

Abstract
In the US and globally, dramatic increases in the prevalence of adult and childhood obesity have been reported during the last 30 years. In addition to cardiovascular disease, type II diabetes, and liver disease, obesity has recently been recognized as an important risk factor for influenza pneumonia. During the influenza pandemic of 2009, obese individuals experienced a greater severity of illness from the H1N1 virus. In addition, obese mice have also been shown to exhibit increased lethality and aberrant pulmonary inflammatory responses following influenza infection. In contrast to influenza, the impact of obesity on bacterial pneumonia in human patients is controversial. In this report, we compared the responses of lean WT and obese CPEfat/fat mice following an intratracheal infection with Streptococcus pneumoniae, the leading cause of community-acquired pneumonia. At 16 weeks of age, CPEfat/fat mice develop severe obesity, hyperglycemia, elevated serum triglycerides and leptin, and increased blood neutrophil counts. There were no differences between lean WT and obese CPEfat/fat mice in survival or lung and spleen bacterial burdens following intratracheal infection with S. pneumoniae. Besides a modest increase in TNF-α levels and increased peripheral blood neutrophil counts in CPEfat/fat mice, there were not differences in lung or serum cytokines after infection. These results suggest that obesity, accompanied by hyperglycemia and modestly elevated triglycerides, at least in the case of CPEfat/fat mice, does not impair innate immunity against pneumococcal pneumonia.
Introduction
The prevalence of obesity has increased dramatically during the last three decades with 35 percent of the US adult population having a body mass index (BMI) of 30 kg/m2 or greater [1]. While obesity is recognized as a significant risk factor for type II diabetes, hypertension, and cardiovascular disease, it is also a significant contributing factor to the pathogenesis of pulmonary diseases such as asthma, obstructive sleep apnea, chronic obstructive pulmonary disease (chronic bronchitis), and a greater severity of illness and death due to influenza H1N1 [2]–[6].
The death toll from the H1N1 pandemic of 2009 has been estimated to be more than 284,000 worldwide [7]. Although the majority of cases were relatively mild and self-limiting, the severity of illness and mortality were greater among the 30–50 age group [8]. In particular, the obese were disproportionately represented among influenza-associated hospitalizations and deaths [9]. This association between obesity and severity of illness and death from H1N1 influenza has been confirmed by many other reports [10]–[14]. Furthermore, it appears that obesity is a risk factor for severity of illness from other strains of influenza and viral pathogens known to infect the respiratory tract [15], [16]. Importantly, the ability of obesity to diminish host defense against influenza infections has been confirmed in robust and carefully controlled studies using obese mice challenged with the pandemic H1N1 and H3N2 strains of the influenza virus [15], [17], [18]. Whether or not obesity is associated with a greater severity of illness with bacterial pneumonia is less certain [6], [19].
Previously, we reported that obese leptin-deficient ob/ob mice exhibited greater mortality following an intratracheal challenge with either K. pneumoniae or S.pneumoniae [20], [21]. In these studies, greater mortality in the ob/ob mouse was associated with impaired pulmonary bacterial clearance and attenuated alveolar macrophage and neutrophil phagocytosis and killing of bacteria, and the elaboration of reactive oxygen intermediates [22]. In addition, many other reports have demonstrated that ob/ob mice exhibit host defense defects in response to several other bacterial, mycobacterial, amoeba, and fungal infections [23]–[28]. However, leptin deficiency disables host defense, in the absence of obesity, and has been demonstrated to restore antimicrobial functions in the presence of obesity in ob/ob mice [21], [29]. The effect of obesity on host defense against community-acquired pneumonia in humans is controversial and appropriate animal models have not been used to address this important question [19]. In the current study, we compared the responses of lean wild type (WT) and obese CPEfat/fat mice, which lack a functional carboxypeptidase enzyme, following an intratracheal infection with Streptococcus pneumoniae, the most common cause of community-acquired pneumonia [30].
Materials and Methods
Ethics statement
All animals were treated according to National Institutes of Health guidelines for the use of experimental animals with the approval of the University of Michigan Committee for the Use and Care of Animals (Protocol Number: #PRO00003932).
Animals
Female CPEfat/fat mice, bred on a C57BL/6 background and age-matched, female C57BL/6 wild type (WT) animals, were purchased from The Jackson Laboratory, Bar Harbor, ME. All mice were 16–18 weeks of age prior to their use in all the experiments performed for these studies. All mice were maintained in the University of Michigan Unit for Laboratory Animal Medicine, maintained on Formulab 5008 rodent chow (LabDiet, Brentwood, MO).
Murine model of pneumococcal pneumonia
S. pneumoniae serotype 3, 6303 (American Type Culture Collection, Manassas, VA) was grown to mid-log phase in Todd-Hewett broth, washed in PBS, and serially diluted in sterile saline. Following anesthesia with ketamine (80 mg/kg) and xylazine (10 mg/kg) delivered via an intraperitoneal injection, a midline incision was made to expose the trachea, a 30-µl inoculum containing 50,000 CFU S. pneumoniae was administered via the trachea using a 26-gauge needle, and the wound was closed using surgical glue (Vetbond, 3 M, St. Paul, MN) [31]. Following infection, mice were warmed by placing their cage on a heating pad and closely observed every 10 min until they recovered from the anesthesia. For the duration of the lethality study, they were observed for survival every 4 hours during the day. Moribund animals (i.e. staggered gate, ruffled fur, unable to reach water or food) were euthanized by CO2 asphyxiation to ameliorate suffering. In a separate group of mice, bacterial growth in lung and spleen homogenates was determined 24 and 48 h after infection using serial dilutions plated on blood agar as previously described [29].
Lung bronchoalveolar lavage fluid (BALF) and blood cytokine determinations
BALF and blood obtained from mice 24 and 48 h after pneumococcal infection were evaluated for GM-CSF(blood only), IL-1β, IL-6, IL-10, IL-12 p70, MIP-2, MCP-1, and TNF-α by ELISA (R&D Duoset, R&D Systems) performed by the University of Michigan Cancer Center Cellular Immunology Core as previously described [32]. Leptin levels (blood only) were determined according to the manufacturer's instructions using a commercially available ELISA kit (Millipore, St. Charles, MO).
Blood glucose, serum triglycerides, and leukocyte determinations
Blood was obtained from mice by cardiac puncture following euthanasia by an overdose of CO2 for glucose measurements and leukocyte counts. Blood glucose was assessed at baseline using a glucometer (Glucometer Elite; Bayer, Elkhart, IN). Serum triglyceride levels were measured using the GPO kit from Raichem (Clinica Corp., San Marcos, CA), with glycerol as a standard according to the manufacturer's instructions. Leukocyte counts were performed after red blood cell lysis (Unopette Microcollection System; Becton-Dickson, Rutherford, NJ) and a Hemavet cell analyzer (Drew Scientific) operated by the University of Michigan Unit for Laboratory Animal Medicine Animal Diagnostic Laboratory.
Statistical analyses
Statistical analyses were conducted using Prism 6.0 software (GraphPad Software, La Jolla, CA). Survival differences were assessed using the Mantel-Cox log-rank test. Where appropriate, mean values were compared using a Student t-test. Differences were considered significant if P≤0.05. All experiments were performed at least three separate times unless otherwise noted in the figure legend. Data are presented as mean values ± standard error of the mean unless noted otherwise.
Results
Obesity and metabolic abnormalities in CPEfat/fat mice
In CPEfat/fat mice, the obese phenotype arises from the lack of carboxypeptidase E, an enzyme that plays an essential role in processing prohormones and proneuropeptides known to regulate appetite and energy expenditure [33]–[35]. As a consequence of the absence of carboxypeptidase, CPEfat/fat mice exhibit hyperphasia, reduced locomotor activity, and reduced energy expenditure compared with wild type animals [34]. As shown in Figure 1, the body weights of CPEfat/fat mice were 2-fold greater than that of WT animals and the increased body mass has previously been attributed to greater fat mass [34]. In addition, the CPEfat/fat mice were hyperglycemic at 16 wks of age and had substantially higher levels of leptin and serum (Figure 1B, C and D). Interestingly, we also observed greater total white blood cell (WBC) and neutrophil (PMN) counts in CPEfat/fat mice (Figure 1E). In total, the CPEfat/fat mouse is obese and hyperglycemic with elevated triglycerides, leptin, and peripheral WBC and PMN counts when maintained on a normal chow diet at 16 wks of age.
Figure 1. Body weight (A), blood glucose (B), serum leptin (C), triglycerides (D), and white blood cell counts (E) in 16-wk-old female wild type (WT) and carboxypeptidase E fat/fat (CPEfat/fat) mice at baseline.

*,<0.05 vs WT using student's t-test. N = 4 mice per group (A-D) and N = 10 mice per group (E).
Differences in weight loss but not survival or bacterial burdens in WT and CPEfat/fat mice following intratracheal S. pneumoniae infection
Since the risk of community-acquired pneumonia among obese individuals in clinical and epidemiologic studies is uncertain [6], we assessed survival and weight loss in WT and CPEfat/fat mice following S. pneumonia challenge. As shown in Figure 2, the differences between WT and CPEfat/fat mice in survival were modest and did not reach statistical significance (p = 0.37). However, we did find greater absolute weight loss after infection in CPEfat/fat mice at both 24 and 48 h post-infection (Figure 2B). In contrast, we did not find differences in % weight loss (from baseline) post-infection (data not shown). Since differences in survival may not reflect potential differences in host defense, we also examined lung and spleen bacterial burdens after infection. As shown in Figure 3, pulmonary and spleen bacterial loads were not different between WT and CPEfat/fat mice at 24 or 48 h after S. pneumoniae challenge. Based on these results, CPEfat/fat did not exhibit impairments in survival or pulmonary clearance of S. pneumoniae.
Figure 2. Survival of WT and CPEfat/fat mice following S. pneumoniae infection (A). Body weight loss, expressed as total weight loss, 24 and 48 h post-infection (B).

Mice were infected with 5×104 CFUs of S. pneumonia via the intratracheal route and monitored for survival for 10 days. N = 6–9 mice per group from 2 independent experiments. Survival was evaluated using the log-rank test. *, p<0.05, total weight loss for WT vs CPEfat/fat mice post-infection using a student's t-test, n = 5 (24 h) and 15 (48 h) mice per group.
Figure 3. Bacterial burdens in WT and CPEfat/fat mice 24 (A) and 48 h (B) following S. pneumoniae challenge.
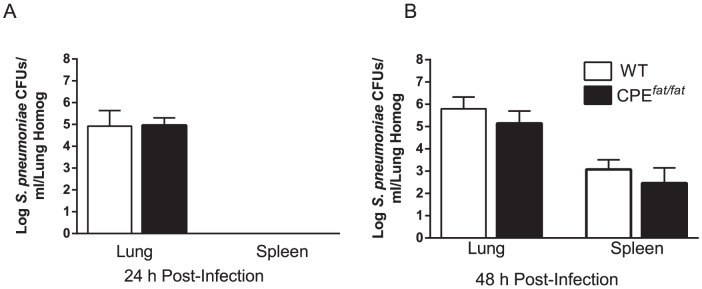
Mice were infected with 5×104 CFUs of S. pneumoniae via the intratracheal route and bacterial loads in tissue homogenates were determined by enumerating CFUs as described in the Materials and Methods section. Statistical comparisons were performed using the students t-test. n = 5 (24 h) and 15 (48 h) mice per group.
Effect of S. pneumoniae challenge on cytokines in BALF
Excess adipose tissue has been shown to produce cytokines that significantly contribute to a chronic state of low-grade systemic inflammation in obese humans and animals [36]. Whether or not pulmonary cytokine production is differentially regulated in CPEfat/fat mice during pneumococcal pneumonia has not been evaluated. Cytokine levels at baseline in BALF were below the limit of detection (data not shown). As shown in Figure 4, higher levels of TNF-α were observed in the BALF of CPEfat/fat mice with modestly elevated levels of IL-6, IL-10, IL-12, and MIP-2 that did not reach statistical significance 24 h after infection. In contrast, IL-10 and IL-12 levels were lower in CPEfat/fat mice 48 h post-infection. As was observed at 24 h, IL-6 was elevated in CPEfat/fat mice 48 h post-infection but this difference was not statistically significant. In total, there were modest differences between WT and CPEfat/fat mice in pulmonary cytokines following S. pneumoniae challenge.
Figure 4. Cytokines in bronchoalveolar lavage fluid (BALF) from WT and CPEfat/fat mice 24 (A) and 48 h (B) post-infection.
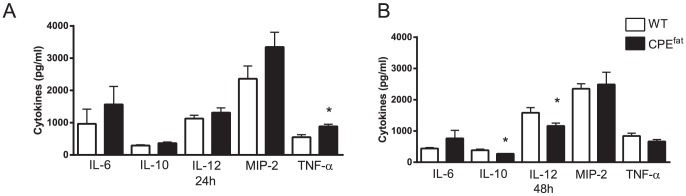
Mice were challenged with 5×104 CFUs of S. pneumoniae via the intratracheal route and BALF was obtained 24 and 48 h post- infection and cytokine levels were determined as described in the Materials and Methods section. n = 5–7 mice per group. *, p<0.05 vs WT using a student's t-test.
Impact of obesity on the systemic inflammatory response following S. pneumoniae challenge
Since the elevated levels of systemic proinflammatory cytokines (MCP-1, IL-1β, and IL-6) and increased peripheral blood leukocyte counts have been reported in obese humans and CPEfat/fat mice, we assessed these cytokines in serum and peripheral blood leukocyte counts after infection [35], [37]. Although there was a trend for elevated levels of IL-1β, IL-6, and MCP-1 in CPEfat/fat mice 24 h post infection, none of these differences reached statistically significance (Figure 5A). In addition, we did not observe differences in blood GM-CSF (data not shown). In contrast, 48 h after infection, we observed a non-significant trend for higher levels of all cytokines in WT animals which was consistent with the trend for elevated spleen bacterial counts (non-significant trend) in WT mice at this time point (Figure 5B). PMN counts were elevated in CPEfat/fat mice 48 h after infection. (Figure 5C). In total, we observed elevated PMN counts in the CPEfat/fat mice with no differences in peripheral blood cytokines during pneumococcal pneumonia.
Figure 5. Serum cytokines (A and B) and peripheral blood cell counts (C) in WT and CPEfat/fat mice following S. pneumoniae challenge.

WT and CPEfat/fat mice were challenged with 105 CFUs of S. pneumoniae via the intratracheal route and serum was prepared from blood samples obtained 24 and 48 h post-infection as mentioned in the Materials and Methods section. n = 5–7 mice per group *, p<0.05 vs WT using a student's t-test.
Discussion
Obese humans and mice are known to experience a greater severity of illness and death from influenza pneumonia [8], [15], [17], [18], [38], [39]. In this report, we compared the responses of lean and obese CPEfat/fat mice following an intratracheal challenge with S. pneumoniae. Despite severe obesity, hyperglycemia, elevated peripheral blood neutrophil counts, and modest differences in lung cytokines following infection, there were no differences between CPEfat/fat mice and WT animals in survival or pulmonary and spleen bacterial burdens following S. pneumoniae challenge. These results suggest that obesity in the CPEfat/fat mouse, even in the presence of hyperglycemia, does not impair host defense against pneumococcal pneumonia.
While we did not observe differences in bacterial burdens in obese CPEfat/fat and WT mice following S. pneumoniae challenge, it is possible that pulmonary host defense may be impaired in other models of obesity. CPEfat/fat mice become hyperphagic due to the lack of functional hormones that regulate food intake [40]. The greater weight loss in the CPEfat/fat mice was likely due to a reduction in hyperphagia since pneumonia is well known to decrease appetite [29]. However, the % weight loss from baseline, which was not different, is a better measure of the severity of infection in murine models of bacterial pneumonia. The mean blood glucose level of 16-wk-old CPEfat/fat mice was approximately 180 mg/dL. Murine diet induced obesity (DIO) is produced by feeding animals a high fat diet (40–60% kcals from fat) for several weeks (usually 10–26 wks) resulting in obesity and diabetes (blood glucose >200 mg/dL) [41], [42]. DIO mice have been shown to exhibit greater renal bacterial burdens in a model of S. aureus-induced sepsis and higher oral bacterial counts following P. gingivalus infection [43]. Diabetes, a common comorbidity of obesity, is a well-known risk factor for pneumococcal infections, and poor glucose control in diabetes is known to increase the risk of pneumococcal pneumonia [44], [45]. Hyperglycemia in type II diabetes is known to compromise host defense against cutaneous infections by impairing wound healing, antimicrobial peptide (LL-37)(cathelicidin) production, and epithelial cell proliferation following tissue injury [15], [46], [47]. Therefore, the combination of obesity and diabetes may be required to impair host defense during bacterial pneumonia.
Leukocytosis is frequently observed in obese humans and mice [37], [48]–[51]. While having a greater number of peripheral blood leukocytes, such as PMNs known to ingest and killing bacteria, may enhance host defense against bacterial infections, a profound recruitment of these cells to a site of infection may also induce collateral tissue damage that diminishes bacterial clearance [50]. The elevated PMNs in CPEfat/fat mice at baseline and after infection did not seem to contribute to differences in bacterial burdens in our study.
Excess adipose tissue may contribute to pulmonary inflammation since levels of systemic cytokines (IL-1β, IL-6, MCP-1, and TNF-α) and acute phase proteins (C-reactive protein and serum amyloid A) are elevated in obese humans and mice [52]. Johnston and co-workers reported enhanced pulmonary inflammatory responses in CPEfat/fat mice following exposure to ozone that was characterized by increased BAL protein, cytokines (IL-6, KC, MCP-1, and MIP-2), and neutrophil recruitment [35]. In addition, peripheral blood neutrophils and MCP-1 were also elevated in the CPEfat/fat mice in response to ozone. In our studies, we observed modest increases in TNF-α in BALF at 24 h and elevated peripheral blood neutrophils in CPEfat/fat mice at baseline and 48 h following an intratracheal challenge with S.pneumoniae. While there were trends for increased IL-6, IL-1β, and MCP-1 in CPEfat/fat mice 24 h after infection, these differences did not reach statistical significance. At 48 h post-infection, the lower levels of IL-10 and IL-12 and trend for reduced TNF-α in CPEfat/fat mice may have been due to the non-significant trend for lower spleen CFUs. In total, the low-grade inflammatory state observed in the CPEfat/fat mice did not affect pulmonary host defense or substantially enhance pulmonary, or systemic cytokine production following infection.
In summary, obese CPEfat/fat mice do not exhibit impairments in host defense against S. pneumoniae. These results suggest that obesity, accompanied with modest metabolic abnormalities, does not compromise pulmonary innate immunity against pneumococcal pneumonia.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are included within the paper.
Funding Statement
This research was supported by grants from the National Institutes of Health HL077417 (PM), Institutional Training Grant T32 ES007062 (EO), and the Flight Attendants Medical Research Institute CIA-103071(PM and DMA). The funders had not role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Ogden CL, Carroll MD, Kit BK, Flegal KM (2013) Prevalence of obesity among adults: United States, 2011–2012. NCHS Data Brief: 1–8. [PubMed]
- 2. Cecere LM, Littman AJ, Slatore CG, Udris EM, Bryson CL, et al. (2011) Obesity and COPD: associated symptoms, health-related quality of life, and medication use. COPD 8: 275–284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Gidding SS, Nehgme R, Heise C, Muscar C, Linton A, et al. (2004) Severe obesity associated with cardiovascular deconditioning, high prevalence of cardiovascular risk factors, diabetes mellitus/hyperinsulinemia, and respiratory compromise. J Pediatr 144: 766–769. [DOI] [PubMed] [Google Scholar]
- 4. Marcus JA, Pothineni A, Marcus CZ, Bisognano JD (2014) The role of obesity and obstructive sleep apnea in the pathogenesis and treatment of resistant hypertension. Curr Hypertens Rep 16: 411. [DOI] [PubMed] [Google Scholar]
- 5. Sideleva O, Black K, Dixon AE (2013) Effects of obesity and weight loss on airway physiology and inflammation in asthma. Pulm Pharmacol Ther 26: 455–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Mancuso P (2013) Obesity and respiratory infections: does excess adiposity weigh down host defense? Pulm Pharmacol Ther 26: 412–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Dawood FS, Iuliano AD, Reed C, Meltzer MI, Shay DK, et al. (2012) Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: a modelling study. Lancet Infect Dis 12: 687–695. [DOI] [PubMed] [Google Scholar]
- 8. Singanayagam A, Wood V, Chalmers JD (2011) Factors associated with severe illness in pandemic 2009 influenza a (H1N1) infection: implications for triage in primary and secondary care. J Infect 63: 243–251. [DOI] [PubMed] [Google Scholar]
- 9. LaRussa P (2011) Pandemic Novel 2009 H1N1 Influenza: What Have We Learned? Semin Respir Crit Care Med 32: 393–399. [DOI] [PubMed] [Google Scholar]
- 10. Centers for Disease Control and Prevention (CDC) (2009) Intensive-care patients with severe novel influenza A (H1N1) virus infection - Michigan, June 2009. MMWR Morb Mortal Wkly Rep 58: 749–752. [PubMed] [Google Scholar]
- 11.Vaillant L, La Ruche G, Tarantola A, Barboza P (2009) Epidemiology of fatal cases associated with pandemic H1N1 influenza 2009. Euro Surveill 14. [DOI] [PubMed]
- 12. Fuhrman C, Bonmarin I, Bitar D, Cardoso T, Duport N, et al. (2011) Adult intensive-care patients with 2009 pandemic influenza A(H1N1) infection. Epidemiol Infect 139: 1202–1209. [DOI] [PubMed] [Google Scholar]
- 13.Fuhrman C, Bonmarin I, Paty AC, Duport N, Chiron E, et al.. (2010) Severe hospitalised 2009 pandemic influenza A(H1N1) cases in France, 1 July-15 November 2009. Euro Surveill 15. [DOI] [PubMed]
- 14. Jain S, Kamimoto L, Bramley AM, Schmitz AM, Benoit SR, et al. (2009) Hospitalized Patients with 2009 H1N1 Influenza in the United States, April–June 2009. New England Journal of Medicine 361: 1935–1944. [DOI] [PubMed] [Google Scholar]
- 15. O′Brien KB, Vogel P, Duan S, Govorkova EA, Webby RJ, et al. (2012) Impaired wound healing predisposes obese mice to severe influenza virus infection. J Infect Dis 205: 252–261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Akiyama N, Segawa T, Ida H, Mezawa H, Noya M, et al. (2011) Bimodal effects of obesity ratio on disease duration of respiratory syncytial virus infection in children. Allergol Int 60: 305–308. [DOI] [PubMed] [Google Scholar]
- 17. Easterbrook JD, Dunfee RL, Schwartzman LM, Jagger BW, Sandouk A, et al. (2011) Obese mice have increased morbidity and mortality compared to non-obese mice during infection with the 2009 pandemic H1N1 influenza virus. Influenza Other Respir Viruses 5: 418–425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Smith AG, Sheridan PA, Harp JB, Beck MA (2007) Diet-induced obese mice have increased mortality and altered immune responses when infected with influenza virus. J Nutr 137: 1236–1243. [DOI] [PubMed] [Google Scholar]
- 19. Phung DT, Wang Z, Rutherford S, Huang C, Chu C (2013) Body mass index and risk of pneumonia: a systematic review and meta-analysis. Obesity Reviews 14: 839–857. [DOI] [PubMed] [Google Scholar]
- 20. Mancuso P, Gottschalk A, Phare SM, Peters-Golden M, Lukacs NW, et al. (2002) Leptin-deficient mice exhibit impaired host defense in Gram-negative pneumonia. J Immunol 168: 4018–4024. [DOI] [PubMed] [Google Scholar]
- 21. Hsu A, Aronoff DM, Phipps J, Goel D, Mancuso P (2007) Leptin improves pulmonary bacterial clearance and survival in ob/ob mice during pneumococcal pneumonia. Clin Exp Immunol 150: 332–339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Moore SI, Huffnagle GB, Chen GH, White ES, Mancuso P (2003) Leptin modulates neutrophil phagocytosis of Klebsiella pneumoniae. Infect Immun 71: 4182–4185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Ikejima S, Sasaki S, Sashinami H, Mori F, Ogawa Y, et al. (2005) Impairment of host resistance to Listeria monocytogenes infection in liver of db/db and ob/ob mice. Diabetes 54: 182–189. [DOI] [PubMed] [Google Scholar]
- 24. Madan R, Guo X, Naylor C, Buonomo EL, Mackay D, et al. (2014) Role of Leptin-Mediated Colonic Inflammation in Defense against Clostridium difficile Colitis. Infect Immun 82: 341–349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Ordway D, Henao-Tamayo M, Smith E, Shanley C, Harton M, et al. (2008) Animal model of Mycobacterium abscessus lung infection. J Leukoc Biol 83: 1502–1511. [DOI] [PubMed] [Google Scholar]
- 26. Wieland CW, Florquin S, Chan ED, Leemans JC, Weijer S, et al. (2005) Pulmonary Mycobacterium tuberculosis infection in leptin-deficient ob/ob mice. Int Immunol 17: 1399–1408. [DOI] [PubMed] [Google Scholar]
- 27. Guo X, Roberts MR, Becker SM, Podd B, Zhang Y, et al. (2011) Leptin signaling in intestinal epithelium mediates resistance to enteric infection by Entamoeba histolytica. Mucosal Immunol 4: 294–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Fernandez-Riejos P, Najib S, Santos-Alvarez J, Martin-Romero C, Perez-Perez A, et al. (2010) Role of leptin in the activation of immune cells. Mediators Inflamm 2010: 568343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Mancuso P, Huffnagle GB, Olszewski MA, Phipps J, Peters-Golden M (2006) Leptin corrects host defense defects after acute starvation in murine pneumococcal pneumonia. Am J Respir Crit Care Med 173: 212–218. [DOI] [PubMed] [Google Scholar]
- 30. Bartlett JG (2011) Diagnostic tests for agents of community-acquired pneumonia. Clin Infect Dis 52 Suppl 4S296–304. [DOI] [PubMed] [Google Scholar]
- 31. Mancuso P, Peters-Golden M, Goel D, Goldberg J, Brock TG, et al. (2011) Disruption of leptin receptor-STAT3 signaling enhances leukotriene production and pulmonary host defense against pneumococcal pneumonia. J Immunol 186: 1081–1090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Phipps JC, Aronoff DM, Curtis JL, Goel D, O′Brien E, et al. (2010) Cigarette smoke exposure impairs pulmonary bacterial clearance and alveolar macrophage complement-mediated phagocytosis of Streptococcus pneumoniae. Infect Immun 78: 1214–1220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Naggert JK, Fricker LD, Varlamov O, Nishina PM, Rouille Y, et al. (1995) Hyperproinsulinaemia in obese fat/fat mice associated with a carboxypeptidase E mutation which reduces enzyme activity. Nat Genet 10: 135–142. [DOI] [PubMed] [Google Scholar]
- 34. Cawley NX, Zhou J, Hill JM, Abebe D, Romboz S, et al. (2004) The carboxypeptidase E knockout mouse exhibits endocrinological and behavioral deficits. Endocrinology 145: 5807–5819. [DOI] [PubMed] [Google Scholar]
- 35. Johnston RA, Theman TA, Shore SA (2006) Augmented responses to ozone in obese carboxypeptidase E-deficient mice. Am J Physiol Regul Integr Comp Physiol 290: R126–133. [DOI] [PubMed] [Google Scholar]
- 36. Xu H, Barnes GT, Yang Q, Tan G, Yang D, et al. (2003) Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest 112: 1821–1830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Herishanu Y, Rogowski O, Polliack A, Marilus R (2006) Leukocytosis in obese individuals: possible link in patients with unexplained persistent neutrophilia. Eur J Haematol 76: 516–520. [DOI] [PubMed] [Google Scholar]
- 38. Morgan OW, Bramley A, Fowlkes A, Freedman DS, Taylor TH, et al. (2010) Morbid obesity as a risk factor for hospitalization and death due to 2009 pandemic influenza A(H1N1) disease. PLoS One 5: e9694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Paich HA, Sheridan PA, Handy J, Karlsson EA, Schultz-Cherry S, et al. (2013) Overweight and obese adult humans have a defective cellular immune response to pandemic H1N1 Influenza a virus. Obesity(Silver Spring) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Cawley NX, Wetsel WC, Murthy SR, Park JJ, Pacak K, et al. (2012) New roles of carboxypeptidase E in endocrine and neural function and cancer. Endocr Rev 33: 216–253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Surwit RS (1988) Diet-induced type II diabetes in C57BL/6J mice. Diabetes (New York, NY) 37: 1163–1167. [DOI] [PubMed] [Google Scholar]
- 42. Hariri N, Thibault L (2010) High-fat diet-induced obesity in animal models. Nutrition Research Reviews 23: 270–299. [DOI] [PubMed] [Google Scholar]
- 43. Strandberg L, Verdrengh M, Enge M, Andersson N, Amu S, et al. (2009) Mice Chronically Fed High-Fat Diet Have Increased Mortality and Disturbed Immune Response in Sepsis. PLoS ONE 4: e7605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Seminog OO, Goldacre MJ (2013) Risk of pneumonia and pneumococcal disease in people with severe mental illness: English record linkage studies. Thorax 68: 171–176. [DOI] [PubMed] [Google Scholar]
- 45. Rueda AM, Ormond M, Gore M, Matloobi M, Giordano TP, et al. (2010) Hyperglycemia in diabetics and non-diabetics: effect on the risk for and severity of pneumococcal pneumonia. J Infect 60: 99–105. [DOI] [PubMed] [Google Scholar]
- 46. Rivas-Santiago B, Trujillo V, Montoya A, Gonzalez-Curiel I, Castaneda-Delgado J, et al. (2012) Expression of antimicrobial peptides in diabetic foot ulcer. J Dermatol Sci 65: 19–26. [DOI] [PubMed] [Google Scholar]
- 47. Cianfarani F, Toietta G, Di Rocco G, Cesareo E, Zambruno G, et al. (2013) Diabetes impairs adipose tissue-derived stem cell function and efficiency in promoting wound healing. Wound Repair Regen 21: 545–553. [DOI] [PubMed] [Google Scholar]
- 48. Louriz M, Mahraoui C, Azzouzi A, El Fassy Fihri MT, Zeggwagh AA, et al. (2010) Clinical features of the initial cases of 2009 pandemic influenza A (H1N1) virus infection in an university hospital of Morocco. Int Arch Med 3: 26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Nanji AA, Freeman JB (1985) Relationship between body weight and total leukocyte count in morbid obesity. Am J Clin Pathol 84: 346–347. [DOI] [PubMed] [Google Scholar]
- 50. Hodgson KA, Govan BL, Walduck AK, Ketheesan N, Morris JL (2013) Impaired Early Cytokine Responses at the Site of Infection in a Murine Model of Type 2 Diabetes and Melioidosis Comorbidity. Infection and Immunity 81: 470–477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Faggioni R, Jones-Carson J, Reed DA, Dinarello CA, Feingold KR, et al. (2000) Leptin-deficient (ob/ob) mice are protected from T cell-mediated hepatotoxicity: Role of tumor necrosis factor alpha and IL-18. PNAS 97: 2367–2372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Tilg H, Moschen AR (2008) Role of adiponectin and PBEF/visfatin as regulators of inflammation: involvement in obesity-associated diseases. Clin Sci (Lond) 114: 275–288. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are included within the paper.