

Summary
The authors identify a pentatricopeptide repeat (PPR) protein SOAR1 as a crucial player of ABA signalling, which localizes to both the cytosol and nucleus probably to regulate nuclear gene expression.
Key words: Abscisic acid signalling, Arabidopsis thaliana, Mg-chelatase H subunit, pentatricopeptide repeat (PPR) protein, post-germination growth, seed germination.
Abstract
A dominant suppressor of the ABAR overexpressor, soar1-1D, from CHLH/ABAR [coding for Mg-chelatase H subunit/putative abscisic acid (ABA) receptor (ABAR)] overexpression lines was screened to explore the mechanism of the ABAR-mediated ABA signalling. The SOAR1 gene encodes a pentatricopeptide repeat (PPR) protein which localizes to both the cytosol and nucleus. Down-regulation of SOAR1 strongly enhances, but up-regulation of SOAR1 almost completely impairs, ABA responses, revealing that SOAR1 is a critical, negative, regulator of ABA signalling. Further genetic evidence supports that SOAR1 functions downstream of ABAR and probably upstream of an ABA-responsive transcription factor ABI5. Changes in the SOAR1 expression alter expression of a subset of ABA-responsive genes including ABI5. These findings provide important information to elucidate further the functional mechanism of PPR proteins and the complicated ABA signalling network.
Introduction
Pentatricopeptide repeat (PPR) proteins are a class of RNA-binding proteins characterized by the presence of a degenerate 35 amino acid repeat, the PPR motif, which is repeated in tandem 2–50 times. The PPR motifs form a helical structure and are considered to be RNA-binding motifs (Aubourg et al., 2000; Small and Peeters, 2000; Lurin et al., 2004). The first PPR gene was identified in Saccharomyces cerevisiae (Manthey and McKewen, 1995), and it is known that all sequenced eukaryotic genomes have been found to encode PPR proteins, though the numbers of PPR genes in both animal and fungal genomes are relatively small. The PPR domain protein family is particularly large in land plants. In the Arabidopsis thaliana genome, 450 putative PPR genes were identified, and >600 PPR genes have been predicted to occur in the rice genome (Small and Peeters, 2000; Lurin et al., 2004; Rivals et al., 2006; Schmitz-Linneweber and Small, 2008).
It has been known that PPR proteins are mostly targeted to mitochondria and/or chloroplasts in plants, and they are involved in many aspects of RNA processing in these two organelles, such as RNA splicing, editing, 5′ and 3′ end processing, stability and cleavage, and translation (Meierhoff et al., 2003; Williams and Barkan, 2003; Lurin et al., 2004). The mitochondrial/chloroplast PPR proteins play diverse and crucial roles in plant developmental processes and responses to environmental stresses (Small and Peeters, 2000; Lurin et al., 2004; Oguchi et al., 2004; Tzafrir et al., 2004; Cushing et al., 2005; Ding et al., 2006; Wang et al., 2006; Gutierrez-Marcos et al., 2007; Koussevitzky et al., 2007; Chi et al., 2008; Fujii and Small, 2011; Hu et al., 2012; Nakamura et al., 2012). Few PPR proteins, however, have been found to localize to cellular compartments other than mitochondria and chloroplasts. To the authors’ knowledge, thus far, two PPR proteins have been identified as localizing to the nucleus, of which one PPR protein was found only in the nucleus and another in both the mitochondrion and the nucleus, and they regulate embryogenesis probably by modulating nuclear gene transcription and RNA processing (Ding et al., 2006; Hammani et al., 2011).
The phytohormone abscisic acid (ABA) regulates many developmental processes and plant adaptation to adverse conditions (reviewed in Finkelstein et al., 2002; Adie et al., 2007; Cutler et al., 2010). Numerous ABA signalling components, including receptors or candidate receptors for ABA, have been identified (Finkelstein et al., 2002, Gao et al., 2007; Johnston et al., 2007; Liu et al., 2007a , b ; Guo et al., 2008; Ma et al., 2009; Pandey et al., 2009; Park et al., 2009; Santiago et al., 2009; Cutler et al., 2010). The START-domain family proteins PYR/PYL/RCAR are the best characterized cytosolic ABA receptors, which mediate a core ABA signalling pathway involving the downstream components such as the type 2C protein phosphatases (PP2Cs), SNF1-related protein kinase 2s (SnRK2s), and a clade of bZIP-domain transcription factors (Fujii et al., 2009; Ma et al., 2009; Park et al., 2009; Santiago et al., 2009; Cutler et al., 2010). In the highly complex ABA signalling network, five Arabidopsis PPR proteins, PPR4 (Zsigmond et al., 2008), ABO5 (Liu et al., 2010), PGN (Laluk et al., 2011), SLG1 (Yuan and Liu, 2012), and AHG11 (Murayama et al., 2012), have been identified to play an important role. All five PPR proteins were found to localize to mitochondria and probably regulate reactive oxygen species (ROS) production in this organelle to be involved in ABA signalling (Zsigmond et al., 2008; Liu et al., 2010; Laluk et al., 2011; Murayama et al., 2012; Yuan and Liu, 2012).
It was previously reported that the chloroplast magnesium-protoporphyrin IX chelatase large subunit [Mg-chelatase H subunit CHLH/putative ABA receptor (ABAR)] functions as a candidate receptor for ABA in Arabidopsis (Shen et al., 2006; Wu et al., 2009; Shang et al., 2010; Du et al., 2012; Liu et al., 2012; Yan et al., 2013; Zhang et al., 2013, 2014). Although it is still controversial whether CHLH/ABAR binds ABA (Shen et al., 2006; Muller and Hansson, 2009; Wu et al., 2009; Tsuzuki et al., 2011; Wang et al., 2011; Du et al., 2012), it has been well supported that CHLH/ABAR functions positively in ABA signalling by regulating a complicated pathway, in which the chloroplast protein cochaperonin CPN20 and cytosolic–nuclear protein WRKY18/40/60 transcription repressors are involved (Shen et al., 2006; Wu et al., 2009; Shang et al., 2010; Du et al., 2012; Liu et al., 2012; Yan et al., 2013; Zhang et al., 2013, 2014). There are four abar mutant alleles in Arabidopsis, abar-2, abar-3, cch, and rtl1, which all show altered ABA responses (Shen et al., 2006; Wu et al., 2009; Tsuzuki et al., 2011, 2013; Du et al., 2012). It was shown that CHLH/ABAR also regulates guard cell signalling in response to ABA in tobacco (Nicotiana benthamiana) leaves (Du et al., 2012). Other independent groups demonstrated that CHLH/ABAR mediates ABA signalling in guard cells in both Arabidopsis (Legnaioli et al., 2009; Tsuzuki et al., 2011, 2013) and peach (Prunus persica) leaves (Jia et al., 2011a ). Tsuzuki and co-workers (2013) recently showed that CHLH/ABAR mediates ABA inhibition of blue light (BL)-induced phosphorylation of H+-ATPase in Arabidopsis guard cells, suggesting that CHLH/ABAR regulates not only ABA-induced stomatal closure but also ABA inhibition of BL-mediated stomatal opening. Interestingly, it has been demonstrated that CHLH/ABAR mediates ABA signalling in fruit ripening of both peach (Jia et al., 2011a ) and strawberry (Fragaria ananassa) (Jia et al., 2011b ). These data demonstrate that CHLH/ABAR is an essential ABA signalling regulator in plant cells.
To explore further the mechanism of the CHLH/ABAR-mediated ABA signalling, a suppressor of the ABAR overexpressor (named soar1-1D), in which the expression levels of both the CHLH/ABAR gene and the SOAR1 gene encoding a PPR-motif protein, are up-regulated, was screened. It was shown that SOAR1 localizes to both the cytosol and nucleus, and functions as a critical, negative, regulator of the ABA signalling pathway in seed germination and seedling growth. Genetic evidence revealed that SOAR1 acts downstream of ABAR and probably upstream of a nuclear ABA-responsive bZIP transcription factor ABA-INSENSITIVE5 (ABI5). These findings provide important information to elucidate further the functional mechanism of PPR proteins and the highly complicated ABA signalling network.
Materials and methods
Plant materials and growth conditions
ABAR overexpression lines were generated by introducing an ABAR gene (At5g13630) fragment (encoding a truncated ABAR with amino acid residues 631–999) into A. thaliana ecotype Columbia-0 (Col) plants as a green fluorescent protein (GFP) fusion protein. It was previously shown that the N-terminally truncated ABAR tagged with GFP functions similarly to full-length ABAR in transgenic plants, leading to ABA hypersensitivity in the major ABA responses (Wu et al., 2009). Therefore, the truncated ABAR overexpression lines were used as ABAR overexpressors. The cDNA isolation and transgenic manipulation were as previously described (Wu et al., 2009). From the population of the ABAR overexpression transgenic lines, the lines with ABA-insensitive or wild-type phenotypes in seed germination and post-germination growth were screened, which gave candidate suppressors of the ABAR overexpressor (named soar mutant) lines. The soar1-1D mutant was identified from these candidate soar mutant lines.
The soar1-2 and soar1-3 (stock nos FLAG_546D07and FLAG_500B04, respectively, with the Col ecotype as background) were obtained from Versailles Genetics and Plant Breeding Laboratory, Arabidopsis thaliana Resource Centre (INRA; http://dbsgap.versailles.inra.fr/portail/). The seed of the abi5-1 (stock no. CS8105, with the Wassilewskija ecotype as background; locus of the ABI5 gene, At2g36270) mutant was obtained from the Arabidopsis Biological Resource Center (ABRC; http://abrc.osu.edu/), and the background of the abi5-1 mutant was changed to the Col ecotype by crossing as described previously (Shang et al., 2010). The soar1-2 abi5-1 double mutant was created by crossing. The seeds of abi1-3 (stock no. SALK_076309) and abi2-2 (stock no. SALK_015166) mutants were also obtained from the ABRC, and the mutants are T-DNA insertion knockout alleles in the ABI1 (At4g26080) and ABI2 (At5g57050) genes, respectively. The background of both mutants is the Col ecotype. The abi1-3 abi2-2 double mutant is a generous gift from Y. Guo’s laboratory (College of Biological Sciences, China Agricultural University). All the primers for identification of the above-mentioned mutants are listed in Supplementary Table S1 available at JXB online.
The A. thaliana ecotype Col was used to generate transgenic plants. To generate the SOAR1 (At5g11310) overexpression lines, the full-length SOAR1 cDNA, amplified by PCR with the primers listed in Supplementary Table S1 at JXB online, was cloned into the binary vector pCAMBIA1300 (http://www.cambia.org), which contains the Cauliflower mosaic virus (CaMV) 35S promoter and the C-terminal GFP flag. Also, an ABAR and SOAR1 double gene overexpression line was created by crossing an ABAR overexpressor with a SOAR1 overexpressor (OE1) to test genetic interaction of these two genes. A previously generated ABI2 overexpression line (Sun et al., 2011; the ABI2-2 line harbouring GFP-tagged full-length ABI2 under the Col background) was used as a control in phenotypic analysis of the SOAR1 overexpression lines, which showed strong ABA-insensitive phenotypes (Sun et al., 2011) and was renamed ABI2-OE in the present study. To generate the transgenic complementation lines of the soar1-2 and soar1-3 mutants, the native promoter, isolated by PCR with the primers listed in Supplementary Table S1 was used, to replace the CaMV 35S promoter in the above-mentioned construct to create the native promoter-driven SOAR1 construct. These constructs were introduced, respectively, into Agrobacterium tumefaciens strain GV3101 and transformed into the wild-type plants (for the SOAR1-overexpressiong lines) or soar1 mutants (for the complementation lines of the soar1-2 and soar1-3 mutants) by the floral dip infiltration method (Clough and Bent, 1998). The homozygous T3 seeds of the transgenic plants were used for analysis.
Plants were grown on Murashige and Skoog (MS) medium (Murashige and Skoog, 1962) containing 3% (w/v) sucrose and 0.8% agar or in compost soil under a 16h photoperiod in a growth chamber at ~20 °C. Plants were grown under a 12h photoperiod at ~20 °C for protoplast preparation.
SOAR1 promoter–GUS transformation assay
The promoter of SOAR1 (pSOAR1) was amplified by PCR using forward primer 5ʹ-AACTGCAGTTCCGACAAACATAAAATGG TA-3ʹ and reverse primer 5ʹ-CGGGATCCTCCGCCGAGAAAATT AGGACA-3ʹ. The PCR product was digested and cloned into the pCAMBIA1391 vector. The construct pSOAR1-GUS (β-glucuronidase) was transformed into Arabidopsis Col plants by floral infiltration. Histochemical staining was performed, as described previously (Jefferson et al., 1987), by soaking whole plants or tissues in a solution consisting of 1mM X-gluc, 100mM sodium phosphate buffer (pH 7.0), 0.05mM K3Fe(CN)6, 0.05mM K4Fe(CN)6, 2mM EDTA, and 0.1% (v/v) Triton X-100 at 37 °C for 5–8h. After GUS staining, chlorophyll was cleared from the tissues with a mixture of 30% acetic acid and 70% ethanol, and then the samples were investigated under a stereomicroscope (Olympus).
Quantitative real-time PCR and TAIL-PCR
Quantitative real-time PCR for mRNA expression levels of various genes (see Supplementary Table S1 at JXB online for the gene-specific primers) was performed as previously described (Shang et al., 2010) essentially according to the instructions provided for the Bio-Rad Real-Time System CFX96TM C1000 thermal cycler (Bio-Rad, Hercules, CA, USA). The T-DNA flanking sequence in the soar1-1 dominant mutant was determined by TAIL-PCR (thermal asymmetric interlaced PCR) with pCAMBIA1300-specific left border and random primers that are listed in Supplementary Table S1. The reaction program for rounds was described previously (Liu et al., 1995).
Seeds were stratificated for 3 d at 4 °C, incubated on filter paper imbibed with ABA-free or ABA-containing solution for 24h in a light growth chamber at 20 °C, and collected for RNA extraction. Total RNA was isolated from these germinating seeds with the RNasy plant mini kit (Qiagen) supplemented with an on-column DNA digestion (Qiagen RNase-Free DNase set) according to the manufacturer’s instructions, and then the RNA sample was reverse-transcribed with the Superscript II RT kit (Invitrogen) in a 25ml volume at 42 °C for 1h. Amplification of ACTIN2/8 genes was used as an internal control. The cDNA was amplified using SYBR Premix Ex Taq (TaKaRa) with a DNA Engine Opticon 2 thermal cycler in a 10ml volume. The Ct (threshold cycle), defined as the PCR cycle at which a statistically significant increase of reporter fluorescence was detected, was used as a measure for the starting copy numbers of the target gene. Relative quantitation of the target gene expression level was performed using the comparative Ct method.
Arabidopsis protoplast and onion epidermis transformation
Transient transgenic manipulation in both Arabidopsis protoplasts and onion epidermis was used to assay the subcellular localization of the SOAR1 protein essentially as described previously (Shang et al., 2010). The full-length SOAR1 and a fragment of 106–1809bp downstream of the transcription start site of SOAR1 (SOAR1 106–1809, which encodes a truncated SOAR1 from amino acid residue 36 to 603 with the N-terminal 35 amino acid residues deleted) were amplified by PCR, and the products were cloned into the p-EASY-T1 vector (Transgen, Beijing, China) for sequencing, and then fused with GFP and inserted into the pROK219 vector, driven by the CaMV 35S promoter. The positive control cytosolic–nuclear marker PYR1 (At4g17870; Park et al., 2009) and nuclear marker FBI1 (At1g02340; Lee et al., 2002) were used as described previously (Zhao et al., 2011), and their cDNAs were amplified and fused with mCherry (Shaner et al., 2004) in-frame into the pROK219 vector, driven by the CaMV 35S promoter, respectively. The primers for cloning the full-length SOAR1, SOAR1 106–1809, PYR1, and FBI1 are listed in Supplementary Table S1 at JXB online. Protoplasts were transiently transformed using the polyethylene glycol-mediated transformation protocol (Yoo et al., 2007). The onion epidermal cells were transformed by particle bombardment-mediated transformation with the biolistic PDS-1000/HE gene gun system (Bio-Rad). Bombarded samples were cultured at 26 °C for 16h, and then observed with a confocal laser scanning microscope (ZEISS, Oberkochen, Germany).
Isolation of cytosolic and nuclear fractions
The cytosolic and nuclear fractions were isolated essentially according to the protocol described previously (Shang et al., 2010). Ten-day-old Arabidopsis seedlings were ground to fine powder using liquid nitrogen and pre-chilled using a mortar and pestle. Cytosolic protein isolation buffer is composed of 10mM HEPES, pH 8.0, 250mM sucrose, 0.5% (v/v) Triton X-100, 1mM EDTA, 5mM MgCl2, 50mM NaCl, 1mM phenylmethylsulphonyl fluoride (PMSF), and 1× Roche Cocktail (protease inhibitor cocktail). The buffer was added at 1ml g–1 to powder to generate the homogenate. After centrifuging at 10 000 g for 15min, the supernatant was mixed with 2× SDS sample buffer and denatured for 10min in boiling water. The isolated cytosolic fraction was examined by immunodetecting the presence of the nuclear marker histone H3 with anti-histone H3 antibody (Sigma-Aldrich) to verify that the cytosolic fraction was not contaminated by the nuclear fraction. The nuclear fraction was isolated according to the protocol of Cold Spring Harbor Laboratory as described on its website, and examined by immunodetecting the presence of the cytosolic marker PEPC (phosphoenolpyruvate carboxylase) with anti-PEPC antibody (Agrisera) to ensure that the nuclear fraction was not contaminated by the cytosolic fraction.
Antiserum production, protein extraction, and immunoblotting
The antisera against ABAR and SOAR1 were produced and tested for specificity essentially with the same procedures as described previously (Wu et al., 2009; Shang et al., 2010). A truncated SOAR1 (303 amino acid residues from 299 to 602) was used as antigen for production of the anti-SOAR1 serum. The primers for cloning the cDNA fragment to produce the truncated SOAR1 in Escherichia coli are listed in Supplementary Table S1 at JXB online. The produced anti-SOAR1 serum was tested and shown to be specific for SOAR1 protein (Supplementary Fig. S3). The extraction of the Arabidopsis total protein, SDS–PAGE, and immunoblotting were done essentially according to previously described procedures (Wu et al., 2009; Shang et al., 2010).
Ten-day-old Arabidopsis seedlings were harvested, ground in liquid nitrogen, then transferred into an Eppendorf tube containing ice-cold extraction buffer composed of 50mM TRIS-HCl, pH 7.5, 150mM NaCl, 1mM EDTA, 0.1% Triton X-100, 10% glycerol, and 1× protease inhibitor cocktail (Roche). The sample was extracted for 15min in ice, and centrifuged three times for 10min each at 16 000 g; the supernatant was transferred to a new Eppendorf tube and centrifuged again at 12 000 g for 20min, and then the concentration of the supernatant was detected by Coomassie Brilliant Blue G-250 (Amresco). The samples were either kept at 0 °C for immediate use or frozen and stored at –80 °C until use.
For the immunoblotting assays, proteins were separated by SDS–PAGE on 10% polyacrylamide gels, and transferred to nitrocellulose membranes (0.45 μm; Amersham Life Science) in a medium consisting of 25mM TRIS-HCl (pH 8.3), 192mM glycine, and 20% (v/v) methanol. After rinsing in TRIS-buffered saline (TBS) containing 10mM TRIS-HCl (pH 7.5) and 150mM NaCl, the blotted membranes were pre-incubated for 3h in a blocking buffer containing 3% (w/v) bovine serum albumin dissolved in TBS supplemented by 0.05% (v/v) Tween-20 (TBST1) and then incubated with gentle shaking for 2h at room temperature with appropriate antibodies. The anti-GFP serum (mouse, YTHX Biotechnology Beijing Limited Company, http://www.ythxbio.com/) and anti-actin serum (rabbit) were diluted ~1:3000, and the anti-SOAR1 serum (rabbit) was diluted ~1:2000 in the blocking buffer. Following extensive washes by TBST1, the membranes were incubated with goat anti-rabbit (or anti-mouse for GFP immunoblot) IgG (Cell Signaling Technology, http://www.cellsignal.com/) conjugated with alkaline phosphatase (diluted ~1:1000 in TBST1) at room temperature for 1h and then washed with TBST2 [50mM TRIS-HCl, pH 7.5, 150mM NaCl, and 0.1% (v/v) Tween-20] and TBS. The locations of antigenic proteins were visualized by incubating the membranes with nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate.
Nucleus and mitochondrion staining in Arabidopsis roots
The roots of the 7-day-old OE1 seedlings were sampled and stained for 30min in 1 μg ml–1 4′,6-diamidino-2-phenylindole (DAPI; Sigma; a nuclear marker), or 15min in 0.250 μM Mito Tracker Red CMXRos (Invitrogen). DAPI was dissolved in ddH2O directly. The Mito Tracker Red CMXRos (a mitochondrion marker) stock solution (250 μM) was dissolved in dimethylsulphoxide, then diluted 1000 times with ddH2O or 10mM phosphate-buffered saline (PBS) (pH 7.4, 10mM Na2HPO4, 10mM NaH2PO4, 8.5g l–1 NaCl) when used for staining. After the staining process, the samples were rinsed several times with ddH2O (for DAPI staining) or 10mM PBS (for the Mito Tracker Red staining). Samples were examined with a Leica TCS SP5 confocal microscope under a 63.0×1.40 oil immersion objective.
Phenotypic analysis
Phenotypic analysis was carried out essentially as previously described (Shen et al., 2006; Wu et al., 2009; Shang et al., 2010). Seeds were harvested and stored at room temperature for ~3–6 months before being used in the experiments. To assay germination and post-germination growth, the MS medium (Sigma-Aldrich, St Louis, MO, USA; full-strength MS) contained 3% (w/v) sucrose and 0.8% (w/v) agar, pH 5.8–6.0, and was supplemented or not with different concentrations of (±)-ABA. The seeds were sown and stratified in the MS medium at 4 °C for 3 d, and then they were placed at 20 °C under light conditions. Germination (emergence of radicals) was scored at the indicated times. Seedling growth was assessed by directly sowing the seeds in ABA-containing MS medium to investigate the response of seedling growth to ABA after germination. Another method was used to assay seedling growth in response to ABA: seeds were germinated after stratification on common MS medium and transferred to MS medium supplemented with different concentrations of (±)-ABA. The time for transfer was 48h or 4 d (as indicated) after stratification. Seedling growth was investigated at the indicated times after the transfer.
Results
Down-regulation of SOAR1 increases, and up-regulation of SOAR1 abolishes, ABA sensitivity in seed germination and post-germination growth
The ABAR overexpression lines show ABA-hypersensitive phenotypes (Shen et al., 2006; Wu et al., 2009; see also the Materials and methods). From the ABAR overexpression lines (see the Materials and methods), which were identified by PCR analysis, a putative soar1 mutant named soar1-1D was screened, which showed an ABA-insensitive phenotype (Fig. 1A). PCR analysis showed that the construct for overexpressing the ABAR gene was inserted into the promoter of the SOAR1 gene in the soar1-1D mutant (Supplementary Fig. S1 at JXB online). The SOAR1 gene (At5g11310) encodes a PPR protein with tandem arrays of 10 predicted PPR motifs (Supplementary Fig. S2). The SOAR1 protein appears not to be a P-type member of the PPRs as it has a C-terminal extension (Supplementary Fig. S2) which is not related to other domains of PPRs according to its sequence. The GFP-tagged, functional, truncated ABAR was detected by immunoblot analysis in this soar1-1D mutant (Fig. 1B). To characterize the mutant further, the antibody against a truncated SOAR1 protein (303 amino acid residues from 299 to 602, see the Materials and methods) was generated, which was shown to be specific for SOAR1 (Supplementary Fig. S3). Immunoblot assays by using this anti-SOAR1 serum showed that the amount of SOAR1 protein in the soar1-1D mutant was enhanced >2-fold compared with the wild-type plants (Fig. 1B), which results from this ‘T-DNA’ (construct for overexpressing the ABAR gene) insertion (Supplementary Fig. S1). The soar1-1D is a dominant allele, of which both the homozygous and heterozygous progeny showed strong ABA-insensitive phenotypes in ABA-induced seed germination inhibition and post-germination growth arrest (Fig. 1A–E). Further, T-DNA insertion mutant alleles, soar1-2 and soar1-3, which down-regulate the SOAR1 expression level were obtained (Fig. 1B; Supplementary Fig. S1). A null allele of the SOAR1 gene was not isolated, probably because the loss-of-function soar1 mutant is lethal. It was observed that, in contrast to the soar1-1D mutant, the soar1-2 and soar1-3 mutants showed ABA-hypersensitive phenotypes in ABA-induced seed germination inhibition and post-germination growth arrest (Fig. 1C–E; Supplementary Fig. S4A–E). The seeds of the soar1-2 and soar1-3 mutants germinated more slowly than the wild-type seeds in the exogenous ABA-free medium, suggesting that these mutant seeds may be overly sensitive to the endogenous ABA at a low, physiological level (Fig. 1C; Supplementary Fig. S4A available at JXB online). The intensity of the ABA-hypersensitive phenotypes of the soar1-2 and soar1-3 mutants was similar to, or stronger than, that of the well-characterized abi1 abi2 double-knockout mutant.
Fig. 1.
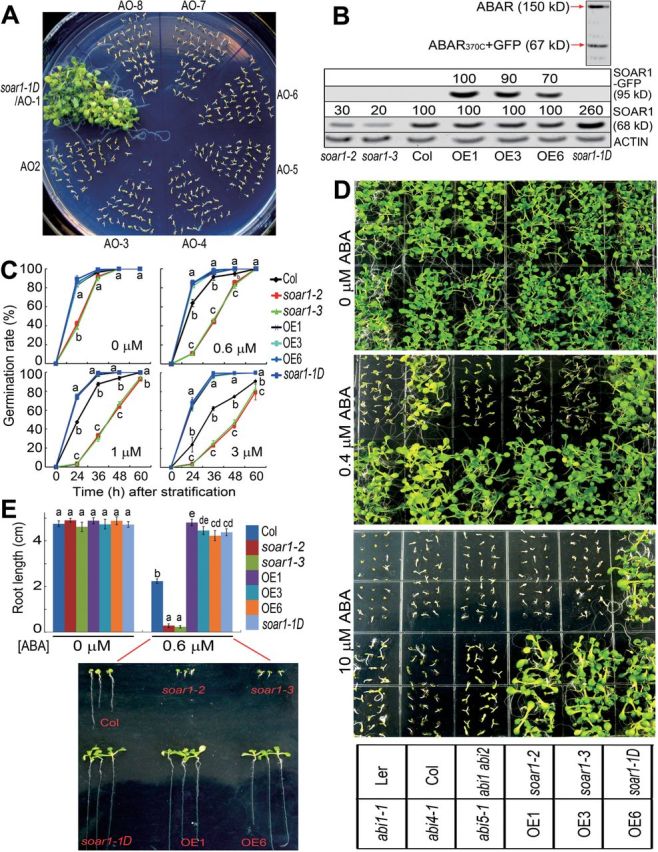
SOAR1 negatively regulates ABA signalling. (A) Screen of the soar1-1D mutant. AO1–AO8 indicate the ABAR overexpression lines 1–8. The seeds were directly planted in 1 μM ABA-containing MS medium, and seedling growth was investigated 10 d after stratification. (B) Immunoblotting assays for the SOAR1–GFP fusion (95kDa) and SOAR1 protein (68kDa) levels in the 10-day-old seedlings of the soar1-1D, soar1-2, and soar1-3 mutants, and SOAR1 overexpression lines (OE1, OE3, and OE6). The SOAR1–GFP fusion and SOAR1 protein amounts were evaluated by scanning the protein bands, and relative band intensities, normalized relative to the band intensity (as 100%) from the sample of the OE1 (for the SOAR1–GFP fusion) or the wild-type Col plants (for the SOAR1 protein), are indicated by numbers above the bands. Actin was used as a loading control. In the soar1-1D mutant, the expression of the truncated ABAR (ABAR370C) tagged by GFP (67kDa), introduced by transgenic manipulation, was tested, and the 150kDa wild-type ABAR was also detected. (C) Seed germination: germination rates of the wild-type Col, soar1-1D, soar1-2, and soar1-3 mutants, and three SOAR1 overexpression lines (OE1, OE3, and OE6) were recorded on ABA-free (0 μM) and ABA-containing (0.6, 1, or 3 μM) MS medium from 24h to 60h after stratification. (D) Early seedling growth: seeds from the wild types Col and Ler, the abi1-1 dominant mutant, the abi1-3 abi2-2 double-knockout mutant, abi4-1 and abi5-1 mutants, and the different genotypes as described in (C) were directly planted in the MS medium supplemented with 0 (top), 0.5 (middle), or 10 μM (±)ABA (bottom), and the growth was investigated 10 d after stratification. (E) Statistical values of the early seedling growth described in (D) from the wild-type Col, soar1-1D, soar1-2, and soar1-3 mutants, and three SOAR1 overexpression lines (OE1, OE3, and OE6) in the MS medium supplemented with 0 and 0.6 μM (±)ABA. The bottom panel shows the pictures of early seedling growth of these genotypes. Each value in (C) and (E) is the mean ±SE of five biological determinations, and different letters indicate significant differences at P<0.05 (Duncan’s multiple range test) when comparing the germination rates among different genotypes at the same time point after stratification (C) or comparing the root lengths among different genotypes in the ABA-free and 0.6 μM ABA-containing medium (E).
SOAR1-overexpressing (OE) lines, in which SOAR1 was fused with GFP, were also generated. Immunoblot analysis detected both the natural SOAR1 and the SOAR1–GFP fusion protein in these OE lines (Fig. 1B; Supplementary Fig. S5B at JXB online). The OE lines showed strong ABA-insensitive phenotypes in ABA-induced seed germination inhibition and post-germination growth arrest (Figs 1C–E, 2A, B; Supplementary Figs S5A, S6A–D). The intensity of ABA-insensitive phenotypes of the OE lines was much stronger than that of the abi1-1 dominant mutant, abi4 and abi5 loss-of-function mutants, and a strong ABI2-overexpressing line ABI2-OE (Figs 1C–E and 2A, B; Supplementary Figs S5A, S6A–D), all of which are well-characterized strong ABA-insensitive mutants (Gosti et al., 1999; Leung et al., 1997; Finkelstein et al., 1998; Finkelstein and Lynch, 2000; Sun et al., 2011). It is noteworthy that the seeds of the SOAR1 OE lines germinated and their post-germination seedlings continued to grow in the medium containing >200 μM (±)ABA, while the wild-type Col seeds generally do not germinate if the medium contains >3 μM (±)ABA, and the strong ABA-insensitive ABI2-OE line did not grow in the 100 μM (±)ABA-containing medium (Fig. 2A; Supplementary Figs. S5A, S6A–D available at JXB online). The 48-hour-old germinating seeds of the SOAR1 OE lines even grew in the medium containing >500 μM (±)ABA (Fig. 2B). These data showed that SOAR1 up-regulation almost completely abolished ABA responses of seeds and young seedlings, indicating the critical role of SOAR1 in ABA signalling.
Fig. 2.
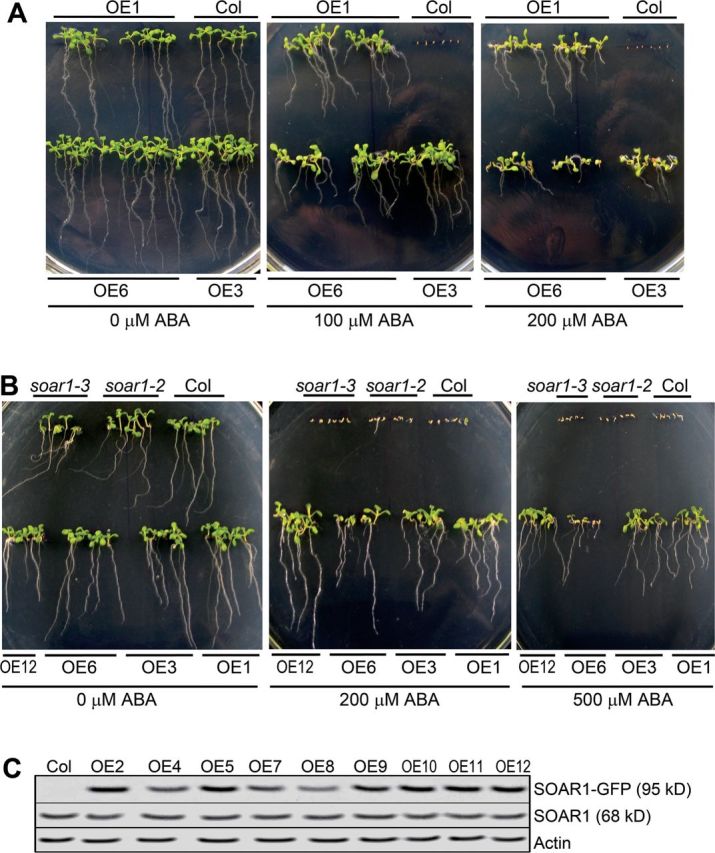
Overexpression of SOAR1 essentially abolishes ABA responses of seeds and young seedlings. (A) Early seedling growth: seeds from the wild-type Col and three SOAR1 overexpression lines (OE1, OE3, and OE6) were directly planted in MS medium supplemented with 0 (left), 100 (middle), or 200 μM (±)ABA (right), and the growth was investigated 2 weeks after stratification. (B) Early seedling growth: germinating seeds of the wild-type Col, soar1-2 and soar1-3 mutants, and four SOAR1 overexpression lines (OE1, OE3, OE6, and OE12) were transferred, 48h after stratification, from ABA-free MS medium to the MS medium supplemented with 0 (left), 200 (middle), or 500 μM (±)ABA, and the growth was investigated 2 weeks after the transfer. (C) Immunoblot analysis of the SOAR1–GFP fusion protein (95kDa) and SOAR1 protein (68kDa) in the 10-day-old seedlings of the wild-type Col and transgenic lines OE2, OE4, OE5, and OE7–OE12, of which the ABA-related phenotypes are presented in B (OE12) and Supplementary Fig. S5 at JXB online. Actin was used as a loading control.
The GFP transgenic lines showed wild-type phenotypes (Supplementary Fig. S5C at JXB online), revealing that the ABA-related phenotypes of the transgenic SOAR1-GFP fusion lines were specific. The complementation lines of the soar1-2 and soar1-3 mutants rescued the ABA-hypersensitive phenotypes of these mutants (Supplementary Fig. S7), demonstrating that the mutation in the SOAR1 gene is responsible for the altered ABA responses of the mutants. Additionally, it was shown that ABA concentrations were not significantly changed in the soar1-2 mutant and the SOAR1-overexpressiing line OE1 compared with the wild-type plants (Supplementary Fig. S8), revealing that ABA metabolism is not significantly affected by changes in SOAR1 expression.
SOAR1 overexpression suppresses ABA hypersensitive phenotypes of the ABAR overexpression lines
The SOAR1 and ABAR double overexpression lines were generated by using an ABAR overexpression line that expressed a truncated ABAR and showed ABA-hypersensitive phenotypes as described previously (Wu et al., 2009). Immunoblot analysis detected the truncated ABAR and SOAR1–GFP fusion proteins in the SOAR1 and ABAR double overexpression lines (Fig. 3A). These double overexpression lines showed strong ABA-insensitive phenotypes in ABA-induced seed germination inhibition and post-germination growth arrest, which were similar to the SOAR1 overexpression lines (Fig. 3B–E). These findings, together with the discovery of the soar1-1D dominant mutant (Fig. 1A, B), reveal that SOAR1 functions downstream of ABAR in the ABA signalling pathway.
Fig. 3.
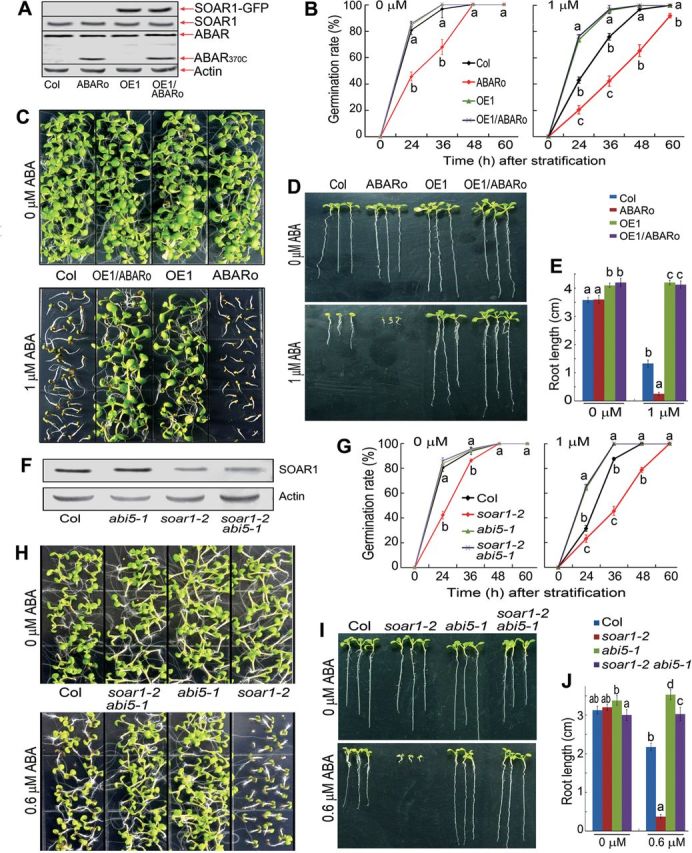
SOAR1 acts genetically downstream of ABAR and upstream of ABI5 in ABA signalling. (A) Immunoblot analysis of the SOAR1–GFP fusion, SOAR1, ABAR, and truncated ABAR (ABAR370C) proteins in 10-day-old seedlings of the wild-type Col, ABAR overexpressor (ABARo), SOAR1 overexpressor (OE1), and ABARo/OE1 double overexpression line. Actin was used as a loading control. (B) Seed germination rates of the different genotypes described in (A), which were recorded on ABA-free (0 μM) and ABA-containing (1 μM) MS medium from 24h to 60h after stratification. (C–E) Early seedling growth of the different genotypes described in (A). Seeds were directly planted in MS medium supplemented with 0 or 1 μM (±)ABA; the growth was investigated (C and D) and root length (E) was measured 10 d after stratification. (F) Immunoblot analysis of the SOAR1 protein in 10-day-old seedlings of the wild-type Col, abi5-1, soar1-2 mutants, and the soar1-2 abi5-1 double mutant. Actin was used as a loading control. (G) Seed germination rates of the different genotypes described in (F), which were recorded on ABA-free (0 μM) and ABA-containing (1 μM) MS medium from 24h to 60h after stratification. (H–J) Early seedling growth of the different genotypes described in (F). Seeds were directly planted in MS medium supplemented with 0 or 0.6 μM (±)ABA; the growth was investigated (H and I) and root length (J) was measured 10 d after stratification. Each value in (B), (E), (G), and (J) is the mean ±SE of five biological determinations, and different letters indicate significant differences at P<0.05 (Duncan’s multiple range test) when comparing the germination rates among different genotypes at the same time point after stratification (B, G) or comparing the root lengths among different genotypes in the ABA-free and ABA-containing medium (E, J).
The SOAR1 gene is expressed in the whole plant but preferentially in seeds, and the SOAR1 protein is localized to both the cytosol and nucleus
The gene expression data in the public websites at http://bar.utoronto.ca and http://www.genevestigator.com showed that the SOAR1 gene is expressed in different organs/tissues, with the highest level in seeds, and this expression profile was confirmed with the SOAR1 promoter–GUS transgenic lines (Supplementary Fig. S9 at JXB online).
As regards the subcellular localization of SOAR1, a bioinformatics search allowed the prediction that SOAR1 may localize to the mitochondrion, chloroplast, or nucleus (Supplementary Fig. S10 at JXB online). The transient expression assays in Arabidopsis protoplasts showed that SOAR1 co-localized with the cytosol–nucleus dual-localized PYR1 (Fig. 4A), which is a member of the PYR/PYL/RCAR receptors for ABA (Ma et al., 2009; Park et al., 2009; Santiago et al., 2009), while the SOAR1 fluorescence was not seen in the chloroplasts (Fig. 4A). However, the cytosolic SOAR1 disappeared, and the SOAR1 fluorescence was seen only in the nucleus and co-localized with a nuclear marker, bHLH (basic helix–loop–helix) transcription factor FBI1 (At1g02340; Fairchild et al., 2000; Jang et al., 2005) when an N-terminal 35 amino acid fragment was deleted (Fig. 4A), suggesting that the N-terminal 35 amino acid fragment is required for the cytosolic localization of the SOAR1 protein. The data from the transient expression assays in onion epidermis cells are consistent with those from the transgenic Arabidopsis protoplasts (Fig. 4B).
Fig. 4.
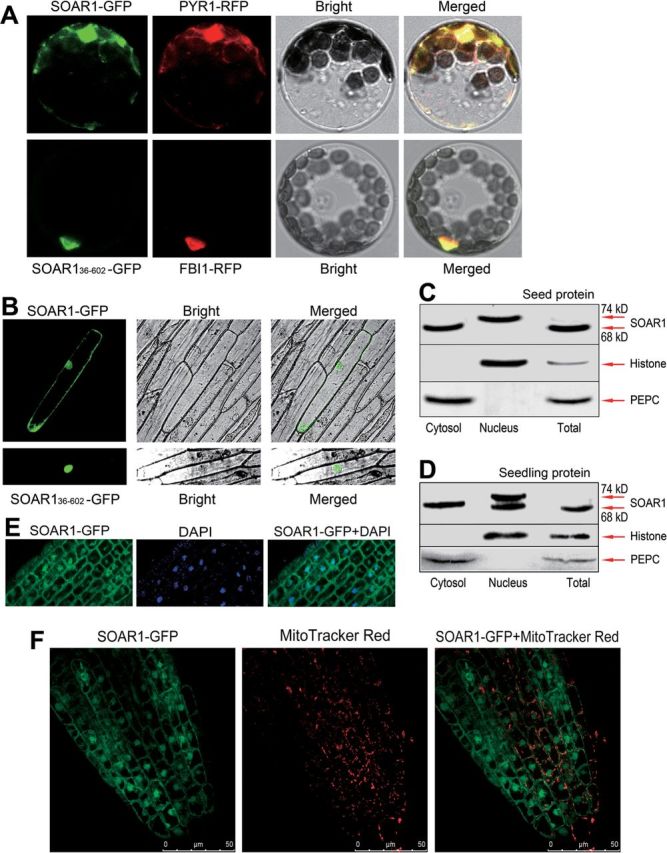
SOAR1 is localized in both the cytosol and nucleus. (A) Transient expression in Arabidopsis protoplasts. Top panels: SOAR1 is co-localized with a nuclear–cytosol-localized protein PYR1 (At4g17870; Park et al., 2009). SOAR1 was tagged with green fluorescent protein (SOAR1–GFP), and PYR1 protein was tagged with mCherry (PYR1–RFP). Bottom panels: the N-terminal truncated SOAR1 (SOAR136–602, with 35 amino acid residues deleted at the N-terminus of the SOAR1 protein; see Supplementary Fig. S1A at JXB online) is co-localized with a nuclear-localized FBI1 bHLH transcription factor (At1g02340; Fairchild et al., 2000; Jang et al., 2005). SOAR136–602 was tagged with GFP (SOAR136–602–GFP), and FBI1 protein was tagged with mCherry (FBI1–RFP). Bright, bright-field; Merged, merged image. The experiments were repeated five times with the same results. (B) Transient expression of the above-mentioned constructs in onion epidermis cells. (C) and (D) Immunoblot analysis of the SOAR1 protein in the total protein (Total), cytosolic (Cytosol), and nuclear (Nucleus) fractions from the seeds sampled 24h after stratification (C) and 10-day-old seedlings (D). Histone H3 (nuclear marker) and PEPC (phosphoenolpyruvate carboxylase; cytosolic marker) were tested in the cytosolic and nuclear fractions to estimate the purity of the fractions. (E) Transgenic expression of the SOAR1–GFP fusion protein in Arabidopsis whole plants. A portion of the SOAR1–GFP fusion protein (left) and a nuclear marker DAPI (middle) are co-localized to the nucleus (right, SOAR1–GFP+DAPI) in the root of the transgenic line OE1. The experiments were repeated five times with the same results. (F) Transgenic expression of the SOAR1–GFP fusion protein in Arabidopsis whole plants. Investigations were performed in the root of the OE1 line. SOAR1–GFP, the distribution pattern of SOAR1–GFP fusion protein (left); Mito Tracker Red, the profile of mitochondria stained by Mito Tracker Red (middle); SOAR1–GFP+Mito Tracker Red, merged image of SOAR1–GFP and Mito Tracker Red (right). The images show that SOAR1–GFP localization is distinct from the Mito Tracker Red-stained mitochondrial profile. The experiments were repeated five times with the same results.
The localization of SOAR1 was further verified by immunoblot assays with the purified cytosolic and nuclear fractions from Arabidopsis, in which the cytosolic marker could not be detected in the nuclear fraction and the nuclear marker histone H3 could not be detected in the cytosolic fraction (Fig. 4C, D), showing the purity of the cytosolic and nuclear fraction. Using the antibody specific for SOAR1 (Supplementary Fig. S3 at JXB online), we immunodetected the SOAR1 protein in both the purified cytosolic and nuclear fractions (Fig. 4C, D), which consistently confirmed that SOAR1 is a cytosol and nucleus dual-localized protein.
It is noteworthy that, in the proteins sampled from the germinating seeds 24h after stratification, a form of SOAR1 protein was detected in the nuclear fraction with a higher molecular mass (74kDa) than the normal protein in the cytosolic fraction (68kDa) (Fig. 4C); and in the proteins sampled from the 2-week-old seedlings, both forms of SOAR1 protein were detected in the nuclear fraction with molecular masses of 68kDa and 74kDa, respectively (Fig. 4D). However, only a very weak signal of the 74kDa protein was detected that often could be scarcely seen in the total proteins (Fig. 4C, D) probably because of too low a concentration of the SOAR1 in the total protein extracts. These data indicate that the SOAR1 protein is subjected to a post-translational modification before or after it enters the nucleus, which may be associated with its function in the nucleus. This aspect remains an open question and needs further studies in the future.
Given that SOAR1 protein is present in the cytosolic space surrounding the mitochondria, it was further tested whether SOAR1 resides in the mitochondrion by using the above-mentioned SOAR1-GFP transgenic line OE1 (Fig. 1A). The nuclear localization of the SOAR1–GFP fusion protein was first verified in the root of the OE1 plants, as visualized by co-localization of the GFP fluorescence with the DAPI-stained nuclei (Fig. 4E). It was further shown that the localization pattern of the SOAR1 protein visualized by GFP fluorescence is distinct from the distribution profile of the mitochondria labelled by a mitochondrial marker (MitoTracker Red) (Fig. 4F). Taken together, these data demonstrate that SOAR1 localizes to the cytosol and nucleus, but not to chloroplasts or mitochondria.
Changes in SOAR1 expression alter expression of a subset of ABA-responsive genes
The expression levels of a subset of the ABA-responsive genes were tested in the soar1-2 mutant and the SOAR1 overexpressor OE1. These genes include ABF4/AREB2 (Choi et al., 2000; Uno et al., 2000), ABI1 (Leung et al., 1994; Meyer et al., 1994; Gosti et al., 1999), ABI2 (Leung et al., 1997), ABI3 (Giraudat et al., 1992), ABI4 (Finkelstein et al., 1998), ABI5 (Finkelstein and Lynch, 2000), DREB1A, DREB2A (Liu et al., 1998), MYB2 (Abe et al., 2003), PYR1/RCAR11, PYL2/RCAR14, PYL4/RCAR10, PYL7/RCAR2, PYL9/RCAR1 (Ma et al., 2009; Park et al., 2009), RD29A, RD29B (Yamaguchi-Shinozaki and Shinozaki, 1994), RAB18 (Lang and Palva, 1992), SnRK2.2, and SnRK2.3 (Fujii et al., 2007; Fujii and Zhu, 2009). The expression of the positive ABA signalling regulator-encoding genes (or positively ABA-responsive genes) ABI3, ABI4, ABI5, ABF4, DREB2A, PYR1, RAB18, RD29A, RD29B, SnRK2.2, and SnRK2.3 was significantly up-regulated in the soar1-2 mutant, while it was repressed in the OE1 line (Fig. 5A–D). However, the expression levels of the other positive ABA signalling regulator-encoding genes PYL2, PYL4, DREB1A, and MYB2 was not changed much in the soar1-2 mutant and the OE1 line compared with the wild-type plants, though some significant differences were still detected (Fig. 5B). The expression of another two genes encoding ABA receptors was altered differently: PYL9 was remarkably repressed, while PYL7 was significantly up-regulated in the soar1-2 mutant (Fig. 5B). It is noteworthy that the ABI3 and ABI5 genes, encoding two critical, positive regulators of ABA-responsive seed germination and post-germination growth, were substantially suppressed to a null level in the OE1 line while they were considerably up-regulated in the soar1-2 mutant (Fig. 5A, D), suggesting that these genes are potential, main targets of the SOAR1 protein. The expression of the negative ABA signalling regulator-encoding gene ABI1 was significantly repressed in the soar1-2 mutant, while ABI2 was remarkably up-regulated in the OE1 line (Fig. 5A, C).
Fig. 5.
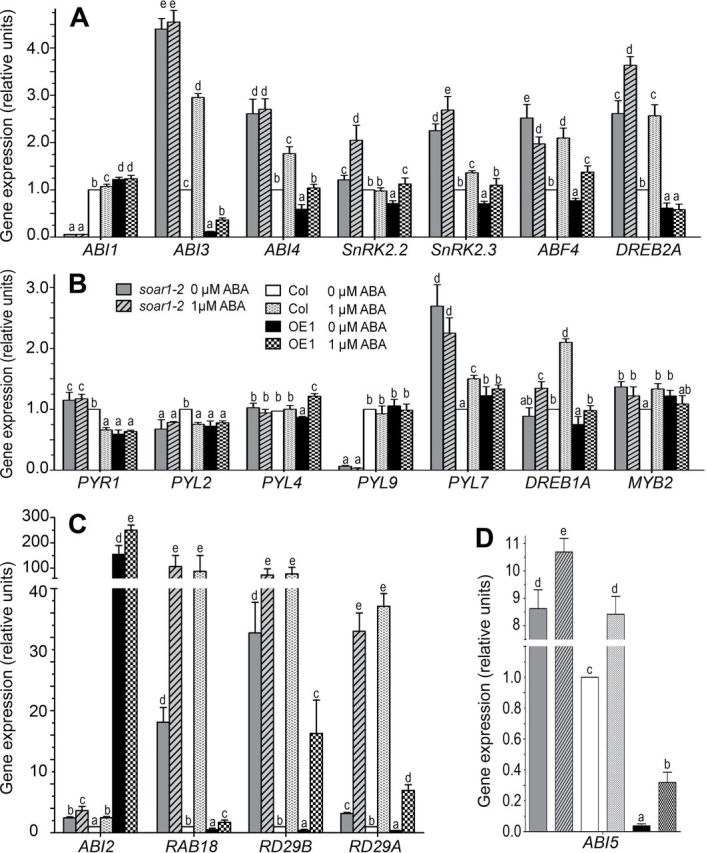
Down- or up-regulation of SOAR1 alters expression of a subset of genes involved in ABA signalling. The RNA sample was extracted from the germinating seeds 24h after a 3 d stratification, and the gene expression levels were analysed by quantitative real-time PCR (A–D). The gene expression levels were relative units normalized relative to the value from the sample of the wild-type Col plants (as 1). Each column is the same in all panels, and a key is provided in B. Each value is the mean ±SE of three biological determinations, and different letters indicate significant differences at P<0.05 (Duncan’s multiple range test) when comparing the expression levels for the same gene among different genotypes treated with the ABA-free (0 μM) or 1 μM ABA-containing solution.
The exogenous ABA treatment enhanced the expression levels of the ABA-responsive genes including ABI1, ABI2, ABI3, ABI4, ABF4, DREB1A, DREB2A, MYB2, PYL7, RAB18, RD29A, RD29B, and SnRK2.3 in the wild-type Col plants, whereas such ABA responsiveness of gene expression was significantly altered in the soar1-2 mutant and OE1 line (Fig. 5A–C). It is noteworthy that, with the ABA treatment, the expression levels of ABI3 and ABI5 still remained lower in the OE1 line (Fig. 5A, D), and the levels of most of the positive ABA signalling regulator-encoding genes such as ABI3, ABI4, ABI5, ABF4, DREB2A, PYL7, SnRK2.2, and SnRK2.3 remained higher in the soar1-2 mutant than in wild-type plants (Fig. 5A–D). The low expression of ABI1 was not significantly changed by ABA treatment in the soar1-2 mutant, and an even higher level of ABI2 was observed in the ABA-treated OE1 line (Fig. 5A, C).
Overall, these gene expression data are globally consistent with ABA-insensitive phenotypes of the SOAR1 overexpression lines and ABA-hypersensitive phenotypes of the soar1-2 and soar1-3 mutants. In particular, the remarkably high level of ABI2, together with low levels of ABI3 and ABI5 in the OE1 line, may be linked directly to the strong ABA-insensitive phenotypes of the SOAR1 overexpression lines, and the significantly high levels of ABI3, ABI5, ABF4, PYR7, and SnRK2.3 and the low level of ABI1 may be associated directly with the ABA-hypersensitive phenotypes of the soar1-2 and soar1-3 mutants (Figs 1, 2; Supplementary Figs S4–S7 at JXB online). Additionally, the expression profile of the ABA-responsive genes in the soar1-2 mutant and OE1 line suggests the presence of a complicated feed-forward and feed-back mechanism that may balance positive and negative regulation of gene expression to optimize ABA signalling.
Loss of function of ABI5 suppresses ABA-hypersensitive phenotypes of the soar1-2 T-DNA insertion mutant
Given that the ABI5 transcription factor is a key player regulating seed germination and post-germination growth in response to ABA (Finkelstein and Lynch, 2000; Lopez-Molina et al., 2001), and the ABI5 gene expression is significantly up-regulated in the soar1-2 mutant and nearly knocked out in the OE1 line (Fig. 5C), it was tested whether ABI5 functions downstream of SOAR1 as a potential target of this PPR protein. The abi5-1 soar1-2 double mutant was generated, in which abi5-1 is a knockout allele of the ABI5 gene and soar1-2 a knockdown allele of the SOAR1 gene. The abi5-1 soar1-2 double mutant showed ABA-insensitive phenotypes in ABA-induced seed germination inhibition and post-germination growth arrest, which are similar to the abi5-1 mutant (Fig. 3F–J). These findings suggest that SOAR1 may function upstream of ABI5 in the ABA signalling pathway.
Discussion
SOAR1 is a critical, negative, regulator acting downstream of ABAR and probably upstream of ABI5 in ABA signalling
It was shown that down-regulation of SOAR1 strongly increases, and up-regulation of SOAR1 almost completely impairs, ABA sensitivity in seed germination and post-germination growth. The intensity of the ABA overly-sensitive phenotypes of the two mutants was similar to, or stronger than, that of the well-characterized abi1 abi2 double-knockout mutant (Fig. 1; Supplementary Fig. S4 at JXB online). It could be expected that a complete loss of SOAR1 function would lead to even stronger ABA-hypersensitive phenotypes or dormant seeds; however, the null allele of the SOAR1 gene is likely to be lethal. The intensity of ABA-insensitive phenotypes of the SOAR1-overexpressing lines was much stronger than that of the abi1-1 dominant mutant, abi4 and abi5 loss-of-function mutants, and a strong ABI2-overexpressing line ABI2-OE (Figs 1, 2; Supplementary Figs S5, S6). In particular, it is surprising to note that the seeds of the SOAR1 overexpressors germinated and their post-germination seedlings continued to grow in the medium containing >200 μM (±)ABA (Fig. 2; Supplementary Figs S5, S6), and the 48-hour-old germinating seeds of the SOAR1 overexpressors even grew in the medium containing >500 μM (±)ABA (Fig. 2). Previous studies reported that the seeds of the srk2dei mutant, a triple-knockout mutant of three SnRK2 members SnRK2.2, SnRK2.3, and SnRK2.6, germinated and continued to grow in the presence of 50 μM or 100 μM exogenous ABA, which is believed to impair the ABA response completely (Fujii and Zhu, 2009; Nakashima et al., 2009; Umezawa et al., 2009). In this regard, the intensity of ABA insensitivity of the SOAR1 overexpression lines in this study is comparable with that of the triple loss-of-function mutant of the SnRK2 members (Fig. 2; Supplementary Figs S5, S6). However, the seeds of the SOAR1 overexpression lines are not viviparous despite their strong ABA insensitivity, suggesting that there may be a sophisticated mechanism to balance and optimize the function of SOAR1 under basal growth conditions.
All the findings reveal that SOAR1 is a critical, negative, regulator of ABA signalling, which regulates key processes of cell signalling in response to ABA. The ABA hypersensitivity of the ABAR overexpressors was suppressed by up-regulation of SOAR1 (Figs 1, 5), showing that SOAR1 functions in ABA signalling downstream of ABAR. Loss of function of ABI5 suppressed ABA-hypersensitive phenotypes of a SOAR1 knockdown mutant (soar1-2), suggesting that SOAR1 may function upstream of ABI5. These findings suggest a possible ABAR–SOAR1–ABI5-linked signalling cascade in the ABA signalling pathway, though it still remains unknown whether a direct interconnection exists between ABAR and SOAR1 or between SOAR1 and ABI5.
How does SOAR1 work in ABA signalling?
Currently, it remains largely unknown whether and how the PPR proteins regulate nuclear gene expression. In the two identified Arabidopsis nucleus-localized PPR proteins, GRP23 interacts physically with RNA polymerase II, suggesting its potential function as a transcription regulator (Ding et al., 2006); PNM1, dual localized to both the nucleus and mitochondrion, physically interacts with the nucleosome assembly protein NAP1 and the transcription factor TCP8, suggesting its potential roles in the coordination of mitochondrial and nuclear gene expression (Hammani et al., 2011). However, whether and how these nuclear PPR proteins regulate nuclear gene expression, and what their downstream, nuclear target genes are, have not been reported (Ding et al., 2006; Hammani et al., 2011).
In the present experiments, it was observed that down-regulation of the SOAR1 gene enhanced, but up-regulation of the SOAR1 gene repressed, the expression levels of the ABA-responsive genes ABI3, ABI4, ABI5, ABF4, DREB2A, PYR1, RAB18, RD29A, RD29B, SnRK2.2, and SnRK2.3. It is particularly noteworthy that the expression levels of the ABI3, ABI5, RAB18, and RD29B genes were markedly increased, while the level of ABI1 was almost completely suppressed by down-regulation of the SOAR1 gene, and the ABI3 and ABI5 genes were almost completely suppressed by overexpression of the SOAR1 gene (Fig. 5). It is also surprising to observe that the level of ABI2 expression was increased by >100-fold with up-regulation of the SOAR1 gene (Fig. 5). The marked increase of the ABI2 mRNA in the SOAR1 overexpression lines is likely to be caused by a decrease of some repressive factors of ABI2 to which is SOAR1 targeted, protecting the ABI2 mRNA from degradation. An ABA receptor member-encoding gene PYL9 was repressed, while the gene of another member PYL7 was up-regulated in the soar1-2 mutant (Fig. 5), suggesting a SOAR1-related balance mechanism that may function in RNA processing to maintain homeostasis of these family proteins. These gene expression data strongly suggest that these genes are most probably potential, direct or indirect targets of the SOAR1 protein, and explain, at least partly, the strong ABA-related phenotypes of the soar1 mutants and SOAR1 overexpressors. The gene expression profile under treatment with exogenous ABA is consistent with this conclusion. Genetic evidence that ABI5 may function downstream of SOAR1 (Fig. 3) strongly supports that ABI5 mRNA may be a target of the SOAR1 protein, which may function as a SOAR1–ABI5 directly coupled signalling module in the ABA signalling pathway.
It remains largely unclear whether SOAR1 participates in the PYR/PYL/RCAR-mediated ABA signalling pathway, a well-characterized core ABA signalling pathway (Fujii et al., 2009; Ma et al., 2009; Park et al., 2009; Santiago et al., 2009; Cutler et al., 2010). However, the expression data of the ABA-responsive genes in the soar1-2 mutant and SOAR1 overexpressor OE1 (Fig. 5) showed that the alteration in SOAR1 expression significantly changes the expression of a subset of genes, of which the encoded proteins have been identified to be directly involved in the PYR/PYL/RCAR-mediated ABA signalling, including the PYR/PYL/RCAR family members PYR1/RCAR11, PYL7/RCAR2, and PYL9/RCAR1, and some key signalling components ABI1, ABI2, SnRK2.2, SnRK2.3, ABI5, and ABF4 (Fig. 5). If the ABI5 mRNA is a direct target of SOAR1, ABI5 may be a common target of SOAR1 and SnRK2.2/3/6 in the ABA signalling pathway where the SnRK2 members regulate ABI5 by a phosphorylation process, a post-translation modification (Fujii et al., 2007; Fujii and Zhu, 2009), while SOAR1 may regulate ABI5 by post-trancriptional RNA processing. Thus, they may cooperate to regulate ABA signalling. Additionally, the gene expression data suggest that SOAR1 may regulate RNA processing of other key signalling component-encoding genes including SnRK2.2 and SnRK2.3. All the gene expression data support the idea that SOAR1 may also be involved in the PYR/PYL/RCAR-mediated ABA signalling which may possibly interact with ABAR/CHLH-mediated signalling through SOAR1. However, whether and how PYR/PYL/RCAR function as ABA receptors to regulate SOAR1, and how PYR/PYL/RCAR-mediated signalling interacts with ABAR/CHLH-mediated signalling through SOAR1, need further studies. Exploration of the detailed mechanisms of the cytosol–nuclear dual-localized SOAR1 protein that functions in the nuclear events as a critical component of ABA signalling, such as the nuclear mechanism by which SOAR1 regulates ABI5 mRNA processing, will be of particular importance in the future to understand the functional mechanism of PPR proteins and the highly complicated ABA signalling network.
Supplementary data
Supplementary data are available at JXB online.
Figure S1. Diagrams of the three T-DNA insertion mutants in the SOAR1 gene (At5g11310).
Figure S2. SOAR1 is a member of the pentatricopeptide repeat protein family.
Figure S3. Test of the specificity of the anti-SOAR1 serum.
Figure S4. The soar1-2 and soar1-3 mutants are hypersensitive to ABA in seed germination and early seedling growth.
Figure S5. The ABA-insensitive phenotypes in early seedling growth of 12 SOAR1 overexpression lines.
Figure S6. Phenotypes of the soar1-1D, soar1-2, and soar1-3 mutants, and SOAR1 and ABI overexpression lines in response to ABA.
Figure S7. Transgenic expression of SOAR1 rescues the ABA-hypersensitive phenotypes of the soar1-2 and soar1-3 mutant.
Figure S8. ABA concentrations in the germinating seeds of different genotypes.
Figure S9. SOAR1 is expressed in different organs/tissues, with the highest expression level in seeds.
Figure S10. Prediction of the subcellular localization of SOAR1 protein.
Table S1. Primers used in this study.
Acknowledgements
We thank Drs Yan Guo and Zhong-Jun Zhang (China Agricultural University, Beijing, China), and Drs Dong Liu and Li Yu (Tsinghua University, Beijing, China) for help with materials and equipment. This research was supported by the National Key Basic Research Program of China (2012CB114300-002), the National Natural Science Foundation of China (grant nos 31200213 and 31170268), and the Ministry of Agriculture of China (grant no. 2013ZX08009003).
References
- Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. 2003. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcription activators in abscisic acid signaling. The Plant Cell 15, 63–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adie BAT, Pérez-Pérez J, Pérez-Pérez MM, Godoy M, Sánchez-Serrano JJ, Schmelz EA, Solano R. 2007. ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis . The Plant Cell 19, 1665–1681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aubourg S, Boudet N, Kreis M, Lecharny A. 2000. In Arabidopsis thaliana, 1% of the genome codes for a novel protein family unique to plants. Plant Molecular Biology 42, 603–613. [DOI] [PubMed] [Google Scholar]
- Chi W, Ma J, Zhang D, Guo J, Chen F, Lu C, Zhang L. 2008. The pentratricopeptide repeat protein DELAYED GREENING1 is involved in the regulation of early chloroplast development and chloroplast gene expression in Arabidopsis. Plant Physiology 147, 573–584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi HJ, Hong J, Ha JO, Kang JY, Kim SY. 2000. ABFs, a family of ABA-responsive element binding factors. Journal of Biological Chemistry 275, 1723–1730. [DOI] [PubMed] [Google Scholar]
- Clough SJ, Bent AF. 1998. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana . The Plant Journal 16, 735–743. [DOI] [PubMed] [Google Scholar]
- Cushing DA, Forsthoefel NR, Gestaut DR, Vernon DM. 2005. Arabidopsis emb175 and other ppr knockout mutants reveal essential roles for pentatricopeptide repeat (PPR) proteins in plant embryogenesis. Planta 221, 424–436. [DOI] [PubMed] [Google Scholar]
- Cutler SR, Rodriguez PL, Finkelsteion RR, Abrams SR. 2010. Abscisic acid: emergence of a core signaling network. Annual Review of Plant Biology 61, 651–679. [DOI] [PubMed] [Google Scholar]
- Ding YH, Liu NY, Tang ZS, Liu J, Yang WC. 2006. Arabidopsis GLUTAMINE-RICH PROTEIN23 is essential for early embryogenesis and encodes a novel nuclear PPR motif protein that interacts with RNA polymerase II subunit III. The Plant Cell 18, 815–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du SY, Zhang XF, Lu Z, Xin Q, Wu Z, Jiang T, Lu Y, Wang XF, Zhang DP. 2012. Roles of the different components of magnesium chelatase in abscisic acid signal transduction. Plant Molecular Biology 80, 519–537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fairchild CD, Schumaker MA, Quail PH. 2000. HFR1 encodes an atypical bHLH protein that acts in phytochrome A signal transduction. Genes and Development 14, 2377–2391. [PMC free article] [PubMed] [Google Scholar]
- Finkelstein RR, Gampala S, Rock C. 2002. Abscisic acid signaling in seeds and seedlings. The Plant Cell 14 (suppl.), S15–S45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finkelstein RR, Lynch TJ. 2000. The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zipper transcription factor. The Plant Cell 12, 599–609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finkelstein RR, Wang ML, Lynch TJ, Rao S, Goodman HM. 1998. The Arabidopsis abscisic acid response locus ABI4 encodes an APETALA2 domain protein. The Plant Cell 10, 1043–1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujii H, Chinnusamy V, Rodrigues A, Rubio S, Antoni R, Park SY, Cutler SR, Sheen J, Rodriguez PL, Zhu JK. 2009. In vitro reconstitution of an abscisic acid signaling pathway. Nature 462, 660–664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujii H, Verslues PE, Zhu JK. 2007. Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis. The Plant Cell 19, 485–494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujii H, Zhu JK. 2009. Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction, and stress. Proceedings of the National Academy of Sciences, USA 106, 8380–8385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujii S, Small I. 2011. The evolution of RNA editing and pentatricopeptide repeat genes. New Phytologist 191, 37–47. [DOI] [PubMed] [Google Scholar]
- Gao Y, Zeng Q, Guo J, Cheng J, Ellis BE, Chen JG. 2007. Genetic characterization reveals no role for the reported ABA receptor, GCR2, in ABA control of seed germination and early seedling development in Arabidopsis. The Plant Journal 52, 1001–1013. [DOI] [PubMed] [Google Scholar]
- Giraudat J, Hauge BM, Valon C, Smalle J, Parcy F, Goodman HM. 1992. Isolation of the Arabidopsis ABI3 gene by positional cloning. The Plant Cell 4, 1251–1261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gosti F, Beaudoin N, Serizet C, Webb AAR, Vartanian N, Giraudat J. 1999. ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signaling. The Plant Cell 11, 1897–1909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo J, Zeng Q, Emami M, Ellis BE, Chen JG. 2008. The GCR2 gene family is not required for ABA control of seed germination and early seedling development in Arabidopsis. PLoS One 3, e2982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutiérrez-Marcos JF, Prà MD, Giulini A, et al. 2007. empty pericarp4 encodes a mitochondrion-targeted pentatricopeptide repeat protein necessary for seed development and plant growth in maize. The Plant Cell 19, 196–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammani K, Gobert A, Hleibieh K, Choulier L, Small I, Giegé P. 2011. An Arabidopsis dual-localized pentatricopeptide repeat protein interacts with nuclear proteins involved in gene expression regulation. The Plant Cell 23, 730–740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu J, Wang K, Huang W, et al. 2012. The rice pentatricopeptide repeat protein RF5 restores fertility in Hong-Lian cytoplasmic male-sterile lines via a complex with the glycine-rich protein GRP162. The Plant Cell 24, 109–122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang IC, Yang JY, Seo HS, Chua NH. 2005. HFR1 is targeted by COP1 E3 ligase for post-translational proteolysis during phytochrome A signaling. Genes and Development 19, 593–602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jefferson RA, Kavanagh TA, Bevan MW. 1987. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO Journal 20, 3901–3907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia HF, Chai YM, Li CL, Lu D, Luo JJ, Qin L, Shen YY. 2011. b Abscisic acid plays an important role in the regulation of strawberry fruit ripening. Plant Physiology 157, 188–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia HF, Chai YM, Li CL, Qin L, Shen YY. 2011. a Cloning and characterization of the H subunit of a magnesium chelatase gene (PpCHLH) in peach. Journal of Plant Growth Regulation 30, 445–455. [Google Scholar]
- Johnston CA, Temple BR, Chen JG, Gao Y, Moriyama EN, Jones AM, Siderovski DP, Willard FS. 2007. Comment on ‘A G protein-coupled receptor is a plasma membrane receptor for the plant hormone abscisic acid’. Science 318, 914. [DOI] [PubMed] [Google Scholar]
- Koussevitzky S, Nott A, Mockler TC, Hong F, Sachetto-Martins G, Surpin M, Lim J, Mittler R, Chory J. 2007. Signals from chloroplasts converge to regulate nuclear gene expression. Science 316, 715–719. [PubMed] [Google Scholar]
- Laluk K, AbuQamar S, Mengiste T. 2011. The Arabidopsis mitochondria-localized pentatricopeptide repeat protein PGN functions in defense against necrotrophic fungi and abiotic stress tolerance. Plant Physiology 156, 2053–2068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang V, Palva ET. 1992. The expression of a rab-related gene, rab18, is induced by abscisic acid during the cold acclimation process of Arabidopsis thaliana (L.) Heynh. Plant Molecular Biology 20, 951–962. [DOI] [PubMed] [Google Scholar]
- Lee DK, Suh D, Edenberg HJ, Hur MW. 2002. POZ domain transcription factor, FBI-1, represses transcription of ADH5/FDH by interacting with the zinc finger and interfering with DNA binding activity of Sp1. Journal of Biological Chemistry 277, 26761–26768. [DOI] [PubMed] [Google Scholar]
- Legnaioli T, Cuevas J, Mas P. 2009. TOC1 functions as a molecular switch connecting the circadian clock with plant responses to drought. EMBO Journal 28, 3745–3757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung J, Bouvier-Durand M, Morris PC, Guerrier D, Chefdor F, Giraudat J. 1994. Arabidopsis ABA response gene ABI1: features of a calcium-modulated protein phosphatase. Science 264, 1448–1452. [DOI] [PubMed] [Google Scholar]
- Leung J, Merlot S, Giraudat J. 1997. The Arabidopsis ABSCISIC ACD-INSENSITIVE2 (ABI2) and ABI1 encode homologous protein phosphatase 2C involved in abscisic acid signal transduction. The Plant Cell 9, 759–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K, Shinozaki K. 1998. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA-binding domain separate two cellular signal transduction pathways in drought- and low temperature-responsive gene expression in Arabidopsis. The Plant Cell 10, 1391–1406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Yue Y, Li B, Nie Y, Li W, Wu WH, Ma L. 2007. a A G protein-coupled receptor is a plasma membrane receptor for the plant hormone abscisic acid. Science 315, 1712–1716. [DOI] [PubMed] [Google Scholar]
- Liu X, Yue Y, Li W, Ma LG. 2007. b Response to comment on ‘A G protein coupled receptor is a plasma membrane receptor for the plant hormone abscisic acid’. Science 318, 914. [DOI] [PubMed] [Google Scholar]
- Liu Y, He J, Chen Z, Ren X, Hong X, Gong Z. 2010. ABA overly-sensitive 5 (ABO5), encoding a pentatricopeptide repeat protein required for cis-splicing of mitochondrial nad2 intron 3, is involved in the abscisic acid response in Arabidopsis. The Plant Journal 63, 749–765. [DOI] [PubMed] [Google Scholar]
- Liu YG, Mitsukawa N, Oosumi T, Whittier RF. 1995. Efficient isolation and mapping of Arabidopsis thaliana T-DNA insert junctions by thermal asymmetric interlaced PCR. The Plant Journal 8, 457–463. [DOI] [PubMed] [Google Scholar]
- Liu ZQ, Yan L, Wu Z, Mei C, Lu K, Yu YT, Liang S, Zhang XF, Wang XF, Zhang DP. 2012. Cooperation of three WRKY-domain transcription factors WRKY18, WRKY40, and WRKY60 in repressing two ABA responsive genes ABI4 and ABI5 in Arabidopsis . Journal of Experimental Botany 63, 6371–6392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Molina L, Mongrand S, Chua NH. 2001. A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis . Proceedings of the National Academy of Sciences, USA 98, 4782–4787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lurin C, Andrés C, Auberge S, et al. 2004. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. The Plant Cell 16, 2089–2103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E. 2009. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 324, 1064–1068. [DOI] [PubMed] [Google Scholar]
- Manthey GM, McEwen JE. 1995. The product of the nuclear gene PET309 is required for translation of mature mRNA and stability or production of intron-containing RNAs derived from the mitochondrial COX1 locus of Saccharomyces cerevisiae . EMBO Journal 14, 4031–4043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meierhoff K, Felder S, Nakamura T, Bechtold N, Schuster G. 2003. HCF152, an Arabidopsis RNA binding pentatricopeptide repeat protein involved in the processing of chloroplast psbB-psbT-psbH-petB-petD RNAs. The Plant Cell 15, 1480–1495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer K, Leube MP, Grill E. 1994. A protein phosphatase 2C involved in ABA signal transduction in Arabidopsis thaliana . Science 264, 1452–1455. [DOI] [PubMed] [Google Scholar]
- Müller AH, Hansson M. 2009. The barley magnesium chelatase 150-kD subunit is not an abscisic acid receptor. Plant Physiology 150, 157–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 15, 473–497. [Google Scholar]
- Murayama M, Hayashi S, Nishimura N, Ishide M, Kobayashi K, Yagi Y, Asami T, Nakamura T, Shinozaki K, Hirayama T. 2012. Isolation of Arabidopsis ahg11, a weak ABA hypersensitive mutant defective in nad4 RNA editing. Journal of Experimental Botany 63, 5301–5310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura T, Yagi Y, Kobayashi K. 2012. Mechanistic insight into pentatricopeptide repeat proteins as sequence-specific RNA-binding proteins for organellar RNAs in plants. Plant and Cell Physiology 53, 1171–1179. [DOI] [PubMed] [Google Scholar]
- Nakashima K, Fujita Y, Kanamori N, et al. 2009. Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant and Cell Physiology 50, 1345–1363. [DOI] [PubMed] [Google Scholar]
- Oguchi T, Sage-Ono K, Kamada H, Ono M. 2004. Genomic structure of a novel Arabidopsis clock-controlled gene, AtC401, which encodes a pentatricopeptide repeat protein. Gene 330, 29–37. [DOI] [PubMed] [Google Scholar]
- Pandey S, Nelson DC, Assmann SM. 2009. Two novel GPCR-type G proteins are abscisic acid receptors in Arabidopsis . Cell 136, 136–148. [DOI] [PubMed] [Google Scholar]
- Park SY, Fung P, Nishimura N, et al. 2009. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 324, 1068–1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivals E, Bruyère C, Toffano-Nioche C, Lecharny A. 2006. Formation of the Arabidopsis pentatricopeptide repeat family. Plant Physiology 141, 825–839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santiago J, Rodrigues A, Saez A, Rubio S, Antoni R, Dupeux F, Park SY, Márquez JA, Cutler SR, Rodriguez PL. 2009. Modulation of drought resistance by the abscisic acid receptor PYL5 through inhibition of clade A PP2Cs. The Plant Journal 16, 575–588. [DOI] [PubMed] [Google Scholar]
- Schmitz-Linneweber C, Small I. 2008. Pentatricopeptide repeat proteins: a socket set for organelle gene expression. Trends in Plant Science 13, 663–670. [DOI] [PubMed] [Google Scholar]
- Shaner NC, Campbell RE, Steinbach PA, Giepmans BN, Palmer AE, Tsien RY. 2004. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nature Biotechnology 22, 1567–1572. [DOI] [PubMed] [Google Scholar]
- Shang Y, Yan L, Liu ZQ, et al. 2010. The Mg-chelatase H subunit of Arabidopsis antagonizes a group of WRKY transcription repressors to relieve ABA-responsive genes of inhibition. The Plant Cell 22, 1909–1935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen YY, Wang XF, Wu FQ, et al. 2006. The Mg-chelatase H subunit is an abscisic acid receptor. Nature 443, 823–826. [DOI] [PubMed] [Google Scholar]
- Small ID, Peeters N. 2000. The PPR motif—a TPR-related motif prevalent in plant organellar proteins. Trends in Biochemical Sciences 25, 45–47. [DOI] [PubMed] [Google Scholar]
- Sun HL, Wang XJ, Ding WH, Zhu SY, Zhao R, Zhang YX, Xin Q, Wang XF, Zhang DP. 2011. Identification of an important site for function of the type 2C protein phosphatase ABI2 in abscisic acid signalling in Arabidopsis . Journal of Experimental Botany 62, 5713–5725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuzuki T, Takahashi K, Inoue S, Okigaki Y, Tomiyama M, Hossain MA, Shimazaki K, Murata Y, Kinoshita T. 2011. Mg-chelatase H subunit affects ABA signaling in stomatal guard cells, but is not an ABA receptor in Arabidopsis thaliana . Journal of Plant Research 124, 527–538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuzuki T, Takahashi K, Tomiyama M, Inoue S, Kinoshita T. 2013. Overexpression of the Mg-chelatase H subunit in guard cells confers drought tolerance via promotion of stomatal closure in Arabidopsis thaliana . Frontiers in Plant Science 4, 440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tzafrir I, Pena-Muralla R, Dickerman A, et al. 2004. Identification of genes required for embryo development in Arabidopsis . Plant Physiology 135, 1206–1220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umezawa T, Sugiyama N, Mizoguchi M, Hayashi S, Myouga F, Yamaguchi-Shinozaki K, Ishihama Y, Hirayama T, Shinozaki K. 2009. Type 2C protein phosphatases directly regulate abscisic acid-activated protein kinases in Arabidopsis. Proceedings of the National Academy of Sciences, USA 106, 17588–17593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K. 2000. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proceedings of the National Academy of Sciences, USA 97, 11632–11637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang XF, Xin Q, Shen YY, Zhang DP. 2011. Approaches to the identification of ABAR as an abscisic acid receptor. Methods in Molecular Biology 773, 83–97. [DOI] [PubMed] [Google Scholar]
- Wang Z, Zou Y, Li X, et al. 2006. Cytoplasmic male sterility of rice with boro II cytoplasm is caused by a cytotoxic peptide and is restored by two related PPR motif genes via distinct modes of mRNA silencing. The Plant Cell 18, 676–687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams PM, Barkan A. 2003. A chloroplast-localized PPR protein required for plastid ribosome accumulation. The Plant Journal 36, 675–686. [DOI] [PubMed] [Google Scholar]
- Wu FQ, Xin Q, Cao Z, et al. 2009. The magnesium-chelatase H subunit binds abscisic acid and functions in abscisic acid signaling: new evidence in Arabidopsis. Plant Physiology 150, 1940–1954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi-Shinozaki K, Shinozaki K. 1994. A nove1 cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. The Plant Cell 6, 251–264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan L, Liu ZQ, Xu YH, Lu K, Wang XF, Zhang DP. 2013. Auto- and cross-repression of three Arabidopsis WRKY transcription factors WRKY18, WRKY40, and WRKY60 negatively involved in ABA signaling. Journal of Plant Growth Regulation 32, 399–416. [Google Scholar]
- Yoo SD, Cho YH, Sheen J. 2007. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nature Protocols 2, 1565–1572. [DOI] [PubMed] [Google Scholar]
- Yuan H, Liu D. 2012. Functional disruption of the pentatricopeptide protein SLG1 affects mitochondrial RNA editing, plant development, and response to abiotic stresses in Arabidopsis . The Plant Journal 70, 432–444. [DOI] [PubMed] [Google Scholar]
- Zhang XF, Jiang T, Wu Z, Du SY, Yu YT, Jiang SC, Lu K, Feng XJ, Wang XF, Zhang DP. 2013. Cochaperonin CPN20 negatively regulates abscisic acid signaling in Arabidopsis . Plant Molecular Biology 83, 205–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang XF, Jiang T, Yu YT, et al. 2014. Arabidopsis co-chaperonin CPN20 antagonizes Mg-chelatase H subunit to derepress ABA-responsive WRKY40 transcription repressor. Science China Life Sciences 57, 11–21. [DOI] [PubMed] [Google Scholar]
- Zhao R, Sun HL, Mei C, Wang XJ, Yan L, Liu R, Zhang XF, Wang XF, Zhang DP. 2011. The Arabidopsis Ca2+-dependent protein kinase CPK12 negatively regulates abscisic acid signaling in seed germination and post-germination growth. New Phytologist 192, 61–73. [DOI] [PubMed] [Google Scholar]
- Zsigmond L, Rigó G, Szarka A, Székely G, Ötvös K, Darula Z, Medzihradszky KF, Koncz C, Koncz Z, Szabados L. 2008. Arabidopsis PPR40 connects abiotic stress responses to mitochondrial electron transport. Plant Physiology 146, 1721–1737. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.