) mice and mice lacking the mdr1a gene (KO) were fed diets supplemented with either SBI (2% w/w) or milk proteins (control diet) starting on day 19 (weaning). At day 56, SBI reduced the production of the proinflammatory cytokines and chemokines IL-17, IL-6 and CCL4 (P < 0.05), prevented the expression of IFN-γ (P < 0.05), and blocked the increase in colon crypt permeability that was found in the mdr1a KO model. SBI treatment produced increases in mucosal concentrations of anti-inflammatory TGF-β and in the percentage of regulatory T lymphocytes (both P < 0.05), thus reducing the activated Th1 to regulatory Treg lymphocyte ratio. PPC and immunoglobulin-enriched protein isolates have also been demonstrated to affect Peyer’s patch lymphocyte populations in weaned rats challenged with S. aureus superantigen B (SEB)[70]. In this study, it was shown that the mild intestinal inflammation associated with the SEB model was reduced by dietary PPC as measured by decreased diarrhea. Furthermore, the administration of PPC significantly increased the number of T-helper cells, while reducing the number of activated T-helper cells as compared with animals not fed PPC or immunoglobulin-enriched protein isolates[70]. This same trend was observed for changes in the population of γδ-T cells and natural killer (NK) T cells in the Peyer’s patches of rats fed diets containing PPC or immunoglobulin isolates.
Table 4.
Effects of serum-derived immunoglobulin/protein isolates administration on immune and inflammatory markers
| Species | Model/indication | Impact of dietary supplementation with SBI | Ref. |
| Pig | ETEC K88 | Reduced expression of TNF-α and IL-8 in the gut | Bosi et al[44] |
| Postweaning | Reduced TNF-α in the colon | Peace et al[46] | |
| Reduced IFNγ levels in the ileum and colon day 7, but not day 14 post weaning | |||
| Rat | SEB | Prevented the SEB-induced increase in IFN-γ, IL-6, and LTB4 in Peyer's patches and in the mucosa | Pérez-Bosque et al[69] |
| Increased anti-inflammatory cytokines (IL-10 and mature TGF-β) in intestinal mucosa | Pérez-Bosque et al[68] | ||
| Reduced SEB-induced increase in cytotoxic lymphocyte populations of γδ-T cells, natural killer cells, and the number of activated T lymphocytes in lamina propria. | |||
| Mouse | Mdr1-/- knockout mouse model of spontaneous colitis | Reduced the percentage of activated Th lymphocytes | Moretó et al[48] |
| Reduced INF-γ and TNF-α expression in the colon | |||
| Significantly reduced the expression of cytokines IL-2 and IL-17, chemokines MCP-1 and MIP-1b, and iNOS in the mucosa | |||
| Mouse | 2% DSS-induced IBD model | Reduced elevation of IL-1α, IL-4, IL-6, IL-10, MCP-1, and KC | Jiang et al[67] |
| Human (HIV+ adults) | HIV enteropathy | I-FABP fell below baseline in 4/5 patients who continued receiving SBI (P < 0.12) out to 48 wk | Asmuth et al[31] |
| MMP-9/TIMP-1 ratios in subjects were significantly lower than controls at baseline (P < 0.007) | |||
| MCP-1 levels decreased in 5/5 patients who continued receiving SBI (P < 0.06) out to 48 wk |
ETEC K88: Enterotoxigenic Escherichia coli, K88 strain; TNF-α: Tumor necrosis factor α; IL: Interleukin; SEB: Staphylococcus aureus enterotoxin B; IFNγ: Interferon-γ; LTB4: Leukotriene B4; TGF-β: Transforming growth factor β; MCP 1: Monocyte chemotactic protein 1; MIP-1b: Macrophage inflammatory protein; iNOS: Inducible nitric oxide synthase; KC: Keratinocyte-derived cytokine; HIV: Human immunodeficiency virus; I-FABP: Intestinal-fatty acid binding protein; MMP-9: Matrix metalloproteinas-9; TIMP: Tissue inhibitor of metalloproteinase.
In an initial clinical trial in HIV patients with decimated lamina propria CD4+ counts, SBI ingestion significantly increased jejunal CD4+ lymphocyte densities over 8 wk, but had no effect on circulating CD4+ counts[31]. In addition, levels of I-FABP, a marker for enterocyte damage, initially rose in 7/8 subjects after 8 wk, but then fell below baseline in 4/5 who continued taking SBI after 48 additional weeks on product, suggesting that damage to enterocytes caused by inflammation had ceased (Table 4). Collectively, data from these preclinical and clinical studies support the hypothesis that the distinctive protein composition of SBI can play a role in the modulation of the immune response in the intestine.
MECHANISM OF ACTION
SBI contains immunoglobulins, particularly IgG, that are directed against a wide array of pathogens and foreign antigens due to the fact that SBI is prepared from plasma obtained from thousands of animal donors. The Fab regions of IgG recognize antigenic targets and provide diversity to antibodies, while the Fc region interacts with Fc gamma receptors on certain immune cells to enhance phagocytic activity by macrophages, monocytes, and polymorphonuclear neutrophils (PMNs). Several mechanisms may explain how oral immunoglobulins modulate intestinal inflammation and support gut barrier function. For example, immunoglobulins in SBI may simply bind directly to specific microbial pathogens or their toxins, thereby interfering with their ability to migrate through the mucus layer and enter or damage epithelial cells.
A more likely explanation of how oral immunoglobulins work to maintain intestinal homeostasis may involve binding to highly-conserved microbial antigens such as bacterial lipopolysaccharide (LPS), also known as microbe-associated molecular patterns (MAMPs), and interfere with signaling pathways that lead to inflammation (Figure 3). Under normal conditions, cells of the innate immune system play a crucial role in maintaining intestinal homeostasis through a highly-regulated process involving the recognition of MAMPs through pattern recognition receptors (PRRs), such as toll-like receptors (TLRs). TLRs are differentially expressed by various cells of the GI tract (e.g., macrophages, dendritic cells, endothelial cells, myofibroblasts)[72,73] and play a key role in signaling the recognition of MAMPs by activating several inflammatory pathways, including the NF-κB pathway which is a key regulator of proinflammatory TNF-α, IL-1β, IL-6 and IL-8 cytokine production[74]. Prolonged recognition of MAMPs can lead to a persistent state of inflammation associated with numerous chronic inflammatory disorders such as IBS, IBD, and HIV enteropathy. Studies have shown that the IgG, IgA, and IgM contained in SBI bind to bacterial endotoxins and a wide array of other bacterial, viral, and fungal MAMPs[75,76]. Therefore, it is possible that SBI binding of microbiota components results in less binding of MAMPs by macrophages and dendritic cells which may interfere with release of IL-1, IL-6, and TNF-α[77]. Similarly, less presentation of antigens by dendritic cells and macrophages may result in a decrease in activated T cell populations and more regulatory cell phenotypes that produce IL-10 to dampen inflammation[70].
Figure 3.
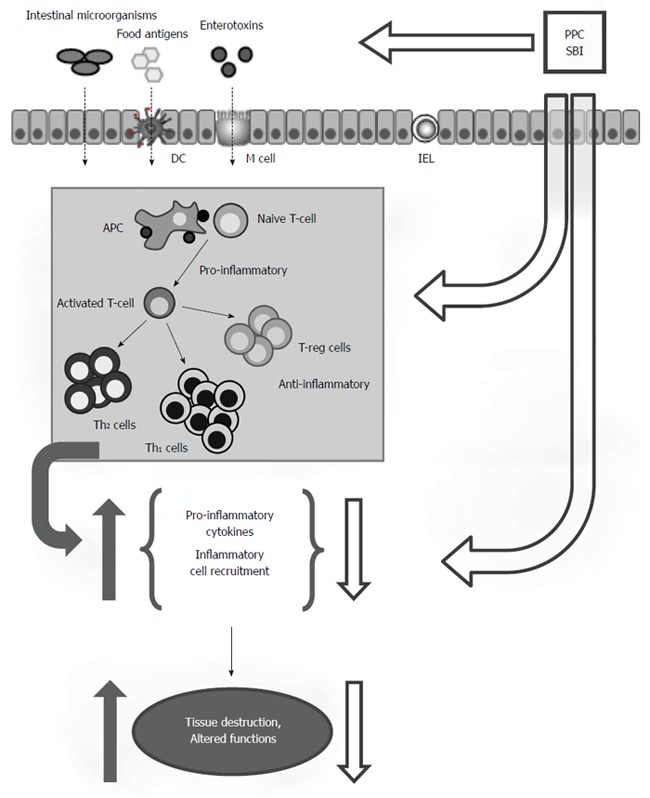
Summary of the postulated mode of action for serum-derived immunoglobulin/protein isolates. Immunoglobulins in SBI support intestinal homeostasis by binding MAMPs (endotoxins, etc.), toxins or other antigens in the lumen of the intestinal tract. Immunoglobulin binding interferes with downstream antigen detection by cell surface receptors on IELs or APCs such as DCs and macrophages that influence T cell activation, cytokine production, and barrier fortification. Additionally, biologically active compounds in immunoglobulin isolates may interact directly with mucosal immune cells in the lamina propria to influence mucosal inflammatory responses and epithelial cells to influence barrier function. SBI: Serum-derived immunoglobulin/protein isolates; MAMP: Microbe-associated molecular patterns; IEL: Intraepithelial lymphocytes; APC: Antigen presenting cells; DC: Dendritic cells; T-reg: Regulator T-lymphocytes; Th1: T helper type 1; Th2: T helper type 2. Reprinted with permission from Moretó et al[48].
Alternatively, SBI may contain a large fraction of natural antibodies that work in other ways to maintain immune homeostasis. For example, studies with intravenous immunoglobulin (IVIG) have shown that binding of the Fc portion of IgG to Fc receptors on target cells may govern some of the anti-inflammatory mechanisms involved with IVIG therapy by up-regulating the expression of inhibitory classes of Fc receptors and down-regulating the activating class of Fc receptors[78]. Autoreactive antibodies in IVIG have also been shown to modulate Th1 and Th2 cytokine production[79], trigger the production of interleukin-1 receptor antagonist, abrogate the capacity of mature dendritic cells to secrete IL-12 upon activation in vitro, and enhance anti-inflammatory IL-10 production[80]. Collectively, such immune modulating effects of SBI might explain previous reports of reduced expression of pro-inflammatory cytokines and altered lymphocyte response to immune activation in weaned piglets[46] and experimental models of intestinal inflammation[66-69].
The ability of SBI to modulate inflammation may also benefit the patient with enteropathy by improving gut barrier function. A developing body of evidence indicates that intermittent or even minor inflammation in the intestinal mucosa can elicit changes in intestinal structure and function, leading to increased mucosal permeability[12,81]. For example, increased production of pro-inflammatory cytokines such as TNF-α, IFN-γ, and various interleukins during certain chronic inflammatory disorders[4,82,83] have been shown to increase paracellular permeability by impacting the expression or degradation of claudin and occludin tight junction proteins[84,85]. Conversely, certain anti-inflammatory cytokines such as IL-10 and TGF-β appear to maintain tight junction barrier and protect against intestinal inflammation[82].
In addition to the IgG content of PPC and SBI, the effect on lean body mass may also be in part due to the amino acid content of the complex protein mixture. Plasma protein concentrate and SBI contain amino acids which have been identified to be important for recovery after intestinal damage from infectious agents[43,44,47]. For example, glutamine serves as a preferential energy source for rapidly proliferating immune cells and enterocytes, is a nontoxic transporter of ammonia, and has been linked with maintenance of gut barrier function and cell differentiation[86]. Amino acids absorbed into the blood from PPC or SBI may also play an anabolic role in the body; for example, tryptophan may support the generation of serotonin or metabolite formation in the kynurenine pathway[22-24].
CONCLUSION
Chronic intestinal disorders or enteropathies occur in a variety of human disease conditions such as IBS, IBD, and HIV infection which are characterized by intestinal inflammation, increased gut permeability, and reduced capacity to absorb nutrients. Most therapies used to treat enteropathy are aimed at managing symptoms or target single pathways. However, a multifaceted approach may be needed to manage enteropathy associated with these complicated disease states, or in some cases the side effects of pharmaceutical treatment protocols.
There is a developing body of evidence indicating that intermittent or even relatively minor inflammation can lead to changes in intestinal structure and barrier function[87]. Translocation of bacterial antigens may result in increased production and secretion of pro-inflammatory cytokines, including TNFα, IFN-γ, and interleukins[4,82], which contribute to processes that degrade structural tight junction proteins (e.g., occludins[84], claudins[85]), and contribute to symptoms associated with enteropathy. Inflammation-driven disruption of barrier function has been shown to negatively influence growth in young animals, and also has a range of health consequences in humans[1]. For example, post-infectious IBS is recognized to have inflammatory involvement which may persist months after the initial resolution of infection[88,89] with associated intestinal histological changes and increased intestinal permeability[12,83,90,91]. HIV-associated enteropathy has long been associated with inflammatory damage, decreased barrier function, increased permeability and malabsorption of nutrients[92-94]. Due to increased permeability, microbial translocation markers in HIV patients have been shown to be significant predictors for disease progression and death[95,96]. Serum-derived bovine immunoglobulin/protein isolate may provide a distinctive protein composition to counter intestinal inflammation and the resulting changes in barrier function as well as tight junction permeability to help maintain proper functioning of the intestine. Enteropathy is also associated with chronic undernutrition[97,98]. Malabsorption of nutrients such as bile acids, polyols, fructose, and lactose has been reported to contribute to increased symptoms in patients with IBS[99-101]. A nutritional deficiency of vitamin B6 may also be correlated with IBS symptoms[102]. Nutritional interventions may be needed alongside current drug treatments to effectively manage these complicated disorders.
Results from numerous research studies consistently demonstrate beneficial physiological effects for IgG-containing PPC and SBI protein preparations[27,28,30,33]. SBI contains distinctive nutritional factors that may impart growth and protective benefits by several different mechanisms including binding endotoxin, supporting intestinal barrier function, fostering the growth and maintenance of the normal microbiota, reducing pro-inflammatory cytokine production, and maintaining epithelial tight junctions. In environmentally stressed or disease states, increased cytokine production can promote an increase in enteric epithelial tight junction permeability with resultant antigenic penetration of the gut barrier. These effects may be ameliorated through PPC and SBI preparations via the reduction of pro-inflammatory cytokine expression, including TNF-α, IFN-γ, IL-1β, IL-6, IL-8, IL-17, thus facilitating restoration of normal GI function and improved nutritional utilization of accompanying SBI proteins[44,66-70,103].
Safety in humans has been confirmed in five clinical studies in which SBI has been administered. Four clinical trials have reported results that are consistent with the hypothesis that SBI may improve intestinal dysfunction. The study of SBI in HIV-enteropathy patients[31] in which D-xylose uptake was increased and in infants with malnutrition[41] where fecal wet/dry weights as well as lower fecal fat and energy losses were prevented suggest that this distinct and specially formulated protein mixture is able to restore intestinal structure and functional damage caused by proteolytic enzymes, lymphocytic cytokines, or chemokine-induced damage. Oral SBI may represent a safe and effective option with multiple modes of action to provide for distinctive nutritional requirements in patients with disease-related enteropathy to increase digestion, absorption, metabolism, and utilization of a variety of macro- and micronutrients and facilitate resolution of their gastrointestinal symptoms associated with enteropathy, compared to other protein sources.
Footnotes
P- Reviewer: Hauer-Jensen M, Howarth GS, Mazzarella G S- Editor: Ding Y L- Editor: A E- Editor: Wang CH
References
- 1.Farhadi A, Banan A, Fields J, Keshavarzian A. Intestinal barrier: an interface between health and disease. J Gastroenterol Hepatol. 2003;18:479–497. doi: 10.1046/j.1440-1746.2003.03032.x. [DOI] [PubMed] [Google Scholar]
- 2.Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, Magris M, Hidalgo G, Baldassano RN, Anokhin AP, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486:222–227. doi: 10.1038/nature11053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sonnenburg JL, Angenent LT, Gordon JI. Getting a grip on things: how do communities of bacterial symbionts become established in our intestine? Nat Immunol. 2004;5:569–573. doi: 10.1038/ni1079. [DOI] [PubMed] [Google Scholar]
- 4.Martin GR, Wallace JL. Gastrointestinal inflammation: a central component of mucosal defense and repair. Exp Biol Med (Maywood) 2006;231:130–137. doi: 10.1177/153537020623100202. [DOI] [PubMed] [Google Scholar]
- 5.Green PH, Cellier C. Celiac disease. N Engl J Med. 2007;357:1731–1743. doi: 10.1056/NEJMra071600. [DOI] [PubMed] [Google Scholar]
- 6.Greenwald DA. Protein losing enteropathy. In: Feldman M, Friedman LS, Brandt LJ, editors. Sleisenger & Fordtran’s Gastrointestinal and Liver Disease. 9th ed. Philadelphia, PA: Saunders Elsevier; 2010. p. 437. [Google Scholar]
- 7.Korpe PS, Petri WA. Environmental enteropathy: critical implications of a poorly understood condition. Trends Mol Med. 2012;18:328–336. doi: 10.1016/j.molmed.2012.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Prendergast A, Kelly P. Enteropathies in the developing world: neglected effects on global health. Am J Trop Med Hyg. 2012;86:756–763. doi: 10.4269/ajtmh.2012.11-0743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hauer-Jensen M, Denham JW, Andreyev HJ. Radiation enteropathy-pathogenesis, treatment and prevention. Nat Rev Gastroenterol Hepatol. 2014;11:470–479. doi: 10.1038/nrgastro.2014.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sostres C, Gargallo CJ, Arroyo MT, Lanas A. Adverse effects of non-steroidal anti-inflammatory drugs (NSAIDs, aspirin and coxibs) on upper gastrointestinal tract. Best Pract Res Clin Gastroenterol. 2010;24:121–132. doi: 10.1016/j.bpg.2009.11.005. [DOI] [PubMed] [Google Scholar]
- 11.Camilleri M, Lasch K, Zhou W. Irritable bowel syndrome: methods, mechanisms, and pathophysiology. The confluence of increased permeability, inflammation, and pain in irritable bowel syndrome. Am J Physiol Gastrointest Liver Physiol. 2012;303:G775–G785. doi: 10.1152/ajpgi.00155.2012. [DOI] [PubMed] [Google Scholar]
- 12.Matricon J, Meleine M, Gelot A, Piche T, Dapoigny M, Muller E, Ardid D. Review article: Associations between immune activation, intestinal permeability and the irritable bowel syndrome. Aliment Pharmacol Ther. 2012;36:1009–1031. doi: 10.1111/apt.12080. [DOI] [PubMed] [Google Scholar]
- 13.Fasano A, Shea-Donohue T. Mechanisms of disease: the role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases. Nat Clin Pract Gastroenterol Hepatol. 2005;2:416–422. doi: 10.1038/ncpgasthep0259. [DOI] [PubMed] [Google Scholar]
- 14.MacArthur RD, DuPont HL. Etiology and pharmacologic management of noninfectious diarrhea in HIV-infected individuals in the highly active antiretroviral therapy era. Clin Infect Dis. 2012;55:860–867. doi: 10.1093/cid/cis544. [DOI] [PubMed] [Google Scholar]
- 15.Tsianos EV, Katsanos KH, Tsianos VE. Role of genetics in the diagnosis and prognosis of Crohn’s disease. World J Gastroenterol. 2011;17:5246–5259. doi: 10.3748/wjg.v17.i48.5246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Barrett M, Chandra SB. A review of major Crohn’s disease susceptibility genes and their role in disease pathogenesis. Genes Genom. 2011;33:317–325. [Google Scholar]
- 17.Jansson J, Willing B, Lucio M, Fekete A, Dicksved J, Halfvarson J, Tysk C, Schmitt-Kopplin P. Metabolomics reveals metabolic biomarkers of Crohn’s disease. PLoS One. 2009;4:e6386. doi: 10.1371/journal.pone.0006386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Thompson AI, Lees CW. Genetics of ulcerative colitis. Inflamm Bowel Dis. 2011;17:831–848. doi: 10.1002/ibd.21375. [DOI] [PubMed] [Google Scholar]
- 19.Lin HM, Helsby NA, Rowan DD, Ferguson LR. Using metabolomic analysis to understand inflammatory bowel diseases. Inflamm Bowel Dis. 2011;17:1021–1029. doi: 10.1002/ibd.21426. [DOI] [PubMed] [Google Scholar]
- 20.Saito YA. The role of genetics in IBS. Gastroenterol Clin North Am. 2011;40:45–67. doi: 10.1016/j.gtc.2010.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bjerrum JT, Nielsen OH, Hao F, Tang H, Nicholson JK, Wang Y, Olsen J. Metabonomics in ulcerative colitis: diagnostics, biomarker identification, and insight into the pathophysiology. J Proteome Res. 2010;9:954–962. doi: 10.1021/pr9008223. [DOI] [PubMed] [Google Scholar]
- 22.Clarke G, Fitzgerald P, Cryan JF, Cassidy EM, Quigley EM, Dinan TG. Tryptophan degradation in irritable bowel syndrome: evidence of indoleamine 2,3-dioxygenase activation in a male cohort. BMC Gastroenterol. 2009;9:6. doi: 10.1186/1471-230X-9-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kilkens TO, Honig A, van Nieuwenhoven MA, Riedel WJ, Brummer RJ. Acute tryptophan depletion affects brain-gut responses in irritable bowel syndrome patients and controls. Gut. 2004;53:1794–1800. doi: 10.1136/gut.2004.041657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shufflebotham J, Hood S, Hendry J, Hince DA, Morris K, Nutt D, Probert C, Potokar J. Acute tryptophan depletion alters gastrointestinal and anxiety symptoms in irritable bowel syndrome. Am J Gastroenterol. 2006;101:2582–2587. doi: 10.1111/j.1572-0241.2006.00811.x. [DOI] [PubMed] [Google Scholar]
- 25.Murray MF. Tryptophan depletion and HIV infection: a metabolic link to pathogenesis. Lancet Infect Dis. 2003;3:644–652. doi: 10.1016/s1473-3099(03)00773-4. [DOI] [PubMed] [Google Scholar]
- 26.Ciorba MA. Indoleamine 2,3-dioxygenase in intestinal disease. Curr Opin Gastroenterol. 2013;29:146–152. doi: 10.1097/MOG.0b013e32835c9cb3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Coffey RD, Cromwell GL. Use of spray-dried animal plasma in diets for weanling pigs. Pig News Inform. 2001;22:39N–48N. [Google Scholar]
- 28.Pierce JL, Cromwell GL, Lindemann MD, Russell LE, Weaver EM. Effects of spray-dried animal plasma and immunoglobulins on performance of early weaned pigs. J Anim Sci. 2005;83:2876–2885. doi: 10.2527/2005.83122876x. [DOI] [PubMed] [Google Scholar]
- 29.Nofrarías M, Manzanilla EG, Pujols J, Gibert X, Majó N, Segalés J, Gasa J. Effects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J Anim Sci. 2006;84:2735–2742. doi: 10.2527/jas.2005-414. [DOI] [PubMed] [Google Scholar]
- 30.Torrallardona D. Spray dried animal plasma as an alternative to antibiotics in weanling pigs - A review. Asian-Australasian J Anim Sci. 2010;23:131–148. [Google Scholar]
- 31.Asmuth DM, Ma ZM, Albanese A, Sandler NG, Devaraj S, Knight TH, Flynn NM, Yotter T, Garcia JC, Tsuchida E, et al. Oral serum-derived bovine immunoglobulin improves duodenal immune reconstitution and absorption function in patients with HIV enteropathy. AIDS. 2013;27:2207–2217. doi: 10.1097/QAD.0b013e328362e54c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wilson D, Evans M, Weaver E, Shaw AL, Klein GL. Evaluation of serum-derived bovine immunoglobulin protein isolate in subjects with diarrhea-predominant irritable bowel syndrome. Clin Med Insights Gastroenterol. 2013;6:49–60. doi: 10.4137/CGast.S13200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gatnau R, Paul PS, Zimmerman DR. Spray dried porcine plasma as a source of immunoglobulins for newborn piglets. J Anim Sci. 1989;67:244. [Google Scholar]
- 34.Borg BS, Campbell JM, Russel LE, Rodríguez C, Ródenas J. Evaluation of the chemical and biological characteristics of spray-dried plasma protein collected from various locations around the world. Proc Am Assoc Swine Vet. 2002;33:97–100. [Google Scholar]
- 35.Food and Drug Administration. Bovine Globulin -Agency Response Letter GRAS Notice No. GRN 000255. 2008. Available from: http://www.fda.gov/Food/FoodIngredientsPackaging/GenerallyRecognizedasSafeGRAS/GRASListings/ucm154991.htm.
- 36.EnteraGam Package Insert, 2014. Available from: http://www.enterahealth.com/enteragam.
- 37.Jiang R, Chang X, Stoll B, Ellis KJ, Shypailo RJ, Weaver E, Campbell J, Burrin DG. Dietary plasma protein is used more efficiently than extruded soy protein for lean tissue growth in early-weaned pigs. J Nutr. 2000;130:2016–2019. doi: 10.1093/jn/130.8.2016. [DOI] [PubMed] [Google Scholar]
- 38.Hanning RM, Drew M. Bovine Immunoglobulin Feeding Trial. 1994 [Google Scholar]
- 39.Morel PCH, Shollum LM, Buwalda TR, Pearson G. Digestibility of bovine immunoglobulin in the piglet. In: Hennessy DP, Cranwell PD, editors. Manipulating Pig Production; 1995; Canberra, Australia. Australia: CSIRO Publishing; 1995. p. 181. [Google Scholar]
- 40.Rodriguez C, Blanch F, Romano V. Porcine immunoglobulins survival in the intestinal tract of adult dogs and cats fed dry food kibbles containing spray-dried porcine plasma (SDPP) or porcine immunoglobulin concentrate (PIC) Anim Feed Sci Technol. 2007;139:201–211. [Google Scholar]
- 41.Lembcke JL, Peerson JM, Brown KH. Acceptability, safety, and digestibility of spray-dried bovine serum added to diets of recovering malnourished children. J Pediatr Gastroenterol Nutr. 1997;25:381–384. doi: 10.1097/00005176-199710000-00003. [DOI] [PubMed] [Google Scholar]
- 42.Bégin F, Santizo MC, Peerson JM, Torún B, Brown KH. Effects of bovine serum concentrate, with or without supplemental micronutrients, on the growth, morbidity, and micronutrient status of young children in a low-income, peri-urban Guatemalan community. Eur J Clin Nutr. 2008;62:39–50. doi: 10.1038/sj.ejcn.1602682. [DOI] [PubMed] [Google Scholar]
- 43.Corl BA, Harrell RJ, Moon HK, Phillips O, Weaver EM, Campbell JM, Arthington JD, Odle J. Effect of animal plasma proteins on intestinal damage and recovery of neonatal pigs infected with rotavirus. J Nutr Biochem. 2007;18:778–784. doi: 10.1016/j.jnutbio.2006.12.011. [DOI] [PubMed] [Google Scholar]
- 44.Bosi P, Casini L, Finamore A, Cremokolini C, Merialdi G, Trevisi P, Nobili F, Mengheri E. Spray-dried plasma improves growth performance and reduces inflammatory status of weaned pigs challenged with enterotoxigenic Escherichia coli K88. J Anim Sci. 2004;82:1764–1772. doi: 10.2527/2004.8261764x. [DOI] [PubMed] [Google Scholar]
- 45.Hunt E, Fu Q, Armstrong MU, Rennix DK, Webster DW, Galanko JA, Chen W, Weaver EM, Argenzio RA, Rhoads JM. Oral bovine serum concentrate improves cryptosporidial enteritis in calves. Pediatr Res. 2002;51:370–376. doi: 10.1203/00006450-200203000-00017. [DOI] [PubMed] [Google Scholar]
- 46.Peace RM, Campbell J, Polo J, Crenshaw J, Russell L, Moeser A. Spray-dried porcine plasma influences intestinal barrier function, inflammation, and diarrhea in weaned pigs. J Nutr. 2011;141:1312–1317. doi: 10.3945/jn.110.136796. [DOI] [PubMed] [Google Scholar]
- 47.Pérez-Bosque A, Amat C, Polo J, Campbell JM, Crenshaw J, Russell L, Moretó M. Spray-dried animal plasma prevents the effects of Staphylococcus aureus enterotoxin B on intestinal barrier function in weaned rats. J Nutr. 2006;136:2838–2843. doi: 10.1093/jn/136.11.2838. [DOI] [PubMed] [Google Scholar]
- 48.Moretó M, Pérez-Bosque A. Dietary plasma proteins, the intestinal immune system, and the barrier functions of the intestinal mucosa. J Anim Sci. 2009;87:E92–100. doi: 10.2527/jas.2008-1381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Campbell JM, Polo J, Russell LE, J D. C. Review of spray-dried plasma’s impact on intestinal barrier function. Livestock Sci. 2010;133:239–241. [Google Scholar]
- 50.Tana C, Umesaki Y, Imaoka A, Handa T, Kanazawa M, Fukudo S. Altered profiles of intestinal microbiota and organic acids may be the origin of symptoms in irritable bowel syndrome. Neurogastroenterol Motil. 2010;22:512–519, e114-e115. doi: 10.1111/j.1365-2982.2009.01427.x. [DOI] [PubMed] [Google Scholar]
- 51.Rajilić-Stojanović M, Biagi E, Heilig HG, Kajander K, Kekkonen RA, Tims S, de Vos WM. Global and deep molecular analysis of microbiota signatures in fecal samples from patients with irritable bowel syndrome. Gastroenterology. 2011;141:1792–1801. doi: 10.1053/j.gastro.2011.07.043. [DOI] [PubMed] [Google Scholar]
- 52.Kassinen A, Krogius-Kurikka L, Mäkivuokko H, Rinttilä T, Paulin L, Corander J, Malinen E, Apajalahti J, Palva A. The fecal microbiota of irritable bowel syndrome patients differs significantly from that of healthy subjects. Gastroenterology. 2007;133:24–33. doi: 10.1053/j.gastro.2007.04.005. [DOI] [PubMed] [Google Scholar]
- 53.Parkes GC, Rayment NB, Hudspith BN, Petrovska L, Lomer MC, Brostoff J, Whelan K, Sanderson JD. Distinct microbial populations exist in the mucosa-associated microbiota of sub-groups of irritable bowel syndrome. Neurogastroenterol Motil. 2012;24:31–39. doi: 10.1111/j.1365-2982.2011.01803.x. [DOI] [PubMed] [Google Scholar]
- 54.Saulnier DM, Riehle K, Mistretta TA, Diaz MA, Mandal D, Raza S, Weidler EM, Qin X, Coarfa C, Milosavljevic A, et al. Gastrointestinal microbiome signatures of pediatric patients with irritable bowel syndrome. Gastroenterology. 2011;141:1782–1791. doi: 10.1053/j.gastro.2011.06.072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Roediger WE, Moore J, Babidge W. Colonic sulfide in pathogenesis and treatment of ulcerative colitis. Dig Dis Sci. 1997;42:1571–1579. doi: 10.1023/a:1018851723920. [DOI] [PubMed] [Google Scholar]
- 56.Ott SJ, Musfeldt M, Wenderoth DF, Hampe J, Brant O, Fölsch UR, Timmis KN, Schreiber S. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut. 2004;53:685–693. doi: 10.1136/gut.2003.025403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Frank DN, St Amand AL, Feldman RA, Boedeker EC, Harpaz N, Pace NR. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc Natl Acad Sci USA. 2007;104:13780–13785. doi: 10.1073/pnas.0706625104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Khor B, Gardet A, Xavier RJ. Genetics and pathogenesis of inflammatory bowel disease. Nature. 2011;474:307–317. doi: 10.1038/nature10209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Morgan XC, Tickle TL, Sokol H, Gevers D, Devaney KL, Ward DV, Reyes JA, Shah SA, LeLeiko N, Snapper SB, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012;13:R79. doi: 10.1186/gb-2012-13-9-r79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Brown K, DeCoffe D, Molcan E, Gibson DL. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients. 2012;4:1095–1119. doi: 10.3390/nu4081095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Jeffery IB, O’Toole PW. Diet-microbiota interactions and their implications for healthy living. Nutrients. 2013;5:234–252. doi: 10.3390/nu5010234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Asmuth DM, Stombaugh J, Ma ZM, Albanese A, Hodzic E, Troia-Cancio P, Flynn NM, Yotter T, Miller CJ, Knight R. Changes in stool microbiota, bacterial translocation and mucosal immunity after oral serum-derived bovine immunoglobulin (SBI) administration. 20th Conference on Retroviruses and Opportunistic Infections (CROI); Mar 3-6; Atlanta, GA. Atlanta: CROI; 2013. pp. B–186. [Google Scholar]
- 63.Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444:1022–1023. doi: 10.1038/4441022a. [DOI] [PubMed] [Google Scholar]
- 64.Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, Fernandes GR, Tap J, Bruls T, Batto JM, et al. Enterotypes of the human gut microbiome. Nature. 2011;473:174–180. doi: 10.1038/nature09944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Dinarello CA. Role of pro- and anti-inflammatory cytokines during inflammation: experimental and clinical findings. J Biol Regul Homeost Agents. 1997;11:91–103. [PubMed] [Google Scholar]
- 66.Moretó M, Miró L, Maijó M, Weaver E, Crenshaw JD, Russell L, Campbell J, Perez-Bosque A. Dietary supplementation with porcine plasma proteins reduce lymphocyte recruitment and cytokine and chemokine expression in a mouse model of spontaneous colitis. Gastroenterology. 2010;138:S–743.W1801. [Google Scholar]
- 67.Jiang H, Becker C, Przybyszewski J, MacDonald RS. Dietary immunoglobulins affect colon cytokines in mouse model of inflammatory bowel disease. FASEB J. 2010;24:720.1. [Google Scholar]
- 68.Pérez-Bosque A, Miró L, Polo J, Russell L, Campbell J, Weaver E, Crenshaw J, Moretó M. Dietary plasma proteins modulate the immune response of diffuse gut-associated lymphoid tissue in rats challenged with Staphylococcus aureus enterotoxin B. J Nutr. 2008;138:533–537. doi: 10.1093/jn/138.3.533. [DOI] [PubMed] [Google Scholar]
- 69.Pérez-Bosque A, Miró L, Polo J, Russell L, Campbell J, Weaver E, Crenshaw J, Moretó M. Dietary plasma protein supplements prevent the release of mucosal proinflammatory mediators in intestinal inflammation in rats. J Nutr. 2010;140:25–30. doi: 10.3945/jn.109.112466. [DOI] [PubMed] [Google Scholar]
- 70.Pérez-Bosque A, Pelegrí C, Vicario M, Castell M, Russell L, Campbell JM, Quigley JD, Polo J, Amat C, Moretó M. Dietary plasma protein affects the immune response of weaned rats challenged with S. aureus Superantigen B. J Nutr. 2004;134:2667–2672. doi: 10.1093/jn/134.10.2667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Pérez-Bosque A, Miró L, Maijó M, Polo J, Campbell J, Russell L, Crenshaw J, Weaver E, Moretó M. Dietary inclusion of serum-derived bovine immunoglobulins ameliorates colitis in mdr1a-/- mice. J Crohn’s Colitis. 2014 [Google Scholar]
- 72.Abreu MT, Fukata M, Arditi M. TLR signaling in the gut in health and disease. J Immunol. 2005;174:4453–4460. doi: 10.4049/jimmunol.174.8.4453. [DOI] [PubMed] [Google Scholar]
- 73.Liu Y, Rhoads JM. Communication between B-Cells and Microbiota for the Maintenance of Intestinal Homeostasis. Antibodies. 2013;2:535–553. [Google Scholar]
- 74.Tak PP, Firestein GS. NF-kappaB: a key role in inflammatory diseases. J Clin Invest. 2001;107:7–11. doi: 10.1172/JCI11830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Weaver EM, Klein GL, DeVries BK, Maas KJ, Shaw AL. Endotoxin Neutralization activity (ENA) of bovine plasma and bovine Immunoglobulin (IgG)-rich fractions as compared to human plasma. FASEB J. 2013;27:1079.58. [Google Scholar]
- 76.Navarro A, Eslava C, García de la Torre G, León LA, Licona D, León L, Zarco LA, Cravioto A. Common epitopes in LPS of different Enterobacteriaceae are associated with an immune response against Escherichia coli O157 in bovine serum samples. J Med Microbiol. 2007;56:1447–1454. doi: 10.1099/jmm.0.47201-0. [DOI] [PubMed] [Google Scholar]
- 77.Pérez-Bosque A, Moretó M. A rat model of mild intestinal inflammation induced by Staphylococcus aureus enterotoxin B. Proc Nutr Soc. 2010;69:447–453. doi: 10.1017/S0029665110001849. [DOI] [PubMed] [Google Scholar]
- 78.Kaneko Y, Nimmerjahn F, Madaio MP, Ravetch JV. Pathology and protection in nephrotoxic nephritis is determined by selective engagement of specific Fc receptors. J Exp Med. 2006;203:789–797. doi: 10.1084/jem.20051900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Mouzaki A, Theodoropoulou M, Gianakopoulos I, Vlaha V, Kyrtsonis MC, Maniatis A. Expression patterns of Th1 and Th2 cytokine genes in childhood idiopathic thrombocytopenic purpura (ITP) at presentation and their modulation by intravenous immunoglobulin G (IVIg) treatment: their role in prognosis. Blood. 2002;100:1774–1779. [PubMed] [Google Scholar]
- 80.Negi VS, Elluru S, Sibéril S, Graff-Dubois S, Mouthon L, Kazatchkine MD, Lacroix-Desmazes S, Bayry J, Kaveri SV. Intravenous immunoglobulin: an update on the clinical use and mechanisms of action. J Clin Immunol. 2007;27:233–245. doi: 10.1007/s10875-007-9088-9. [DOI] [PubMed] [Google Scholar]
- 81.Brenchley JM, Douek DC. Microbial translocation across the GI tract. Annu Rev Immunol. 2012;30:149–173. doi: 10.1146/annurev-immunol-020711-075001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Al-Sadi R, Boivin M, Ma T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front Biosci (Landmark Ed) 2009;14:2765–2778. doi: 10.2741/3413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Camilleri M. Peripheral mechanisms in irritable bowel syndrome. N Engl J Med. 2012;367:1626–1635. doi: 10.1056/NEJMra1207068. [DOI] [PubMed] [Google Scholar]
- 84.Cummins PM. Occludin: one protein, many forms. Mol Cell Biol. 2012;32:242–250. doi: 10.1128/MCB.06029-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Prasad S, Mingrino R, Kaukinen K, Hayes KL, Powell RM, MacDonald TT, Collins JE. Inflammatory processes have differential effects on claudins 2, 3 and 4 in colonic epithelial cells. Lab Invest. 2005;85:1139–1162. doi: 10.1038/labinvest.3700316. [DOI] [PubMed] [Google Scholar]
- 86.Jian ZM, Cao JD, Zhu XG, Zhao WX, Yu JC, Ma EL, Wang XR, Zhu MW, Shu H, Liu YW. The impact of alanyl-glutamine on clinical safety, nitrogen balance, intestinal permeability, and clinical outcome in postoperative patients: a randomized, double-blind, controlled study of 120 patients. JPEN J Parenter Enteral Nutr. 1999;23:S62–S66. doi: 10.1177/014860719902300516. [DOI] [PubMed] [Google Scholar]
- 87.Peuhkuri K, Vapaatalo H, Korpela R. Even low-grade inflammation impacts on small intestinal function. World J Gastroenterol. 2010;16:1057–1062. doi: 10.3748/wjg.v16.i9.1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Spiller R, Garsed K. Infection, inflammation, and the irritable bowel syndrome. Dig Liver Dis. 2009;41:844–849. doi: 10.1016/j.dld.2009.07.007. [DOI] [PubMed] [Google Scholar]
- 89.Spiller RC. Role of infection in irritable bowel syndrome. J Gastroenterol. 2007;42 Suppl 17:41–47. doi: 10.1007/s00535-006-1925-8. [DOI] [PubMed] [Google Scholar]
- 90.Martínez C, Lobo B, Pigrau M, Ramos L, González-Castro AM, Alonso C, Guilarte M, Guilá M, de Torres I, Azpiroz F, et al. Diarrhoea-predominant irritable bowel syndrome: an organic disorder with structural abnormalities in the jejunal epithelial barrier. Gut. 2013;62:1160–1168. doi: 10.1136/gutjnl-2012-302093. [DOI] [PubMed] [Google Scholar]
- 91.Ohman L, Simrén M. Pathogenesis of IBS: role of inflammation, immunity and neuroimmune interactions. Nat Rev Gastroenterol Hepatol. 2010;7:163–173. doi: 10.1038/nrgastro.2010.4. [DOI] [PubMed] [Google Scholar]
- 92.Kapembwa MS, Fleming SC, Sewankambo N, Serwadda D, Lucas S, Moody A, Griffin GE. Altered small-intestinal permeability associated with diarrhoea in human-immunodeficiency-virus-infected Caucasian and African subjects. Clin Sci (Lond) 1991;81:327–334. doi: 10.1042/cs0810327. [DOI] [PubMed] [Google Scholar]
- 93.Bjarnason I, Sharpstone DR, Francis N, Marker A, Taylor C, Barrett M, Macpherson A, Baldwin C, Menzies IS, Crane RC, et al. Intestinal inflammation, ileal structure and function in HIV. AIDS. 1996;10:1385–1391. doi: 10.1097/00002030-199610000-00011. [DOI] [PubMed] [Google Scholar]
- 94.Sharpstone D, Neild P, Crane R, Taylor C, Hodgson C, Sherwood R, Gazzard B, Bjarnason I. Small intestinal transit, absorption, and permeability in patients with AIDS with and without diarrhoea. Gut. 1999;45:70–76. doi: 10.1136/gut.45.1.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Marchetti G, Cozzi-Lepri A, Merlini E, Bellistrì GM, Castagna A, Galli M, Verucchi G, Antinori A, Costantini A, Giacometti A, et al. Microbial translocation predicts disease progression of HIV-infected antiretroviral-naive patients with high CD4+ cell count. AIDS. 2011;25:1385–1394. doi: 10.1097/QAD.0b013e3283471d10. [DOI] [PubMed] [Google Scholar]
- 96.Sandler NG, Wand H, Roque A, Law M, Nason MC, Nixon DE, Pedersen C, Ruxrungtham K, Lewin SR, Emery S, et al. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J Infect Dis. 2011;203:780–790. doi: 10.1093/infdis/jiq118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Knights D, Lassen KG, Xavier RJ. Advances in inflammatory bowel disease pathogenesis: linking host genetics and the microbiome. Gut. 2013;62:1505–1510. doi: 10.1136/gutjnl-2012-303954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.El-Salhy M. Irritable bowel syndrome: diagnosis and pathogenesis. World J Gastroenterol. 2012;18:5151–5163. doi: 10.3748/wjg.v18.i37.5151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wong BS, Camilleri M, Carlson P, McKinzie S, Busciglio I, Bondar O, Dyer RB, Lamsam J, Zinsmeister AR. Increased bile acid biosynthesis is associated with irritable bowel syndrome with diarrhea. Clin Gastroenterol Hepatol. 2012;10:1009–1015.e3. doi: 10.1016/j.cgh.2012.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Yao CK, Tan HL, van Langenberg DR, Barrett JS, Rose R, Liels K, Gibson PR, Muir JG. Dietary sorbitol and mannitol: food content and distinct absorption patterns between healthy individuals and patients with irritable bowel syndrome. J Hum Nutr Diet. 2014;27 Suppl 2:263–275. doi: 10.1111/jhn.12144. [DOI] [PubMed] [Google Scholar]
- 101.Putkonen L, Yao CK, Gibson PR. Fructose malabsorption syndrome. Curr Opin Clin Nutr Metab Care. 2013;16:473–477. doi: 10.1097/MCO.0b013e328361c556. [DOI] [PubMed] [Google Scholar]
- 102.Ligaarden SC, Farup PG. Low intake of vitamin B6 is associated with irritable bowel syndrome symptoms. Nutr Res. 2011;31:356–361. doi: 10.1016/j.nutres.2011.04.001. [DOI] [PubMed] [Google Scholar]
- 103.Maijó M, Miró L, Polo J, Campbell J, Russell L, Crenshaw J, Weaver E, Moretó M, Pérez-Bosque A. Dietary plasma proteins attenuate the innate immunity response in a mouse model of acute lung injury. Br J Nutr. 2012;107:867–875. doi: 10.1017/S0007114511003655. [DOI] [PubMed] [Google Scholar]