

Abstract
Aaptamine (8,9-dimethoxy-1H-benzo[de][1,6]naphthyridine) is a marine natural compound possessing antioxidative, antimicrobial, antifungal, and antiretroviral activity. Earlier, we have found that aaptamine and its derivatives demonstrate equal anticancer effects against the human germ cell cancer cell lines NT2 and NT2-R and cause some changes in the proteome of these cells. In order to explore further the mechanism of action of aaptamine and its derivatives, we studied the effects of aaptamine (1), demethyl(oxy)aaptamine (2), and isoaaptamine (3) on human cancer cell lines and on AP-1-, NF-κB-, and p53-dependent transcriptional activity in murine JB6 Cl41 cells. We showed that compounds 1–3 demonstrate anticancer activity in THP-1, HeLa, SNU-C4, SK-MEL-28, and MDA-MB-231 human cancer cell lines. Additionally, all compounds were found to prevent EGF-induced neoplastic transformation of murine JB6 Cl41 cells. Nuclear factors AP-1, NF-κB, and p53 are involved in the cellular response to high and nontoxic concentrations of aaptamine alkaloids 1–3. Furthermore, inhibition of EGF-induced JB6 cell transformation, which is exerted by the compounds 1–3 at low nontoxic concentrations of 0.7–2.1 μM, cannot be explained by activation of AP-1 and NF-κB.
1. Introduction
Aaptamines are a group of bioactive benzo[de][1,6]-naphthyridine alkaloids, initially isolated from marine sponges mostly belonging to the genus Aaptos. These compounds have been found to possess a variety of biological activities (for review see [1]). Among those, an anticancer effect has been the most frequently reported for aaptamines, although the underlying mechanism is poorly understood. Aaptamine has been shown to have DNA intercalating activity [2] as well as the ability to induce a p21-mediated (but p53-independent) G2/M cell cycle arrest [3–5]. Our recently reported global proteome screen of proteins differentially regulated upon treatment with aaptamine (1) and its derivatives 2, 3 in the human embryonal carcinoma cell lines NT2 and NT2-R, a cisplatin-resistant subline, uncovered several possible protein targets of these alkaloids [5, 6]. In the present study, the effects of the alkaloids 1–3 (Figure 1) on AP-1, NF-κB, and p53 transcriptional activity were investigated in order to further explore the mechanism of action of these compounds [7]. As a result, we present a more detailed picture of the biological action of aaptamines, extending the knowledge gained from previous research [5–11].
Figure 1.
Structures of aaptamine (1), demethyl(oxy)aaptamine (2), and isoaaptamine (3).
2. Materials and Methods
2.1. Reagents
Aaptamine compounds 1–3 were isolated from the marine sponge Aaptos sp. as described before [7, 8]. Epidermal growth factor (EGF) was purchased from Collaborative Research (Bedford, MA, USA), the Cell Titer 96 Aqueous One Solution Reagent [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt (MTS)] Kit was purchased from Promega (Madison, WI, USA), and D-luciferin was purchased from Anaspec (Waddinxveen, The Netherlands).
2.2. Cell Culture
The murine epidermal cell line JB6 P+ Cl41 and its stable transfectants JB6-Luc AP-1, JB6-Luc NF-κB, or JB6-Luc p53 (PG-13) cells were cultured at 37°C and 5% CO2 in MEM, containing 5% FBS, 2 mM L-glutamine, and 1% penicillin/streptomycin (Invitrogen, Paisley, UK). The human cancer cell lines HeLa (cervical carcinoma), SNU-C4 (colon cancer), and THP-1 (monocytic leukemia) were cultured at 37°C and 5% CO2 in RPMI medium containing 10% FBS, 2 mM L-glutamine, and 1% penicillin/streptomycin. The human cancer cell lines MDA-MB-231 (breast cancer) and SK-MEL-28 (melanoma) were cultured at 37°C and 5% CO2 in DMEM medium containing 10% FBS, 2 mM L-glutamine, and 1% penicillin/streptomycin. The THP-1 cell line was cultured in suspension; other cell lines were cultured in monolayers. JB6 cell lines were kindly provided by Dr. Zigang Dong, Hormel Institute, University of Minnesota, MN, USA. The cancer cell lines were purchased from the ATCC collection. Information regarding the genetic background of these cell lines is available online at the ATCC website.
2.3. Cytotoxicity Assay (MTS Test)
The effect of the substances on cell viability was evaluated using the MTS test [12]. The cells were preincubated overnight in 96-well plates (6 × 103 per well), 100 μL/well for adherent cells, or 50 μL/well for suspension (THP-1 cells). Then, the medium was replaced with fresh medium containing the substances at various concentrations in a total volume of 100 μL/well for adherent cells, and for suspension cells, 50 μL/well of fresh substance-containing medium was added and the cells were incubated for 24 h. Then 20 μL of Cell Titer 96 Aqueous One Solution Reagent was added into each well, and MTS reduction was measured 2 h later spectrophotometrically at 492 and 690 nm as background using μQuant equipment (Bio-Tek Instruments, Winooski, VT, USA). Results are represented as IC50 of the substances against corresponding untreated cells.
2.4. Anchorage-Independent Neoplastic Transformation Assay
The cancer preventive effect of aaptamine derivatives was evaluated using an anchorage-independent neoplastic transformation assay, as described previously [13]. EGF (10 ng/mL) was used to induce neoplastic transformation of JB6 P+ Cl41 cells. JB6 P+ Cl41 cells (8 × 103 cells/mL) were treated in 6-well plates with various concentrations of the substances in 1 mL of 0.33% basal medium Eagle- (BME-) agar containing 10% FBS over 3 mL of 0.5% BME-agar containing 10% FBS and various concentrations of the substances. The plates were incubated at 37°C in a humidified atmosphere with 5% CO2 for 1 week, before cell colonies were scored using an Olympus CKX31 inverted research microscope (Olympus, Center Valley, PA, USA). The ability of the substances to inhibit neoplastic transformation of JB6 P+ Cl41 cells is represented as concentration-dependent correlation of number of cell colonies.
2.5. Determination of the Effect of the Substances on the Basal Transcriptional Activity of AP-1, NF-κB, or p53 Nuclear Factors
The effects of the substances on the basal transcriptional activities of AP-1, NF-κB, or p53 nuclear factor were evaluated using JB6 Cl41 cell lines stably expressing a luciferase reporter gene controlled by an AP-1-, NF-κB-, or p53-DNA binding sequence, as described previously [14]. Briefly, cells were preincubated overnight in 96-well plates (20 × 103 cells/well) in 100 μL/well of culture medium. Then the medium was replaced with fresh medium containing different concentrations of the substances tested. After incubation for 6 h or 24 h, cell viability and the effect on the transcriptional activities of the nuclear factors were determined. To determine the transcriptional activities, cells were lysed for 1 h at RT with lysis buffer (0.1 M PBS (pH 7.8), 1% Triton X-100, 1 mM DTT, 2 mM EDTA). Then, 50 μL of lysate from each well was transferred into a luminescent analysis plate, and luciferase activity was measured using luciferase assay buffer (100 μL/well) (0.47 mM D-luciferin, 20 mM Tricin, 1.07 mM (MgCO3)4 × Mg(OH)2 × 5H2O, 2.67 mM MgSO4 × 7H2O, 33.3 mM DTT, 0.53 mM ATP, 0.27 mM CoA, and 0.1 mM EDTA (pH 7.8)) and the Luminoscan Ascent Type 392 microplate reader (Labsystems, Helsinki, Finland). The results are presented as a concentration-dependent correlation of transcriptional activity of AP-1, NF-κB, or p53 nuclear factors.
2.6. Apoptosis Assay
The onset of early and late apoptosis was analyzed by flow cytometry using Annexin V-FITC and propidium iodide (PI) double staining. 1 × 106 cells per 10 cm dish containing 10% FBS-RPMI were treated with various concentrations of substances 1–3 for 24 hours. After incubation, cells were washed with PBS by centrifugation at 1000 rpm for 5 min and processed for detection of apoptosis using Annexin V-FITC and PI staining according to the manufacturer's protocol. In brief, 1 × 105–5 × 105 cells were resuspended in 500 μL of 1× binding buffer (Annexin V-FITC Apoptosis Detection Kit). Then, 5 μL of Annexin V-FITC and 5 μL of PI were added, and the cells were incubated at room temperature for 15 min in the dark and were analyzed by flow cytometry.
3. Results
3.1. Aaptamines Alter AP-1-, NF-κB-, and p53-Dependent Transcriptional Activity in JB6 Cl41 Cells
The effects of aaptamines 1–3 on the transcriptional activities of the nuclear factors AP-1, NF-κB, and p53 were examined by using the luciferase assay and JB6 Cl41 cells stably expressing a luciferase reporter gene controlled by AP-1, NF-κB, or p53 DNA binding sequences. The short-term effect of aaptamines on the transcriptional activity after 6 h of treatment and the long-term effect after 24 h of treatment were examined. Results are shown in Figure 2 as concentration-dependent correlations of transcriptional activities (as percentage of untreated control cells). Unexpectedly, we found that aaptamine alkaloids 1–3 consistantly activate AP-1- and NF-κB-dependent transcriptional activity at nontoxic concentrations after both 6 h and 24 h of treatment (Figure 2). The observed effects were similar to those for cisplatin (data not shown), a widely used anticancer cytotoxic drug, which also activates AP-1- and NF-κB-dependent transcriptional activity at noncytotoxic concentration in JB6 Cl41 cells.
Figure 2.
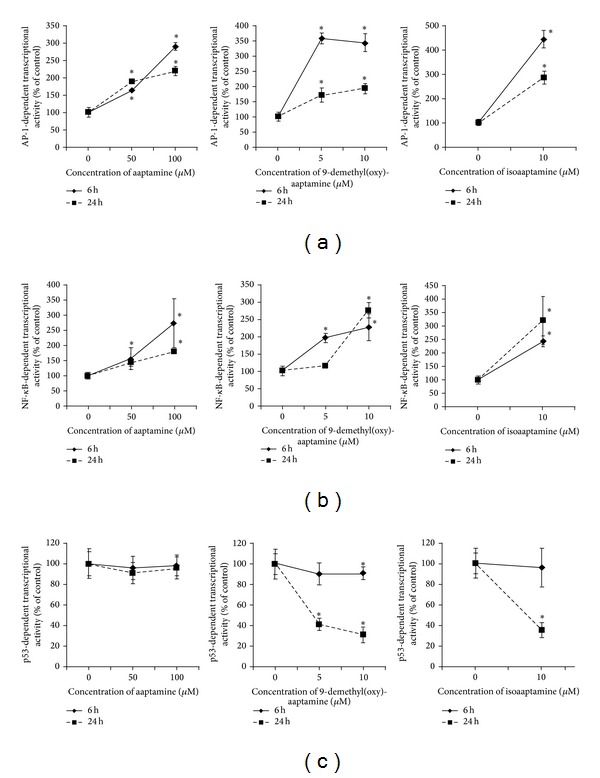
Effects of aaptamines 1–3 on the transcriptional activity of AP-1 (a), NF-κB (b), or p53 (c) in JB6 Cl41 cells stably expressing a luciferase reporter gene controlled by AP-1, NF-κB, or p53 DNA binding sequences, respectively. Cells were treated with the indicated concentrations of the substances for 6 h or 24 h. All experiments were performed in triplicate and repeated at least two times. “∗”-P < 0.05, statistically significant differences between treated and untreated control cells (Student's t-test).
Neither aaptamine (1) nor demethyl(oxy)aaptamine (2) or isoaaptamine (3) activated p53-dependent transcriptional activity. Moreover, derivatives 2 and 3 as well as cisplatin downregulated p53 transcriptional activity at noncytotoxic concentrations. In fact, this finding is in line with previous observations suggesting p53-independent cell cycle arrest in aaptamine treated cells at noncytotoxic concentrations of the drug [3–5]. Therefore, we speculate that apoptosis induced by demethyl(oxy)aaptamine (2) and isoaaptamine (3) [8] is not p53-dependent, similar to the situation reported after treatment of fibroblasts with cisplatin [15], although additional experiments are necessary to confirm this assumption.
In addition, we carried out similar experiments using a number of significantly lower concentrations (0.25; 0.5; 1.0; 2.0 μM) of the substances 1–3. Neither aaptamine (1) nor demethyl(oxy)aaptamine (2) or isoaaptamine (3) affects AP-1, NF-κB, or p53-dependent transcriptional activities in this range of low, nontoxic concentrations.
3.2. Analysis of Anticancer Activity of Aaptamines
The effects of the aaptamines 1–3 on the viability of five human cancer cell lines were studied using the MTS assay. It was shown that demethyl(oxy)aaptamine (2) and isoaaptamine (3) demonstrate higher anticancer activity than the mother compound aaptamine (1) (Table 1).
Table 1.
Anticancer activity of aaptamines 1–3 against several human cancer cell lines. Cells were treated with the indicated concentrations of the substances for 24 h. All experiments were performed in triplicate.
| Cancer type | Cell line | Compound (IC50, μM) | ||
|---|---|---|---|---|
| 1 | 2 | 3 | ||
| Monocytic leukemia | THP-1 | 161.3 ± 20.2 | 40.9 ± 9.9 | 32.2 ± 6.8 |
| Cervical carcinoma | HeLa | 151.1 ± 10.8 | 18.6 ± 1.7 | 50.7 ± 3.6 |
| Colon cancer | SNU-C4 | 267 ± 24.4 | 22.3 ± 6.9 | 35.8 ± 5.8 |
| Melanoma | SK-MEL-28 | 156.5 ± 6.7 | 35.0 ± 2.2 | 70.3 ± 3.3 |
| Breast cancer | MDA-MB-231 | 147.2 ± 3.9 | 9.1 ± 1.4 | 10.6 ± 2.8 |
3.3. Aaptamines Induce Apoptosis in THP-1 Human Leukemia Cells
To study whether aaptamine alkaloids 1–3 induce apoptosis in human cancer cells, we analysed THP-1 cells by flow cytometry after treatment with the compounds. The results show that aaptamine, demethyl(oxy)aaptamine, and isoaaptamine induce apoptosis in THP-1 cells in a dose-dependent manner (Figure 3). In these experiments, aaptamine (1) was also less active than the other two alkaloids 2 and 3.
Figure 3.
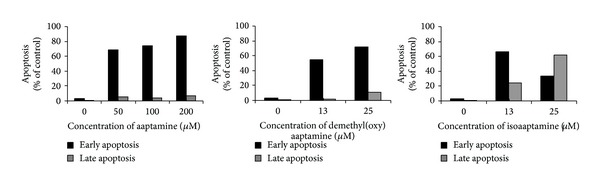
The induction of apoptosis by aaptamines 1–3 in THP-1 human cancer cells. Cells were treated with the indicated concentrations of the substances for 24 h.
3.4. Aaptamines Prevent EGF-Induced Transformation of JB6 P+ Cl41 Cells
The ability of the aaptamine alkaloids 1–3 to prevent EGF-induced neoplastic transformation and colony formation of murine epithelial JB6 P+ Cl41 cells was studied using anchorage independent soft agar assay.
This clone of JB6 cells is sensitive (P+) to tumor promoters like EGF or 12-O-tetradecanoylphorbol-13-acetate (TPA) and shows neoplastic transformation and anchorage independent colony formation upon stimulation with these agents. As shown in Figure 4, the substances examined were able to inhibit EGF-induced neoplastic transformation and colony formation of JB6 P+ Cl41 cells at low, noncytotoxic concentrations. Demethyl(oxy)aaptamine (2) possessed the strongest activity in preventing colony formation, INCC50 = 0.7 μM, whereas both aaptamine and isoaaptamine showed INCC50 = 2.1 μM (Figure 4).
Figure 4.
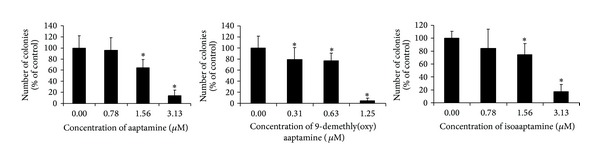
Effects of aaptamines 1–3 on the EGF-induced neoplastic transformation and colony formation of murine epithelial JB6 P+ Cl41 cells. Cells in a soft agar were treated with the indicated concentrations of the substances for one week. All experiments were performed in triplicate and repeated at least two times. “∗”-P < 0.05, statistically significant differences between treated and untreated control cells (Student's t-test).
4. Discussion
The transcription factors activator protein-1 (AP-1) and nuclear factor kappa B (NF-κB) are strongly involved in regulation of a wide range of cellular processes, including cell migration, proliferation, differentiation, inflammation, survival, and immunity [16–22].
Both nuclear factors are implicated not only in cell transformation and tumor promotion, but also in the induction of apoptosis and tumor suppression [16, 21, 23–33]. Some of the AP-1 proteins, such as Jun-B and c-Fos, were shown to have tumor-suppressor activity both in vitro and in vivo [34, 35]. Activation of another AP-1 protein, c-Jun, is required for the induction of Fas L-mediated apoptosis in PC12 and human leukemia HL-60 cells [36, 37]. Transactivated AP-1 protein inhibits proliferation of activated T cells [38]. Activation of both AP-1 and NF-κB nuclear factors is necessary for apoptosis by DNA damaging agents and ceramide in T lymphocytes and Jurkat T cells [31, 32]. NF-κB activation is required for apoptosis in fibrocystin/polyductin-depleted kidney epithelial cells [33]. One member of the AP-1 protein family, activating transcription factor 2 (ATF2), has tumor suppressor activities in nonmalignant skin tumors and breast cancer [39]. The balance between AP-1 family members, c-Jun, and ATF2 governs the choice between differentiation and apoptosis in PC12 cells [40]. The ultimate fate of the cells relies on the relative abundance of AP-1 or NF-κB complexes, their compositions, cell type, and cellular environment [41]. It was reported that some cancer preventive and therapeutic compounds, as well as DNA damaging agents, including those of marine origin, can induce AP-1 and/or NF-κB activities. For example, anticancer drug vinblastine, cancer preventive flavonoids kaempferol and genistein, anti-inflammatory drug tolfenamic acid, and marine alkaloids 3- and 10-bromofascaplysins all induce AP-1 activity [31, 42–46]. Marine compound 3-demethylubiquinone Q2 from ascidian Aplidium glabrum and its synthetic analogs, as well as the cancer preventive terpenoid dactylone, induce AP-1 and NF-κB and at the same time inhibit p53-dependent transcriptional activities [47–50].
Our investigations demonstrate that aaptamine alkaloids 1–3 induce AP-1 and NF-κB- dependent transcriptional activity at high nontoxic concentrations (100% viable cells) (Figure 2). For aaptamine (1), such concentrations are 50–100 μM; for 9-demethyl(oxy)aaptamine (2) -5–10 μM; and for isoaaptamine (3) about 10 μM. On the other hand, it was shown that neither aaptamine (1) nor demethyl(oxy)aaptamine (2) or isoaaptamine (3) affects the AP-1, NF-κB, or p53-dependent transcriptional activity at low nontoxic concentrations of 0.25–2.0 μM.
As was also demonstrated, aaptamines 1–3 show inhibition of anchorage-independent EGF-induced JB6 cell transformation and colony formation in soft agar at low nontoxic concentrations (Figure 3) of 0.7–2.1 μM. Therefore, inhibition of transformation of JB6 cells by aaptamines 1–3 cannot be explained by the induction of AP-1 and NF-κB-dependent transcriptional activity. Therefore, the molecular mechanisms underlying the cancer preventive effects of aaptamine and its derivatives at low nontoxic concentrations still remain unknown and await further investigations.
We showed that aaptamine (1) and its derivatives 2, 3 demonstrate anticancer effects against five human tumor cell lines. The IC50 for aaptamine is about 150 μM, and for alkaloids 2, 3 from 10 to 70 μM. At similar concentrations, these substances induced apoptosis in THP-1 human leukemia cells. Therefore, the anticancer effect of aaptamine, 9-demethyl(oxy)aaptamine, and isoaaptamine, can be at least in part explained by the induction of classical apoptosis.
5. Conclusions
Our study results indicate that the nuclear factors AP-1, NF-κB, and p53 are involved in the cellular response following treatment with high nontoxic (but not with low nontoxic) concentrations of aaptamine alkaloids 1–3. It was also found that aaptamine (1) at high nontoxic concentrations exerts biological action independently of p53-dependent transcriptional activation, whereas aaptamine analogues 2 and 3 inhibited p53 activation. We also provide evidence for cancer preventive activity of all aaptamines, which is exerted at low nontoxic concentrations and therefore independently of AP-1 and NF-κB activation.
Acknowledgments
The study was supported by the Program of the Presidium of RAS “Molecular and Cell Biology” (Grant 12-IP6-11), Grant no. 13-03-00986 from the RFBR, and Grant of President of Russia no. 148.2014.4 supporting leading Russian scientific schools, by the Grant of President of Russian Federation MK-6019.2014.4. The authors are grateful to Professor Zigang Dong (Hormel Institute of Minnesota University, USA) who kindly donated the JB6 cell lines, which were used in the present study.
Conflict of Interests
The authors declare no conflict of interests.
Authors' Contribution
Gunhild Keller-von Amsberg and Friedemann Honecker contributed equally to this work.
References
- 1.Larghi EL, Bohn ML, Kaufman TS. Aaptamine and related products. Their isolation, chemical syntheses, and biological activity. Tetrahedron. 2009;65(22):4257–4282. [Google Scholar]
- 2.Bowling JJ, Pennaka HK, Ivey K, et al. Antiviral and anticancer optimization studies of the DNA-binding marine natural product aaptamine. Chemical Biology and Drug Design. 2008;71(3):205–215. doi: 10.1111/j.1747-0285.2008.00628.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aoki S, Kong D, Suna H, et al. Aaptamine, a spongean alkaloid, activates p21 promoter in a p53-independent manner. Biochemical and Biophysical Research Communications. 2006;342(1):101–106. doi: 10.1016/j.bbrc.2006.01.119. [DOI] [PubMed] [Google Scholar]
- 4.Jin MH, Zhao WN, Zhang YW, Kobayashi M, Duan HQ, Kong DX. Antiproliferative effect of aaptamine on human chronic myeloid leukemia K562 cells. International Journal of Molecular Sciences. 2011;12(11):7352–7359. doi: 10.3390/ijms12117352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dyshlovoy SA, Naeth I, Venz S, et al. Proteomic profiling of germ cell cancer cells treated with aaptamine, a marine alkaloid with antiproliferative activity. Journal of Proteome Research. 2012;11(4):2316–2330. doi: 10.1021/pr300170p. [DOI] [PubMed] [Google Scholar]
- 6.Dyshlovoy SA, Venz S, Shubina LK, et al. Activity of aaptamine and two derivatives, demethyloxyaaptamine and isoaaptamine, in cisplatin-resistant germ cell cancer. Journal of Proteomics. 2014;96:223–239. doi: 10.1016/j.jprot.2013.11.009. [DOI] [PubMed] [Google Scholar]
- 7.Shubina LK, Kalinovsky AI, Fedorov SN, et al. Aaptamine alkaloids from the Vietnamese sponge Aaptos sp. Natural Product Communications. 2009;4(8):1085–1088. [PubMed] [Google Scholar]
- 8.Shubina LK, Makarieva TN, Dyshlovoy SA, Fedorov SN, Dmitrenok PS, Stonik VA. Three new aaptamines from the marine sponge Aaptos sp. and their proapoptotic properties. Natural Product Communications. 2010;5(12):1881–1884. [PubMed] [Google Scholar]
- 9.Dyshlovoy SA, Fedorov S, Shubina L, Honecker F, Stonik V. Anticancer activity of aaptamine and its derivatives isolated from marine Vietnamese sponge Aaptos sp. Annals of Oncology. 2011;22:p. 33. [Google Scholar]
- 10.Dyshlovoy SA, Naeth I, Venz S, et al. Aaptamine, demethyloxyaaptamine, and isoaaptamine: a proteomic- based screening of protein targets in cisplatin-resistant tumor cells. Annals of Oncology. 2012;23:28–28. [Google Scholar]
- 11.Dyshlovoy SA, Naeth I, Venz S, et al. Proteomic-based screening of protein targets of aaptamine, a marine alkaloid with antiproliferative activity. Annals of Oncology. 2012;23:p. 32. [Google Scholar]
- 12.Baltrop JA, Owen TC, Cory AH, Cory JG. 5-(3-carboxymethoxyphenyl)-2-(4,5-dimethylthiazolyl)-3- (4-sulfophenyl)tetrazolium, inner salt (MTS) and related analogs of 3-(4,5-dimethylthiazolyl)-2,5-diphenyltetrazolium bromide (MTT) reducing to purple water-soluble formazans as cell-viability indicators. Bioorganic and Medicinal Chemistry Letters. 1991;1(11):611–614. [Google Scholar]
- 13.Fedorov SN, Dyshlovoy SA, Shubina LK, Guzii AG, Kuzmich AS, Makarieva TN. C11 cyclopentenone from the ascidian Diplosoma sp. prevents epidermal growth factor-induced transformation of JB6 cells. Drugs and Therapy Studies. 2012;2(article e4) [Google Scholar]
- 14.Fedorov SN, Krasokhin VB, Shubina LK, Dyshlovoy SA, Nam NH, Minh CV. The extracts of some marine invertebrates and algae collected off the coast waters of vietnam induce the inhibitory effects on the activator protein-1 transcriptional activity in JB6 Cl41 cells. Journal of Chemistry. 2013;2013:6 pages.896709 [Google Scholar]
- 15.Mckay BC, Becerril C, Ljungman M. P53 plays a protective role against UV- and cisplatin-induced apoptosis in transcription-coupled repair proficient fibroblasts. Oncogene. 2001;20(46):6805–6808. doi: 10.1038/sj.onc.1204901. [DOI] [PubMed] [Google Scholar]
- 16.Eferl R, Wagner EF. AP-1: a double-edged sword in tumorigenesis. Nature Reviews Cancer. 2003;3(11):859–868. doi: 10.1038/nrc1209. [DOI] [PubMed] [Google Scholar]
- 17.Young MR, Li JJ, Rincón M, et al. Transgenic mice demonstrate AP-1 (activator protein-1) transactivation is required for tumor promotion. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(17):9827–9832. doi: 10.1073/pnas.96.17.9827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bernstein LR, Colburn NH. AP1/jun function is differentially induced in promotion-sensitive and resistant JB6 cells. Science. 1989;244(4904):566–569. doi: 10.1126/science.2541502. [DOI] [PubMed] [Google Scholar]
- 19.Tulchinsky E. Fos family members: regulation, structure and role in oncogenic transformation. Histology and Histopathology. 2000;15(3):921–928. doi: 10.14670/HH-15.921. [DOI] [PubMed] [Google Scholar]
- 20.Foletta VC, Segal DH, Cohen DR. Transcriptional regulation in the immune system: all roads lead to AP-1. Journal of Leukocyte Biology. 1998;63(2):139–152. doi: 10.1002/jlb.63.2.139. [DOI] [PubMed] [Google Scholar]
- 21.Shen H-M, Tergaonkar V. NFκB signaling in carcinogenesis and as a potential molecular target for cancer therapy. Apoptosis. 2009;14(4):348–363. doi: 10.1007/s10495-009-0315-0. [DOI] [PubMed] [Google Scholar]
- 22.Oeckinghaus A, Ghosh S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harbor Perspectives in Biology. 2009;1(4) doi: 10.1101/cshperspect.a000034.a000034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shaulian E, Karin M. AP-1 in cell proliferation and survival. Oncogene. 2001;20(19):2390–2400. doi: 10.1038/sj.onc.1204383. [DOI] [PubMed] [Google Scholar]
- 24.Shaulian E. AP-1 - The Jun proteins: oncogenes or tumor suppressors in disguise? Cellular Signalling. 2010;22(6):894–899. doi: 10.1016/j.cellsig.2009.12.008. [DOI] [PubMed] [Google Scholar]
- 25.Ameyar M, Wisniewska M, Weitzman JB. A role for AP-1 in apoptosis: the case for and against. Biochimie. 2003;85(8):747–752. doi: 10.1016/j.biochi.2003.09.006. [DOI] [PubMed] [Google Scholar]
- 26.Liss AS, Tiwari R, Kralova J, Bose HR., Jr. Cell transformation by v-Rel reveals distinct roles of AP-1 family members in Rel/NF-B oncogenesis. Oncogene. 2010;29(35):4925–4937. doi: 10.1038/onc.2010.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bhoumik A, Ronai Z. ATF2: a transcription factor that elicits oncogenic or tumor suppressor activities. Cell Cycle. 2008;7(15):2341–2345. doi: 10.4161/cc.6388. [DOI] [PubMed] [Google Scholar]
- 28.Sen R, Smale ST. Selectivity of the NF-{kappa}B response. Cold Spring Harbor Perspectives in Biology. 2010;2(4) doi: 10.1101/cshperspect.a000257.a000257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cadoret A, Bertrand F, Baron-Delage S, et al. Down-regulation of NF-κB activity and NF-κB p65 subunit expression by ras and polyoma middle T oncogenes in human colonic Caco-2 cells. Oncogene. 1997;14(13):1589–1600. doi: 10.1038/sj.onc.1200992. [DOI] [PubMed] [Google Scholar]
- 30.Kucharczak J, Simmons MJ, Fan Y, Gélinas C. To be, or not to be: NF-κB is the answer: role of Rel/NF-κB in the regulation of apoptosis. Oncogene. 2003;22(56):8961–8982. doi: 10.1038/sj.onc.1207230. [DOI] [PubMed] [Google Scholar]
- 31.Kasibhatla S, Brunner T, Genestier L, Echeverri F, Mahboubi A, Green DR. DNA damaging agents induce expression of Fas ligand and subsequent apoptosis in T lymphocytes via the activation of NF-κB and AP-1. Molecular Cell. 1998;1(4):543–551. doi: 10.1016/s1097-2765(00)80054-4. [DOI] [PubMed] [Google Scholar]
- 32.Manna SK, Sah NK, Aggarwal BB. Protein tyrosine kinase p56(lck) is required for ceramide-induced but not tumor necrosis factor-induced activation of NF-κB, AP-1, JNK, and apoptosis. The Journal of Biological Chemistry. 2000;275(18):13297–13306. doi: 10.1074/jbc.275.18.13297. [DOI] [PubMed] [Google Scholar]
- 33.Mangolini A, Bogo M, Durante C, et al. NF-κB activation is required for apoptosis in fibrocystin/polyductin-depleted kidney epithelial cells. Apoptosis. 2010;15(1):94–104. doi: 10.1007/s10495-009-0426-7. [DOI] [PubMed] [Google Scholar]
- 34.Chiu R, Angel P, Karin M. Jun-B differs in its biological properties from, and is a negative regulator of, c-Jun. Cell. 1989;59(6):979–986. doi: 10.1016/0092-8674(89)90754-x. [DOI] [PubMed] [Google Scholar]
- 35.Passegué E, Jochum W, Schorpp-Kistner M, Möhle-Steinlein U, Wagner EF. Chronic myeloid leukemia with increased granulocyte progenitors in mice lacking JunB expression in the myeloid lineage. Cell. 2001;104(1):21–32. doi: 10.1016/s0092-8674(01)00188-x. [DOI] [PubMed] [Google Scholar]
- 36.Le-Niculescu H, Bonfoco E, Kasuya Y, Claret F, Green DR, Karin M. Withdrawal of survival factors results in activation of the JNK pathway in neuronal cells leading to Fas ligand induction and cell death. Molecular and Cellular Biology. 1999;19(1):751–763. doi: 10.1128/mcb.19.1.751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kondo T, Matsuda T, Kitano T, et al. Role of c-jun expression increased by heat shock- and ceramide-activated caspase-3 in HL-60 cell apoptosis: possible involvement of ceramide in heat shock-induced apoptosis. The Journal of Biological Chemistry. 2000;275(11):7668–7676. doi: 10.1074/jbc.275.11.7668. [DOI] [PubMed] [Google Scholar]
- 38.Yan Y, Zhang GX, Williams MS, et al. TCR stimulation upregulates MS4a4B expression through induction of AP-1 transcription factor during T cell activation. Molecular Immunology. 2012;52(2):71–78. doi: 10.1016/j.molimm.2012.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lau E, Ronai ZA. ATF2 - at the crossroad of nuclear and cytosolic functions. Journal of Cell Science. 2012;125(12):2815–2824. doi: 10.1242/jcs.095000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Leppä S, Eriksson M, Saffrich R, Ansorge W, Bohmann D. Complex functions of AP-1 transcription factors in differentiation and survival of PC12 cells. Molecular and Cellular Biology. 2001;21(13):4369–4378. doi: 10.1128/MCB.21.13.4369-4378.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gopalakrishnan A, Tony Kong A. Anticarcinogenesis by dietary phytochemicals: cytoprotection by Nrf2 in normal cells and cytotoxicity by modulation of transcription factors NF-κB and AP-1 in abnormal cancer cells. Food and Chemical Toxicology. 2008;46(4):1257–1270. doi: 10.1016/j.fct.2007.09.082. [DOI] [PubMed] [Google Scholar]
- 42.Berry A, Goodwin M, Moran CL, Chambers TC. AP-1 activation and altered AP-1 composition in association with increased phosphorylation and expression of specific Jun and Fos family proteins induced by vinblastine in KB-3 cells. Biochemical Pharmacology. 2001;62(5):581–591. doi: 10.1016/s0006-2952(01)00694-3. [DOI] [PubMed] [Google Scholar]
- 43.Fan M, Goodwin ME, Birrer MJ, Chambers TC. The c-Jun NH2-terminal protein kinase/AP-1 pathway is required for efficient apoptosis induced by vinblastine. Cancer Research. 2001;61(11):4450–4458. [PubMed] [Google Scholar]
- 44.Gopalakrishnan A, Xu C, Nair SS, Chen C, Hebbar V, Kong AT. Modulation of activator protein-1 (AP-1) and MAPK pathway by flavonoids in human prostate cancer PC3 cells. Archives of Pharmacal Research. 2006;29(8):633–644. doi: 10.1007/BF02968247. [DOI] [PubMed] [Google Scholar]
- 45.Lee S-H, Bahn JH, Whitlock NC, Baek SJ. Activating transcription factor 2 (ATF2) controls tolfenamic acid-induced ATF3 expression via MAP kinase pathways. Oncogene. 2010;29(37):5182–5192. doi: 10.1038/onc.2010.251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kuzmich AS, Fedorov SN, Shastina VV, et al. The anticancer activity of 3- and 10-bromofascaplysins is mediated by caspase-8, -9, -3-dependent apoptosis. Bioorganic and Medicinal Chemistry. 2010;18(11):3834–3840. doi: 10.1016/j.bmc.2010.04.043. [DOI] [PubMed] [Google Scholar]
- 47.Shubina LK, Fedorov SN, Radchenko OS, et al. Desmethylubiquinone Q2 from the Far-Eastern ascidian Aplidium glabrum: Structure and synthesis. Tetrahedron Letters. 2005;46(4):559–562. [Google Scholar]
- 48.Fedorov SN, Radchenko OS, Shubina LK, et al. Evaluation of cancer-preventive activity and structure-activity relationships of 3-demethylubiquinone Q2, isolated from the ascidian Aplidium glabrum, and its synthetic analogs. Pharmaceutical Research. 2006;23(1):70–81. doi: 10.1007/s11095-005-8813-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fedorov SN, Radchenko OS, Shubina LK, et al. Anticancer activity of 3-demethylubiquinone Q2. In vivo experiments and probable mechanism of action. Anticancer Research. 2008;28(2):927–932. [PubMed] [Google Scholar]
- 50.Fedorov SN, Shubina LK, Bode AM, Stonik VA, Dong ZG. Dactylone inhibits epidermal growth factor-induced transformation and phenotype expression of human cancer cells and induces G1-S arrest and apoptosis. Cancer Research. 2007;67(12):5914–5920. doi: 10.1158/0008-5472.CAN-06-3723. [DOI] [PubMed] [Google Scholar]