

Abstract
Two non-pigmented, motile, Gram-negative marine bacteria designated R9SW1T and A3d10T were isolated from sea water samples collected from Chazhma Bay, Gulf of Peter the Great, Sea of Japan, Pacific Ocean, Russia and St. Kilda Beach, Port Phillip Bay, the Tasman Sea, Pacific Ocean, respectively. Both organisms were found to grow between 4°C and 40°C, between pH 6 to 9, and are moderately halophilic, tolerating up to 20% (w/v) NaCl. Both strains were found to be able to degrade Tween 40 and 80, but only strain R9SW1T was found to be able to degrade starch. The major fatty acids were characteristic for the genus Marinobacter including C16:0, C16:1 ω7c, C18:1 ω9c and C18:1 ω7c. The G+C content of the DNA for strains R9SW1T and A3d10T were determined to be 57.1 mol% and 57.6 mol%, respectively. The two new strains share 97.6% of their 16S rRNA gene sequences, with 82.3% similarity in the average nucleotide identity (ANI), 19.8% similarity in the in silico genome-to-genome distance (GGD), 68.1% similarity in the average amino acid identity (AAI) of all conserved protein-coding genes, and 31 of the Karlin's genomic signature dissimilarity. A phylogenetic analysis showed that R9SW1T clusters with M. algicola DG893T sharing 99.40%, and A3d10T clusters with M. sediminum R65T sharing 99.53% of 16S rRNA gene sequence similarities. The results of the genomic and polyphasic taxonomic study, including genomic, genetic, phenotypic, chemotaxonomic and phylogenetic analyses based on the 16S rRNA, gyrB and rpoD gene sequence similarities, the analysis of the protein profiles generated using MALDI-TOF mass spectrometry, and DNA-DNA relatedness data, indicated that strains R9SW1T and A3d10T represent two novel species of the genus Marinobacter. The names Marinobacter salarius sp. nov., with the type strain R9SW1T ( = LMG 27497T = JCM 19399T = CIP 110588T = KMM 7502T) and Marinobacter similis sp. nov., with the type strain A3d10T ( = JCM 19398T = CIP 110589T = KMM 7501T), are proposed.
Introduction
The genus Marinobacter (family Alteromonadaceae, order Alteromonadales, class Gammaproteobacteria) was created by Gauthier et al. for a hydrocarbon degrading bacterium. At the time of writing, the genus comprises 33 validly described species, http://www.bacterio.net/marinobacter.html [1], which accommodates Gram-negative, chemoheterotrophic and halophilic, rod-shaped bacteria [2], [3]. The important role played by Marinobacter spp. in metabolizing hydrocarbons has long been noted, with M. hydrocarbonoclasticus [2], M. aquaeolei [4], [5], M. maritimus [6], and M. algicola [7] having been characterized as being able to utilise aromatic and aliphatic hydrocarbons as their sole carbon and energy sources. It was also shown that bacteria of the genus Marinobacter are one of the dominant bacterial community groups constantly recovered from hydrocarbon polluted sites [8]–[10]. For example, it was recently demonstrated that M. vinifirmus was able to effectively degrade toluene, benzene, ethylbenzene, and p-xylene [11].
The objectives of this study were to classify two newly isolated marine bacteria; strain R9SW1T, which was derived from a water sample collected from Chazhma Bay (Gulf of Peter the Great, Sea of Japan, Pacific Ocean) during taxonomic studies of microbial communities developed in sea water contaminated by radionuclides [12]; and strain A3d10T, which was isolated from Port Philip Bay (the Tasman Sea, Pacific Ocean) during the course of polymer biodegradation studies [13]. The comparative taxonomic investigations of these bacteria, together with their close relatives, revealed their distinct taxonomic standing. This suggests that strain R9SW1T and strain A3d10T represent two novel species of the genus Marinobacter.
Materials and Methods
Isolation procedures, bacterial strains, and growth conditions
Strain R9SW1T was isolated from a sea water sample collected from Chazhma Bay in the Sea of Japan, Pacific Ocean, in 2000. Water sample collection was within the research program funded by the Federal Agency for Science of the Ministry of Education and Science of the Russian Federation, grant 2–2.16 and by the Russian Foundation for Basic Research and grant ‘Molecular and Cell Biology’ from the Presidium of the Russian Academy of Sciences, grant 02-04-48211". The specific location of the studies (GPS coordinates) was 42°53′38″ N 132°22′02″ E. The permit issued by the Department of Marine Expeditions, Ministry of Education and Science of the Russian Federation. Strain A3d10T was isolated from a sea water sample collected one metre below the water surface in Port Philip Bay, the Tasman Sea, Pacific Ocean, in 2008. Sea water collected from St Kilda Beach which is a publicly accessible beach area in Melbourne, not part of any protected area of land or sea. Furthermore, the field studies did not involve endangered or protected species. The specific location of the studies (GPS coordinates) was 37°51′50″S 144°58′55″E. The sample handling and isolation procedures used were as previously described [12], [13]. Samples were plated on marine agar 2216 (BD, USA) and incubated aerobically at approximately 22–25°C for 5, 7 or 10 days. The isolation and purification procedure has been described elsewhere [14], [15]. Ten type strains of the Marinobacter species were obtained from various culture collections and used as the reference strains; M. lipolyticus CIP 107627T, M. gudaonensis CIP 109534T, M. adhaerens CIP 110141T, M. salsuginis CIP 109893T and M. flavimaris CIP 108615T were obtained from Collection de l’Institut Pasteur (CIP) culture collection, M. algicola LMG 23835T, M. guineae LMG 24048T and M. sediminum LMG 23833T were obtained from The Belgian Co-ordinated Collections of Micro-organisms (BCCM/LMG), M. goseongensis KCTC 12515T was obtained from Korean Collection for Type Cultures (KCTC) and M. xestospongiae JCM 17469T was obtained from RIKEN BRC-Japan Collection of Microorganisms (JCM). The type species of the genus, M. hydrocarbonoclasticus SP. 17T was kindly provided by Dr. Stan-Lotter. Strains were stored at −80°C in marine broth 2216 (BD, USA) that had been supplemented with 20% (v/v) glycerol.
16S rDNA, gyrB, rpoD sequencing and phylogenetic analysis
Genomic DNAs were isolated using a Wizard Genomic DNA Purification Kit (Promega, USA) according to the manufacturer's specifications. The 16S rRNA gene sequences for strains R9SW1T and A3d10T were extracted from the whole genome sequences [16] while gyrB and rpoD genes were amplified using primers (see Supporting Information, Table S1 in File S1) that have been previously described [17], [18]. The 16S rRNA gene sequences of validly described Marinobacter species were retrieved from GenBank and aligned using the CLUSTAL W program [19]. Evolutionary phylogenetic trees were constructed using the neighbour-joining (NJ) [20], maximum-likelihood (ML) [21] and maximum-parsimony (MP) [22] algorithms. Genetic distances were calculated using Kimura's two-parameter model [23] by using the MEGA 5 software [24]. The GenBank/EMBL/DDBJ accession numbers of 16S rRNA gene, gyrB, rpoD and whole genome sequences were presented as in Table 1.
Table 1. GenBank/EMBL/DDBJ accession numbers of 16S rDNA, gyrB, rpoD and whole genome sequences for strains R9SW1T, A3d10T and phylogenetically related type strains and type species of the genus Marinobacter.
| Species name | GenBank/EMBL/DDBJ accession numbers | |||
| 16S rDNA | gyrB | rpoD | whole genome | |
| Strain R9SW1T | KJ547705 | KF811464 | KF811478 | CP007152 |
| Strain A3d10T | KJ547704 | KF811465 | KF811471 | CP007151 |
| M. algicola LMG 23835T | AY258110* | KF811463 | KF811474 | - |
| M. sediminum LMG 23833T | AJ609270* | KF811466 | KF811477 | - |
| M. adhaerens CIP 110141T | AY241552* | KF811467 | KF811473 | NC_017506* |
| M. flavimaris CIP 108615T | AY517632* | KF811468 | KF811475 | - |
| M. salsuginis CIP 109893T | EF028328* | KF811469 | KF811476 | - |
| M. hydrocarbonoclasticus SP.17T | X67022* | KF811470 | KF811472 | NC_017067* |
*Accession numbers from previous publications.
MALDI-TOF MS analysis
The sample preparation and MALDI-TOF MS analysis was carried out according to the techniques described elsewhere [25]. Briefly, 5 µL of the cultures grown overnight were transferred into microcentrifuge tubes and subjected to ethanol and formic acid protein extraction. One µL aliquots of the supernatant were transferred onto the MALDI target plate and air dried at room temperature, followed by the addition of 1 µL of matrix solution, then air dried. Samples were then subjected to analysis using a Microflex MALDI-TOF mass spectrometer (Bruker Daltonik GmbH, Leipzig, Germany) equipped with a 60 Hz nitrogen laser. Spectra were recorded in the positive linear mode for the mass range of 2,000 to 20,000 Da at the maximum laser frequency. The raw spectra were then analysed using the MALDI Biotyper 3.0 software package (Bruker Daltonik GmbH, Bremen, Germany) under the default settings. Measurements were performed via the automatic mode, without any user intervention.
GC content and DNA-DNA hybridization
The GC content of strains R9SW1T and A3d10T was calculated on the basis of their whole genome sequences [16], [26], and these have been deposited at GenBank/EMBL/DDBJ under the accession number of CP007152 and CP007151, respectively. The DNA-DNA hybridizations between strain R9SW1T and M. algicola LMG 23835T, and strain A3d10T and M. sediminum LMG 23833T were performed by the Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ) identification service, where cells were initially disrupted using a Constant Systems TS 0.75 KW (IUL Instruments, Germany), followed by purification of the extracted DNA in the crude lysate form by chromatography on hydroxyapatite as described by Cashion et. al. (1977) [27]. DNA-DNA hybridization was carried out in duplicate using a 2× saline sodium citrate (SSC) buffer with 5% formamide as described by De Ley et al. [28], with consideration of the modifications described by Huss et. al. (1983) [29], using a model Cary 100 Bio UV/VIS-spectrophotometer equipped with a Peltier-thermostatted 6×6 multi-cell changer and a temperature controller with an in-situ temperature probe (Varian).
Genome comparison and genomic signatures analyses
Complete genome sequences for only two validly described species of Marinobacter, M. hydrocarbonoclasticus ATCC 49840T [30] and M. adhaerens HP15T [31], which have previously been assembled, were used in this study for genomic analysis. The fully sequenced and assembled genomes of both these species were retrieved from GenBank, and compared to those of R9SW1T and A3d10T. Genome comparison between strains R9SW1T, A3d10T, M. adhaerens HP15T and M. hydrocarbonoclasticus ATCC 49840T was carried out using reciprocal BLAST analysis, according to the method described by Goris et al. [32]. A map of the percentage identity between each of M. adhaerens HP15T, R9SW1T and A3d10T to the type species was generated using the BLAST Ring Image Generator (BRIG) software [33]. The in-silico genome-to-genome distance (GGD) between the four strains was also calculated using the genome-to-genome distance calculator 2.0 (GGDC) provided by DSMZ, http://ggdc.dsmz.de [34], [35]. The average amino acid identity (AAI) of all conserved protein-coding genes was calculated as previously described [36]. The conserved genes between a pair of genomes were determined by whole-genome pairwise sequence comparison using the BLAST algorithm release 2.2.5 [37] using a minimum cut-off of 40% identity and 70% of the length of the query gene. The difference in genome signature between two individual sequences is expressed in terms of the Karlin's genomic signature dissimilarity (δ*), which was calculated by dividing the genomic dinucleotide frequencies with the corresponding mononucleotide content using the equation described by Karlin et al. [38]. Phylogenomic relationship between the four strains were also elucidated using Mauve multiple alignment software (v2.3.1) [39] and ClonalFrame software v1.2 [40], with Alteromonas sp. DE [41] used as an outgroup.
Genotype to phenotype analyses of a few distinctive phenotypes were also carried using the whole genome sequences of strains R9SW1T, A3d10T, M. hydrocarbonoclasticus ATCC 49840T and M. adhaerens HP15T using the methods as previously described [42].
Physiological and biochemical analysis
Six reference type strains, along with strains R9SW1T and A3d10T, were used for the phenotypic and biochemical tests (Table 2). The cell morphology and motility were determined using scanning electron and light microscopies. Gram stain reaction, catalase (5% H2O2) and starch hydrolysis analyses were performed according to the method described by Smibert and Krieg (1994) [43]. Determination of the oxidase activity was performed using Bactident oxidase strips (Merck Millipore, Germany). The capacity of the strains to oxidize and to ferment D-glucose and lactose was carried out according to the method described by Smibert and Krieg (1994) [43], using a modified semi-solid medium containing: 9.4 g L−1 O/F medium (Oxoid, UK), 20 g L−1 Sea Salt (Sigma-Aldrich, USA) and 1% carbohydrate. The strains were incubated at 30°C and the results were obtained after 48 hours. The temperature and pH tolerance ranges were determined via marine agar growth tests subjected to different temperature (4, 10, 15, 20, 25, 30, 37, 40, 45 and 50°C) and pH (4, 6, 7, 8, 9 and 11, adjusting the pH with HCl and NaOH) conditions. The NaCl tolerance was determined using different concentrations of NaCl (0, 0.5, 1, 3, 6, 10, 15, 20 and 25%) in modified salinity agar (SA) containing: 5 g L−1 peptone, 1 g L−1 yeast extract, 0.1 g L−1 ferric citrate, 3.24 g L−1 magnesium sulphate (MgSO4), 0.55 g L−1 dipotassium phosphate (K2HPO4), 15 g L−1 agar, and the respective NaCl concentration, each at a pH of 7.6±0.2. Plates were incubated under optimal temperature conditions and the results were recorded daily for 7 days.
Table 2. Differential characteristics between strains R9SW1T, A3d10T, their close phylogenetic neighbors and type species of the genus Marinobacter.
| Characteristics | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 |
| Cell length (µm) | 1.9–3.2 | 1.6–2.5 | 1.3–2.1 | 1.8–2.5 | 2.0–4.0 | 1.7–2.4 | 1.5–3.0 | 2.5–3.5 | 1.2–1.8 | 1.6–2.0 | 2.0–2.5 | 1.4–4.0 | 2.0–3.0 |
| Cell width (µm) | 0.40–0.72 | 0.45–0.55 | 0.40–0.45 | 0.3–0.4 | 1.0 | 0.6–0.8 | 0.6–0.9 | 0.3–0.5 | 0.3–0.5 | 0.5–0.8 | 0.6–0.8 | 0.4 | 0.3–0.6 |
| Growth temperature (°C) | 4–40 | 5–40 | 4–40 | 4–42 | 10–45 | 4–45 | 4–45 | 15–40 | 10–45 | 10–37 | 15–42 | 4–42 | 10–45 |
| pH range | 6–9 | 5–10 | 6–9 | ND | 6.5–9.5 | 5.5–10.0 | >5.5 | 5.0–10.0 | 6.0–9.5 | 5.3–9.3 | 5.0–10.0 | 5.0–9.5 | 6–9.5 |
| Salinity range (%, w/v) | 0.5–20 | 1–12 | 0.5–20 | 0.5–18 | 1–20 | 0.5–20 | 1–20 | 1–15 | 0–15 | 1–25 | 0.5–6.0 | 1–15 | 1–20 |
| Nitrate reduction | - | + (-) | + | + | + | - | + | - | + | ND | + | + | + |
| Nitrite reduction | - | + (-) | - | + (-) | - (+) | - | - | - | - | ND | - | + | - (+) |
| Hydrolysis of starch | + | + | - | - | - | - | - | - | + | - | - | - | - |
| Indole production | - | - | - | - | - | - | - | - | ND | ND | + | - | - |
| Fermentation of: | |||||||||||||
| D-Glucose | - | - | w | w | - | w | - | ND | ND | ND | + | + | - |
| Lactose | - | - | - | - | - | w | - | ND | ND | ND | ND | ND | - |
| Utilisation of: | |||||||||||||
| Glycogen | + | + | + | - | - | - | - | + | ND | - | ND | ND | - |
| Mono-methyl-succinate | + | - | + | + (-) | + | + | w | - | ND | - | ND | ND | + |
| γ-Hydroxy-butyric acid | + | + | + | - | - | w (+) | w | - | ND | - | ND | ND | - |
| Succinic acid | + | - (+) | - | - | + | w | - (+) | - | + | - | + | ND | + |
| L-Glutamic acid | + | + | + | w (-) | + | + | + (-) | - | - | - | + | ND | + |
| L-Phenylalanine | - | + | - | - | - (+) | - | - | - | ND | - | ND | ND | - |
| L-Serine | + | - | - | - (+) | - | - | - | - | ND | - | ND | ND | - |
| Glycerol | + | - (+) | - | - | w (+) | + (-) | - | - | + | - | + | + | - |
| DNA G+C content (mol%) | 57.1 | 55.0 | 57.6 | 56.5 | 55.9 | 56.9 | 58.0 | 57.0 | 57.9 | ND | 57.1 | 57.1 | 52.7 |
Strains: 1, strain R9SW1T; 2, M. algicola LMG 23835T; 3, strain A3d10T; 4, M. sediminum LMG 23833T; 5, M. salsuginis CIP 109893T; 6, M. adhaerens CIP 110141T; 7, M. flavimaris CIP 108615T; 8, M. lipolyticus SM19T; 9, M. gudaonensis SL014B61AT; 10, M. goseongensis En6T; 11, M. xestospongiae UST090418-1611T; 12, M. guineae M3BT; 13, M. hydrocarbonoclasticus SP.17T.
Data for nitrate and nitrite reduction, starch hydrolysis, fermentation, indole and acid production, organic substrates utilisation, and enzyme activities for strains R9SW1T, M. algicola LMG 23835T, A3d10T, M. sediminum LMG 23833T, M. salsuginis CIP 109893T, M. adhaerens CIP 110141T, M. flavimaris CIP 108615T and M. hydrocarbonoclasticus SP. 17T are from this study. The data in brackets are from previously published work [2], [7], [62]–[70].
+, Positive; -, Negative; w, Weak reaction; ND, No Data available.
The susceptibility of the bacteria to antibiotics was tested using modified media containing: 21 g L−1 Mueller-Hinton medium (Oxoid, UK); 7.5% Sea salt and 15 g L−1 bacteriological agar (Agar No. 1, Oxoid, UK). The antibiotics tested were penicillin G (10 µg), chloramphenicol (30 µg), streptomycin (10 µg), tetracycline (30 µg), ampicillin (10 µg) and oxacillin (1 µg). The strains were incubated under optimal temperature conditions and results were obtained after 24 hours of incubation.
The ability of the strains to oxidise a range of organic substrates was investigated using a 96-well Biolog GN2 microplate (Biolog, USA), in triplicate. Inoculates were prepared by suspending culture that had been grown overnight into 3% (w/v) saline solution, then adjusting the density of the suspension to McFarland standard no. 1, followed by pipetting 150 µL aliquots of the suspension into each well. All the plates were incubated at 30°C and results were manually obtained after 24 h and 48 h. Enzymatic tests were performed using API ZYM test strips (bioMérieux, France) in two individual experiments. Inoculations were prepared by suspending culture that had been grown overnight into 3% (w/v) saline solution and adjusting the density to McFarland standard no. 5. A Microbact 24E Gram-negative identification system (Oxoid, UK) was also used to test other biochemical reactions, namely: lysine and ornithine decarboxylase; H2S production; glucose, mannitol and xylose fermentation; hydrolysis of o-nitrophenyl-β-D-galactopyranoside (ONPG); indole production; urea hydrolysis; acetoin production (Voges-Proskaüer reaction); citrate utilisation; production of indolepyruvate; gelatin liquefaction; malonate inhibition; inositol, sorbitol, rhamnose, sucrose, lactose, arabinose, adonitol, raffinose and salicin fermentation; and arginine dihydrolase. All tests were carried out according to the manufacturer's specifications unless otherwise stated.
Fatty acids analysis
Fatty acid (FA) methyl esters were prepared as described elsewhere [44]. The resulting fatty acid methyl esters were analysed using a Shimadzu GC-14A gas chromatograph with a flame ionization detector, using both a nonpolar SPB-5 fused-silica column (30 m×0.25 mm i.d.) at 210°C and a polar Supelcowax-10 fused-silica column (30 m×0.25 mm i.d.) at 200°C.
Results and Discussion
Analysis of the complete 16S rRNA gene sequences of strains R9SW1T and A3d10T revealed that both strains are grouped with species of the genus Marinobacter, with the sequence similarity between strains R9SW1T, A3d10T and all validly described Marinobacter species being in the range of 93.84–99.40% and 93.91–99.53%, respectively. The two new strains, R9SW1T and A3d10T shared 97.6% of their 16S rRNA gene sequences, however, phylogenetic analysis showed that they cluster separately forming two different clusters, one with M. algicola DG893T and another with M. sediminum R65T, where both clusters were supported by the bootstrap value of 99% and 100% in both the NJ and ML methods (Figure 1A and Figure S1 in File S1). The highest 16S rRNA gene sequence similarity between strain R9SW1T and M. algicola DG893T was found to be 99.40% (M. algicola DG893T), whilst strain A3d10T displays the highest 16S rRNA gene sequence similarity with M. sediminum R65T (99.53%).
Figure 1. Neighbour-joining phylogenetic tree showing the taxonomic position of strains R9SW1T and A3d10T according to their (A) 16S rRNA, (B) gyrB and (C) rpoD gene sequences.

Numbers at branching points are percentage bootstrap values based on 1000 replications, with only values above 50% are shown. Scale bar represents 0.005/0.02 substitutions per nucleotide position. The Maximum-likelihood (ML) and maximum Parsimony (MP) algorithms were also used for tree construction, where branches in agreement with ML and MP methods were marked with + and X respectively.
Due to the high 16S rRNA gene sequence similarity between strains R9SW1T and M. algicola DG893T, and between A3d10T and M. sediminum R65T, an extended phylogenetic analysis based on gyrB and rpoD genes was carried out. The use of housekeeping genes in phylogenetic analysis can be beneficial, in that it overcomes the possibility of the presence of nucleotide polymorphisms in the 16S rRNA gene [45], [46]. Two genes, gyrB and rpoD, were selected, since they have been previously reported to be excellent marker genes and sufficient for the identification and classification of various groups of microorganism [25], [47]–[49]. M. sediminum LMG 23833T, M. salsuginis CIP 109893T, M. algicola LMG 23835T, M. adhaerens CIP 110141T, and M. flavimaris CIP 108615T were selected, as they are phylogenetically close to strains R9SW1T and A3d10T according to their 16S rRNA gene sequences. M. hydrocarbonoclasticus SP.17T was also included as representing the type species of the genus. A phylogenetic analysis of the gyrB and rpoD gene sequence similarities reconfirmed the clustering of strain R9SW1T with M. algicola LMG 23835T, and strain A3d10T with M. sediminum LMG 23833T, both of which were supported with 100% bootstrap values (Figure 1(B) and (C)). The gyrB and rpoD sequence similarities for strains R9SW1T, A3d10T and their phylogenetically related species was also determined and found to be in the range of 77.8–94.3% (R9SW1T, gyrB), 80.0–93.5% (A3d10T, gyrB), and 78.6–93.8% (R9SW1T, rpoD), 78.6–96.2% (A3d10T, rpoD), respectively (Table 3). The gene sequence similarity for gyrB and rpoD between the previously described sister species of Marinobacter, i.e., M. adhaerens CIP 110141T and M. flavimaris CIP 108615T was found to be 99.0% and 98.4% respectively (Table 3), which is higher than that found for strains R9SW1T, A3d10T and their respective closest phylogenetic relatives. The sequence similarities of the gyrB gene of 94.3% and 93.5% for strains R9SW1T, A3d10T with their closest relatives were also lower than the previously proposed gyrB sequence similarity cut-off value of 98.95% for genus Amycolatopsis [50] and 98.22% for genus Kribbella [51]. Also, the data reported for the two Vibrio species, V. gigantis LGP 13T and V. crassostreae LGP 7T, were 98% for gyrB and 97% for rpoD [52], which again showed higher similarity values than the gyrB and rpoD sequence similarities of strains R9SW1T, A3d10T and their closest relatives. The sequence similarities for gyrB and rpoD between strains R9SW1T and A3d10T were significantly lower than the values mentioned above, i.e., 81.6% for gyrB and 78.2% for rpoD, suggesting distinct standing of new strains on the species level.
Table 3. The gyrB and rpoD gene sequence similarities of strains R9SW1T, A3d10T and phylogenetically related type strains and type species of the genus Marinobacter.
| Similarity of gyrB/rpoD genes (%) | ||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 1. M. adhaerens CIP 110141T | 100/100 | |||||||
| 2. M. algicola LMG 23835T | 78.0/81.2 | 100/100 | ||||||
| 3. M. flavimaris CIP 108615T | 99.0/98.4 | 77.8/81.0 | 100/100 | |||||
| 4. M. hydrocarbonoclasticus SP. 17T | 80.7/81.7 | 78.2/77.0 | 80.0/81.5 | 100/100 | ||||
| 5. M. salsuginis CIP 109893T | 86.5/93.4 | 76.2/80.3 | 86.1/93.5 | 80.8/80.0 | 100/100 | |||
| 6. M. sediminum LMG 23833T | 84.1/83.5 | 80.3/77.8 | 83.8/84.2 | 83.6/78.7 | 85.8/84.0 | 100/100 | ||
| 7. Marinobacter sp. A3d10T | 83.7/83.8 | 80.0/78.8 | 83.6/84.4 | 82.2/78.6 | 84.7/84.1 | 93.5/96.2 | 100/100 | |
| 8. Marinobacter sp. R9SW1T | 78.2/80.5 | 94.3/93.8 | 78.2/80.3 | 77.8/79.5 | 78.0/80.6 | 81.9/78.6 | 81.6/78.2 | 100/100 |
In order to further assess the taxonomic affiliation of the two new bacteria, a comparative analysis of the total protein profiles was performed using MALDI-TOF mass spectrometry (Figure 2). The results are in agreement with the phylogenetic analyses, clearly indicating that strain R9SW1T is clustering with M. algicola LMG 23835T, and strain A3d10T is clustering with M. sediminum LMG 23833T with a critical distance level below 500. As suggested in the previously reported studies, clustering below the distance level of 500 can be considered as reliable clustering [53], [54], which was also in agreement with the recent studies on Alteromonas spp., where the clustering within the distance level of 500 was shown to be able to differentiate the closely related Alteromonas species [25], [55]. Hence, the results of this study confirmed the confident clustering of the two new isolates within other species of the genus Marinobacter. Also, the clusters of both strains R9SW1T and A3d10T with their nearest neighbour were stable, but exceeded the minimum differences between existing species, e.g., the distance level between species in both clusters were greater than those within a cluster that contained M. gudaonensis CIP 109534T, M. adhaerens CIP 110141T, M. salsuginis CIP 109893T, and M. flavimaris CIP 108615T; so does the position of strains R9SW1T and A3d10T resulting in different clusters in the MALDI dendrogram, provide evidence of the distinctive standing of two new bacteria.
Figure 2. Main spectra library (MSP) dendrogram of MALDI-TOF mass spectral profiles of strains R9SW1T, A3d10T and closely related Marinobacter species.

The dendrogram was generated by MALDI Biotyper 3.0 software with distance is displayed in relative units.
In order to confirm the separate species standing of these two strains, a DNA-DNA hybridization experiment was conducted. DNA-DNA relatedness between strain R9SW1T and M. algicola LMG 23835T was found to be 63.05±1.85%, and between strain A3d10T and M. sediminum LMG 23833T was found to be 67.60±1.3%. Both of these relatedness values are below the 70% cut-off value generally recommended for species differentiation [56]. Recently, information of whole genome sequences have been recommended to be integrated into bacterial systematics [57]–[59]. In this study, whole genome sequences of strains R9SW1T, A3d10T, M. adhaerens HP15T and M. hydrocarbonoclasticus ATCC 49840T were visually compared using BLAST (Figure S2 in File S1) and the average nucleotide identity (ANI), genome-to-genome distance (GGD), average amino acid identity (AAI), and the Karlin's genomic signature dissimilarity (δ*) between the four strains were calculated, the results of which are presented in Table 4. Due to the lack of the availability of the assembled, whole genome sequences for validly named Marinobacter species, genomic signatures between strains R9SW1T, A3d10T and validly described Marinobacter species can only be performed using those of M. adhaerens HP15T [31] and M. hydrocarbonoclasticus ATCC 49840T [30]. As can be seen from the information presented in Table 4, the ANIs between the four strains were in the range of 82.3–83.3%, which is significantly lower than the suggested threshold range of 95–96% [58], [60]; the GGDs were calculated to be in the range of 19.8–20.7% which is lower than the cur-off value of 70% [61]; the AAI and Karlin signature dissimilarity values for the four strains were in the range of 68.1–77.6% and 31–36 respectively, each of which fall outside the range to be consider as same species [42], [61]; and thus again indicating that strains R9SW1T and A3d10T can be considered as two novel species of the genus Marinobacter. The distinct standing of strains R9SW1T and A3d10T can also be confirmed by the phylogenomic relationship analysis using the core proteome of the genomes from the four strains (Figure 3).
Table 4. The genomic signatures between strains R9SW1T, A3d10T, M. adhaerens HP15T and M. hydrocarbonoclasticus ATCC 49840T.
| Genomic signatures | ||||
| 1 | 2 | 3 | 4 | |
| 1. M. hydrocarbonoclasticus ATCC 49840T | 20.1/35 | 20/31 | 19.8/32 | |
| 2. M. adhaerens HP15T | 83.1/74.5 | 20.2/36 | 20.7/35 | |
| 3. Strain R9SW1T | 82.3/69.5 | 82.7/72.6 | 19.8/31 | |
| 4. Strain A3d10T | 82.5/72.7 | 83.3/77.6 | 82.3/68.1 | |
Data in the lower triangular corresponds to ANI/AAI (%) and data in the upper triangular corresponds to GGD (%)/Karlin signature.
Figure 3. Phylogenomic tree of strains R9SW1T, A3d10T, M. hydrocarbonoclasticus ATCC 49840T and M. adhaerens HP15T constructed using concatenated sequence of the core proteome (544,643 bp) of the genomes.
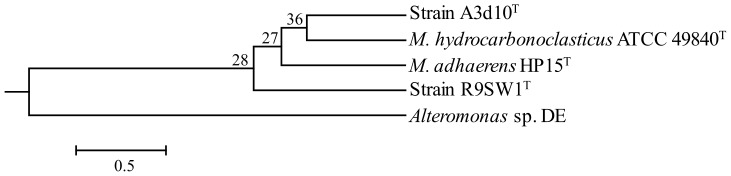
Alteromonas sp. DE was used as outgroup.
The major features of the genomes of strains R9SW1T and A3d10T were identified as described elsewhere [16]. Briefly, they are 4,616,532 bp and 3,975,896 bp in size, composed of 99 and 29 contigs, both have 3 rRNAs, and 44 and 46 tRNAs, for strains R9SW1T and A3d10T, respectively. The DNA G+C content of strains R9SW1T and A3d10T were found to be 57.1 and 57.6 mol%, respectively (Table 2), the values which are consistent with those of the genus Marinobacter.
Both bacteria were found to be Gram-negative, aerobic, motile by means of a single flagellum and rod-shaped with the size of 1.9–3.2×0.40–0.72 µm for strain R9SW1T and 1.3–2.1×0.40–0.45 µm for strain A3d10T (Figure S3 in File S1). The catalase and oxidase tests were found to be positive, H2S and indole tests were found to be negative. It can be seen that strain R9SW1T can be clearly differentiated from M. algicola LMG 23835T by its inability to reduce nitrate and nitrite, its ability to utilise mono-methyl succinate and L-serine, its inability to utilise L-phenylalanine and the absence of lipase (C14); while strain A3d10T can be clearly differentiated from M. sediminum LMG 23833T by its inability to reduce nitrite, its ability to utilise glycogen, γ-hydroxy-butyric acid and L-glutamic acid, and its weak activities for valine arylamidase and cystine arylamidase. The major phenotypic difference between strains R9SW1T and A3d10T are nitrate reduction, hydrolysis of starch, fermentation of D-glucose, and their utilisation of dextrin, D-fructose, maltose, acetic acid, propionic acid, succinic acid, L-serine and glycerol. Other phenotypic characteristics which differentiate the two novel strains from each other and their closest phylogenetic neighbours are shown in Table 2, Table S2 in File S1, and in their respective species descriptions. Both strains were found to be sensitive to penicillin G (10 µg), chloramphenicol (30 µg), and ampicillin (10 µg), and resistant to streptomycin (10 µg) and tetracycline (30 µg). The fatty acid composition of strains R9SW1T and A3d10T are shown in Table S3 in File S1, where the predominant fatty acids were identified as being C16:0, C16:1 ω7c, C18:1 ω9c and C18:1 ω7c.
The genotype to phenotype analyses were also carried out based on the whole genome sequences of the four strains, the results of which are presented in Table 5. It can be seen that of the results of physiological and biochemical tests match when comparing the in silico results, however a few discrepancies are noted. A similar level of deviation previously reported in the case of Vibrio species and it was suggested that expression of certain genes may be restricted by stop codon, repressor genes, regulatory proteins, global regulators, genome coverage or sequencing errors [42].
Table 5. Comparative identification of phenotypic characteristics based on genomic analysis and physiological and biochemical tests.
| Strain R9SW1T | Strain A3d10T | M. hydrocarbonoclasticus ATCC 49840T | M. adhaerens HP15T | |||||
| In vitro | In silico | In vitro | n silico | In vitro | In silico | In vitro | In silico | |
| Nitrate reduction | - | + | + | + | + | + | - | - |
| Nitrite reduction | - | - | - | - | - | + | - | - |
| Hydrolysis of starch | + | - | - | - | - | - | - | - |
| Indole production | - | - | - | - | - | - | - | - |
| D-Glucose | - | - | w | - | - | - | w | - |
| Lactose | - | - | - | - | - | - | w | - |
| Glycogen | + | - | + | - | - | + | - | + |
| Mono-methyl-succinate | + | + | + | + | + | + | + | + |
| γ-Hydroxy-butyric acid | + | - | + | - | - | - | w | + |
| Succinic acid | + | + | - | - | + | + | w | + |
| L-Glutamic acid | + | + | + | + | + | + | + | + |
| L-Phenylalanine | - | - | - | - | - | - | - | - |
| L-Serine | + | + | - | + | - | - | - | - |
| Glycerol | + | + | - | - | - | - | + | - |
In summary, the comparative genomic and phylogenetic analysis based on the full-length of 16S rRNA gene sequence similarities, pheno- and chemotaxonomic properties revealed that strains R9SW1T and A3d10T can be affiliated to the genus Marinobacter. A further dual-locus sequence analysis based on gyrB and rpoD gene sequence similarities, the comparative analysis of whole cells protein profiles based on MALDI-TOF mass spectrometry analysis, their phenotypic characteristics and their DNA-DNA hybridization values below 70% confirmed that strains R9SW1T and A3d10T should be classified as two novel species of the genus Marinobacter for which the name Marinobacter salarius sp. nov. and Marinobacter similis sp. nov. are proposed.
Description of Marinobacter salarius sp. nov
Marinobacter salarius (sa.la'ri.us, L. masc. adj., salarius, of or belonging to salt, pertaining to salt tolerance)
Cells are Gram-negative rods (approximately 1.9–3.2×0.40–0.72 µm). Motile by means of a single polar flagellum. Colonies on marine agar are semi-translucent, non-pigmented, circular to slightly irregular (0.8–1.0 mm) and smooth after 48 hours of incubation. Colonies turn to creamy in colour with increasing incubation time. Growth occurs at 4°C–40°C (optimum, 25°C–30°C), between pH 6–9 (optimum, pH 7.5) and in the presence of 0.5–20% (w/v) NaCl. No growth was observed at 0 or 25% (w/v) NaCl. Catalase and oxidase tests are positive. Starch, Tween 40 and 80 are positive, while nitrate and nitrite reduction are negative. Indole, lysine decarboxylase, ornithine decarboxylase, β-galactosidase, tryptophan deaminase, gelatinase, arginine dihydrolase, acetoin, urea and H2S are not produced. Acid is not produced from glucose, mannitol, xylose, inositol, sorbitol, rhamnose, sucrose, lactose, arabinose, adonitol, raffinose and salicin. According to API ZYM, strain R9SW1T is positive for alkaline phosphatase, esterase (C4), esterase lipase (C8), leucine arylamidase, valine arylamidase, cystine arylamidase and N-acetyl-β-glucosaminidase; weakly positive for acid phosphatase, naphthol-AS-BI-phosphohydrolase and α-glucosidase; negative for lipase (C14), trypsin, α-chymotrypsin, α-galactosidase, β-galactosidase, β-glucuronidase, β-glucosidase, α-mannosidase and α-fucosidase. Positive for the utilization of dextrin, glycogen, D-fructose, maltose, methyl-pyruvate, mono-methyl-succinate, acetic acid, β-hydroxybutyric acid, γ-hydroxybutyric acid, DL-lactic acid, propionic acid, succinic acid, L-glutamic acid, L-proline, L-serine and glycerol; weakly positive for the utilization of L-alaninamide, D-alanine, L-alanine and L-leucine; negative for the utilization of α-cyclodextrin, N-acetyl-D-galactosamine, N-acetyl-D-glucosamine, adonitol, L-arabinose, D-arabitol, D-cellobiose, i-erythritol, L-fucose, D-galactose, gentiobiose, α-D-glucose, m-inositol, α-D-lactose, lactulose, D-mannitol, D-mannose, D-melibiose, β-methyl-D-glucoside, D-psicose, D-raffinose, L-rhamnose, D-sorbitol, sucrose, D-trehalose, turanose, xylitol, cis-aconitic acid, citric acid, formic acid, D-galactonic acid lactone, D-galacturonic acid, D-gluconic acid, D-glucosaminic acid, D-glucuronic acid, α-hydroxybutyric acid, p-hydroxyphenylacetic acid, itaconic acid, α-ketoglutaric acid, α-ketobutyric acid, α-ketovaleric acid, malonic acid, quinic acid, D-saccharic acid, sebacic acid, bromosuccinic acid, succinamic acid, glucuronamide, L-alanyl-glycine, L-asparagine, L-aspartic acid, glycyl-L-aspartic acid, glycyl-L-glutamic acid, L-histidine, hydroxyl-L-proline, L-ornithine, L-phenylalanine, L-pyroglutamic acid, D-serine, L-threonine, DL-carnitine, γ-aminobutyric acid, urocanic acid, inosine, uridine, thymidine, phenyethylamine, putrescine, 2-aminoethanol, 2,3-butanediol, DL-α-glycerol, glucose-1-phosphate and glucose-6-phosphate as the sole carbon and energy source. The main cellular fatty acids are C16:0, C16:1 ω7c, C18:1 ω9c and C18:1 ω7c. The G+C content of the type strain is 57.1 mol%. The type strain is R9SW1T ( = LMG 27497T = JCM 19399T = CIP 110588T = KMM 7502T), isolated from sea water from Chazhma Bay in the Sea of Japan, Pacific Ocean. The accession number for the whole genome sequence of strain R9SW1T is CP007152.
Description of Marinobacter similis sp. nov
Marinobacter similis (si'mi.lis, L. masc. adj., similis, like, resembling, similar, pertaining to close similarity with other species)
Cells are Gram-negative rods (approximately 1.3 - 2.1×0.40 - 0.45 µm). Motile by means of a single polar flagellum. Colonies on marine agar are semi-translucent, non-pigmented, circular to slightly irregular (0.5 – 1.0 mm) and smooth after 48 hours of incubation. Colonies turn to creamy in colour with increasing incubation time. Growth occurs at 4°C - 40°C (optimum, 25°C - 30°C), between pH 6 to 9 (optimum, pH 7.5) and in the presence of 0.5–20% (w/v) NaCl. No growth was observed at 0 or 25% w/v NaCl. Catalase and oxidase tests are positive. Tween 40 and 80 are positive, while starch is not. Nitrate is reduced but not nitrite. Indole, lysine decarboxylase, ornithine decarboxylase, β-galactosidase, tryptophan deaminase, gelatinase, arginine dihydrolase, acetoin, urea and H2S are not produced. Acid is not produced from glucose, mannitol, xylose, inositol, sorbitol, rhamnose, sucrose, lactose, arabinose, adonitol, raffinose and salicin. According to API ZYM, strain A3d10T is positive for alkaline phosphatase, esterase (C4), esterase lipase (C8), leucine arylamidase, naphthol-AS-BI-phosphohydrolase, and N-acetyl-β-glucosaminidase; weakly positive for lipase (C14), valine arylamidase, cystine arylamidase and acid phosphatase; negative for trypsin, α-chymotrypsin, α-galactosidase, β-galactosidase, β-glucuronidase, β-glucosidase, α-glucosidase, α-mannosidase and α-fucosidase. Positive for the utilization of, glycogen, methyl-pyruvate, mono-methyl-succinate, β-hydroxybutyric acid, γ-hydroxybutyric acid, DL-lactic acid, D-alanine, L-alanine, L-glutamic acid and L-proline; negative for the utilization of α-cyclodextrin, dextrin, N-acetyl-D-galactosamine, N-acetyl-D-glucosamine, adonitol, L-arabinose, D-arabitol, D-cellobiose, i-erythritol, D-fructose, L-fucose, D-galactose, gentiobiose, α-D-glucose, m-inositol, α-D-lactose, lactulose, maltose, D-mannitol, D-mannose, D-melibiose, β-methyl-D-glucoside, D-psicose, D-raffinose, L-rhamnose, D-sorbitol, sucrose, D-trehalose, turanose, xylitol, acetic acid, cis-aconitic acid, citric acid, formic acid, D-galactonic acid lactone, D-galacturonic acid, D-gluconic acid, D-glucosaminic acid, D-glucuronic acid, α-hydroxybutyric acid, p-hydroxyphenylacetic acid, itaconic acid, α-ketoglutaric acid, α-ketobutyric acid, α-ketovaleric acid, malonic acid, propionic acid, quinic acid, D-saccharic acid, sebacic acid, succinic acid, bromosuccinic acid, succinamic acid, glucuronamide, L-alaninamide, L-alanyl-glycine, L-asparagine, L-aspartic acid, glycyl-L-aspartic acid, glycyl-L-glutamic acid, L-histidine, hydroxyl-L-proline, L-leucine, L-ornithine, L-phenylalanine, L-pyroglutamic acid, D-serine, L-serine, L-threonine, DL-carnitine, γ-aminobutyric acid, urocanic acid, inosine, uridine, thymidine, phenyethylamine, putrescine, 2-aminoethanol, 2,3-butanediol, glycerol, DL-α-glycerol, glucose-1-phosphate and glucose-6-phosphate as the sole carbon and energy source. The main cellular fatty acids are C16:0, C16:1 ω7c, C18:1 ω9c and C18:1 ω7c. The G+C content of the type strain is 57.6 mol%. The type strain is A3d10T ( = JCM 19398T = CIP 110589T = KMM 7501T), isolated from sea water from Port Philip Bay of the Tasman Sea, the Pacific Ocean. The accession number for the whole genome sequence for strain A3d10T is CP007151.
Supporting Information
Includes Figures S1–S3 and Tables S1–S3. Figure S1. Neighbour-joining phylogenetic tree showing the taxonomic position of strains R9SW1T and A3d10T according to their 16S rRNA gene sequences. Figure S2. BLAST genome ring (A) and comparison of all proteins in the genomes in terms of the similar composition of the gene families (B) between strains R9SW1T, A3d10T, M. adhaerens HP15T and M. hydrocarbonoclasticus ATCC 49840T. Figure S3. Scanning electron micrographs of strains (A) R9SW1T and (B) A3d10T. Table S1. Genes and the corresponding primer sequences used for the amplification and sequencing. Table S2. Phenotypic characteristics of strains R9SW1T, A3d10T and closely related type strains and type species of the genus Marinobacter. Table S3. Cellular fatty acids composition of strains R9SW1T, A3d10T and closely related type strains and type species of the genus Marinobacter.
(DOCX)
Acknowledgments
We would like to thank Professor Dr Bernhard Schink for verification of species epithet etymology and Bioscreen and Bio21 Institute for the access to the MALDI-TOF MS instrument.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported in part by Genome Information Upgrading Program of National BioResource Project from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan. Water sample collection was within the research program funded by the Federal Agency for Science of the Ministry of Education and Science of the Russian Federation, grant 2-2.16, and by the Russian Foundation for Basic Research and grant ‘Molecular and Cell Biology’ from the Presidium of the Russian Academy of Sciences, grant 02-04-48211. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Euzeby JP (1997) List of bacterial names with standing in nomenclature: a folder available on the Internet. Int J Syst Bacteriol 47: 590–592. [DOI] [PubMed] [Google Scholar]
- 2. Gauthier MJ, Lafay B, Christen R, Fernandez L, Acquaviva M, et al. (1992) Marinobacter hydrocarbonoclasticus gen. nov., sp. nov., a new, extremely halotolerant, hydrocarbon-degrading marine bacterium. Int J Syst Bacteriol 42: 568–576. [DOI] [PubMed] [Google Scholar]
- 3.Bowman J, McMeekin T (2005) Alteromonadales ord. nov. In: Brenner D, Krieg N, Staley J, Garrity G, Boone D, et al.., editors. Bergey's Manual of Systematic Bacteriology: Springer US. pp. 443–491.
- 4. Huu NB, Denner EB, Ha DT, Wanner G, Stan-Lotter H (1999) Marinobacter aquaeolei sp. nov., a halophilic bacterium isolated from a Vietnamese oil-producing well. Int J Syst Bacteriol 49 Pt 2: 367–375. [DOI] [PubMed] [Google Scholar]
- 5. Marquez MC, Ventosa A (2005) Marinobacter hydrocarbonoclasticus Gauthier, et al. 1992 and Marinobacter aquaeolei Nguyen, et al. 1999 are heterotypic synonyms. Int J Syst Evol Microbiol 55: 1349–1351. [DOI] [PubMed] [Google Scholar]
- 6. Shivaji S, Gupta P, Chaturvedi P, Suresh K, Delille D (2005) Marinobacter maritimus sp. nov., a psychrotolerant strain isolated from sea water off the subantarctic Kerguelen islands. Int J Syst Evol Microbiol 55: 1453–1456. [DOI] [PubMed] [Google Scholar]
- 7. Green DH, Bowman JP, Smith EA, Gutierrez T, Bolch CJ (2006) Marinobacter algicola sp. nov., isolated from laboratory cultures of paralytic shellfish toxin-producing dinoflagellates. Int J Syst Evol Microbiol 56: 523–527. [DOI] [PubMed] [Google Scholar]
- 8. Cui Z, Lai Q, Dong C, Shao Z (2008) Biodiversity of polycyclic aromatic hydrocarbon-degrading bacteria from deep sea sediments of the Middle Atlantic Ridge. Environ Microbiol 10: 2138–2149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Dastgheib SMM, Amoozegar MA, Khajeh K, Shavandi M, Ventosa A (2012) Biodegradation of polycyclic aromatic hydrocarbons by a halophilic microbial consortium. Appl Microbiol Biotechnol 95: 789–798. [DOI] [PubMed] [Google Scholar]
- 10.Lal D, Jindal S, Kumari H, Jit S, Nigam A, et al. (2013) Bacterial diversity and real-time PCR based assessment of linA and linB gene distribution at hexachlorocyclohexane contaminated sites. J Basic Microbiol: doi: 10.1002/jobm.201300211. [DOI] [PubMed]
- 11. Berlendis S, Cayol JL, Verhé F, Laveau S, Tholozan JL, et al. (2010) First evidence of aerobic biodegradation of BTEX compounds by pure cultures of Marinobacter . Appl Biochem Biotechnol 160: 1992–1999. [DOI] [PubMed] [Google Scholar]
- 12. Ivanova EP, Gorshkova NM, Mikhailov VV, Sergeev AF, Gladkikh RV, et al. (2005) Distribution of saprophytic bacteria in the atomic submarine accident zone in Chazhma Bay, Sea of Japan. Russ J Mar Biol 31: 65–72. [Google Scholar]
- 13. Webb HK, Crawford RJ, Sawabe T, Ivanova EP (2009) Poly(ethylene terephthalate) polymer surfaces as a substrate for bacterial attachment and biofilm formation. Microbes Environ 24: 39–42. [DOI] [PubMed] [Google Scholar]
- 14. Ivanova EP, Kiprianova EA, Mikhailov VV, Levanova GF, Garagulya AD, et al. (1996) Characterization and identification of marine Alteromonas nigrifaciens strains and emendation of the description. Int J Syst Bacteriol 46: 223–228. [DOI] [PubMed] [Google Scholar]
- 15. Gorshkova NM, Ivanova EP, Sergeev AF, Zhukova NV, Alexeeva Y, et al. (2003) Marinobacter excellens sp. nov., isolated from sediments of the Sea of Japan. Int J Syst Evol Microbiol 53: 2073–2078. [DOI] [PubMed] [Google Scholar]
- 16. Ivanova EP, Ng HJ, Webb HK, Feng G, Oshima K, et al. (2014) Draft genome sequences of Marinobacter similis A3d10T and Marinobacter salarius R9SW1T . Genome Announc 2(3): e00442–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Yamamoto S, Harayama S (1995) PCR amplification and direct sequencing of gyrB genes with universal primers and their application to the detection and taxonomic analysis of Pseudomonas putida strains. Appl Environ Microbiol 61: 1104–1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Yamamoto S, Harayama S (1998) Phylogenetic relationships of Pseudomonas putida strains deduced from the nucleotide sequences of gyrB, rpoD and 16S rRNA genes. Int J Syst Bacteriol 48 Pt 3: 813–819. [DOI] [PubMed] [Google Scholar]
- 19. Thompson JD, Higgins DG, Gibson TJ (1994) Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap enalties and weight matrix choice. Nucleic Acids Res 22: 4673–4680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Saitou N, Nei M (1987) The neighbor-joining method - a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406–425. [DOI] [PubMed] [Google Scholar]
- 21. Felsenstein J (1981) Evolutionary trees from DNA sequences: a maximum likelihood approach. J Mol Evol 17: 368–376. [DOI] [PubMed] [Google Scholar]
- 22. Fitch WM (1971) Toward defining the course of evolution: minimum change for a specific tree topology. Syst Biol 20: 406–416. [Google Scholar]
- 23. Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide-sequences. J Mol Evol 16: 111–120. [DOI] [PubMed] [Google Scholar]
- 24. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, et al. (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28: 2731–2739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Ng HJ, Webb HK, Crawford RJ, Malherbe F, Butt H, et al. (2013) Updating the taxonomic toolbox: classification of Alteromonas spp. using multilocus phylogenetic analysis and MALDI-TOF mass spectrometry. Anton Leeuw Int J G 103: 265–275. [DOI] [PubMed] [Google Scholar]
- 26. Lagier JC, Gimenez G, Robert C, Raoult D, Fournier PE (2012) Non-contiguous finished genome sequence and description of Herbaspirillum massiliense sp. nov. Stand Genomic Sci 7: 200–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Cashion P, Holder-Franklin MA, McCully J, Franklin M (1977) A rapid method for the base ratio determination of bacterial DNA. Anal Biochem 81: 461–466. [DOI] [PubMed] [Google Scholar]
- 28. De Ley J, Cattoir H, Reynaerts A (1970) The quantitative measurement of DNA hybridization from renaturation rates. Eur J Biochem 12: 133–142. [DOI] [PubMed] [Google Scholar]
- 29. Huss VA, Festl H, Schleifer KH (1983) Studies on the spectrophotometric determination of DNA hybridization from renaturation rates. Syst Appl Microbiol 4: 184–192. [DOI] [PubMed] [Google Scholar]
- 30. Grimaud R, Ghiglione JF, Cagnon C, Lauga B, Vaysse PJ, et al. (2012) Genome sequence of the marine bacterium Marinobacter hydrocarbonoclasticus SP17, which forms biofilms on hydrophobic organic compounds. J Bacteriol 194: 3539–3540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Gärdes A, Kaeppel E, Shehzad A, Seebah S, Teeling H, et al. (2010) Complete genome sequence of Marinobacter adhaerens type strain (HP15), a diatom-interacting marine microorganism. Standards in Genomic Sciences 3: 97–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Goris J, Konstantinidis KT, Klappenbach JA, Coenye T, Vandamme P, et al. (2007) DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int J Syst Evol Microbiol 57: 81–91. [DOI] [PubMed] [Google Scholar]
- 33. Alikhan NF, Petty NK, Ben Zakour NL, Beatson SA (2011) BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12: 402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Auch AF, von Jan M, Klenk HP, Goker M (2010) Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand Genomic Sci 2: 117–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Meier-Kolthoff JP, Auch AF, Klenk HP, Goker M (2013) Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics 14: 60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Konstantinidis KT, Tiedje JM (2005) Towards a genome-based taxonomy for prokaryotes. J Bacteriol 187: 6258–6264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389–3402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Karlin S, Mrazek J, Campbell AM (1997) Compositional biases of bacterial genomes and evolutionary implications. J Bacteriol 179: 3899–3913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Darling AC, Mau B, Blattner FR, Perna NT (2004) Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res 14: 1394–1403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Didelot X, Lawson D, Darling A, Falush D (2010) Inference of homologous recombination in bacteria using whole-genome sequences. Genetics 186: 1435–1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Ivars-Martinez E, Martin-Cuadrado AB, D'Auria G, Mira A, Ferriera S, et al. (2008) Comparative genomics of two ecotypes of the marine planktonic copiotroph Alteromonas macleodii suggests alternative lifestyles associated with different kinds of particulate organic matter. ISME J 2: 1194–1212. [DOI] [PubMed] [Google Scholar]
- 42. Amaral GR, Dias GM, Wellington-Oguri M, Chimetto L, Campeao ME, et al. (2014) Genotype to phenotype: identification of diagnostic vibrio phenotypes using whole genome sequences. Int J Syst Evol Microbiol 64: 357–365. [DOI] [PubMed] [Google Scholar]
- 43.Smibert R, Krieg N (1994) Phenotypic characterization. In: Gerhart P, Murray R, Wood W, Krieg N, editors. In Methods for General and Molecular Bacteriology. Washington DC: American Society for Microbiology. pp. 607–654. [Google Scholar]
- 44. Svetashev VI, Vysotskii MV, Ivanova EP, Mikhailov VV (1995) Cellular fatty acids of Alteromonas species. Syst Appl Microbiol 18: 37–43. [Google Scholar]
- 45. Cilia V, Lafay B, Christen R (1996) Sequence heterogeneities among 16S ribosomal RNA sequences, and their effect on phylogenetic analyses at the species level. Mol Biol Evol 13: 451–461. [DOI] [PubMed] [Google Scholar]
- 46. Alperi A, Figueras MJ, Inza I, Martínez-Murcia AJ (2008) Analysis of 16S rRNA gene mutations in a subset of Aeromonas strains and their impact in species delineation. Int Microbiol 11: 185–194. [PubMed] [Google Scholar]
- 47. Yamamoto S, Kasai H, Arnold DL, Jackson RW, Vivian A, et al. (2000) Phylogeny of the genus Pseudomonas: intrageneric structure reconstructed from the nucleotide sequences of gyrB and rpoD genes. Microbiology 146 (Pt 10): 2385–2394. [DOI] [PubMed] [Google Scholar]
- 48. Puthucheary SD, Puah SM, Chua KH (2012) Molecular characterization of clinical isolates of Aeromonas species from Malaysia. PLoS ONE 7(2): e30205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Táncsics A, Benedek T, Farkas M, Máthé I, Márialigeti K, et al. (2014) Sequence analysis of 16S rRNA, gyrB and catA genes and DNA-DNA hybridization reveal that Rhodococcus jialingiae is a later synonym of Rhodococcus qingshengii . Int J Syst Evol Microbiol 64: 298–301. [DOI] [PubMed] [Google Scholar]
- 50. Everest GJ, Meyers PR (2009) The use of gyrB sequence analysis in the phylogeny of the genus Amycolatopsis . Anton Leeuw Int J G 95: 1–11. [DOI] [PubMed] [Google Scholar]
- 51. Kirby BM, Everest GJ, Meyers PR (2010) Phylogenetic analysis of the genus Kribbella based on the gyrB gene: proposal of a gyrB-sequence threshold for species delineation in the genus Kribbella . Anton Leeuw Int J G 97: 131–142. [DOI] [PubMed] [Google Scholar]
- 52. Le Roux F, Goubet A, Thompson FL, Faury N, Gay M, et al. (2005) Vibrio gigantis sp. nov., isolated from the haemolymph of cultured oysters (Crassostrea gigas). Int J Syst Evol Microbiol 55: 2251–2255. [DOI] [PubMed] [Google Scholar]
- 53. Sauer S, Freiwald A, Maier T, Kube M, Reinhardt R, et al. (2008) Classification and identification of bacteria by mass spectrometry and computational analysis. PLoS One 3: e2843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Dubois D, Leyssene D, Chacornac JP, Kostrzewa M, Schmit PO, et al. (2010) Identification of a variety of Staphylococcus species by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J Clin Microbiol 48: 941–945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Ivanova EP, Ng HJ, Webb HK, Kurilenko VV, Zhukova NV, et al. (2013) Alteromonas australica sp. nov., isolated from the Tasman Sea. Antonie Van Leeuwenhoek 103: 877–884. [DOI] [PubMed] [Google Scholar]
- 56. Wayne LG, Brenner DJ, Colwell RR, Grimont PAD, Kandler O, et al. (1987) Report of the ad-hoc-committee on reconciliation of approaches to bacterial systematics. Int J Syst Bacteriol 37: 463–464. [Google Scholar]
- 57. Chun J, Rainey FA (2014) Integrating genomics into the taxonomy and systematics of the Bacteria and Archaea. Int J Syst Evol Microbiol 64: 316–324. [DOI] [PubMed] [Google Scholar]
- 58. Kim M, Oh HS, Park SC, Chun J (2014) Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int J Syst Evol Microbiol 64: 346–351. [DOI] [PubMed] [Google Scholar]
- 59. Ramasamy D, Mishra AK, Lagier JC, Padhmanabhan R, Rossi M, et al. (2014) A polyphasic strategy incorporating genomic data for the taxonomic description of novel bacterial species. Int J Syst Evol Microbiol 64: 384–391. [DOI] [PubMed] [Google Scholar]
- 60. Richter M, Rossello-Mora R (2009) Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci U S A 106: 19126–19131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Thompson CC, Chimetto L, Edwards RA, Swings J, Stackebrandt E, et al. (2013) Microbial genomic taxonomy. BMC Genomics 14: 913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Romanenko LA, Schumann P, Rohde M, Zhukova NV, Mikhailov VV, et al. (2005) Marinobacter bryozoorum sp. nov. and Marinobacter sediminum sp. nov., novel bacteria from the marine environment. Int J Syst Evol Microbiol 55: 143–148. [DOI] [PubMed] [Google Scholar]
- 63. Antunes A, Franca L, Rainey FA, Huber R, Nobre MF, et al. (2007) Marinobacter salsuginis sp. nov., isolated from the brine-seawater interface of the Shaban Deep, Red Sea. Int J Syst Evol Microbiol 57: 1035–1040. [DOI] [PubMed] [Google Scholar]
- 64. Kaeppel EC, Gardes A, Seebah S, Grossart HP, Ullrich MS (2012) Marinobacter adhaerens sp. nov., isolated from marine aggregates formed with the diatom Thalassiosira weissflogii . Int J Syst Evol Microbiol 62: 124–128. [DOI] [PubMed] [Google Scholar]
- 65. Yoon JH, Yeo SH, Kim IG, Oh TK (2004) Marinobacter flavimaris sp. nov. and Marinobacter daepoensis sp. nov., slightly halophilic organisms isolated from sea water of the Yellow Sea in Korea. Int J Syst Evol Microbiol 54: 1799–1803. [DOI] [PubMed] [Google Scholar]
- 66. Martin S, Marquez MC, Sanchez-Porro C, Mellado E, Arahal DR, et al. (2003) Marinobacter lipolyticus sp. nov., a novel moderate halophile with lipolytic activity. Int J Syst Evol Microbiol 53: 1383–1387. [DOI] [PubMed] [Google Scholar]
- 67. Gu J, Cai H, Yu SL, Qu R, Yin B, et al. (2007) Marinobacter gudaonensis sp. nov., isolated from an oil-polluted saline soil in a Chinese oilfield. Int J Syst Evol Microbiol 57: 250–254. [DOI] [PubMed] [Google Scholar]
- 68. Roh SW, Quan ZX, Nam YD, Chang HW, Kim KH, et al. (2008) Marinobacter goseongensis sp. nov., from seawater. Int J Syst Evol Microbiol 58: 2866–2870. [DOI] [PubMed] [Google Scholar]
- 69. Lee OO, Lai PY, Wu HX, Zhou XJ, Miao L, et al. (2012) Marinobacter xestospongiae sp. nov., isolated from the marine sponge Xestospongia testudinaria collected from the Red Sea. Int J Syst Evol Microbiol 62: 1980–1985. [DOI] [PubMed] [Google Scholar]
- 70. Montes MJ, Bozal N, Mercade E (2008) Marinobacter guineae sp. nov., a novel moderately halophilic bacterium from an Antarctic environment. Int J Syst Evol Microbiol 58: 1346–1349. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Includes Figures S1–S3 and Tables S1–S3. Figure S1. Neighbour-joining phylogenetic tree showing the taxonomic position of strains R9SW1T and A3d10T according to their 16S rRNA gene sequences. Figure S2. BLAST genome ring (A) and comparison of all proteins in the genomes in terms of the similar composition of the gene families (B) between strains R9SW1T, A3d10T, M. adhaerens HP15T and M. hydrocarbonoclasticus ATCC 49840T. Figure S3. Scanning electron micrographs of strains (A) R9SW1T and (B) A3d10T. Table S1. Genes and the corresponding primer sequences used for the amplification and sequencing. Table S2. Phenotypic characteristics of strains R9SW1T, A3d10T and closely related type strains and type species of the genus Marinobacter. Table S3. Cellular fatty acids composition of strains R9SW1T, A3d10T and closely related type strains and type species of the genus Marinobacter.
(DOCX)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.