

Abstract
Janus kinase-signal transducer and activator of transcription (JAK/STAT) signalling, pivotal in Philadelphia-negative (Ph-ve) myeloproliferative neoplasm (MPN), is negatively regulated by molecules including SOCSs, CISH and SHP1. SOCS1, SOCS2 and SOCS3 methylation have been studied in MPN with discordant results. Herein, we studied the methylation status of SOCS1, SOCS2 and SOCS3, CISH and SHP1 by methylation-specific polymerase chain reaction (MSP) in cell lines and 45 diagnostic marrow samples of Ph-ve MPN. Moreover, we attempted to explain the discordance of methylation frequency by mapping the studied MSP primers to the respective genes. Methylation was detected in normal controls using SOCS2 MSP primers in the 3′translated exonic sequence, but not primers around the transcription start site in the 5′ untranslated regions (5′UTR). SOCS1, SOCS2, SOCS3 and CISH were completely unmethylated in primary MPN samples and cell lines. In contrast, methylation of SHP1 was detected in 8.9% primary marrow samples. Moreover, SHP1 was completely methylated in K562 cell line, leading to reversible SHP1 silencing. A review of methylation studies of SOCS1 and SOCS3 showed that spuriously high rates of SOCS methylation had been reported using MSP primers targeting CpG sites in the 3′translated exonic sequence, which is also methylated in normal controls. However, using MSP primers localized to the 5′UTR, methylation of SOCS1, SOCS2 and SOCS3 is infrequent across all studies. In summary, methylation of SOCS1, SOCS2, SOCS3 and CISH is infrequent in Ph-ve MPN. Appropriate MSP primers are important for accurate estimation of the methylation frequency. The role of SHP1 methylation in the pathogenesis of MPN warrants further investigation.
Keywords: myeloproliferative neoplasms, methylation, SOCS1, SOCS2, SOCS3, CISH, SHP1, MSP primer
Introduction
Philadelphia-negative (Ph-ve) myeloproliferative neoplasm (MPN) is a stem cell disease with proliferation of myeloid compartment, leading to development of distinct clinical entities such as polycythemia vera (PV), essential thrombocythemia (ET) and primary myelofibrosis (PMF) [1, 2]. Recently, Janus kinase 2 (JAK2) V617F mutation was detected in most of the patients with PV and about half of the patients with ET and PMF, leading to constitutive activation of Janus kinase-signal transducer and activator of transcription (JAK/STAT) signalling [3, 4]. However, pathogenesis in those without JAK2 V617F mutation remains unknown.
The JAK/STAT signalling pathway is important for the transmission of cytokine signals from cell surface to the nucleus [5]. Binding of cytokines to their cognate receptors results in the dimerization of receptor complexes and activation of the Janus family of protein tyrosine kinases [6, 7], followed by phosphorylation of the cytoplasmic STATs. Upon phosphorylation, STATs form homo- or hetero-dimers migrate to the nucleus and activate gene transcription. This JAK/STAT pathway is subject to negative regulation by PIAS members, JAK inhibitors like SOCS family proteins and protein tyrosine phosphatases (PTP) such as SHP1 [6–8]. The SOCS family comprises eight members, including SOCS1-7 and CISH, and is characterized by the presence of a central Src homology (SH2) domain that is flanked by a variable length N-terminal domain, and a conserved 40 amino acid carboxy terminal ‘SOCS box’ domain [6, 7]. SOCS proteins suppress JAK/STAT signalling by binding with their SH2 domain to phosphotyrosine residues in cytokine receptors or activated JAKs [9]. Among them, CISH, localized at 3p21.2; SOCS1, at 16p13.13; SOCS2 at 12q22; and SOCS3 at 17q25.3 are most thoroughly studied in haematological malignancies. SOCS1 and SOCS3 can bind the activated JAKs and cytokine receptors, respectively, through SH2 domain to inhibit JAK tyrosine kinase activation. CISH and SOCS2 inhibit STAT activation by binding to phosphorylated tyrosine residues on activated cytokine receptors and compete with STAT or hinder the STAT-binding sites of receptors [9]. SOCS members are cytokine-inducible negative regulators of the cytokine signalling [9]. SOCS1, SOCS2, SOCS3 and CISH can be induced by a multitude of cytokines, and hence serve as negative feedback to curb excessive cytokine signalling [9]. SOCS1 and SOCS3 comprise two exons with the first exon being untranslated, whereas SOCS2 consists of three exons with the first exon being untranslated [10–12]. All these three genes are embedded in a huge CpG island of more than two kilobases extending from the 5′UTR to the 3′translated exonic sequence, as demonstrated in the corresponding websites in NCBI NIH Roadmap Epigenomics project [13].
SHP1, also known as HCP, SHPTP1 and PTP1C, is a 68-kd, cytoplasmic PTP [14]. The human SHP1 gene is located on chromosome 12p13, consists of 17 exons and spans ∼17 KB of DNA. It contains two tandem Src homology (SH2) domains, a catalytic domain and a C-terminal tail of about 100 amino acid residues [14]. In contrast to the ubiquitous expression of the structurally related SHP2, SHP1 is primarily expressed in haematopoietic cells, and considered a putative tumour suppressor gene in lymphoma and leukaemia, as it antagonizes the growth-promoting and oncogenic potentials of protein tyrosine kinase [14].
Hypermethylation of promoter-associated CpG islands of tumour suppressor genes and recently microRNA [15–17], resulting in gene silencing, and hence inactivation of tumour suppressor genes, has been implicated in the pathogenesis of haematological malignancies [18]. Moreover, aberrant DNA methylation of SOCS1, SOCS2, SOCS3 and SHP1 has been studied in Ph-ve MPN with discordant results [19–23]. Our previous study has shown that while normal cells possess unmethylated CpG islands around the transcription start site (TSS) in the 5′UTR, CpG sites inside the coding exonic sequence are methylated for SOCS1, thereby emphasizing the importance of methylation-specific polymerase chain reaction (MSP) primer selection [24, 25]. Moreover, there is scanty data on the methylation of CISH in Ph-ve MPN. In this study, we investigated the methylation profile of SOCS1, SOCS2, SOCS3, CISH and SHP1 in MPN by the use of MSP primers in the both 5′UTR and translated exonic sequence of the genes. Finally, we attempted to explain the discordance of methylation frequency of SOCS1 and SOCS3 in MPN by mapping the studied MSP primers to the respective genes.
Materials and methods
Patient samples
DNA was extracted from primary bone marrow samples at diagnosis of 45 patients with Ph-ve MPN. Diagnosis of Ph-ve MPN including ET, PV and PMF was based on WHO criteria. MSP on DNA extracted from primary marrow samples at diagnosis was performed on 45 patients with MPN [ET, N = 34 (75.6%); PV, N = 7 (15.6%) and PMF, N = 4 (8.9%)]. The clinical features of these patients have been previously described [26–28]. In brief, there were 24 (53.5%) male and 21 (46.7%) female patients with a median age of 67.5 years (range: 28–89 years) The median presenting platelet count of 848 × 109/l, presenting Hb of 9–22 g/dl (median 13.3 g/dl), median presenting leucocyte count of 14.4 × 109/l (range: 7–28 × 109/l). Apart from 5 (11.1%) patients in whom the presenting symptoms was unavailable, 25 (62.5%) were asymptomatic at diagnosis, 4 (10%) with bleeding, 4 (10%) with erythromelalgia, 2 (5%) with minor stroke, 3 (7.5%) with abdominal pain and one each (2.5%) with blurred vision and weight loss. JAK2 V617F mutation was presented in 26 of the 40 patients. DNA from three normal bone marrow donors and five normal peripheral blood donors was used as negative control, while enzymatically methylated control DNA (CpGenome Universal Methylated DNA, Millipore, Billerica, MA, USA) was considered as positive control. The study has been approved by Institutional Review Board of Queen Mary Hospital, and written informed consent has been obtained.
Cell lines and culture
Cell lines and culture were described in previous study [28]. SET-2 cells were purchased from Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DMSZ) (Braunschweig, Germany). HEL cells is a human erythroleukemia cell line generously provided by Dr Zhang Dong-Er, Department of Pathology and Molecular Biology, Moores Cancer Center, University of California San Diego, USA. MEG-01 and K562 cells were kindly provided by Dr Mo Yang, Department of Paediatrics, Queen Mary Hospital, The University of Hong Kong, Hong Kong. SET-2 was derived from ET at megakaryoblastic leukaemic transformation. HEL was derived from peripheral blood of a patient with erythroleukemia. Both SET-2 and HEL carried JAK2 V617F mutation. MEG-01 and K562 were derived from blastic transformation of patients with CML and carried BCR/ABL fusion gene. Cell cultures were cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA), supplemented with 10% foetal bovine serum (20% for SET-2; Invitrogen), 50 U/ml penicillin and 50 μg/ml streptomycin (Invitrogen) in a humidified atmosphere of 5% CO2 at 37°C.
Methylation-specific polymerase chain reaction
DNA extraction from bone marrow samples, peripheral blood and cell lines was performed with QIAamp DNA Blood Mini Kit according to the manufacturer's instructions. DNA bisulphite conversion was performed with a commercially available kit (EpiTect Bisulfite Kit, Qiagen, Duesseldorf, Germany). Methylation status of SOCS1, SOCS2, SOCS3, CISH and SHP1 gene promoter CpG islands was investigated by MSP as previously reported [24]. Primers used for the methylated MSP (M-MSP) and unmethylated MSP (U-MSP) were listed in the Table S1. For SOCS1, based on our previous study [24], MSP primer inside exon 1 was methylated in normal controls, and hence unsuitable for methylation analysis. In contrast, MSP primers upstream of the translation start region (MSP-5′) were unmethylated in normal controls, and hence used in this study. For SOCS2, two sets of primers, located 5′ (SOCS2-5′) and 3′ (SOCS2-3′) to the translation start site of SOCS2 were used in this study. SOCS2-5′ primers were used according to previous study [29]. SOCS2-3′ primers were mapped to the region, in which methylation of SOCS2 has been studied by bisulphite genomic sequencing and methylation-sensitive restriction enzyme assay in MPN patients samples by Quentmeier et al. [30]. SOCS3 primers were adopted from He et al. [31]. All MSP were performed in thermal cycler (9700, Applied Biosystems, Foster City, CA, USA) under the conditions: 95°C for 10 min., followed by specific cycles of 95°C for 30 sec., specific annealing temperature for 30 sec., 72°C for 30 sec. and a final extension of 10 min. at 72°C (Table S1). The MSP mixture contained 30 ng of bisulphite-treated DNA, 0.2 mM dNTPs, MgCl2 (Table S1), 10 pmol of each primer, 1× PCR buffer and 1 unit of FastStart Taq DNA polymerase (Roche, Mannheim, Germany) in a final volume of 25 μl. Ten microlitre of PCR products was loaded onto 6% polyacrylamide gels, electrophoresed, stained with ethidium bromide and visualized under ultraviolet light.
5-Aza-2′-deoxycytidine (5-AzadC) treatment of the cell line K562
K562 cells were completely methylated for SHP1. For treatment with 5-AzadC, K562 cells (1 × 106 cells/ml) were seeded into six-well plates on day 0 and cultured with 0.5 μM of 5-AzadC from day 1 to day 3. Cells on day 0 and day 3 of treatment were harvested.
Reverse transcription-PCR for SHP1
MirVana™ miRNA Isolation Kit (Ambion, Austin, TX, USA) and QuantiTect Reverse Transcription kit (Qiagen, Duesseldorf, Germany) were used for isolation of total RNA and RT-PCR, respectively, according to the manufacturers’ instructions. For SHP1 RT–PCR, the following primers: forward 5′-GGC ACT GGG AGC TGC ATC TGA GGC-3′; reverse 5′-CTC GCA CAT GAC CTT GAT GTG-3′ were used. For GAPDH RT–PCR, the following primers: forward 5′-ACC ACA GTC CAT GCC ATC ACT-3′; reverse 5′-TCC ACC ACC CTG TTG CTG TA-3′ were used.
Statistical analysis
Correlation between SHP1 gene methylation status and the presence of JAK2 V617F mutation was computed by the Chi-square test (or Fisher Exact test) using Statistical Package for the Social Sciences (SPSS, IBM, New York, NY, USA) version 18.0. All P values were two-sided.
Results
Controls
As mentioned in the section for materials and methods, two sets of MSP primers, one in the 5′UTR and the other in the 3′translated exon sequence, were available for SOCS1 (SOCS1-5′ and SOCS1-3′) and SOCS2 (SOCS2-5′ and SOCS2-3′). SOCS1-3′ has been shown to be methylated in normal controls [24], and hence SOCS1-5′ MSP primers was used in this study. Using the MSP primers in the 5′UTR near the TSS as described previously, SOCS1 was unmethylated in normal controls, but methylated in the positive control [24]. For SOCS2, using downstream MSP primers inside the translated exonic sequence, i.e. SOCS2-3′, methylation was detected in four of five normal control DNA samples (Fig. 1A). Moreover, sequencing of the M-MSP products confirmed authenticity of methylation in these normal controls, which was demonstrated by the presence of unconverted, and hence methylated, cytosine molecules at ‘CpG’ dinucleotide (Fig. 1B). On the other hand, using MSP primers in the 5′UTR near the TSS, SOCS2-5′ MSP primers, SOCS2 was unmethylated in all of the normal control samples, but methylated in the positive control DNA (Fig. 2). Therefore, we studied methylation of the SOCS2 gene using MSP with primer set in the 5′UTR near the TSS instead of inside the translated exon sequence. For the SOCS3, SHP1 and CISH genes, M-MSP was negative in normal DNA, while positive for the methylated control DNA. Conversely, U-MSP was positive in normal DNA, while negative for the methylated control DNA (Fig. 2). Sequencing of the M-MSP products of these four genes from the methylated control DNA showed the expected nucleotide changes, and hence confirmed complete bisulphite conversion and specificity of MSP primers.
Fig. 1.
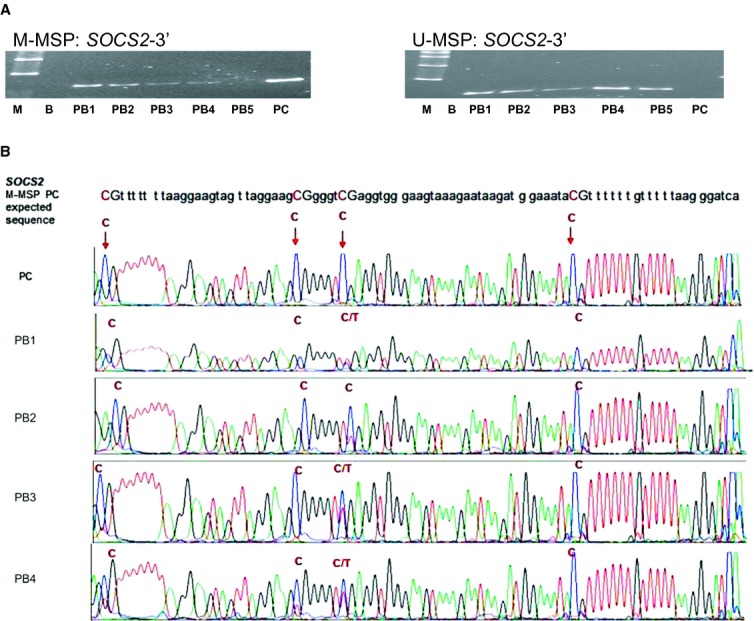
(A) Methylation-specific PCR of normal controls using SOCS2-3′ methylation-specific polymerase chain reaction (MSP) primers. M-/U-MSP analysis showed that four of five normal peripheral blood controls (PB1-5) were methylated with the SOCS2-3′ primers. (M: DNA marker; B: blank; PC: positive control; PB, normal peripheral blood control). (B) Sequencing of MSP products using SOCS2-3′ MSP primers in normal controls and positive control showing methylation signals. Methylated cytosine residues [C] in CpG dinucleotide remained as C, whereas unmethylated cytosine read as [T] after bisulphite conversion. (PC: positive control; PB: normal peripheral blood control).
Fig. 2.
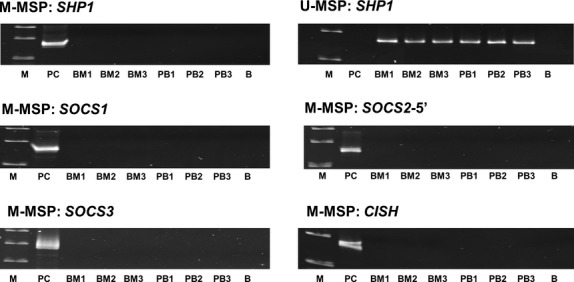
Methylation-specific polymerase chain reaction (MSP) of SOCS1, SOCS2-5′, SOCS3, CISH and SHP1 in normal controls. M-/U-MSP analysis showed that three normal peripheral blood controls (PB1-3) and three bone marrow controls (BM1-3) were unmethylated with the SOCS1, SOCS2-5′, SOCS3, CISH and SHP1 primers. (M: DNA marker; B: blank; PC: positive control; PB, normal peripheral blood control; BM: normal bone marrow control).
Cell lines
SHP1 was homozygously methylated in K562, but completely unmethylated in HEL, MEG-01 and SET-2. On the other hand, SOCS1, SOCS2, SOCS3 and CISH genes were completely unmethylated in these four cell lines (Fig. 3).
Fig. 3.
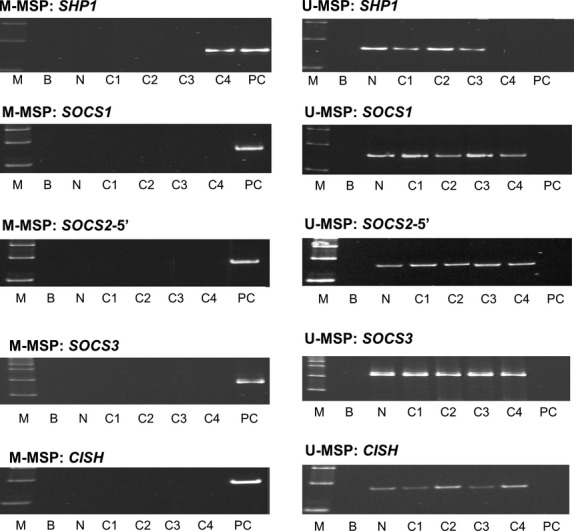
Methylation-specific polymerase chain reaction of SOCS1, SOCS2-5′, SOCS3, CISH and SHP1 in cell lines. For SHP1, K562 was homozygously methylated, while HEL, SET-2 and MEG-01 were completely unmethylated. HEL, SET-2, MEG-01 and K562 were completely unmethylated for SOCS1, SOCS2-5′, SOCS3 and CISH. (M: DNA marker; B: blank; PC: positive control; N: normal control; C1: HEL; C2: MEG-01; C3: SET-2; C4: K562).
Primary samples
Methylation-specific polymerase chain reaction was performed in 45 primary samples of Ph-ve MPN patients for these five genes. None of the patients carried hypermethylation of SOCS1, SOCS2, SOCS3 and CISH (Fig. 4A). Nevertheless, methylation of SHP1 was found in four ET patients, which was confirmed by direct sequencing of M-MSP products of positive control and primary samples (Figs 4A and B). Of these, three (75%) patients carried JAK2 V617F mutation. Two were female. The median age was 63.3 years. There was no association of SHP1 methylation and JAK2 V617F mutation (P = 0.99). One presented with cerebral infarction, one with peptic ulcer and the other two were asymptomatic at diagnosis. The median diagnostic Hb was 12.7 g/dl, platelet 962 × 109/L and leucocyte 15.4 × 109/L. Median survival was 48.6 months. At the time of writing, two had died, one of cerebral infarction and the cause of death of the other was unknown. No myeloid transformation had occurred.
Fig. 4.
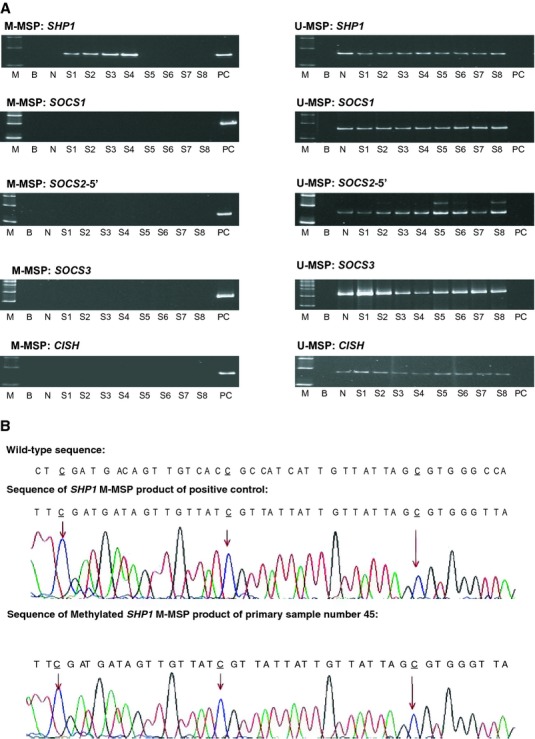
Methylation-specific polymerase chain reaction (MSP) of SOCS1, SOCS2-5′, SOCS3, CISH and SHP1 in MPN primary samples. (A) M-/U-MSP analysis showed that for SOCS1, SOCS2-5′, SOCS3, CISH, methylation was absent in MPN marrow samples, whereas for SHP1, methylation was found in four MPN patients. (M: DNA marker; B: blank; PC: positive control; N: normal control; S: primary sample). (B) DNA sequencing of SHP1 M-MSP products from bisulphite-converted methylated positive and MPN primary samples showing methylated cytosine [C] residues in CpG dinucleotide remained unchanged, unmethylated C residues were converted into [T], whereas all the non-CpG C residues were unmethylated and were converted to thymidine [T].
5-AzadC treatment of K562 cells
Untreated K562 cell line showed complete methylation of SHP1. Treatment of cells with 0.5 μM 5-AzadC for 3 days led to demethylation of SHP1 as demonstrated by the emergence of SHP1 U-MSP signal (Fig. 5A). Moreover, RT–PCR demonstrated that SHP1 gene was re-expressed after 5-AzadC treatment of K562 cells (Fig. 5B).
Fig. 5.
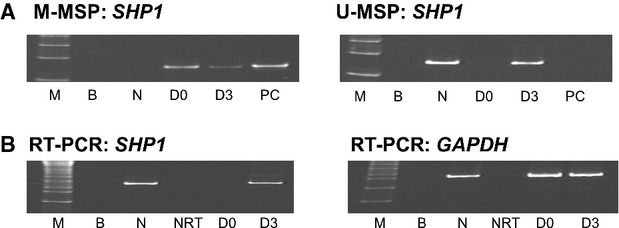
Effect of 5-AzadC treatment on K562 cells. (A) M-/U-methylation-specific polymerase chain reaction analysis of SHP1 promoter methylation status showed that 5-AzadC treatment led to progressive demethylation of SHP1 promoter in K562 cells. (M: DNA marker; B: blank; PC: positive control; N: normal control; D0, day 0; D3, day 3 culture in 5-AzadC with 0.5 μM). (B) Reverse transcription–PCR (RT-PCR) analysis of the GAPDH status and SHP1 expression after 5-AzadC treatment. (M: DNA marker; B: blank; N: normal control; NRT: negative control without reverse transcriptase; D0, day 0; D3, day 3 culture in 5-AzadC with 0.5 μM).
Discussion
To account for the discrepancy among the frequency of methylation of SOCS1, SOCS2 and SOCS3 in MPN, we have mapped the MSP primers used in different studies onto the gene map of SOCS1, SOCS2 and SOCS3 (Figures S1–3; Table 1) and derived the following observations.
Table 1.
Literature about the frequencies of SOCS1, SOCS2, SOCS3, SHP1 and CISH methylation in Ph-ve MPN. Primers positions of SOCS1, SOCS2 and SOCS3 genes are represented by arrows in supplementary figures
| Author | Zhang | Jost [23] | Fourouclas [22] | Capello [20] | Quentmeier* [30] | Fernandez -Mercado [21] | Teofili [29] | Barrio† [19] | Fodermayr [34] |
|---|---|---|---|---|---|---|---|---|---|
| No of case | 46 | 39 | 73 | 112 | 7 | 39 | 81 | 35 | 39 |
| Genes studied | SOCS1, SOCS2, SOCS3, CISH, SHP1 | SOCS1, SHP1 | SOCS1, SOCS3 | SOCS1, SOCS3, SHP1 | SOCS2 | SOCS1, SOCS3 | SOCS1, SOCS2, SOCS3, | SOCS1, SOCS2, SOCS3,, CISH | SOCS1, SOCS3, SHP1 |
| SOCS1 Primer site | 5′ UTR: Figure S1-A | exon2: Figure S1-C | 5′ UTR: Figure S1-B exon2: Figure S1-C | exon2: Figure S1-C | — | 5′ UTR: Figure S1-A exon2 : no mention | exon2: Figure S1-C | No mention | exon2: Figure S1-C |
| % SOCS1 methylation | 0 | 15.4 | 5′ UTR : 0; exon2: 38.4 | 13.4 | — | 5′ UTR : 0 exon2 region 1: 72.2 exon2 region 2: 0 | 13.5 | 34.3 | 12.8 |
| SOCS2 Primer site | 5′ UTR: Figure S2-A | — | — | — | 5′ UTR: Figure S2-B | — | 5′ UTR: Figure S2-A | No mention | — |
| % SOCS2 methylation | 0 | — | — | — | 28.6 | — | 0 | 0 | — |
| SOCS3 Primer site | 5′ UTR: Figure S3-C | — | 5′ UTR: Figure S3-A | exon 2:Figure S3-D | — | 5′UTR: Figure S3-B intron 1 | 5′ UTR: Figure S3-C | No mention | Intron 1 |
| % SOCS3 methylation | 0 | — | None of PV or ET but 27% of PMF | 41.1 | — | 5′UTR: 7.7 intron 1: 23.1 | 13.5 | 0 | 15.4 |
| % SHP-1 methylation | 8.7 | 0 | — | 7.1 | — | — | — | — | 12.8 |
| % CISH methylation | 0 | — | — | — | — | — | — | 0 | — |
Using bisulphite genomic sequencing and methylation-sensitive restriction enzyme assay to detect methylation status.
Using Human-Methylation27 DNA Analysis BeadChip’ (Illumina) to detect methylation status.
UTR: untranslated region; PV: polycythemia vera; ET: essential thrombocytosis; PMF: primary myelofibrosis.
Firstly, SOCS1, SOCS2 and SOCS3 genes have multiple exons with the first exon being untranslated. Moreover, all of them are embedded in a huge CpG island spanning >2 kb that extends from the 5′UTR into the 3′translated protein-coding exonic sequences (Figures S1–3).
In a previous study, we have shown that using MSP primers of SOCS1-3′ located in the translated exon 2, as described by Watanabe et al. [32], methylation was detected in six of 12 normal peripheral blood and two of three normal marrow samples [24]. These results were confirmed by sequencing, suggesting that methylation within SOCS1 exon 2 might not be involved in the regulation of gene transcription. The lack of impact of SOCS1 exon 2 methylation on gene expression has been illustrated in HL60 and U937 cell lines, in which complete methylation of CpG islands within SOCS1 exon 2 was associated with significant expression of SOCS1 [32] (Figure S1), and hence a lack of correlation between methylation of CpG sites in the 3′translated exonic sequence and gene silencing. On the contrary, methylation of CpG islands in the 5′UTR, using MSP primer targeting CpG sites in the 5′UTR, has been shown to be associated with inhibition of expression of, and hence silencing of SOCS1 by immunohistochemistry in hepatoblastoma [33]. Therefore, methylation of SOCS1 should be studied with MSP primers mapping to the 5′UTR (SOCS1-5′) as performed in this study. Using this SOCS1-5′ MSP primers, SOCS1 methylation was not detected in any primary MPN samples or cell lines. Similarly, other studies using primers targeting CpG sites in the 5′UTR also showed that none of a total of 112 MPN cases (73 in Fourouclas et al. and 39 in Fernández-Mercado et al.) had methylation of SOCS1 [21, 22] (Figure S1). In contrast, studies using MSP primers inside the 3′translated exonic sequence showed methylation frequencies ranging from 12.8% to 72% (median: 14.5%) [20–23, 29, 34] (Figure S1). Therefore, these findings were consistent with the notion that a boundary exists between methylated and unmethylated CpG dinucleotide within a CpG island, and that CpG dinucleotide inside the 3′translated protein-coding sequences is commonly methylated in normal cells.
Similarly, SOCS2 methylation has been reported to be frequently methylated in MPN [30]. In contrast, using MSP primers near the TSS in the 5′UTR, we and others found the absence of SOCS2 methylation [19, 29]. Quentmeier et al., using methylation-sensitive enzymes targeting CpG sites inside the 3′translated exon 2 sequence, showed that 28.6% of MPN patients had methylation of SOCS2 [30] (Figure S2). In this study, we have designed MSP primers mapping to the same exon 2 sequence, and confirmed that CpG sites in this region is in fact methylated in normal controls, and hence unimportant for epigenetic regulation of SOCS2 transcription, and unsuitable for methylation study.
Moreover, using MSP primers close to the TSS in the 5′UTR (Figure S3), we and others showed infrequent methylation of SOCS3 in MPN (this study, 0%; Fourouclas et al., 27%; Teofilli et al., 13.5% and Fernández-Mercado et al., 7.7%) [21, 22, 29], in contrast to frequent methylation of 41.1% samples when MSP primers inside the translated exonic DNA sequence were used [20]. In our study, we adopted MSP primers from He et al. [31] near the TSS (Figure S3) for several reasons. Firstly, our MSP primers were located close to the TSS in the 5′UTR region. Secondly, methylation in CpG islands within this region has been shown to be associated with SOCS3 silencing and aberrant activation of JAK/STAT signalling in lung cancer cell lines, and hence biologically relevant [31]. Using MSP primers in the 5′UTR, SOCS3 methylation was not detected in any of the patients with PV or ET [22]. However, Fourouclas et al. demonstrated SOCS3 methylation in 27% patients with PMF, suggesting a disease-specific SOCS3 methylation among MPN [22]. Unfortunately, in our series, we had too few patients with PMF to verify this finding.
Overall, our findings and literature review showed that MSP primer selection is important in the study of methylation as a methylation boundary may occur in some genes, in which methylation of CpG inside the translated exonic DNA sequence, unrelated to gene silencing, may occur in normal cells. Indeed, in a study of the HIC1 gene, CpG island methylation inside exon 3 was present in normal controls, which was not associated with down-regulation of HIC1, and hence unimportant for transcriptional regulation of HIC1. In contrast, CpG sites in the 5′UTR was unmethylated in normal marrow control, but aberrantly methylated in some AML samples, thereby emphasizing the importance of methylation of 5′UTR, promoter-associated CpG islands, instead of downstream CpG sites in the translated exonic region in epigenetic regulation of gene expression [35]. Moreover, a possible boundary between methylated and unmethylated sequence in SOCS1 gene was suggested by the observation that methylation was absent in the 5′UTR of SOCS1, but was detected in 85% of normal DNA using MSP primers targeting CpG sites in the 3′translated exon 2 sequence [21]. Similarly, in SOCS3, methylation was infrequent using primers near the TSS in the 5′UTR, but present in 41% of primary MPN samples when MSP primers inside the translated exon 2 sequence were used [20–22, 29]. Therefore, one has to select MSP primers in the 5′UTR region, particularly close to the TSS, or else, methylation unrelated to epigenetic regulation might give rise to spuriously high frequency of gene methylation.
In contrast to the SOCS family, SHP1, another negative regulator of JAK/STAT signalling, has been infrequently studied for methylation in MPN. We have shown that in myeloma, SHP1 methylation was frequent, leading to reversible SHP1 silencing and constitutive JSK/STAT activation [24]. Here, we showed methylation of SHP1 in primary MPN samples and K562 cells, which were verified by direct sequencing of M-MSP PCR products. Therefore, SHP1 methylation, leading to reversible gene silencing, is implicated in the constitutive activation of JAK/STAT in MPN, and hence warrants further study in larger number of patients. Similarly, CISH methylation is absent in MPN.
Conclusion
In conclusion, this is a comprehensive study of methylation profile of SOCS1, SOCS2, SOCS3, CISH and SHP1 in MPN. Among these, only SHP1 was methylated in MPN, and hence implicated in the constitutive activation of JAK/STAT signalling in MPN. Moreover, careful selection of MSP primers is important.
Conflicts of interest
The authors confirm that there are no conflicts of interest.
Supporting information
Additional Supporting Information may be found in the online version of this article:
Figure S1 Schematic diagram of the SOCS1 gene and the positions of primers for MSP. Primers positions are represented by arrows.
Figure S2 Schematic diagram of the SOCS2 gene and the positions of primers for MSP. Primers positions are represented by arrows.
Figure S3 Schematic diagram of the SOCS3 gene and the positions of primers for MSP. Primers positions are represented by arrows.
Table S1 MSP primer sequences and reaction conditions.
References
- 1.Tefferi A. Essential thrombocythemia, polycythemia vera, and myelofibrosis: current management and the prospect of targeted therapy. Am J Hematol. 2008;83:491–7. doi: 10.1002/ajh.21183. [DOI] [PubMed] [Google Scholar]
- 2.Chim CS, Kwong YL, Lie AK, et al. Long-term outcome of 231 patients with essential thrombocythemia: prognostic factors for thrombosis, bleeding, myelofibrosis, and leukemia. Arch Intern Med. 2005;165:2651–8. doi: 10.1001/archinte.165.22.2651. [DOI] [PubMed] [Google Scholar]
- 3.Tefferi A. Novel mutations and their functional and clinical relevance in myeloproliferative neoplasms: JAK2, MPL, TET2, ASXL1, CBL, IDH and IKZF1. Leukemia. 2010;24:1128–38. doi: 10.1038/leu.2010.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Vannucchi AM, Antonioli E, Guglielmelli P, et al. Clinical correlates of JAK2V617F presence or allele burden in myeloproliferative neoplasms: a critical reappraisal. Leukemia. 2008;22:1299–307. doi: 10.1038/leu.2008.113. [DOI] [PubMed] [Google Scholar]
- 5.Ward AC, Touw I, Yoshimura A. The Jak-Stat pathway in normal and perturbed hematopoiesis. Blood. 2000;95:19–29. [PubMed] [Google Scholar]
- 6.O'Shea JJ, Gadina M, Schreiber RD. Cytokine signaling in 2002: new surprises in the Jak/Stat pathway. Cell. 2002;109(Suppl):S121–31. doi: 10.1016/s0092-8674(02)00701-8. [DOI] [PubMed] [Google Scholar]
- 7.Alexander WS. Suppressors of cytokine signalling (SOCS) in the immune system. Nat Rev Immunol. 2002;2:410–6. doi: 10.1038/nri818. [DOI] [PubMed] [Google Scholar]
- 8.Myers MP, Andersen JN, Cheng A, et al. TYK2 and JAK2 are substrates of protein-tyrosine phosphatase 1B. J Biol Chem. 2001;276:47771–4. doi: 10.1074/jbc.C100583200. [DOI] [PubMed] [Google Scholar]
- 9.Kubo M, Hanada T, Yoshimura A. Suppressors of cytokine signaling and immunity. Nat Immunol. 2003;4:1169–76. doi: 10.1038/ni1012. [DOI] [PubMed] [Google Scholar]
- 10.Lin LI, Tien HF. SOCS1 (Suppressor of cytokine signaling 1) Atlas Genet Cytogenet Oncol Haematol. 2007 Available at http://atlasgeneticsoncology.org/Genes/SOCS1ID42350ch16p13.html. [Google Scholar]
- 11.Fernández-Pérez L, Flores-Morales A. SOCS2 (suppressor of cytokine signaling 2) Atlas Genet Cytogenet Oncol Haematol. 2007 Available at http://atlasgeneticsoncology.org//Genes/SOCS2ID44123ch12q21.html. [Google Scholar]
- 12.Culig Z. SOCS3 (suppressor of cytokine signaling 3) Atlas Genet Cytogenet Oncol Haematol. 2012 Available at http://atlasgeneticsoncology.org//Genes/SOCS3ID44124ch17q25.html. [Google Scholar]
- 13.Bernstein BE, Stamatoyannopoulos JA, Costello JF, et al. The NIH roadmap epigenomics mapping consortium. Nat Biotechnol. 2010;28:1045–8. doi: 10.1038/nbt1010-1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wu C, Sun M, Liu L, et al. The function of the protein tyrosine phosphatase SHP-1 in cancer. Gene. 2003;306:1–12. doi: 10.1016/s0378-1119(03)00400-1. [DOI] [PubMed] [Google Scholar]
- 15.Wang LQ, Liang R, Chim CS. Methylation of tumor suppressor microRNAs: lessons from lymphoid malignancies. Expert Rev Mol Diagn. 2012;12:755–65. doi: 10.1586/erm.12.64. [DOI] [PubMed] [Google Scholar]
- 16.Wong KY, Huang X, Chim CS. DNA methylation of microRNA genes in multiple myeloma. Carcinogenesis. 2012;33:1629–38. doi: 10.1093/carcin/bgs212. [DOI] [PubMed] [Google Scholar]
- 17.Yim RL, Kwong YL, Wong KY, et al. DNA methylation of tumor suppressive miRNAs in non-Hodgkin's lymphomas. Front Genet. 2012;3:233. doi: 10.3389/fgene.2012.00233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chim CS, Liang R, Kwong YL. Hypermethylation of gene promoters in hematological neoplasia. Hematol Oncol. 2002;20:167–76. doi: 10.1002/hon.694. [DOI] [PubMed] [Google Scholar]
- 19.Barrio S, Gallardo M, Albizua E, et al. Epigenomic profiling in polycythaemia vera and essential thrombocythaemia shows low levels of aberrant DNA methylation. J Clin Pathol. 2011;64:1010–3. doi: 10.1136/jclinpath-2011-200175. [DOI] [PubMed] [Google Scholar]
- 20.Capello D, Deambrogi C, Rossi D, et al. Epigenetic inactivation of suppressors of cytokine signalling in Philadelphia-negative chronic myeloproliferative disorders. Br J Haematol. 2008;141:504–11. doi: 10.1111/j.1365-2141.2008.07072.x. [DOI] [PubMed] [Google Scholar]
- 21.Fernández-Mercado M, Cebrián V, Euba B, et al. Methylation status of SOCS1 and SOCS3 in BCR-ABL negative and JAK2V617F negative chronic myeloproliferative neoplasms. Leuk Res. 2008;32:1638–40. doi: 10.1016/j.leukres.2008.03.014. [DOI] [PubMed] [Google Scholar]
- 22.Fourouclas N, Li J, Gilby DC, et al. Methylation of the suppressor of cytokine signaling 3 gene (SOCS3) in myeloproliferative disorders. Haematologica. 2008;93:1635–44. doi: 10.3324/haematol.13043. [DOI] [PubMed] [Google Scholar]
- 23.Jost E, do O N, Dahl E, et al. Epigenetic alterations complement mutation of JAK2 tyrosine kinase in patients with BCR/ABL-negative myeloproliferative disorders. Leukemia. 2007;21:505–10. doi: 10.1038/sj.leu.2404513. [DOI] [PubMed] [Google Scholar]
- 24.Chim CS, Fung TK, Cheung WC, et al. SOCS1 and SHP1 hypermethylation in multiple myeloma: implications for epigenetic activation of the Jak/STAT pathway. Blood. 2004;103:4630–5. doi: 10.1182/blood-2003-06-2007. [DOI] [PubMed] [Google Scholar]
- 25.Chim CS, Kwong YL. How frequent is SOCS1 methylation in acute myeloid leukaemia? Br J Haematol. 2004;127:609–11. doi: 10.1111/j.1365-2141.2004.05246.x. [DOI] [PubMed] [Google Scholar]
- 26.Chim CS, Fung TK, Liang R. Methylation of cyclin-dependent kinase inhibitors, XAF1, JUNB, CDH13 and soluble Wnt inhibitors in essential thrombocythaemia. J Clin Pathol. 2010;3:518–21. doi: 10.1136/jcp.2009.072413. [DOI] [PubMed] [Google Scholar]
- 27.Chim CS, Wan TS, Fung TK, et al. Methylation of TET2, CBL and CEBPA in Ph-negative myeloproliferative neoplasms. J Clin Pathol. 2010;63:942–6. doi: 10.1136/jcp.2010.080218. [DOI] [PubMed] [Google Scholar]
- 28.Chim CS, Wan TS, Wong KY, et al. vMethylation of miR-34a, miR-34b/c, miR-124-1 and miR-203 in Ph-negative myeloproliferative neoplasms. J Transl Med. 2011;9:197. doi: 10.1186/1479-5876-9-197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Teofili L, Martini M, Cenci T, et al. Epigenetic alteration of SOCS family members is a possible pathogenetic mechanism in JAK2 wild type myeloproliferative diseases. Int J Cancer. 2008;123:1586–92. doi: 10.1002/ijc.23694. [DOI] [PubMed] [Google Scholar]
- 30.Quentmeier H, Geffers R, Jost E, et al. SOCS2: inhibitor of JAK2V617F-mediated signal transduction. Leukemia. 2008;22:2169–75. doi: 10.1038/leu.2008.226. [DOI] [PubMed] [Google Scholar]
- 31.He B, You L, Uematsu K, et al. SOCS-3 is frequently silenced by hypermethylation and suppresses cell growth in human lung cancer. Proc Natl Acad Sci USA. 2003;100:14133–8. doi: 10.1073/pnas.2232790100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Watanabe D, Ezoe S, Fujimoto M, et al. Suppressor of cytokine signalling-1 gene silencing in acute myeloid leukaemia and human haematopoietic cell lines. Br J Haematol. 2004;126:726–35. doi: 10.1111/j.1365-2141.2004.05107.x. [DOI] [PubMed] [Google Scholar]
- 33.Nagai H, Naka T, Terada Y, et al. Hypermethylation associated with inactivation of the SOCS-1 gene, a JAK/STAT inhibitor, in human hepatoblastomas. J Hum Genet. 2003;48:65–9. doi: 10.1007/s100380300008. [DOI] [PubMed] [Google Scholar]
- 34.Födermayr M, Zach O, Huber M, et al. The clinical impact of DNA methylation frequencies of JAK2 negative regulators in patients with essential thrombocythemia. Leuk Res. 2012;36:588–90. doi: 10.1016/j.leukres.2012.01.008. [DOI] [PubMed] [Google Scholar]
- 35.Melki JR, Vincent PC, Clark SJ. Cancer-specific region of hypermethylation identified within the HIC1 putative tumour suppressor gene in acute myeloid leukaemia. Leukemia. 1999;13:877–83. doi: 10.1038/sj.leu.2401401. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.