

Abstract
Schistosomiasis is a chronic inflammatory disease whose macrophages are involved in immunopathology modulation. Although P2X7 receptor signaling plays an important role in inflammatory responses mediated by macrophages, no reports have examined the role of P2X7 receptors in macrophage function during schistosomiasis. Thus, we evaluated P2X7 receptor function in peritoneal macrophages during schistosomiasis using an ATP-induced permeabilization assay and measurements of the intracellular Ca2+ concentration. ATP treatment induced significantly less permeabilization in macrophages from S. mansoni-infected mice than in control cells from uninfected animals. Furthermore, P2X7-mediated increases in intracellular Ca2+ levels were also reduced in macrophages from infected mice. TGF-β1 levels were increased in the peritoneal cavity of infected animals, and pretreatment of control macrophages with TGF-β1 reduced ATP-induced permeabilization, mimicking the effect of S. mansoni infection. Western blot and qRT-PCR data showed no difference in P2X7 protein and mRNA between uninfected, infected, and TGF-β1-treated groups. However, immunofluorescence analysis revealed reduced cell surface localization of P2X7 receptors in macrophages from infected and TGF-β1-treated mice compared to controls. Therefore, our data suggest that schistosomiasis reduces peritoneal macrophage P2X7 receptor signaling. This effect is likely due to the fact that infected mice have increased levels of TGF-β1, which reduces P2X7 receptor cell surface expression.
1. Introduction
Macrophages are plastic phagocytic cells that participate in innate and adaptive immunity, with important roles in the response against extra- and intracellular parasites, as well as in tissue homeostasis [1, 2]. These cells have complex functions during acute and chronic inflammation. In inflammatory foci, macrophages contribute to mounting specific immunological responses for host defense, and the plasticity of macrophage phenotypes is modulated by the cytokine profile [1].
Tissue damage during inflammation increases the extracellular levels of otherwise intracellular molecules, resulting in damage-associated molecular patterns (DAMPs) recognized by the immune system [3–5]. Extracellular nucleotides, such as the ATP released by dying cells and by activated immune cells during inflammation, are important danger signals involved in immune response [6], participating in both paracrine [7] and autocrine [8] signaling pathways. In inflammatory processes, high extracellular ATP levels generated by tissue damage or secretion are recognized by the immune system as a danger signal, activating purinergic P2 receptors that contribute to a proinflammatory response [9].
Purinergic P2 receptors can be classified as ionotropic ATP-gated (P2X) or G-protein coupled metabotropic (P2Y) [10]. Extracellular ATP (eATP) induces macrophage-mediated immune responses mainly through the activation of P2X7 receptors [11]. These receptors have a ubiquitous distribution, although the highest levels of receptor expression are observed in immune cells of monocyte/macrophage origin [12]. P2X7 receptor activation induces a myriad of intracellular events, including the production of nitric oxide (NO) and of reactive oxygen species (ROS) and the activation of phospholipase-D (PLD). These events are important for intracellular parasite killing, for the release of proinflammatory cytokines (such as IL-1β), and for inducing apoptosis [13, 14].
The expression and function of P2X7 receptors are regulated by both pro- and anti-inflammatory stimuli [15]. Initially, P2X7 receptor activation in macrophages opens a plasma membrane cation channel that allows a substantial efflux of K+, and influx of Ca2+, with later formation of pores permeable to large molecules [16, 17]. P2X receptor activation may also induce caspase-1 activation, interleukin IL-1β release, apoptosis, phagolysosomal fusion, and the elimination of intracellular pathogens [14, 18, 19]. In murine macrophages, the mature IL-1β secretion by the activation of P2X7 receptors is reduced by the incubation in a free-Ca2+ buffer [20]. Thus, murine macrophage release of mature IL-1β depends on intracellular Ca2+, confirming that Ca2+ signaling is essential for this P2X7 receptor function.
Schistosomiasis is a chronic inflammatory disease caused by Schistosoma mansoni and it represents the second most common tropical parasitic disease related to socioeconomical factors. While migrating through the vascular system of infected mammalian hosts, parasites evolve from the schistosomula migratory form into adult worms, causing endothelial cell activation, immunological responses, and tissue damage. The disease starts with a Th1-type immune response that gradually changes to a Th2 profile [21–24]. Previous data suggest that the Th2 “stage” begins four to six weeks after infection and is related to egg deposition by adult worms.
Macrophages play important roles in both phases of schistosomiasis [24]. In the early phase of the disease, macrophages act as immune system effectors cells killing schistosomula and promoting tissue repair [24]. After egg deposition, the immune response switches to a Th2 profile that is involved in the formation of liver and colonic granulomas and fibrosis. The eggs are laid in mesenteric microcirculation and they may reach peritoneal cavity where the granulomas formed contain mainly macrophages [25]. Although granulomas are important to limit egg-derived potential cytotoxic products, macrophages isolated, for example, from hepatic granulomas, produce lipids mediators and free radicals that are potentially destructive to host tissues [26]. Among cytokines involved in schistosomiasis, transforming growth factor-β1 (TGF-β1) is of particular interest, since high levels of this cytokine are released by peripheral blood mononuclear cells (PBMCs) from S. mansoni-infected mice [27]. TGF-β has an important role in immune modulation later during infection, limiting liver inflammation and favoring host survival [27, 28].
Recently, Bhardwaj and Skelly [29] showed that S. mansoni expresses P2X7 receptors-like molecules and also enzymes responsible for the clearance of extracellular ATP (ectonucleotidases), suggesting that purinergic signaling is conserved in these parasites and is important for the host-parasite interplay. However, the impact of the chronic inflammation trigged by schistosomiasis on macrophage P2X7 receptor function remains unknown.
Here, we evaluated P2X7 receptor function and expression in macrophage from S. mansoni-infected mice. Our data show that peritoneal macrophage P2X7 receptor function is attenuated during schistosomiasis and that this is associated with high peritoneal levels of TGF-β1, the important inflammatory mediator present in the chronic phase of the disease. We also show that TGF-β1 downregulates P2X function in vitro, mimicking the effect of S. mansoni infection.
2. Materials and Methods
2.1. Reagents and Antibodies
The following primary antibodies were used in this work: rabbit polyclonal anti-P2X7 receptors (APR-004 and APR-008; Alomone Labs, Israel); mouse monoclonal anti-β-actin (Santa Cruz Biotechnology, USA); rat anti-F4/80 (Biolegend, USA); rat anti-F4/80 FITC (AbD Serotec, USA). ATP, 3′-O-(4-benzoyl)-ATP (BzATP), PMSF, sodium orthovanadate, aprotinin, leupeptin, BSA, ionomycin, and EGTA were from Sigma Chemical Co. (USA). Fura-2-AM was from Molecular Probes (USA) and A740003 was from Tocris (USA). RPMI, foetal bovine serum (FBS), and penicillin/streptomycin solutions were from GIBCO BRL (USA). TGF-β1 was from R&D Systems (Minneapolis, MN, USA). Stock solutions were prepared in DMSO (2.5 mM Fura-2-AM and 10 mM A740003), water (10 mM BzATP), RPMI (5 ng/mL TGF-β1), or a buffered physiological saline solution (10 mM ATP). The highest concentration of solvent used was 0.1% (v/v). All other reagents used were of analytical grade.
2.2. Mice
Swiss, C57BL/6 (wild type), and P2X7 receptor knockout (P2X7 KO) male mice were used in all procedures. Swiss and C57BL/6 mice were obtained from the animal facility of the Paulo de Goes Microbiology Institute (Federal University of Rio de Janeiro, Rio de Janeiro, Brazil). P2X7 KO mice (originally from The Jackson Laboratory, USA, stock number 005576) were maintained in the transgenic animal house of the Federal University of Rio de Janeiro. All mice were kept under a light/dark cycle of 12/12 h and with access to water and food ad libitum.
All experiments were conducted in compliance with the ethical standards of our institution (Ethics Committee of the Federal University of Rio de Janeiro; approved under the licenses DFBC-ICB-011 and IBCCF 154) and following both the guidelines of the National Council on Experimental Animal Control (CONCEA, Brazil) and the Committee of Care and Use of Laboratory Animals (National Research Council, United States). All efforts were made to minimize both animals suffering and the number of animals used, on the basis of valid statistical evaluation.
2.3. Parasite and Infection
In this work, we used the BH strain of S. mansoni, obtained from infected Biomphalaria glabrata snails. Swiss, C57BL/6, and P2X7 KO mice (7-10 days old) were infected percutaneously with approximately 80 cercariae from both genders, for 8 min, as previously described [30]. Animals were used in experiments at least 45 days after infection (dpi) to allow for full establishment of the infection. Age-matched (60 to 80 days old) uninfected Swiss, C57BL/6, and P2X7 KO mice were used as controls. C57BL/6 and P2X7 KO mice survival was evaluated during infection.
2.4. Peritoneal Macrophages
To obtain mouse peritoneal macrophages, animals were anesthetized, sacrificed by cervical dislocation, and cleaned with 70% ethanol. The peritoneal cavity was washed with 5 mL of sterile phosphate buffered saline (PBS: 137 mM NaCl, 8.1 mM Na2HPO4, 1.5 mM NaH2PO4, and 2.7 mM KCl, pH 7.4), and the peritoneal exudate was collected and centrifuged at 350 ×g, for 5 min, at 4°C. The pellet was resuspended in 1 mL of PBS and used to perform total leukocyte counts (cell viability, as estimated by Trypan blue exclusion, was always ≥95%). For macrophages culture, cells were resuspended in 1 mL of RPMI-1640 medium containing 2 g/L sodium bicarbonate, 1 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin and plated in 35 mm Petri dishes or on 6-well plates. After 1 h of incubation at 37°C with 5% CO2, nonadherent cells were removed by vigorous washing, and RPMI-1640 medium with 10% heat-inactivated FBS was added to the cultures, which were kept at 37°C with 5% CO2 for 24 h until further use.
2.5. ATP-Induced Permeabilization Assay
To investigate macrophage responses to ATP, freshly harvested peritoneal macrophages (105 cells) from Swiss (uninfected or S. mansoni-infected) or C57BL/6 (wild type or P2X7 KO) mice were treated with 0.1, 0.5, or 1 mM ATP and 2.5 μM ethidium bromide (EB) in PBS (15 min; 37°C). EB was used as a tracer for P2X7 receptor activation (i.e., cell permeabilization) triggered by ATP. After incubation with ATP and EB, F4/80 positive cells were analyzed by flow cytometry (10,000 events/sample) using a FACScan system (BD Pharmingen, USA). Flow cytometry data analysis was performed using the WINMDI 2.9 software. In each histogram, a marker was added based on a gate of the F4/80 positive macrophages population, which limited the basal fluorescence. The specific ATP-induced permeabilization threshold was defined by comparison with the “baseline” profile of control cells not treated with ATP (i.e., incubated with EB only). The specific permeabilization is the percentage of EB positive cells after ATP stimulation in the F4/80 positive gated cells.
Alternatively, peritoneal macrophages were plated on 24-well plates (2 × 105 cells/well) and, after 24 h, treated with 5 ng/mL TGF-β1 or vehicle (RPMI), for 24 h, at 37°C and in 5% CO2. Macrophages were washed in PBS (pH 7.4) and then incubated with 1 mM ATP and 5 μM EB in buffer (145 mM NaCl, 5 mM KCl, 1 mM MgCl2, 1 mM CaCl, and 10 mM HEPES, pH 7.4) for 15 min at 37°C to evaluate the P2X7 receptor activation. The ratio between permeabilized (i.e., EB positive) and total cells was determined by direct counting of cells in five randomly chosen fields, using an Axiovert 100 microscope (Karl Zeiss, Oberkochen, Germany) equipped with an Olympus digital camera (Olympus American Inc., PA, USA). The specific ATP-induced permeabilization value (in %) was calculated by subtracting from the total % of permeabilized cells a “baseline” value corresponding to the % of permeabilized cells in control samples not treated with ATP (i.e., incubated with EB only).
2.6. Measurement of [Ca2+]i in Macrophages
Measurements of the intracellular Ca2+ concentration ([Ca2+]i) using Fura-2-AM were performed as previously described [31]. Briefly, macrophages were plated on glass bottom plates (MatTek, USA), at a density of 1 × 105 cells/plate, and maintained for 24 h at 37°C, with 5% CO2. Then, the medium was removed and the cells were incubated for 40 min with 2.5 μM Fura-2-AM at room temperature. Cells were subjected to alternate cycles of illumination with 340 nm and 380 nm excitation wavelengths, and the emission was measured at 500 nm. Cells were washed twice with buffered physiological solution (in mM: NaCl 140, KCl 5, MgCl2 1, CaCl2 2, glucose 5, and HEPES 5, pH 7.4) and then stimulated with 100 μM 3′-O-(4-benzoyl)-ATP (BzATP) (at 37°C) in the absence or in the presence of 50 nM of the P2X7 receptor antagonist A740003 (preincubated for 45 min, at 37°C). After the maximum effect had been achieved, cells were treated with Ca2+ ionophore (10 μM ionomycin) and then with 6 mM EGTA to determine the molar concentration of Ca2+ according to the Grynkiewicz equation [32].
2.7. Determination of TGF-β1 Levels
The estimation of TGF-β1 levels in mouse peritoneal washes was performed using an ELISA immunoassay kit (Peprotech, Rocky Hill, NJ, USA), according to manufacturer's instructions. For peritoneal wash collection, mice were euthanized with CO2 and the peritoneal cavity was washed with 1 mL of sterile PBS. Approximately 60% of the initial volume of PBS was recovered and centrifuged at 350 g for 5 min, at 4°C. Supernatants were collected and stored in liquid N2 until further use (to determine TGF-β1 content).
2.8. Confocal Microscopy
Peritoneal macrophages from control and infected mice (105 cells/well) were treated with 5 ng/mL TGF-β for 24 h, fixed with 4% paraformaldehyde and 4% sucrose for 10 min in PBS, washed, and incubated for 30 min with 50 mM ammonium chloride (pH 8.0). After three washes in PBS, samples were blocked in PBS with 10% FBS and 0.1% BSA for 30 min, washed twice in PBS, and incubated overnight with a rabbit polyclonal antibody that recognizes an extracellular epitope of P2X7 receptors (APR-008; Alomone Labs, Israel; Peptide KKGWMDPQSKGIQTGRC, corresponding to amino acid residues 136–152 of mouse P2X7 receptor; extracellular loop) diluted to 1 : 400 in PBS and 0.1% BSA. Then, cells were washed and incubated with a rat anti-F4/80 antibody diluted to 1 : 50 in PBS and 0.1% BSA for 4 h, at 4°C. Samples were then labelled with secondary anti-rabbit-Alexa 488 and anti-rat-Alexa 546 antibodies diluted to 1 : 500 in PBS and 0.1% BSA for 1 h at 4°C. The secondary antibodies were used in the absence of primary antibodies as negative controls. After labelling, cells were mounted on slides using Vectashield with DAPI (Vector, Burlingame, CA, USA) and examined on a TCS-SP5 AOBS confocal microscope (Leica Microsystems, Germany). Images were analyzed using the ImageJ software. Each cell micrograph was analyzed using ImageJ software (Rasband, W.S., ImageJ, US National Institutes of Health, Bethesda, Maryland, USA, http://imagej.nih.gov/ij/, 1997–2014). The region of interest (ROI) was delimited in each cell and the respective mean fluorescence intensity was measured based on the pixel intensity. 10 cells per image were evaluated and the fluorescent values were used for statistical analysis. Data were also displayed in an orthogonal slice view that shows the raw pixel intensity values found mutually in each of three perpendicular planes.
2.9. Western Blotting
Macrophages adhered to 6-well plates were washed with sterile PBS and lysed with 200 μL RIPA buffer (50 mM Tris-HCl, pH 7.4, containing 1% Nonidet P40, 0.25% sodium deoxycholate, 150 mM NaCl, 1 mM EDTA, 1 mM PMSF, 1 mM sodium orthovanadate, 1 mM NaF, 10 μg/mL aprotinin, and 10 μg/mL leupeptin). Cell lysates were incubated at 4°C for 30 min and centrifuged at 8,100 ×g, for 10 min, at 4°C; the pellets were discarded and the supernatant had their protein content determined by the Lowry method [33]. Samples were run on 10% SDS–PAGE gels (15 μg protein/lane) and then transferred to nitrocellulose membranes using a semidry transfer system. Membranes were blocked for 1 h in Tris-buffered saline (TBS) with 2% nonfat milk and then incubated overnight with rabbit anti-P2X7 receptors (APR-004; Alomone Labs, Israel; 1 : 200; Peptide (C)KIRKEFPKTQGQYSGFKYPY, corresponding to amino acid residues 576–595 of P2X7 receptor intracellular, C-terminus) or mouse monoclonal anti-β-actin (1 : 20.000) primary antibodies. After 3 washes in TBS-Tween, membranes were incubated with HRP-conjugated anti-rabbit or anti-mouse secondary antibody (1 : 1.000) for 1 h and developed using enhanced chemiluminescence (ECL). Ponceau staining and labeling with a monoclonal anti-β-actin antibody were used as internal controls for protein loading. Densitometry of protein bands was performed using the ImageJ software, and the values for relative amounts of protein were normalized to the β-actin loading control.
2.10. Quantitative Real Time PCR (qRT-PCR)
Peritoneal macrophages from uninfected Swiss mice (treated with 5 ng/mL TGF-β1 or with vehicle, for 24 h) or from S. mansoni-infected mice were used to isolate total RNA using the TRIzol reagent (Life Technologies), according to the manufacturer's instructions. Total RNA was quantified using an ND-1000 spectrophotometer (NanoDrop), and cDNA was synthesized from 500 ng of total RNA using the high capacity cDNA reverse transcription kit with RNase inhibitor (Invitrogen). The SYBR Select Master Mix (Applied Biosystems) was used for qRT-PCR, to detect double-stranded DNA synthesis. Reactions were carried out in a final volume of 10 μL, using 2 μL of diluted cDNA (1 : 10) and 300 nM of each of the reverse and forward primers. The following primers were used for qRT-PCR: for P2rx7, 5′ AATCGGTGTGTTTCCTTTGG 3′ (forward) and 5′ CCGGGTGACTTTGTTTGTCT 3′ (reverse); for Actb, 5′ TATGCCAACACAGTGCTGTCTGG 3′ (forward) and 5′ TACTCCTGCTTGCTGATCCACAT 3′ (reverse); and for Gapdh, 5′ GGTCATCCCAGAGCTGAACG 3′ (forward) and 5′ TTGCTGTTGAAGTCGCAGGA 3′ (reverse).
Reactions were performed in a 7500 Fast Real-Time System (Applied Biosystems), using the following PCR conditions: 5 min at 95°C, followed by 40 cycles of 15 s at 95°C, 35 s at 60°C, and 15 s at 72°C. At the end of cycling protocol, a melting-curve analysis (with fluorescence measurements from 60 to 99°C) was performed. Relative expression levels were determined using the Sequence Detection Software v.2.0.5 (Applied Biosystems), and the efficiency per sample was calculated using the LinRegPCR 11.0 software (manufacturer).β-Actin and Gapdh were used as internal controls to calculate relative P2rx7 mRNA levels, by the comparative threshold cycle (ΔΔCT) method with efficiency correction, using the mean qRT-PCR efficiency for each amplicon, as previously described [34, 35].
2.11. Statistical Analysis
Data were expressed as mean and SEM. Differences between two or more groups were analyzed by Student's t-test or one-way analysis of variance (ANOVA) followed by post hoc Newman-Keuls test, respectively, considering P < 0.05.
3. Results
3.1. P2X7 Receptor Function Is Reduced in Peritoneal Macrophages during Schistosomiasis
We used several approaches to examine the impact of the chronic inflammation triggered by S. mansoni infection on macrophage P2X7 receptor function. Activation of P2X7 receptors by ATP opens plasma membrane pores that allow molecules greater than 900 kDa (such as the fluorescent dye EB) to enter the cells [17, 36]. Thus, we used ATP-induced cell permeabilization (as evidenced by staining with EB) as a tool to compare the P2X7 receptor activation in peritoneal macrophage from infected and uninfected Swiss mice. Incubations with 0.1–1 mM ATP induced cell permeabilization in macrophages from uninfected mice; however, this effect was reduced in macrophages from infected mice (Figure 1(a)). ATP also induced permeabilization in macrophages from C57BL/6 wild type mice but, as expected, not in macrophages from P2X7 KO mice (Figure 1(b)).
Figure 1.
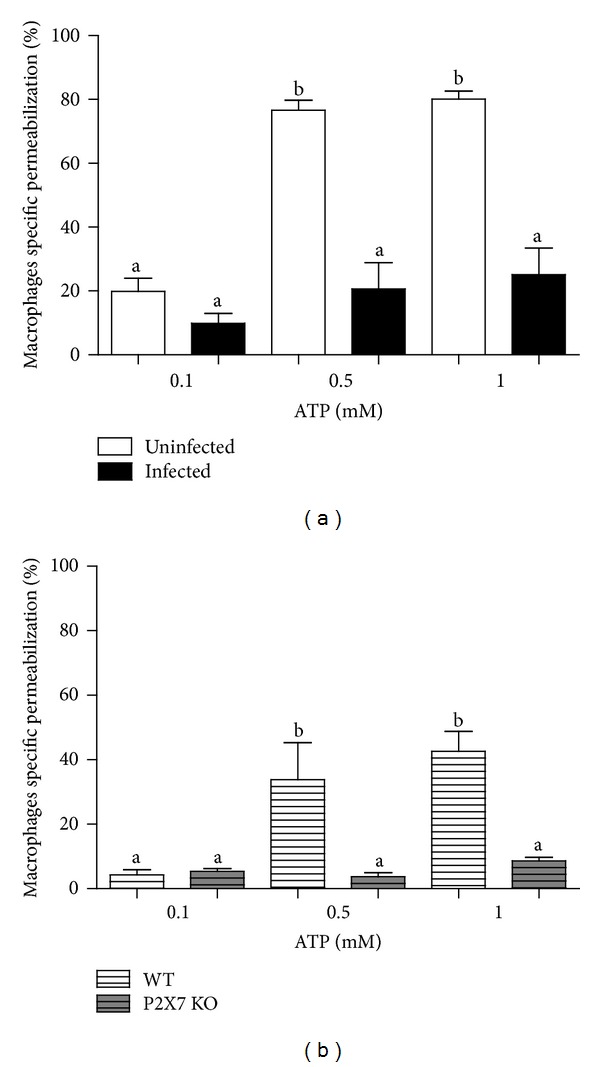
Evaluation of ATP-induced permeabilization in peritoneal macrophages, based on ethidium bromide (EB) uptake. (a) Peritoneal macrophages from S. mansoni-infected Swiss mice (black bars) were less sensitive to ATP-induced permeabilization than those from uninfected mice (white bars). (b) Peritoneal macrophages from C57BL/6 wild type mice (hatched white bars) respond to ATP-induced permeabilization unlike those from P2X7R KO mice (hatched gray bars). ATP-induced permeabilization values represent the % of EB uptake above basal levels by F4/80 positive cells and were expressed as mean and SEM. N = 12 (Swiss) and n = 6 − 9 (C57BL/6) replicates using different animals. (a) and (b) differ from each other with P < 0.001 (one-way ANOVA followed by post hoc Newman-Keuls test).
We also used the potent P2X7 agonist BzATP to verify if P2X7 receptor activation increased the cytosolic Ca2+ concentration in macrophages from infected and uninfected Swiss mice, since ATP-induced Ca2+ influx is a hallmark of P2X7 activation [36]. The basal levels of cytosolic Ca2+ (before BzATP addition) were similar in both groups (Figure 2(a)). After treatment with 100 μM BzATP, macrophages from uninfected mice showed a typical P2X7 receptors activation profile at 37°C, characterized by a pronounced biphasic increase in intracellular Ca2+ levels (Figure 2(a)) [36]. However, the P2X7 receptor-mediated Ca2+ influx was considerably less pronounced in macrophages from infected mice when compared to that observed in macrophages from uninfected mice (Figures 2(a) and 2(b)). We also assessed changes in intracellular Ca2+ in the presence of the selective P2X7 receptor antagonist A740003. Treatment with 50 nM A740003 did not alter basal intracellular Ca2+ levels, but it had a strong negative effect on the BzATP-induced Ca2+ influx, and this effect was similar in macrophages from both groups. These results confirmed that the increase in intracellular Ca2+ levels after addition of BzATP was due to P2X7 receptor activation. These data are in agreement with the reduction in ATP-induced permeabilization observed in macrophages from infected mice (Figure 1(a)) and suggest that P2X7 receptor signaling is downregulated during schistosomiasis.
Figure 2.
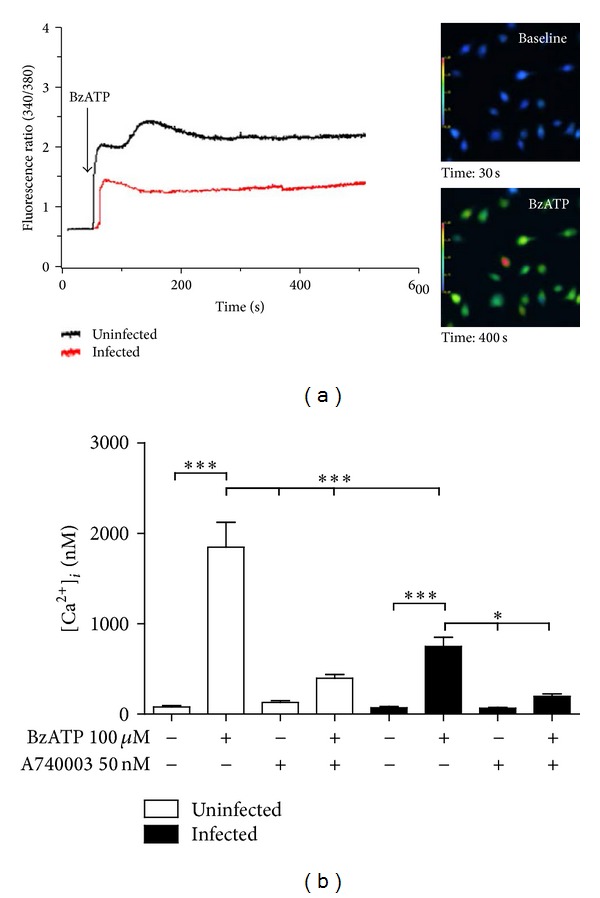
Intracellular Ca2+ measurement (using Fura-2-AM) in macrophages subjected to treatment with the P2X7 agonist BzATP and/or with the antagonist A740003. (a) Changes in intracellular Ca2+ levels in macrophages from control (black line) and S. mansoni-infected (red line) mice. Inset: representative images from macrophages (control) before (baseline) and after treatment with 100 μM BzATP. (b) Changes in intracellular Ca2+ levels ([Ca2+]i) in macrophages from control (white bars) or S. mansoni-infected (black bars) mice. Three plates were used for each condition/animal, 10 cells/plate were chosen randomly for imaging, and cells were obtained from three animals. Data were expressed as mean and SEM. *P < 0.05; ***P < 0.001 (one-way ANOVA followed by post hoc Newman-Keuls test).
3.2. S. Mansoni-Infected Mice Have Higher Levels of Peritoneal TGF-β1, Associated with Decreased P2X7-Dependent Macrophage Permeabilization
The anti-inflammatory cytokine TGF-β1 prevents P2X7 receptor upregulation in monocytes [37]. Moreover, in the chronic phase of schistosomiasis, there is a gradual increase of serum Th2 cytokines [21]. Indeed, we observed a significant increase in TGF-β1 levels in the peritoneal cavity of mice infected with S. mansoni (Figure 3).
Figure 3.
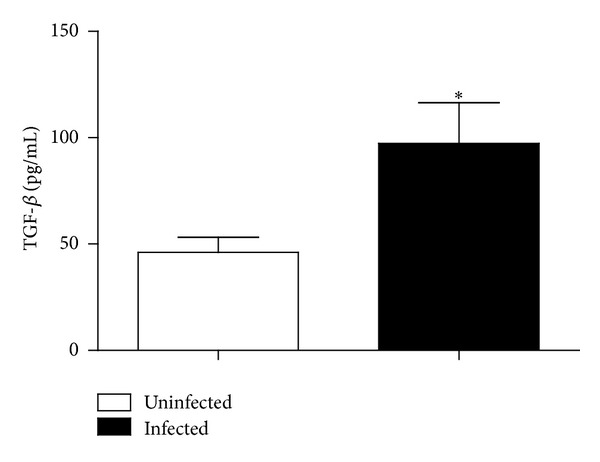
Peritoneal TGF-β1 levels in uninfected (white) and S. mansoni-infected mice (black). Data were expressed as mean and SEM. n = 6 (uninfected) or 4 (infected) mice. *P < 0.05 (Student's t-test).
To investigate if TGF-β1 could modulate P2X7 receptor function in peritoneal macrophages similar to that described for monocytes [37], we harvested peritoneal macrophages from uninfected animals and treated these cells with 5 ng/mL TGF-β1 for 24 h before assessing their sensitivity to ATP-induced permeabilization (using EB as a tracer). In the absence of ATP, TGF-β1 treatment did not induce cell permeabilization; however, TGF-β1 reduced the permeabilizing effect of 1 mM ATP by approximately 50%, when compared to controls treated with ATP only (Figure 4). We also performed the permeabilization after 48 h of TGF-β treatment and we observed that, even after 48 h, the P2X7-induced permeabilization was also reduced (data not shown). The analysis of the concentration-response curve using different concentrations of TGF-β showed that only the concentrations 5 ng/mL and 10 ng/mL were able to reduce the permeabilization intensity in macrophages (data not shown).
Figure 4.
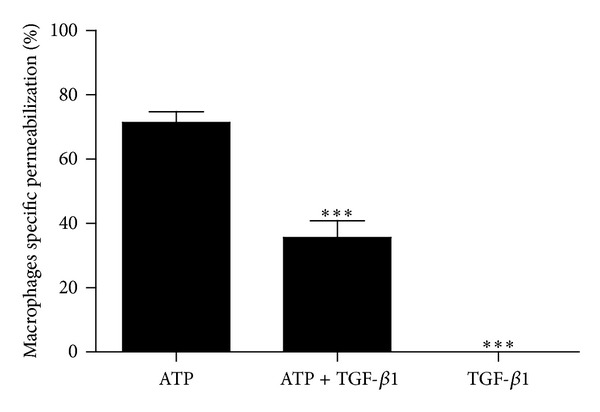
ATP-induced permeabilization assays (based on ethidium bromide uptake) by peritoneal F4/80 positive macrophages of uninfected mice treated with TGF-β1. Cells treated with 5 ng/mL TGF-β1 or vehicle for 24 h were stimulated with 1 mM ATP. ATP-induced permeabilization values represent the % of EB uptake above basal levels and were expressed as mean and SEM. n = 8–21 replicates using 6 animals (***P < 0.001 versus ATP, one-way ANOVA followed by post hoc Newman-Keuls test).
3.3. P2X7 Receptor Protein and mRNA Levels Do Not Change after Infection with S. Mansoni or Treatment with TGF-β1
To evaluate whether the reduction in P2X7 receptors function observed in schistosomiasis and in TGF-β1-treated cells was due, at least in part, to changes in P2X7 receptor expression, we quantified both P2X7 protein expression and the transcription from the P2rx7 locus by Western blotting and qRT-PCR, respectively. We detected similar levels of P2X7 protein and mRNA in macrophages from uninfected and infected groups (Figures 5(a) and 5(c)). Moreover, the levels of P2X7 protein and mRNA in macrophages from uninfected mice were not altered by treatment with 5 ng/mL TGF-β1 for 24 h (Figures 5(b) and 5(c)).
Figure 5.
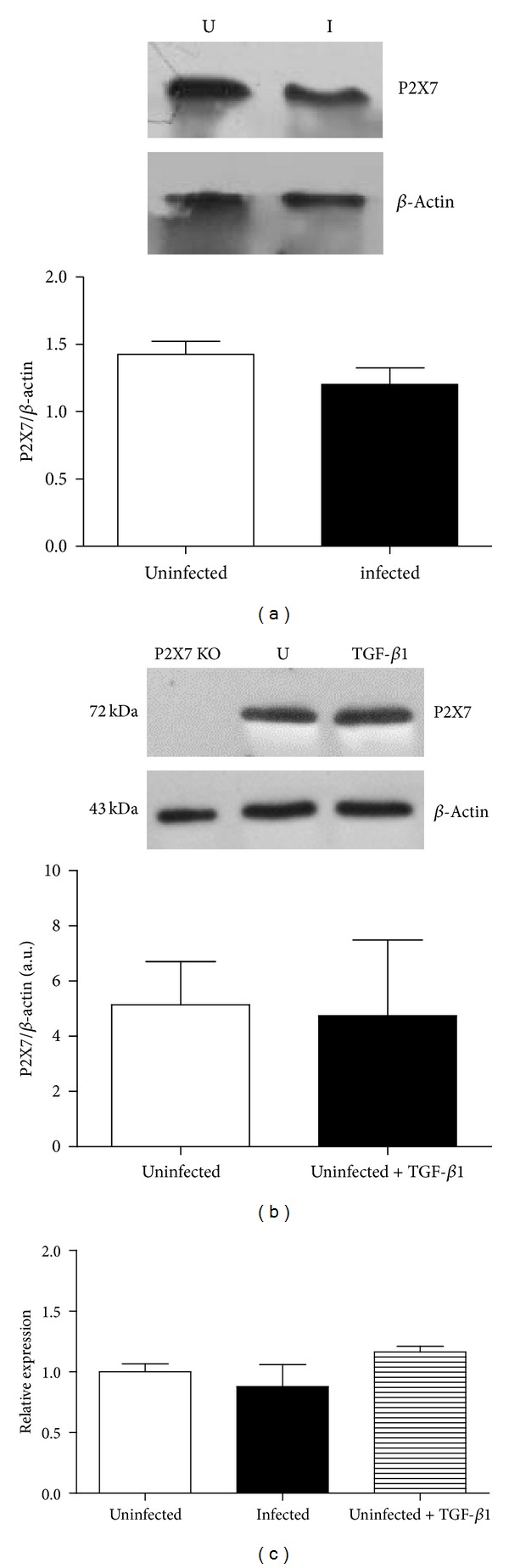
P2X7 receptors expression in peritoneal macrophages. (a) Macrophages from uninfected (white bar) and S. mansoni-infected (black bar) mice express similar levels of P2X7 protein. n = 9 replicates from three different animals, for each group. The images above are representative Western blots (U = uninfected and I = infected). (b) P2X7 receptors expression in peritoneal macrophages from control mice (white bar, untreated) or TGF-β1-treated cells (5 ng/mL, 24 h; black bar) are also similar. The images above are representative Western blots (U = uninfected; TGF-β1= TGF-β1-treated macrophages). Lysates from P2X7 KO macrophages were used as negative controls for P2X7 protein expression. Cells lysates were obtained from three animals for each group. (c) Quantitative RT-PCR (qRT-PCR) analysis of P2X7 receptor mRNA levels in peritoneal macrophages. Similar levels of P2X7 mRNA were found in uninfected (white bar), S. mansoni-infected mice (black bar), and TGF-β1-treated (5 ng/mL, 24 h; hatched bar) mouse macrophages. N = 3–5 samples per group (using different animals).
3.4. Schistosomiasis and Treatment with TGF-β1 Reduce Cell Surface P2X7 Levels in Peritoneal F4/80 Positive Macrophages
P2X7 receptors are targets of posttranslational modifications that regulate their insertion into plasma membrane [38], and the proinflammatory effects of P2X7 receptors (including IL-1β release and NO and ROS production) depend on receptor localization at the cell surface. Moreover, prolonged exposure of macrophages to ATP may induce P2X7 receptors removal from the cell surface by internalization [39, 40].
Since the reduction in P2X7 function during S. mansoni infection was not due to decreases in protein or mRNA levels, we hypothesized that changes in receptor structure or localization occurred during infection and downregulated receptor function, similar to that described for other P2X receptors [41]. To test this hypothesis, we analyzed P2X7 receptor localization in nonpermeabilized F4/80 (a common surface marker of mature macrophages) positive macrophages from uninfected and S. mansoni-infected mice using an antibody that recognizes an extracellular P2X7 epitope. Confocal microscopy analysis of anti-P2X7 labeled cells suggested that the levels of cell surface P2X7 receptors were reduced in macrophages from infected mice (Figures 6(a) and 6(b)). Interestingly, there was also a reduction in cell surface P2X7 in macrophages from uninfected mice treated with TGF-β1 (Figures 6(a) and 6(b)). Relative quantification of the immunofluorescence data showed that there was a 1.6-fold decrease (approximately) in the intensity of cell surface P2X7 fluorescence in macrophages from infected mice, when compared to that observed in uninfected control mice (Figure 6(b)).
Figure 6.
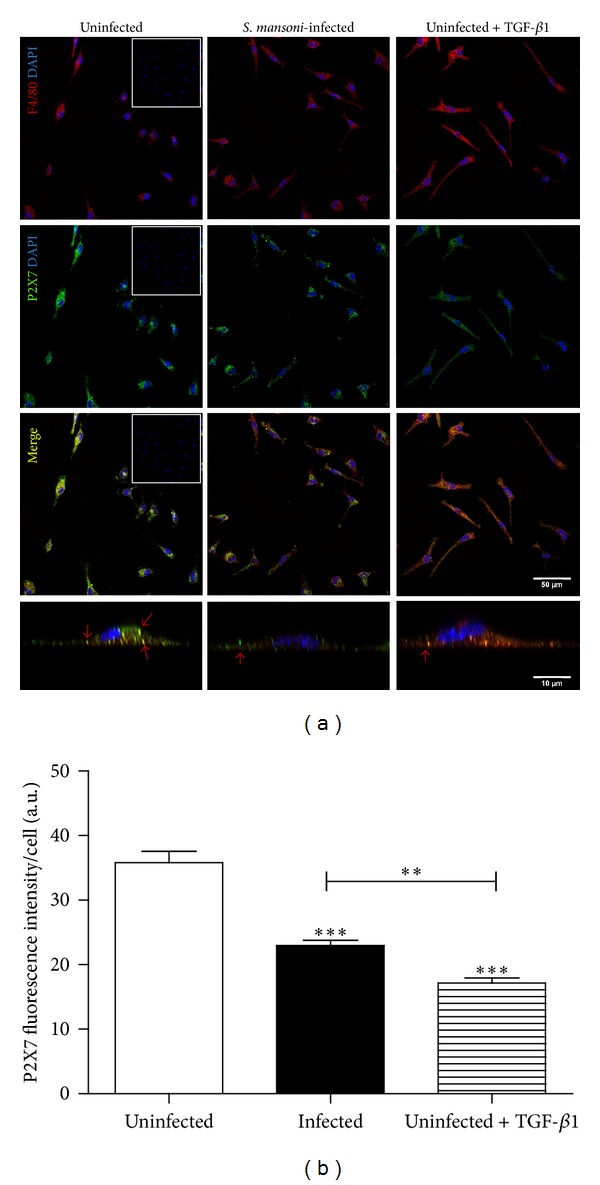
Confocal microscopy analysis of P2X7 receptor expression on the cell surface of macrophages. Cells were labelled with antibodies recognizing P2X7 receptors (green) and F4/80+ (red) as well as with DAPI (blue). (a) Representative images showing macrophages from uninfected (left column), S. mansoni-infected (middle column), and TGF-β1-treated (right column; 5 ng/mL, for 24 h) mice. The bottom row shows orthogonal slices of the representative labelled cells, with red arrows marking P2X7 expression on the plasma membrane. (b) Quantitative analysis of confocal microscopy data. The mean fluorescence intensity, showing cell surface P2X7 receptor expression in peritoneal macrophages from uninfected mice (white bar), S. mansoni-infected (black bar), and TGF-β1-treated (hatched bar; 5 ng/mL, for 24 h) mice, was obtained from pixels intensity using ImageJ software (see methods). Inset: negative controls (macrophages incubated with secondary antibodies only). n = 6 replicates from 3 animals for each condition (***P < 0.001 versus control, P < 0.01 TGF-β1 versus infected group; one-way ANOVA followed by post hoc Newman-Keuls test).
3.5. Reduced Survival of P2X7 KO Mice Infected with S. Mansoni
Next, to evaluate the participation of P2X7 receptors during the disease, we infected C57BL/6 wild type and P2X7 KO mice and followed the progression of infection. The survival curves showed that mortality in P2X7 KO infected mice started at 28 dpi and reached 100% at 60 dpi. In contrast, no death was observed in C57BL/6 wild type mice (Figure 7).
Figure 7.
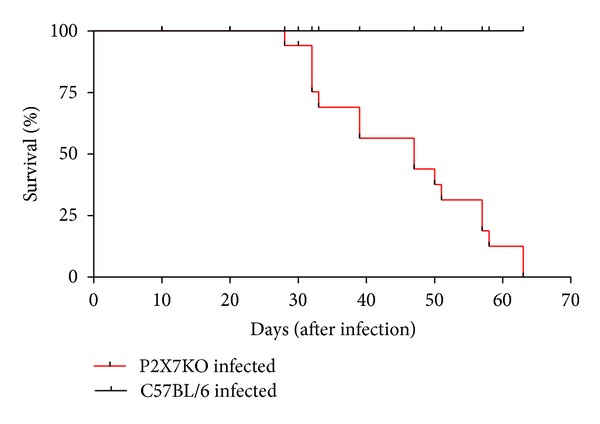
Survival curve of C57BL/6 wild type or P2X7 KO infected with approximately 80 cercariae of S. mansoni. (P2X7 KO: n = 9 females and 7 males; C57BL/6 wild type: 4 females and 10 males). ***P < 0.0001 using the Mantel-Cox log-rank test.
4. Discussion
During schistosomiasis, macrophages may display classical or activated phenotypes. The latter phenotype is present mainly in macrophages surrounding eggs within liver granulomas and was recently shown to be important in the control of tissue fibrosis [42]. Considering that schistosomiasis is a chronic inflammatory disease and that macrophages are involved in schistosomiasis pathogenesis [24], we sought to evaluate the function of P2X7 receptors in macrophages from mice infected with S. mansoni, since these receptors play an important role in inflammatory processes.
Here, we used a combination of ATP-induced permeabilization and intracellular Ca2+ measurement assays to show that, during the chronic phase of schistosomiasis, there is a reduction in the ATP-dependent P2X7 receptor function in macrophages (Figures 1 and 2), similar to that observed in mesenteric endothelial cells using the same experimental model [43].
While the function and expression of macrophage P2X7 receptors are positively regulated by proinflammatory cytokines such as IFN-γ [44, 45], Gadeock and coworkers [37] showed that the upregulation of P2X7 receptors in THP-1 monocytes is negatively regulated by TGF-β. On the other hand, peripheral blood mononuclear cells (PBMCs) from S. mansoni-infected mice produce high levels of TGF-β [27], and this cytokine is important to limit liver inflammation and favor host survival [27, 28]. Overall, these data are in agreement with our results also showing that the levels of peritoneal TGF-β1 are increased in mice infected with S. mansoni (Figure 3) and that TGF-β1, in a concentration close to the ones observed in the serum of chronic patients [46], reduces by approximately 50% the P2X7-dependent macrophage permeabilization triggered by ATP (Figure 4), mimicking the attenuated P2X7 response observed during S. mansoni infection (Figure 1(a)). Taken together, these results support our hypothesis that the downregulation of P2X7 function observed in schistosomiasis is a result of the increase in TGF-β1 levels during infection. Regarding TGF-β levels, despite the difference between the in vitro treatment concentration (5 ng/mL) and the in vivo peritoneal measurement (100 pg/mL), we must consider that we injected 1 mL of PBS in the peritoneal cavity of infected animals before removing the body fluid for cytokine determination. Therefore the samples were diluted, and, as a consequence, peritoneal TGF-β levels in vivo may be higher than the estimation in vitro. Moreover, previous data showed that individuals chronically infected with schistosomiasis have TGF-β serum levels around 20 ng/mL [46] and the serum level estimated in S. mansoni-infected mice (19.98 ± 2.37 ng/mL) is compatible with the value observed in patients [47]. Therefore we conclude that the TGF-β concentration used in present work is compatible with the disease, and the peritoneal TGF-β concentration in the infected animals is probably higher than estimated. Finally, TGF-β in the ng/mL range of concentration is able to modulate P2X7R function.
Western blotting and qRT-PCR data (using whole cell lysates) excluded the possibility that the reduction in P2X7 receptor function during S. mansoni infection or after treatment with TGF-β1 resulted from decreased levels of P2X7 protein or mRNA. In fact, immunofluorescence microscopy analysis of nonpermeabilized macrophages expressing F4/80 on their cell surface revealed that there was a lower density of surface P2X7 receptors in cells from infected mice compared to those from uninfected, control animals (Figure 6). Since total P2X7 protein levels were similar in both groups, we propose that there is reduction of P2X7 receptor in plasma membrane of macrophages from infected mice. The disappearance of immunoreactivity from plasma membrane could be related to some conformational changes of the receptor, an interaction with other proteins masking the extracellular epitope recognized by the antibody or the P2X7 receptor internalization. However, TGF-β treatment in vitro mimicked both the reduced cell surface receptor expression and function observed in macrophages obtained from infected animals. Consequently, a plausible explanation for the reduction in P2X7 function (without corresponding decreases in total protein or mRNA levels) during chronic schistosomiasis could be related to a reduced cell surface expression of P2X7 receptor.
Despite the work of Gadeock et al. [37], data investigating the link between TGF-β1 and P2X7 receptors function and/or expression are still missing. Since control macrophage treatment with TGF-β1 mimicked the profile of cell surface P2X7 receptors expression and function observed in live macrophages obtained from infected mice, it is supposed that TGF-β1 could be involved in the P2X7 receptors reduced function observed in the disease. To the best of our knowledge, this is the first report that shows a direct effect of TGF-β1 on macrophage P2X7 receptor function. However, we do not exclude that other cytokines could contribute to the P2X7 reduced function in macrophages from S. mansoni-infected mice, since in the later phase of the disease beyond the increase of TGF-β levels there is also the increase of IL-4, IL-5, IL-10, and IL-13.. Moreover, it was shown that IL-4 and IL-10 also inhibit EB uptake in rat alveolar macrophage [48]. Furthermore, previous data showed that TGF-β, IL-10, and IL-4 inhibit macrophage cytotoxicity which could be an important strategy used by S. mansoni to evade macrophage-mediated immune destruction [49].
Patients with schistosomiasis may be more susceptible to secondary infections, and prior infection with Schistosoma also increases the severity of secondary infections with Leishmania, Toxoplasma gondii, and Salmonella [50]. In addition, a previous report showed that macrophages from chronically infected mice have reduced phagocytic activity [51]; however, little is known about the mechanisms underlying this phenomenon. Recently, Wiley and Gu [52] showed that P2X7 receptors on the surface of monocytes/macrophages may act as scavenger receptors for bacteria in the absence of ATP. Therefore, the reduction of P2X7 receptor function in S. mansoni-infected animals could limit bacterial (and possibly protozoal) phagocytosis during schistosomiasis. This phenomenon might explain the increased susceptibility of patients infected with S. mansoni to secondary infections with other parasites and also the higher severity of secondary infections in these patients.
Kusner and Adams [53] showed that ATP kills Mycobacterium tuberculosis in human macrophages and that this effect depends on P2X7 receptor-mediated PLD activation. This mechanism is also important for the ATP-mediated killing of Chlamydia trachomatis in murine peritoneal macrophages [13]. Moreover, our group showed that the acute infection of macrophages with Chlamydia psittaci reduces P2X7 receptor-mediated cell permeabilization and Ca2+ influx, thus inhibiting macrophage apoptosis [54]. Considering that C. psittaci are obligatory intracellular parasites, this phenomenon could be an attempt to limit immunological responses by reducing ATP-mediated apoptosis and, consequently, favoring parasite survival. An important new aspect of P2X7 receptors signaling highlighted in the present work is that the function of macrophage P2X7 receptors may also be modulated by the extracellular parasite S. mansoni.
Altogether, our data suggest that, during schistosomiasis, the function of macrophage P2X7 receptors is reduced and that TGF-β1 plays a key putative role in the downregulation of receptor signaling. We hypothesize that macrophage purinergic receptors are differentially modulated during disease progression and that this phenomenon has a key role in the immune response against S. mansoni. In this context, our data show that P2X7 receptor knockout animals (P2X7 KO) are much more susceptible to death during S. mansoni infection than WT mice highlighting this P2 purinergic receptor and reinforcing the necessity of future studies exploring in deep its importance in S. mansoni-induced disease. Given that purinergic signaling is also found in S. mansoni, understanding the interplay between host and parasite purinergic signaling pathways is now important to clarify if future therapeutic approaches targeting these signaling pathways would be useful against schistosomiasis.
5. Conclusions
Altogether, our data show that S. mansoni infection reduced P2X7 function in peritoneal macrophages during the chronic phase of the disease. Furthermore, the peritoneal cavity of infected mice had increased levels of TGF-β1, and this cytokine reduced P2X7 receptor function in macrophages from uninfected mice. Thus, immunomodulation by TGF-β1 could limit P2X7-dependent inflammatory effects in macrophages from S. mansoni patients and may provide an explanation for the increased susceptibility of these patients to infections by others pathogens.
Acknowledgments
The authors thank Dr. Silvana Aparecida Rogel Carvalho Thiengo (Malacology Laboratory, FIOCRUZ, Rio de Janeiro, RJ, Brazil) for the donation of cercariae and Dr. Grasiella V. Matioszek (Institute of Biomedical Sciences, UFRJ) for technical assistance in confocal microscopy. This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Programa de Núcleos de Excelência (PRONEX-CNPq), and Instituto Nacional para Pesquisa Translacional em Saúde e Ambiente na Região Amazônica, Conselho Nacional de Desenvolvimento Científico e Tecnológico/MCT (INCT-INPeTAm/CNPq/MCT), Brazil. The funder had no role in the study design, data collection and analysis, decision to publish, or preparation of the paper.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nature Reviews Immunology. 2008;8(12):958–969. doi: 10.1038/nri2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Galli SJ, Borregaard N, Wynn TA. Phenotypic and functional plasticity of cells of innate immunity: macrophages, mast cells and neutrophils. Nature Immunology. 2011;12(11):1035–1044. doi: 10.1038/ni.2109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bianchi ME. DAMPs, PAMPs and alarmins: all we need to know about danger. Journal of Leukocyte Biology. 2007;81(1):1–5. doi: 10.1189/jlb.0306164. [DOI] [PubMed] [Google Scholar]
- 4.Saïd-Sadier N, Ojcius D. Alarmins, inflammasomes and immunity. Biomedical Journal. 2012;35(6):437–449. doi: 10.4103/2319-4170.104408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kono H, Rock KL. How dying cells alert the immune system to danger. Nature Reviews Immunology. 2008;8(4):279–289. doi: 10.1038/nri2215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.La Sala A, Ferrari D, Di Virgilio F, Idzko M, Norgauer J, Girolomoni G. Alerting and tuning the immune response by extracellular nucleotides. Journal of Leukocyte Biology. 2003;73(3):339–343. doi: 10.1189/jlb.0802418. [DOI] [PubMed] [Google Scholar]
- 7.Elliott MR, Chekeni FB, Trampont PC, et al. Nucleotides released by apoptotic cells act as a find-me signal to promote phagocytic clearance. Nature. 2009;461(7261):282–286. doi: 10.1038/nature08296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Junger WG. Immune cell regulation by autocrine purinergic signalling. Nature Reviews Immunology. 2011;11(3):201–212. doi: 10.1038/nri2938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Morandini AC, Savio LE, Coutinho-Silva R. The role of p2x7 receptor in infectious inflammatory diseases and the influence of ectonucleotidases. Biomedical Journal. 2014;37(4):169–177. doi: 10.4103/2319-4170.127803. [DOI] [PubMed] [Google Scholar]
- 10.Burnstock G. Purine and pyrimidine receptors. Cellular and Molecular Life Sciences. 2007;64(12):1471–1483. doi: 10.1007/s00018-007-6497-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Di Virgilio F. Purinergic signalling in the immune system. A brief update. Purinergic Signalling. 2007;3(1-2):1–3. doi: 10.1007/s11302-006-9048-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Burnstock G, Knight GE. Cellular distribution and functions of P2 receptor subtypes in different systems. International Review of Cytology. 2004;240:31–304. doi: 10.1016/S0074-7696(04)40002-3. [DOI] [PubMed] [Google Scholar]
- 13.Coutinho-Silva R, Stahl L, Raymond M, et al. Inhibition of chlamydial infectious activity due to P2X 7R-dependent phospholipase D activation. Immunity. 2003;19(3):403–412. doi: 10.1016/s1074-7613(03)00235-8. [DOI] [PubMed] [Google Scholar]
- 14.Coutinho-Silva R, Ojcius DM. Role of extracellular nucleotides in the immune response against intracellular bacteria and protozoan parasites. Microbes and Infection. 2012;14(14):1271–1277. doi: 10.1016/j.micinf.2012.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Humphreys BD, Dubyak GR. Modulation of P2X7 nucleotide receptor expression by pro- and anti- inflammatory stimuli in THP-1 monocytes. Journal of Leukocyte Biology. 1998;64(2):265–273. doi: 10.1002/jlb.64.2.265. [DOI] [PubMed] [Google Scholar]
- 16.Coutinho-Silva R, Alves LA, Savino W, Persechini PM. A cation non-selective channel induced by extracellular ATP in macrophages and phagocytic cells of the thymic reticulum. Biochimica et Biophysica Acta—Biomembranes. 1996;1278(1):125–130. doi: 10.1016/0005-2736(95)00200-6. [DOI] [PubMed] [Google Scholar]
- 17.Coutinho-Silva R, Persechini PM. P2Z purinoceptor-associated pores induced by extracellular ATP in macrophages and J774 cells. The American Journal of Physiology—Cell Physiology. 1997;273(6):C1793–C1800. doi: 10.1152/ajpcell.1997.273.6.C1793. [DOI] [PubMed] [Google Scholar]
- 18.Ferrari D, Pizzirani C, Adinolfi E, et al. The P2X7 receptor: a key player in IL-1 processing and release. The Journal of Immunology. 2006;176(12):3877–3883. doi: 10.4049/jimmunol.176.7.3877. [DOI] [PubMed] [Google Scholar]
- 19.Qu Y, Franchi L, Nunez G, Dubyak GR. Nonclassical IL-1β secretion stimulated by P2X7 receptors is dependent on inflammasome activation and correlated with exosome release in murine macrophages. The Journal of Immunology. 2007;179(3):1913–1925. doi: 10.4049/jimmunol.179.3.1913. [DOI] [PubMed] [Google Scholar]
- 20.Gudipaty L, Munetz J, Verhoef PA, Dubyak GR. Essential role for Ca2+ in regulation of IL-1β secretion by P2X7 nucleotide receptor in monocytes, macrophages, and HEK-293 cells. American Journal of Physiology-Cell Physiology. 2003;285(2):C286–C299. doi: 10.1152/ajpcell.00070.2003. [DOI] [PubMed] [Google Scholar]
- 21.Pearce EJ, MacDonald AS. The immunobiology of schistosomiasis. Nature Reviews Immunology. 2002;2(7):499–511. doi: 10.1038/nri843. [DOI] [PubMed] [Google Scholar]
- 22.Wilson MS, Mentink-Kane MM, Pesce JT, Ramalingam TR, Thompson R, Wynn TA. Immunopathology of schistosomiasis. Immunology and Cell Biology. 2007;85(2):148–154. doi: 10.1038/sj.icb.7100014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schramm G, Haas H. Th2 immune response against Schistosoma mansoni infection. Microbes and Infection. 2010;12(12-13):881–888. doi: 10.1016/j.micinf.2010.06.001. [DOI] [PubMed] [Google Scholar]
- 24.Barron L, Wynn TA. Macrophage activation governs schistosomiasis-induced inflammation and fibrosis. European Journal of Immunology. 2011;41(9):2509–2514. doi: 10.1002/eji.201141869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Melro MC, Mariano M. Extra-tissular Schistosoma mansoni egg granulomata in the peritoneal cavity of mice. Memorias do Instituto Oswaldo Cruz. 1987;82(supplement 4):245–252. doi: 10.1590/s0074-02761987000800047. [DOI] [PubMed] [Google Scholar]
- 26.Chuah C, Jones MK, Burke ML, McManus DP, Gobert GN. Cellular and chemokine-mediated regulation in schistosome-induced hepatic pathology. Trends in Parasitology. 2014;30:141–150. doi: 10.1016/j.pt.2013.12.009. [DOI] [PubMed] [Google Scholar]
- 27.Farah IO, Mola PW, Kariuki TM, Nyindo M, Blanton RE, King CL. Repeated exposure induces periportal fibrosis in Schistosoma mansoni- infected baboons: role of TGF-β and IL-4. Journal of Immunology. 2000;164(10):5337–5343. doi: 10.4049/jimmunol.164.10.5337. [DOI] [PubMed] [Google Scholar]
- 28.Herbert DR, Orekov T, Perkins C, Finkelman FD. IL-10 and TGF-β redundantly protect against severe liver injury and mortality during acute schistosomiasis. The Journal of Immunology. 2008;181(10):7214–7220. doi: 10.4049/jimmunol.181.10.7214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bhardwaj R, Skelly PJ. Purinergic signaling and immune modulation at the schistosome surface? Trends in Parasitology. 2009;25(6):256–260. doi: 10.1016/j.pt.2009.03.004. [DOI] [PubMed] [Google Scholar]
- 30.Lenzi HL, Romanha WDS, Santos RM, et al. Four whole-istic aspects of schistosome granuloma biology: fractal arrangement, internal regulation, autopoietic component and closure. Memorias do Instituto Oswaldo Cruz. 2006;101(1):219–231. doi: 10.1590/s0074-02762006000900034. [DOI] [PubMed] [Google Scholar]
- 31.Salomone S, Silva CLM, Morel N, Godfraind T. Facilitation of the vasorelaxant action of calcium antagonists by basal nitric oxide in depolarized artery. Naunyn-Schmiedeberg's Archives of Pharmacology. 1996;354(4):505–512. doi: 10.1007/BF00168443. [DOI] [PubMed] [Google Scholar]
- 32.Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca2+ indicators with greatly improved fluorescence properties. Journal of Biological Chemistry. 1985;260(6):3440–3450. [PubMed] [Google Scholar]
- 33.Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. The Journal of biological chemistry. 1951;193(1):265–275. [PubMed] [Google Scholar]
- 34.Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research. 2001;29(9, article e45) doi: 10.1093/nar/29.9.e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ruijter JM, Ramakers C, Hoogaars WMH, et al. Amplification efficiency: linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Research. 2009;37(6, article e45) doi: 10.1093/nar/gkp045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Persechini PM, Bisaggio RC, Alves-Neto JL, Coutinho-Silva R. Extracellular ATP in the lymphohematopoietic system: P2Z purinoceptors and membrane permeabilization. Brazilian Journal of Medical and Biological Research. 1998;31(1):25–34. doi: 10.1590/s0100-879x1998000100004. [DOI] [PubMed] [Google Scholar]
- 37.Gadeock S, Tran JNSN, Georgiou JG, et al. TGF-β1 prevents up-regulation of the P2X7 receptor by IFN-γ and LPS in leukemic THP-1 monocytes. Biochimica et Biophysica Acta: Biomembranes. 2010;1798(11):2058–2066. doi: 10.1016/j.bbamem.2010.07.022. [DOI] [PubMed] [Google Scholar]
- 38.Gonnord P, Delarasse C, Auger R, et al. Palmitoylation of the P2X7 receptor, an ATP-gated channel, controls its expression and association with lipid rafts. The FASEB Journal. 2009;23(3):795–805. doi: 10.1096/fj.08-114637. [DOI] [PubMed] [Google Scholar]
- 39.Hiken JF, Steinberg TH. ATP downregulates P2X7 and inhibits osteoclast formation in RAW cells. American Journal of Physiology—Cell Physiology. 2004;287(2):C403–C412. doi: 10.1152/ajpcell.00361.2003. [DOI] [PubMed] [Google Scholar]
- 40.Feng YH, Wang L, Wang Q, Li X, Zeng R, Gorodeski GI. ATP stimulates GRK-3 phosphorylation and β-arrestin-2-dependent internalization of P2X7 receptor. The American Journal of Physiology—Cell Physiology. 2005;288(6):C1342–C1356. doi: 10.1152/ajpcell.00315.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Robinson LE, Murrell-Lagnado RD. The trafficking and targeting of P2X receptors. Frontiers in Cellular Neuroscience. 2013;7(article 233) doi: 10.3389/fncel.2013.00233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Pesce JT, Ramalingam TR, Mentink-Kane MM, et al. Arginase-1-expressing macrophages suppress Th2 cytokine-driven inflammation and fibrosis. PLoS Pathogens. 2009;5(4) doi: 10.1371/journal.ppat.1000371.e1000371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Oliveira SDDS, Coutinho-Silva R, Silva CLM. Endothelial P2X7 receptors’ expression is reduced by schistosomiasis. Purinergic Signalling. 2013;9(1):81–89. doi: 10.1007/s11302-012-9332-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Blanchard DK, McMillen S, Djeu JY. IFN-γ enhances sensitivity of human macrophages to extracellular ATP-mediated lysis. Journal of Immunology. 1991;147(8):2579–2585. [PubMed] [Google Scholar]
- 45.Welter-Stahl L, da Silva CM, Schachter J, et al. Expression of purinergic receptors and modulation of P2X7 function by the inflammatory cytokine IFNγ in human epithelial cells. Biochimica et Biophysica Acta—Biomembranes. 2009;1788(5):1176–1187. doi: 10.1016/j.bbamem.2009.03.006. [DOI] [PubMed] [Google Scholar]
- 46.Ferreira RC, Montenegro SM, Domingues AL, et al. Almeida MO: TGF beta and IL13 in schistosomiasis mansoni associated pulmonary arterial hypertension; a descriptive study with comparative groups. BMC Infectious Diseases. 2014;14, article 282 doi: 10.1186/1471-2334-14-282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.El-Lakkany NM, Hammam OA, El-Maadawy WH, Badawy AA, Ain-Shoka AA, Ebeid FA. Anti-inflammatory/anti-fibrotic effects of the hepatoprotective silymarin and the schistosomicide praziquantel against Schistosoma mansoni-induced liver fibrosis. Parasites & Vectors. 2012;5(1, article 9) doi: 10.1186/1756-3305-5-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lemaire I, Leduc N. Purinergic P2X7 receptor function in lung alveolar macrophages: pharmacologic characterization and bidirectional regulation by Th1 and Th2 cytokines. Drug Development Research. 2003;59(1):118–127. [Google Scholar]
- 49.Oswald IP, Gazzinelli RT, Sher A, James SL. IL-10 synergizes with IL-4 and transforming growth factor-β to inhibit macrophage cytotoxic activity. Journal of Immunology. 1992;148(11):3578–3582. [PubMed] [Google Scholar]
- 50.Abruzzi A, Fried B. Coinfection of Schistosoma (Trematoda) with Bacteria, Protozoa and Helminths. Advances in Parasitology. 2011;77:1–85. doi: 10.1016/B978-0-12-391429-3.00005-8. [DOI] [PubMed] [Google Scholar]
- 51.Muniz-Junqueira MI, Prata A, Tosta CE. Phagocytic and bactericidal function of mouse macrophages to Salmonella typhimurium in schistosomiasis mansoni. The American Journal of Tropical Medicine and Hygiene. 1992;46(2):132–136. doi: 10.4269/ajtmh.1992.46.132. [DOI] [PubMed] [Google Scholar]
- 52.Wiley JS, Gu BJ. A new role for the P2X7 receptor: a scavenger receptor for bacteria and apoptotic cells in the absence of serum and extracellular ATP. Purinergic Signalling. 2012;8(3):579–586. doi: 10.1007/s11302-012-9308-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kusner DJ, Adams J. ATP-induced killing of virulent Mycobacterium tuberculosis within human macrophages requires phospholipase D. Journal of Immunology. 2000;164(1):379–388. doi: 10.4049/jimmunol.164.1.379. [DOI] [PubMed] [Google Scholar]
- 54.Coutinho-Silva R, Perfettini J, Persechini PM, Dautry-Varsat A, Ojcius DM. Modulation of P2Z/P2X7 receptor activity in macrophages infected with Chlamydia psittaci . American Journal of Physiology—Cell Physiology. 2001;280(1):C81–C89. doi: 10.1152/ajpcell.2001.280.1.C81. [DOI] [PubMed] [Google Scholar]