

Abstract
Work over the past two decades has led to substantial changes in our understanding of dystonia pathophysiology. Three general abnormalities appear to underlie the pathophysiological substrate. The first line is a loss of inhibition. This makes sense considering that it may be responsible for the excess of movement and for the overflow phenomena seen in dystonia. A second abnormality is sensory dysfunction which is related to the mild sensory complaints in patients with focal dystonias and may be responsible for some of the motor dysfunction. Third, evidence from animal models of dystonia as well as from patients with primary dystonia has revealed significant alterations of synaptic plasticity characterised by a disruption of homeostatic plasticity, with a prevailing facilitation of synaptic potentiation, together with the loss of synaptic inhibitory processes. We speculate that during motor learning this abnormal plasticity may lead to an abnormal sensorimotor integration leading to consolidation of abnormal motor engrams. If so, then removing this abnormal plasticity might have little immediate effect on dystonic movements because bad motor memories have already been “learned” and are difficult to erase. These considerations might explain the delayed clinical effects of DBS in patients with generalized dystonia. Current lines of research will be discussed from a network perspective.
Introduction
There have been great strides in the pathophysiology of dystonia in the last two decades. Three general abnormalities appear to underlie the physiological substrate of dystonia. All three are persuasive and may relate to each other. One abnormality is loss of inhibition.1 The lack of inhibition makes sense considering that it may be responsible for the excess of movement and for the overflow phenomena seen in dystonia. A second abnormality is sensory dysfunction. There are mild sensory findings in patients and the sensory system can drive the motor system. Third, there is a derangement of plasticity in dystonia. In particular, robust evidence from experimental and clinical work supports the hypothesis that dystonia may be considered a disorder linked to the disruption of homeostatic plasticity, with a prevailing facilitation of synaptic potentiation, together with the loss of synaptic inhibitory processes.2, 3. However, despite these findings, an important gap remains in the translation of these insights into an understanding of the fundamental changes in higher-order motor control that underlie dystonic symptoms especially at a network and system level. Therefore, we will discuss the current lines of research in dystonia with an integrative perspective.
Loss of inhibition and the excess of movement
A characteristic clinical feature of dystonia is the excess of movement. This has been demonstrated with electromyographic (EMG) recordings that have shown abnormally long bursts of EMG activity, co contraction of antagonist muscles, and an overflow of activity into muscles not involved in the task.4 Alterations of inhibitory circuits have been reported at the spinal cord, brainstem, and cortex.5 Transcranial magnetic stimulation (TMS) allows the study of different inhibitory circuits at the cortical level.6 Many of these are impaired in dystonia and the abnormal intracortical inhibition may be found in both hemispheres despite unilateral symptoms and even in asymptomatic body parts.7–12 The alterations are nonspecific in that they have also been observed in various other neurological conditions and even in psychogenic dystonia.12, 13 For this reason, there does not appear to be a direct link between reduced intracortical inhibition and dystonia.14. It is also fair to note that physiological abnormalities in asymptomatic body parts could indicate that they are compensatory changes to prevent dystonia; however, this seems unlikely since the abnormalities are generally the same as those in the symptomatic body parts and are in the direction to lead to motor dysfunction.
How can a nonspecific deficit of inhibitory intracortical circuits translate into the typical dystonic unfocussed muscular activation? It is likely that when a specific voluntary movement is generated, the brain has to suppress other possible movements. In this way, the motor cortex can produce a more accurate movement, just as surround inhibition in sensory systems allows a more exact perception.15 There is good evidence for surround inhibition, within sensory-motor cortex, in human movement.16,17
Although it cannot be demonstrated in every normal subject.18 Surround inhibition is reduced in focal hand dystonia and this may contribute to the difficulty in focusing motor command and to overflow phenomena.17, 19, 20 An alteration of surround inhibition can also be demonstrated when dystonic patients imagine abduction of the index finger.21 (fig.1).
Fig. 1. Surround inhibition during motor imagery.
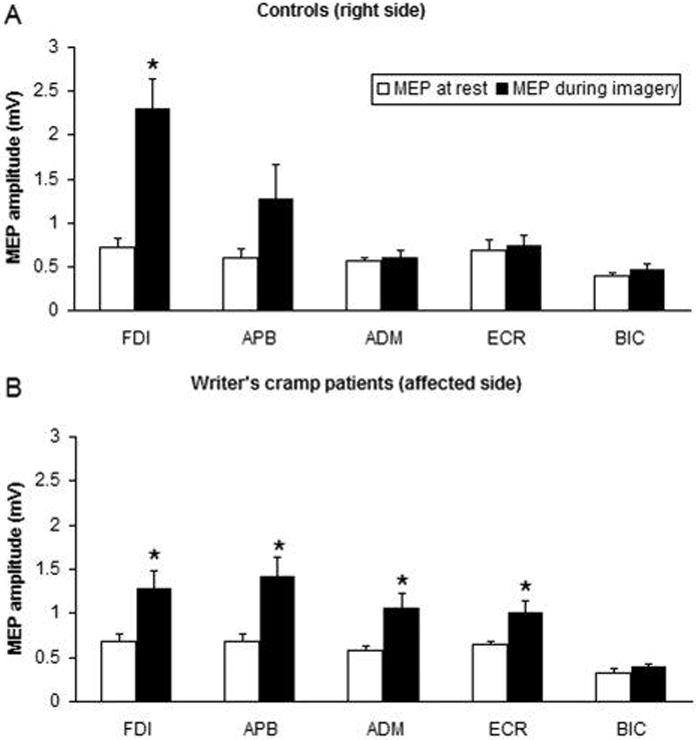
Changes in mean motor evoked potential (MEP) size during motor imagery on the right side of healthy controls and on the affected side of patients. TMS was always given to the motor cortex contralateral to the imagined task. MEP size during motor imagery (MI) compared with rest condition recorded from different target muscles of the right upper limb after stimulation of the left hemisphere in controls (A) and patients with writer’s cramp (B). The bar chart illustrates the mean peak-to-peak amplitude (mV) of MEPs recorded at rest (open columns) and during MI (black columns). Each error bar equals standard error of the mean (SEM). MI elicited an attenuated and less focal increase in MEP amplitude in patients than in controls. FDI, first dorsal interosseus; APB, abductor pollicis brevis; ADM, abductor digiti minimi; ECR, extensor carpi radialis; BIC, biceps. *P < 0.05. (from: Quartarone A, Bagnato S, Rizzo V, Morgante F, Sant’Angelo A, Crupi D, Romano M, Berardelli A, Girlanda P. Corticospinal excitability during motor imagery of a simple tonic finger movement in patients with writer’s cramp. Mov Disord. 2005 Nov;20(11):1488–95).
Considerable work has been done trying to determine the mechanism of surround inhibition. The normal mechanisms are not determined yet, but there are derangements in focal hand dystonia including short intracortical inhibition.22
Loss of inhibition: neuroanatomical correlates
The loss of inhibition has been traditionally interpreted in the context of presumed basal ganglia dysfunction. One hypothesis about basal ganglia organization is that the direct pathway helps command the desired movement, while the indirect pathway inhibits unwanted movements.15 A number of investigators have felt that there is an imbalance in the direct and indirect pathways so that the direct pathway is relatively overactive (or that the indirect pathway is relatively underactive)23. The postulated imbalance should lead to excessive movement and, in particular, a 1oss of surround inhibition. The cerebellum may play a role given its influence on cortical excitability.24–26 Transcranial magnetic stimulation of the cerebellum decreases cortical excitability in normal humans, an effect that is lost in patients with dystonia27, but it does not appear to play a role in surround inhibition.28 At present there is no conclusive evidence linking the loss of inhibition to a specific dysfunction within basal ganglia or cerebellar network. The failure of SICI in focal hand dystonia suggests that there might well be a cortical abnormality of intracortical inhibitory neurons. There was a suggestion of a loss of GABA in the sensorimotor cortex demonstrated with magnetic resonance spectroscopy,29 but this was not confirmed later in a subsequent study.30 Another study, however, showed a loss of flumazenil binding in the sensorimotor cortex, keeping the idea plausible.31
Sensory abnormalities
Another major theme in the pathophysiology of dystonia is a defect in sensory or perceptual function or in “sensorimotor integration”. Although dystonia is generally regarded as a pure32 motor disorder, it is commonly preceded by sensory symptoms. Ill-defined bodily feelings (discomfort, pain, or kinaesthetic sensations) are frequently reported weeks or months before dystonia develops33 and pain can be a component of cervical dystonia. There are also mild sensory deficits. Patients with focal dystonia have difficulty in discriminating sensory stimuli in both spatial and temporal domains.34 Importantly, these abnormalities are present in unaffected body parts such as the unaffected hand in patients with focal hand dystonia. For instance it has been reported that somatosensory temporal discrimination threshold (STDT) is abnormal in all the different forms of primary focal dystonias in all three body regions (eye, hand and neck), regardless of the distribution and severity of motor symptoms.35. These sensory abnormalities, particularly of temporal discrimination,36 have also been documented in some relatives of patients with inherited generalized dystonia.37 These alterations may be related to a deranged somatotopic representation in the sensory cortex as revealed by neurophysiological and neuroimaging studies.36, 38–42
In addition, sensory inputs may modulate dystonic symptoms. A well recognized clinical sign in dystonia is the sensory trick, particularly in cervical dystonia, where the symptoms may improve upon touching the cheek. Tonic vibration of a limb may induce a worsening of hand dystonia whereas anesthetic block may relieve it.43
Sensorimotor integration, which refers to how the sensory system interacts with the motor system, is also abnormal in dystonia.44 For example, sensory modulation in response to movement, so-called sensory gating, is abnormal in focal hand dystonia.
Sensory abnormalities: neuroanatomical correlates
The basal ganglia may play a role in sensory and perceptual defects. Given the influence of striatum over basal ganglia output (via the GPi/SNr) to thalamus and back to motor cortical areas, the basal ganglia have been attributed a role in “sensory gating”, filtering out what sensory information is “passed” to the motor system.45, 46 In this respect, the role played by cholinergic interneurons cannot be neglected. Although they account for 2% or less of the neuronal striatal population, these interneurons have extensive dendritic arbors and are unique for their responsiveness in classical conditioning tasks. Their activity appears to be unrelated to movements while they discharge phasically in response to sensory stimuli serving as a cue for reward delivery and consumption.47 Moreover, thalamic projections to the striatum engage cholinergic interneurons to modulate corticostriatal inputs, thereby supporting their fundamental role in filtering excitatory afferents.48 Cholinergic dysfunction must be important in dystonia considering that partially effective therapies for the disorder are anticholinergic drugs.49 On the other hand, the cerebellum also exerts powerful influences over the somatosensory system. For instance, alterations of cortical excitability after repetitive somatosensory stimulation are mediated by the cerebellum.50, 51 Unlike the basal ganglia that receive sensory information indirectly, the cerebellum is the direct recipient of sensory input from spinal cord. In this way cerebellum may affect somatosensory threshold in the cortex playing also a role in both temporal and spatial discrimination.52, 53
There is also evidence for an abnormality directly in the sensory cortex. Studies using the somatosensory evoked potential recovery curve show that there is a abnormality of short latency intracortical inhibitory mechanisms similar to those in the motor system.32
In addition, studies in patients with task specific dystonia using somatosensory evoked potentials and magnetoencephalography have revealed a dramatic disorganization of the normal homuncular finger representations of both hands in the primary sensory cortex. The abnormal cortical finger representations, which may well arise from a lack of surround inhibition, appear to be an endophenotypic trait of dystonia.39, 42
Maladaptive plasticity in dystonia
Maladaptive plasticity appears to be an important feature for the pathogenesis of dystonia.2, 3, 54 The task specificity in focal dystonia suggests a breakdown in the circuits involved in the encoding of motor memories, which produces abnormal motor engrams. This can also explain the patterned muscular activation, which is typical of dystonia. An important clinical feature is that typically hand dystonia is triggered by period of intensive training of a particular movement.2, 55
In a monkey model of dystonia, overtraining in a specific hand movement induced the appearance of a motor hand impairment similar to problems experienced by patients with focal hand dystonia.56 The somatosensory cortex of these animals was less well organized than that of healthy monkeys, with larger receptive fields and overlapping representations of the individual digits. Therefore it can be postulated that overtraining itself may induce a change in the connectivity in the sensory and motor cortices leading to inappropriate association between sensory input and motor outputs, which in turn would cause errors in selecting muscles, used in voluntary movement.57 However, this experiment showed only that severe over-training could lead to abnormal reorganization of the sensorimotor cortex and dystonia. It does not give clues as to why in humans only some subjects develop dystonia after excessive training whereas others are completely healthy. It seems likely that subtle abnormalities of plasticity may render some individuals susceptible to dystonia if plastic changes are pushed to their extreme by frequent repetition. This suggests a two-factor hypothesis: use dependent environmental factors such as repetitive training and abnormal mechanisms of plasticity within sensorimotor loops.2, 58–60
There is considerable evidence suggesting that both the motor and sensory cortex in primary dystonia exhibits an exaggerated responsiveness to TMS conditioning protocols.59, 61–64
A well-established approach to test plasticity in humans in a non-invasive way is paired associative stimulation (PAS). By using PAS, it has been demonstrated that both LTP-like and LTD-like facilitatory and inhibitory effects on TMS-evoked motor evoked potentials (MEPs) recorded from the target muscle are enhanced in writer’s cramp patients.59, 62
An important feature of PAS-induced associative plasticity in healthy controls is input specificity as PAS after effects are largely confined to the cortical target representation receiving a dual congruent input.65 Instead in writer’s cramp patients, PAS tends to change cortical excitability also of nearby muscle representations (Fig. 2).59, 62, 63,66 This loss of spatial specificity appears to be a relevant finding and could be related to the abnormalities of neuronal inhibition identified previously both in the motor and somatosensory system in dystonic patients.66 These data suggest a failure of GABAergic mechanisms that are recruited during LTP-LTD like phenomena within sensory and motor cortices, and it may be speculated that this phenomenon could underlie the loss of spatial specificity of PAS-induced after effects.54 This abnormal plasticity is not confined to the neural circuits affected by dystonia but is generalized across the entire sensorimotor system representing an endophenotypic trait of dystonia.63 Further evidence of abnormal plasticity of sensorimotor circuits is that cortical excitability, probed by the somatosensory evoked response P27 was enhanced by PAS more in focal hand dystonia than in healthy subjects.61
Fig.2. Paired associative stimulation.
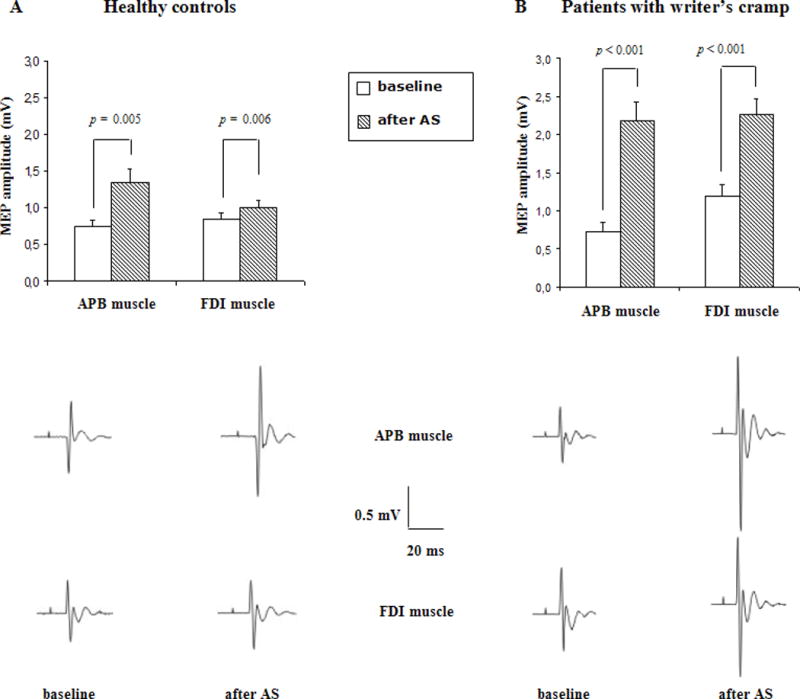
Effect of associative stimulation (PAS) on the size of motor evoked potentials (MEPs) of the right APB and FDI muscle in ten patients with writer’s cramp (right panel) and ten healthy controls (left panel). The bar chart illustrates the mean peak-to-peak amplitude (mV) of MEPs recorded at rest before (open columns) and after associative stimulation (shaded columns). Each error bar equals standard error of the mean (SEM). Representative examples of MEPs evoked in the right APB and FDI muscles are given for each group below each bar chart. Each trace represents an average of five consecutive MEPs. Associative stimulation led to an increase in MEPs size in patients and controls. However, the facilitatory effect was significantly stronger in patients. (from: Quartarone A, Bagnato S, Rizzo V, Siebner HR, Dattola V, Scalfari A, Morgante F, Battaglia F, Romano M, Girlanda P. Abnormal associative plasticity of the human motor cortex in writer’s cramp. Brain. 2003 Dec;126(Pt 12):2586–96).
The alterations of plasticity are not only at cortical level but may also be found at brainstem level. An excess of plasticity can also be found within the blink reflex circuits in patients with blepharospasm.67 This finding was not confirmed by a subsequent study perhaps due to some methodological differences.68
Another paradigm, which can induce plasticity in human cortex, is theta burst stimulation. In one study, there was a loss of TBS after-effect in non-manifesting DYT1 gene carriers, and the investigators speculated that this might protect them from developing dystonia.64 On the other hand, there is impaired motor learning in non-manifesting carriers in behavioural studies.69 The presence of abnormal motor learning even in unaffected carriers strongly suggests that abnormal plasticity is an important endophenotypic trait that might predispose to the subsequent development of dystonia.69
Abnormal plasticity in dystonia: Disruption of synaptic homeostasis
What drives cortical and brainstem plasticity beyond its physiological boundaries? Homeostatic plasticity is an essential requirement to maintain overall synaptic weight in neuronal networks within a useful dynamic range.2, 70 Theoretically, the positive-feedback nature of LTP may potentially trigger an uncontrolled increase in synaptic effectiveness, which can empower and destabilize neural circuits. Evidence in experimental models suggests that this can be prevented by making the amount of LTP dependent on the level of activity in post-synaptic neuron: the greater the ongoing activity, the less effective are processes leading to LTP, whilst processes leading to LTD are enhanced. Conversely, the lower the activity of the postsynaptic neurons, the more effective are processes that lead to LTP. This is recognized as homeostatic plasticity and is formalized in the model described by Bienenstock et al 1982.71 Enhanced plasticity in dystonia might well be the result of a disruption of homeostatic plasticity within sensorimotor circuits. It is possible to study homeostatic mechanisms by using a combined protocol where transcranial direct current stimulation (tDCS) primes the response of the motor cortex to a subsequent period of rTMS.72 In healthy controls preconditioning of the primary motor cortex with 10 minutes of anodal stimulation ca n potentate the inhibitory after-effects of 1 Hz rTMS while 10 minutes of cathodal stimulation changes the effect to facilitation. In patients with focal hand dystonia the response to 1 Hz rTMS was unaffected by preconditioning with TDCS. Specifically, 1 Hz rTMS failed to counteract the increase in cortical excitability induced by anodal TDCS.73 (fig.3)
Fig.3. Homeostatic plasticity.
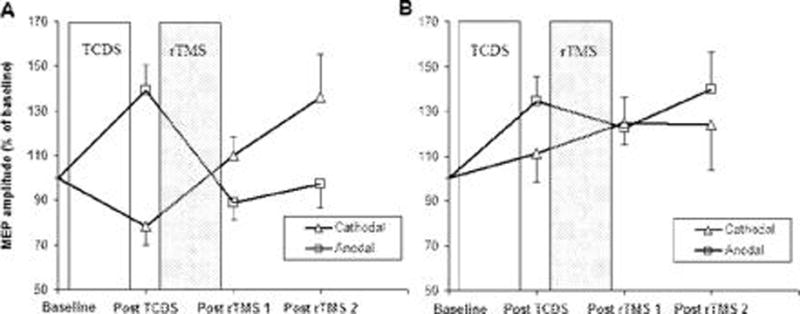
Panel A illustrates the mean amplitude of MEPs after two types of conditioning in healthy controls. There was a “facilitatory” response to anodal TDCS pre-conditioning which was reversed by a subsequent period of 1 Hz rTMS. Conversely, “inhibitory” pre-conditioning with cathodal TDCS resulted in an opposite after effect of 1 Hz rTMS which led to an increase in corticospinal excitability.
Panel B plots the changes in corticospinal excitability in writer’s cramp patients after the two types of conditioning. Patients showed an abnormal responsiveness to TDCS and rTMS. (i) Only anodal TDCS produced a normal facilitatory effect on corticospinal excitability, whereas “inhibitory” cathodal TDCS had no after effect on corticospinal excitability. (ii) Regardless of the type of pre-conditioning, rTMS had no consistent effect on corticospinal excitability. In particular, although anodal TDCS produced a “normal” facilitatory response, subsequent 1 Hz rTMS did not reverse the increase in excitability produced by anodal pre-conditioning. MEP amplitudes are given as a percentage of the MEP size at baseline. Each error bar equals standard error of the mean (SEM). (from: Quartarone A, Rizzo V, Bagnato S, Morgante F, Sant’Angelo A, Romano M, Crupi D, Girlanda P, Rothwell JC, Siebner HR. Homeostatic-like plasticity of the primary motor hand area is impaired in focal hand dystonia. Brain. 2005 Aug;128(Pt 8):1943–50)
To what extent deficient homeostatic metaplasticity applies also to the behavioural level of practice dependent plasticity induced by repetitive and highly stereotyped finger movements has also been addressed. In healthy subjects priming with an excitability-enhancing TMS protocol down regulates practice-dependent plasticity, whereas preconditioning with an excitability-depressing protocol up regulates it.74 In patients with FHD the homeostatic modulation was deficient and the magnitude of this deficiency correlated with clinical severity of the focal hand dystonia.74
Abnormal plasticity: neuroanatomical correlates
There are indirect data suggesting abnormal plasticity within basal ganglia circuits in dystonia. There is excessive oscillatory activity between 3 and 10 Hz in the local field potentials of the pallidum in patients with primary dystonia.75, 76 The magnitude of the pallidal oscillatory activity has been correlated with involuntary dystonic muscle activity.77 Excessive synchrony of neuronal firing within the basal ganglia thalamus and cortex could interfere with correct spatiotemporal processing of sensory input and motor output required for normal voluntary movement. The abnormal oscillatory activity and excessive neuronal synchrony could also be the substrate of aberrant synaptic plasticity within the basal ganglia and cortex. Such a possibility seems likely given that an increase of neuronal oscillations may facilitate cortical plasticity.78
Deep brain stimulation to the GPi can be a very powerful treatment for dystonia79, 80 and recent long-term results demonstrate that benefits are maintained after more than 10 years.81 It is possible that DBS alleviates dystonic symptoms by desynchronizing the excessive synchronized pallidal activity thus restoring a normal plasticity within the sensorimotor loop. An intriguing point is that in contrast to the almost immediate effects of DBS on the majority of symptoms in Parkinson’s disease, it may take several months to achieve maximum clinical benefit in patients with dystonia.80, 82–84 The gradual clinical improvement is paralleled by a similar normalization of several electrophysiological measures of motor inhibition in the brain and spinal cord.82, 85, 86 The slowly progressive nature of changes in clinical status together with the electrophysiological effects, suggests that a process of progressive plasticity and neural reorganization accompanies the long-term effects of globus pallidus DBS. The changes of PAS LTP-like synaptic plasticity after DBS surgery follows an interesting course. After surgery, the plasticity reduces well below normal and then gradually increases toward normal levels at 6 months. If the enhanced plasticity causes inappropriate association between sensorimotor inputs and outputs2, then removing excess plasticity is good, but might have little immediate effect on dystonic movements because bad motor memories have already been “learned” and take time to be reduced.
Finally there is also evidence suggesting that there may be functional and structural changes in the cerebellum of patients with adult onset primary focal dystonia.87, 88 The cerebellum may be involved in abnormalities of sensorimotor integration in dystonia. Patients with focal dystonia show abnormalities of the classic eyeblink conditioning paradigm, which depends on the cerebellum.89 How we can distinguish cause from consequences in physiological studies? Many of the physiological findings appear to be an underlying cause, or etiological factor, of dystonia rather than a consequence or a compensation. This is because they can be seen in asymptomatic body parts, such as in the unaffected arm in a patient with focal hand dystonia or in an arm of a patient with cervical dystonia. Some abnormalities, such as in temporal discrimination, have been identified in fully normal individuals who might be gene carriers. Only very little has been associated only with dystonia. One such example is long afferent inhibition (LAI) which is reduced in patients with writer’s cramp but only on the symptomatic side during muscle activation suggesting a tight relationship with overflow of muscle activity which characterize this condition.90 Concerning plasticity abnormalities, it could be hypothesized that the enhanced facilitation after PAS25 observed in patients with focal hand dystonia is determined by the abnormal dystonic posture. In this case, the enhanced PAS after-effects may be simply due to an activity-dependent lateral shift of the synaptic modification threshold between enhancing and suppression conditions. According to the homeostatic rule, the abnormal hand posture would reduce the efficacy of PAS10 to induce LTD changes. However this was not the case in focal hand dystonia where PAS-induced LTD was also enhanced.62 Additionally, PAS abnormalities can be detected in patients with cranio-cervical dystonia in the unaffected hands.63 Moreover, PAS25-induced plasticity was normal in patients with psychogenic dystonia even in the presence of a fixed dystonic posture.13 These findings support the notion that also the abnormal plasticity is a primary intrinsic abnormality, which can predispose to the development of dystonic movements.
Pathophysiology of dystonia: putting neurophysiological abnormalities in a network
In the past there has been a tendency to ascribe behavioral deficits in patients with movement disorders to pathological deficits in a single node of the motor pathways. However, more recent work at the systems level has focused on how healthy nodes of the brain at a distance from the primary pathology react to the damage. Such plastic reorganization in central nervous system (CNS) pathways can either compensate or exacerbate the primary deficit. In this perspective, despite the traditional pathophysiological models indicating the basal ganglia as a key structure in the pathophysiology of dystonia, it is becoming clear that many others brain regions can contribute including cerebral cortex, cerebellum, thalamus and brainstem. In keeping with this idea, dystonia may result either from a single node dysfunction, from an involvement of multiple nodes or from aberrant communication among the nodes.91, 92 The delay between a lesion and the emergence of secondary dystonia suggests the possibility of a progressive maladaptive plasticity in remote nodes. Similarly the delayed effects of DBS are in keeping with a massive rearrangement within multiple nodes of the motor loop.93, 94 Finally, dystonia may arise from abnormal communications among nodes. Dystonias secondary to thalamic lesions are a good example considering that the thalamus is a region where pallidal and cerebellar afferents may communicate before reaching the cortex.95 Abnormal communication between nodes has been documented with fMRI and EEG. Resting state fMRI, for example, shows reduced connectivity between parietal and dorsal premotor area.96 EEG has been recently used to investigate functional connectivity at rest and during a finger tapping task in patients with FHD. Using mutual information, which is a statistical approach looking at linear and non linear coupling in the various EEG bands, there was a significant reduction of beta band connectivity within sensorimotor area in patients with FHD. Graph theoretical analysis showed decreased efficiency of the beta band network taken overall.97 DBS studies report increased oscillatory activity in the frequency range of 3–20 Hz.75 These data indicate an abnormal brain coupling within motor loop which could underlie the abnormal sensorimotor plasticity. This network model with alterations in single and or multiple nodes could explain and reconcile the different lines of research in the pathophysiology of dystonia: abnormal sensorimotor integration, loss of inhibition and aberrant plasticity. Indeed the model which emerges is one in which misprocessing of sensory feedback coupled with an abnormal excitability within inhibitory motor circuits at different level (spinal cord, brainstem, cerebellum, basal ganglia and cortex) may result in a progressive abnormal plasticity in local and distant nodes culminating in an overt dystonia. This model could also explain the typical clinical course of primary dystonia with onset in one body part and subsequent gradual spread to adjacent body regions, a time course in keeping with progressive aberrant plasticity across the different nodes. The current challenge is to determine whether the alterations across different nodes (spinal cord, somatosensory, basal ganglia, cerebellum, cortical) are causative, compensatory or epiphenomenal.
How does understanding the physiology help us to think about new treatments?
Reshaping the disordered sensory somatotopy in focal dystonias.
Focal hand dystonia is difficult to treat pharmacologically or with injections of botulinum toxin, and an alternative form of treatment is clearly needed. The identification of risk factors along with putative mechanisms has prompted the design of novel therapeutic strategies guided by physiological findings. Since repetitive use of the hand is a crucial factor in the triggering and producing focal hand dystonia, training the hand back to normal is an available option. Sensorimotor retuning is where the healthy fingers are restrained using splints while the affected fingers are engaged in intensive sequential movements. This approach led to an improvement in dystonic symptoms as well as a reshaping of the disordered sensory somatotopy.98 Braille reading, another way of sensory training, has been shown to improve spatial discrimination as well as symptoms in focal dystonia.99 Motor training in the form of retraining individuated finger movements led to a mild improvement in handwriting in patients with writer’s cramp.100, 101 However, none of the treatments reported have led to sustained long term benefit. Since there is impairment in the homeostatic regulation of practice dependent plasticity, treatment protocols that try to retrain movements through intensive practice potentially bear the risk of overloading the system perpetuating a vicious cycle of runaway abnormal plasticity.2 If the loss of inhibition is responsible for the excess of movement in dystonia, then boosting up inhibition might be helpful. Repetitive TMS (rTMS) delivered over M1 at slow rates (between 0.2 and 1 Hz) can induce an increase in inhibition. As a result, rTMS has been a potential candidate to reduce this abnormal cortical excitability. A study delivering 1 Hz rTMS over M1 showed a restoration of intracortical inhibition in dystonic patients associated with a mild improvement in motor performance.102 Premotor cortex is another possible target for neurostimulation since rTMS at 1 Hz can improve the deficit of reciprocal inhibition in dystonia.11 Stimulation of the premotor cortex but not M1 significantly improved the handwriting in the patient group, which lasted up to a few hours in most patients.46 This improvement was not seen in patients receiving sham stimulation. Unfortunately, these promising results have not yet led to a subsequent multiple-session study in focal hand dystonia. Transcranial direct current stimulation (tDCS) is another possible approach to treat patients with dystonia. A randomized, double blind, sham-controlled study investigated the efficacy of cathodal stimulation of the contralateral motor cortex in 3 sessions distributed in 1 week. Cathodal transcranial direct current stimulation had no favourable effects on clinical scales and failed to restore normal handwriting kinematics and cortical inhibition.103 Continuous theta burst TMS over premotor cortex increased short-interval intracortical inhibition and the third phase of spinal reciprocal inhibition, which are typically reduced in dystonia, bringing them back toward the normal range.104 These findings are in line with open data on bilateral epidural premotor cortex stimulation showing a significant improvement after at least 1 month of continuous stimulation.105 The efficacy of rTMS over premotor cortex is not surprising considering the strategic role of the premotor cortex in sensorimotor integration and motor learning. Despite that rTMS appears to be a promising therapeutic tool in dystonia, the overall improvements so far have not been sustained in all studies. It is likely that a limited number of rTMS sessions are not enough to reverse a process, related to an abnormal sensorimotor plasticity that developed over several years. In this context rTMS might be used in the near future to better select dystonic patients as good responders for an eventual epidural cortical implantation. Another possible approach would be to use neuromodulation to reset abnormal oscillations. Transcranial alternating current stimulation (tACS), for example, can entrain regional brain oscillations in a frequency-dependent manner, thereby interacting with specific functions of the stimulated region106–110 This has not yet been undertaken.
Conclusions and future perspectives
There has been considerable progress in understanding the pathophysiology of dystonia. The concept that emerge is a combination of a background of abnormal inhibition and plasticity with environmental influences such as repetitive activity at least in focal hand dystonia. Indeed the relative contribution of abnormal plasticity on one hand, and use dependence on the other, may determine the specific clinical profile of individual patients. For instance, abnormal plasticity may play a stronger role in patients with generalized early onset dystonia and a relatively weaker (but still important) role in focal hand dystonia where additional factors (repetitive hand use or peripheral trauma) are needed to trigger and maintain dystonia. With the loss of homeostatic plasticity, such dysfunction might become permanent, but it might well be possible with prolonged interventions to reverse the abnormalities. DBS certainly is an indication that this can be done, and perhaps non-invasive methods could also be useful in the near future.
References
- 1.Hallett M, Rothwell J. Milestones in clinical neurophysiology. Mov Disord. 2011;26(6):958–967. doi: 10.1002/mds.23572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Quartarone A, Siebner HR, Rothwell JC. Task-specific hand dystonia: can too much plasticity be bad for you? Trends Neurosci. 2006;29(4):192–199. doi: 10.1016/j.tins.2006.02.007. [DOI] [PubMed] [Google Scholar]
- 3.Quartarone A, Pisani A. Abnormal plasticity in dystonia: Disruption of synaptic homeostasis. Neurobiol Dis. 2011;42(2):162–170. doi: 10.1016/j.nbd.2010.12.011. [DOI] [PubMed] [Google Scholar]
- 4.Cohen LG, Hallett M. Hand cramps: clinical features and electromyographic patterns in a focal dystonia. Neurology. 1988;38(7):1005–1012. doi: 10.1212/wnl.38.7.1005. [DOI] [PubMed] [Google Scholar]
- 5.Berardelli A, Rothwell JC, Hallett M, Thompson PD, Manfredi M, Marsden CD. The pathophysiology of primary dystonia. Brain. 1998;121(Pt 7):1195–1212. doi: 10.1093/brain/121.7.1195. [DOI] [PubMed] [Google Scholar]
- 6.Hallett M. Transcranial magnetic stimulation and the human brain. Nature. 2000;406(6792):147–150. doi: 10.1038/35018000. [DOI] [PubMed] [Google Scholar]
- 7.Chen R, Hallett M. Focal dystonia and repetitive motion disorders. Clin Orthop Relat Res. 1998;(351):102–106. [PubMed] [Google Scholar]
- 8.Ridding MC, Sheean G, Rothwell JC, Inzelberg R, Kujirai T. Changes in the balance between motor cortical excitation and inhibition in focal, task specific dystonia. J Neurol Neurosurg Psychiatry. 1995;59(5):493–498. doi: 10.1136/jnnp.59.5.493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Butefisch CM, Boroojerdi B, Chen R, Battaglia F, Hallett M. Task-dependent intracortical inhibition is impaired in focal hand dystonia. Mov Disord. 2005;20(5):545–551. doi: 10.1002/mds.20367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rona S, Berardelli A, Vacca L, Inghilleri M, Manfredi M. Alterations of motor cortical inhibition in patients with dystonia. Mov Disord. 1998;13(1):118–124. doi: 10.1002/mds.870130123. [DOI] [PubMed] [Google Scholar]
- 11.Huang YZ, Edwards MJ, Bhatia KP, Rothwell JC. One-Hz repetitive transcranial magnetic stimulation of the premotor cortex alters reciprocal inhibition in DYT1 dystonia. Mov Disord. 2004;19(1):54–59. doi: 10.1002/mds.10627. [DOI] [PubMed] [Google Scholar]
- 12.Espay AJ, Morgante F, Purzner J, Gunraj CA, Lang AE, Chen R. Cortical and spinal abnormalities in psychogenic dystonia. Ann Neurol. 2006;59(5):825–834. doi: 10.1002/ana.20837. [DOI] [PubMed] [Google Scholar]
- 13.Quartarone A, Rizzo V, Terranova C, et al. Abnormal sensorimotor plasticity in organic but not in psychogenic dystonia. Brain. 2009;132(Pt 10):2871–2877. doi: 10.1093/brain/awp213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Talelli P, Hoffland BS, Schneider SA, et al. A distinctive pattern of cortical excitability in patients with the syndrome of dystonia and cerebellar ataxia. Clin Neurophysiol. 122(9):1816–1819. doi: 10.1016/j.clinph.2011.02.029. [DOI] [PubMed] [Google Scholar]
- 15.Mink JW. The basal ganglia: focused selection and inhibition of competing motor programs. Prog Neurobiol. 1996;50(4):381–425. doi: 10.1016/s0301-0082(96)00042-1. [DOI] [PubMed] [Google Scholar]
- 16.Beck S, Hallett M. Surround inhibition is modulated by task difficulty. Clin Neurophysiol. 2010;121(1):98–103. doi: 10.1016/j.clinph.2009.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hallett M. Neurophysiology of dystonia: The role of inhibition. Neurobiol Dis. 2011;42(2):177–184. doi: 10.1016/j.nbd.2010.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Valls-Sole J, Pascual-Leone A, Wassermann EM, Hallett M. Human motor evoked responses to paired transcranial magnetic stimuli. Electroencephalogr Clin Neurophysiol. 1992;85(6):355–364. doi: 10.1016/0168-5597(92)90048-g. [DOI] [PubMed] [Google Scholar]
- 19.Beck S, Richardson SP, Shamim EA, Dang N, Schubert M, Hallett M. Short intracortical and surround inhibition are selectively reduced during movement initiation in focal hand dystonia. J Neurosci. 2008;28(41):10363–10369. doi: 10.1523/JNEUROSCI.3564-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Beck S, Schubert M, Richardson SP, Hallett M. Surround inhibition depends on the force exerted and is abnormal in focal hand dystonia. J Appl Physiol. 2009;107(5):1513–1518. doi: 10.1152/japplphysiol.91580.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Quartarone A, Bagnato S, Rizzo V, et al. Corticospinal excitability during motor imagery of a simple tonic finger movement in patients with writer’s cramp. Mov Disord. 2005;20(11):1488–1495. doi: 10.1002/mds.20626. [DOI] [PubMed] [Google Scholar]
- 22.Stinear CM, Byblow WD. Impaired modulation of intracortical inhibition in focal hand dystonia. Cereb Cortex. 2004;14(5):555–561. doi: 10.1093/cercor/bhh017. [DOI] [PubMed] [Google Scholar]
- 23.Hallett M. Pathophysiology of dystonia. J Neural Transm Suppl. 2006;(70):485–488. doi: 10.1007/978-3-211-45295-0_72. [DOI] [PubMed] [Google Scholar]
- 24.Luft AR, Manto MU, Ben Taib NO. Modulation of motor cortex excitability by sustained peripheral stimulation: the interaction between the motor cortex and the cerebellum. Cerebellum. 2005;4(2):90–96. doi: 10.1080/14734220410019084. [DOI] [PubMed] [Google Scholar]
- 25.Liepert J, Kucinski T, Tuscher O, Pawlas F, Baumer T, Weiller C. Motor cortex excitability after cerebellar infarction. Stroke. 2004;35(11):2484–2488. doi: 10.1161/01.STR.0000143152.45801.ca. [DOI] [PubMed] [Google Scholar]
- 26.Tamburin S, Fiaschi A, Andreoli A, Marani S, Manganotti P, Zanette G. Stimulus-response properties of motor system in patients with cerebellar ataxia. Clin Neurophysiol. 2004;115(2):348–355. doi: 10.1016/s1388-2457(03)00357-2. [DOI] [PubMed] [Google Scholar]
- 27.Brighina F, Romano M, Giglia G, et al. Effects of cerebellar TMS on motor cortex of patients with focal dystonia: a preliminary report. Exp Brain Res. 2009;192(4):651–656. doi: 10.1007/s00221-008-1572-9. [DOI] [PubMed] [Google Scholar]
- 28.Kassavetis P, Hoffland BS, Saifee TA, et al. Cerebellar brain inhibition is decreased in active and surround muscles at the onset of voluntary movement. Exp Brain Res. 209(3):437–442. doi: 10.1007/s00221-011-2575-5. [DOI] [PubMed] [Google Scholar]
- 29.Levy LM, Hallett M. Impaired brain GABA in focal dystonia. Ann Neurol. 2002;51(1):93–101. [PubMed] [Google Scholar]
- 30.Herath P, Gallea C, van der Veen JW, Horovitz SG, Hallett M. In vivo neurochemistry of primary focal hand dystonia: a magnetic resonance spectroscopic neurometabolite profiling study at 3T. Mov Disord. 25(16):2800–2808. doi: 10.1002/mds.23306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Garibotto V, Romito LM, Elia AE, et al. In vivo evidence for GABA(A) receptor changes in the sensorimotor system in primary dystonia. Mov Disord. 26(5):852–857. doi: 10.1002/mds.23553. [DOI] [PubMed] [Google Scholar]
- 32.Tamura Y, Matsuhashi M, Lin P, et al. Impaired intracortical inhibition in the primary somatosensory cortex in focal hand dystonia. Mov Disord. 2008;23(4):558–565. doi: 10.1002/mds.21870. [DOI] [PubMed] [Google Scholar]
- 33.Martino D, Defazio G, Alessio G, et al. Relationship between eye symptoms and blepharospasm: a multicenter case-control study. Mov Disord. 2005;20(12):1564–1570. doi: 10.1002/mds.20635. [DOI] [PubMed] [Google Scholar]
- 34.Bara-Jimenez W, Shelton P, Hallett M. Spatial discrimination is abnormal in focal hand dystonia. Neurology. 2000;55(12):1869–1873. doi: 10.1212/wnl.55.12.1869. [DOI] [PubMed] [Google Scholar]
- 35.Scontrini A, Conte A, Defazio G, et al. Somatosensory temporal discrimination in patients with primary focal dystonia. J Neurol Neurosurg Psychiatry. 2009;80(12):1315–1319. doi: 10.1136/jnnp.2009.178236. [DOI] [PubMed] [Google Scholar]
- 36.Braun C, Schweizer R, Heinz U, Wiech K, Birbaumer N, Topka H. Task-specific plasticity of somatosensory cortex in patients with writer’s cramp. Neuroimage. 2003;20(2):1329–1338. doi: 10.1016/S1053-8119(03)00375-6. [DOI] [PubMed] [Google Scholar]
- 37.Tinazzi M, Fiorio M, Fiaschi A, Rothwell JC, Bhatia KP. Sensory functions in dystonia: insights from behavioral studies. Mov Disord. 2009;24(10):1427–1436. doi: 10.1002/mds.22490. [DOI] [PubMed] [Google Scholar]
- 38.Nelson AJ, Blake DT, Chen R. Digit-specific aberrations in the primary somatosensory cortex in Writer’s cramp. Ann Neurol. 2009;66(2):146–154. doi: 10.1002/ana.21626. [DOI] [PubMed] [Google Scholar]
- 39.Meunier S, Garnero L, Ducorps A, et al. Human brain mapping in dystonia reveals both endophenotypic traits and adaptive reorganization. Ann Neurol. 2001;50(4):521–527. doi: 10.1002/ana.1234. [DOI] [PubMed] [Google Scholar]
- 40.Delmaire C, Krainik A, Tezenas du Montcel S, et al. Disorganized somatotopy in the putamen of patients with focal hand dystonia. Neurology. 2005;64(8):1391–1396. doi: 10.1212/01.WNL.0000158424.01299.76. [DOI] [PubMed] [Google Scholar]
- 41.Butterworth S, Francis S, Kelly E, McGlone F, Bowtell R, Sawle GV. Abnormal cortical sensory activation in dystonia: an fMRI study. Mov Disord. 2003;18(6):673–682. doi: 10.1002/mds.10416. [DOI] [PubMed] [Google Scholar]
- 42.Bara-Jimenez W, Catalan MJ, Hallett M, Gerloff C. Abnormal somatosensory homunculus in dystonia of the hand. Ann Neurol. 1998;44(5):828–831. doi: 10.1002/ana.410440520. [DOI] [PubMed] [Google Scholar]
- 43.Kaji R, Rothwell JC, Katayama M, et al. Tonic vibration reflex and muscle afferent block in writer’s cramp. Ann Neurol. 1995;38(2):155–162. doi: 10.1002/ana.410380206. [DOI] [PubMed] [Google Scholar]
- 44.Abbruzzese G, Marchese R, Buccolieri A, Gasparetto B, Trompetto C. Abnormalities of sensorimotor integration in focal dystonia: a transcranial magnetic stimulation study. Brain. 2001;124(Pt 3):537–545. doi: 10.1093/brain/124.3.537. [DOI] [PubMed] [Google Scholar]
- 45.Kaji R. Basal ganglia as a sensory gating devise for motor control. J Med Invest. 2001;48(3–4):142–146. [PubMed] [Google Scholar]
- 46.Murase N, Kaji R, Shimazu H, et al. Abnormal premovement gating of somatosensory input in writer’s cramp. Brain. 2000;123(Pt 9):1813–1829. doi: 10.1093/brain/123.9.1813. [DOI] [PubMed] [Google Scholar]
- 47.Graybiel AM. Network-level neuroplasticity in cortico-basal ganglia pathways. Parkinsonism Relat Disord. 2004;10(5):293–296. doi: 10.1016/j.parkreldis.2004.03.007. [DOI] [PubMed] [Google Scholar]
- 48.Ding JB, Guzman JN, Peterson JD, Goldberg JA, Surmeier DJ. Thalamic gating of corticostriatal signaling by cholinergic interneurons. Neuron. 2010;67(2):294–307. doi: 10.1016/j.neuron.2010.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fahn S. High-dosage anticholinergic therapy in dystonia. Adv Neurol. 1983;37:177–188. [PubMed] [Google Scholar]
- 50.Ben Taib NO, Manto M, Pandolfo M, Brotchi J. Hemicerebellectomy blocks the enhancement of cortical motor output associated with repetitive somatosensory stimulation in the rat. J Physiol. 2005;567(Pt 1):293–300. doi: 10.1113/jphysiol.2005.088229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Daskalakis ZJ, Paradiso GO, Christensen BK, Fitzgerald PB, Gunraj C, Chen R. Exploring the connectivity between the cerebellum and motor cortex in humans. J Physiol. 2004;557(Pt 2):689–700. doi: 10.1113/jphysiol.2003.059808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Restuccia D, Valeriani M, Barba C, et al. Functional changes of the primary somatosensory cortex in patients with unilateral cerebellar lesions. Brain. 2001;124(Pt 4):757–768. doi: 10.1093/brain/124.4.757. [DOI] [PubMed] [Google Scholar]
- 53.Pastor MA, Day BL, Macaluso E, Friston KJ, Frackowiak RS. The functional neuroanatomy of temporal discrimination. J Neurosci. 2004;24(10):2585–2591. doi: 10.1523/JNEUROSCI.4210-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Quartarone A, Classen J, Morgante F, Rosenkranz K, Hallett M. Consensus paper: use of transcranial magnetic stimulation to probe motor cortex plasticity in dystonia and levodopa-induced dyskinesia. Brain Stimul. 2009;2(2):108–117. doi: 10.1016/j.brs.2008.09.010. [DOI] [PubMed] [Google Scholar]
- 55.Roze E, Soumare A, Pironneau I, et al. Case-control study of writer’s cramp. Brain. 2009;132(Pt 3):756–764. doi: 10.1093/brain/awn363. [DOI] [PubMed] [Google Scholar]
- 56.Byl NN, Merzenich MM, Jenkins WM. A primate genesis model of focal dystonia and repetitive strain injury: I. Learning-induced dedifferentiation of the representation of the hand in the primary somatosensory cortex in adult monkeys. Neurology. 1996;47(2):508–520. doi: 10.1212/wnl.47.2.508. [DOI] [PubMed] [Google Scholar]
- 57.Quartarone A, Rizzo V, Morgante F. Clinical features of dystonia: a pathophysiological revisitation. Curr Opin Neurol. 2008;21(4):484–490. doi: 10.1097/WCO.0b013e328307bf07. [DOI] [PubMed] [Google Scholar]
- 58.Altenmuller E, Jabusch HC. Focal dystonia in musicians: phenomenology, pathophysiology, triggering factors, and treatment. Med Probl Perform Art. 2010;25(1):3–9. [PubMed] [Google Scholar]
- 59.Quartarone A, Bagnato S, Rizzo V, et al. Abnormal associative plasticity of the human motor cortex in writer’s cramp. Brain. 2003;126(Pt 12):2586–2596. doi: 10.1093/brain/awg273. [DOI] [PubMed] [Google Scholar]
- 60.Torres-Russotto D, Perlmutter JS. Task-specific dystonias: a review. Ann N Y Acad Sci. 2008;1142:179–199. doi: 10.1196/annals.1444.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Tamura Y, Ueki Y, Lin P, et al. Disordered plasticity in the primary somatosensory cortex in focal hand dystonia. Brain. 2009;132(Pt 3):749–755. doi: 10.1093/brain/awn348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Weise D, Schramm A, Stefan K, et al. The two sides of associative plasticity in writer’s cramp. Brain. 2006;129(Pt 10):2709–2721. doi: 10.1093/brain/awl221. [DOI] [PubMed] [Google Scholar]
- 63.Quartarone A, Morgante F, Sant’angelo A, et al. Abnormal plasticity of sensorimotor circuits extends beyond the affected body part in focal dystonia. J Neurol Neurosurg Psychiatry. 2008;79(9):985–990. doi: 10.1136/jnnp.2007.121632. [DOI] [PubMed] [Google Scholar]
- 64.Edwards MJ, Huang YZ, Mir P, Rothwell JC, Bhatia KP. Abnormalities in motor cortical plasticity differentiate manifesting and nonmanifesting DYT1 carriers. Mov Disord. 2006;21(12):2181–2186. doi: 10.1002/mds.21160. [DOI] [PubMed] [Google Scholar]
- 65.Stefan K, Kunesch E, Cohen LG, Benecke R, Classen J. Induction of plasticity in the human motor cortex by paired associative stimulation. Brain. 2000;123(Pt 3):572–584. doi: 10.1093/brain/123.3.572. [DOI] [PubMed] [Google Scholar]
- 66.Belvisi D, Suppa A, Marsili L, et al. Abnormal experimentally- and behaviorally-induced LTP-like plasticity in focal hand dystonia. Exp Neurol. 240:64–74. doi: 10.1016/j.expneurol.2012.11.003. [DOI] [PubMed] [Google Scholar]
- 67.Quartarone A, Sant’Angelo A, Battaglia F, et al. Enhanced long-term potentiation-like plasticity of the trigeminal blink reflex circuit in blepharospasm. J Neurosci. 2006;26(2):716–721. doi: 10.1523/JNEUROSCI.3948-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zeuner KE, Knutzen A, Al-Ali A, et al. Associative stimulation of the supraorbital nerve fails to induce timing-specific plasticity in the human blink reflex. PLoS One. 2010;5(10):e13602. doi: 10.1371/journal.pone.0013602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ghilardi MF, Carbon M, Silvestri G, et al. Impaired sequence learning in carriers of the DYT1 dystonia mutation. Ann Neurol. 2003;54(1):102–109. doi: 10.1002/ana.10610. [DOI] [PubMed] [Google Scholar]
- 70.Turrigiano GG, Leslie KR, Desai NS, Rutherford LC, Nelson SB. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature. 1998;391(6670):892–896. doi: 10.1038/36103. [DOI] [PubMed] [Google Scholar]
- 71.Bienenstock EL, Cooper LN, Munro PW. Theory for the development of neuron selectivity: orientation specificity and binocular interaction in visual cortex. J Neurosci. 1982;2(1):32–48. doi: 10.1523/JNEUROSCI.02-01-00032.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Siebner HR, Lang N, Rizzo V, et al. Preconditioning of low-frequency repetitive transcranial magnetic stimulation with transcranial direct current stimulation: evidence for homeostatic plasticity in the human motor cortex. J Neurosci. 2004;24(13):3379–3385. doi: 10.1523/JNEUROSCI.5316-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Quartarone A, Rizzo V, Bagnato S, et al. Homeostatic-like plasticity of the primary motor hand area is impaired in focal hand dystonia. Brain. 2005;128(Pt 8):1943–1950. doi: 10.1093/brain/awh527. [DOI] [PubMed] [Google Scholar]
- 74.Jung P, Ziemann U. Homeostatic and nonhomeostatic modulation of learning in human motor cortex. J Neurosci. 2009;29(17):5597–5604. doi: 10.1523/JNEUROSCI.0222-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Silberstein P, Kuhn AA, Kupsch A, et al. Patterning of globus pallidus local field potentials differs between Parkinson’s disease and dystonia. Brain. 2003;126(Pt 12):2597–2608. doi: 10.1093/brain/awg267. [DOI] [PubMed] [Google Scholar]
- 76.Liu X, Griffin IC, Parkin SG, et al. Involvement of the medial pallidum in focal myoclonic dystonia: A clinical and neurophysiological case study. Mov Disord. 2002;17(2):346–353. doi: 10.1002/mds.10038. [DOI] [PubMed] [Google Scholar]
- 77.Chen CC, Kuhn AA, Trottenberg T, Kupsch A, Schneider GH, Brown P. Neuronal activity in globus pallidus interna can be synchronized to local field potential activity over 3–12 Hz in patients with dystonia. Exp Neurol. 2006;202(2):480–486. doi: 10.1016/j.expneurol.2006.07.011. [DOI] [PubMed] [Google Scholar]
- 78.Buzsaki G, Draguhn A. Neuronal oscillations in cortical networks. Science. 2004;304(5679):1926–1929. doi: 10.1126/science.1099745. [DOI] [PubMed] [Google Scholar]
- 79.Coubes P, Echenne B, Roubertie A, et al. Treatment of early-onset generalized dystonia by chronic bilateral stimulation of the internal globus pallidus. Apropos of a case. Neurochirurgie. 1999;45(2):139–144. [PubMed] [Google Scholar]
- 80.Coubes P, Roubertie A, Vayssiere N, Hemm S, Echenne B. Treatment of DYT1-generalised dystonia by stimulation of the internal globus pallidus. Lancet. 2000;355(9222):2220–2221. doi: 10.1016/S0140-6736(00)02410-7. [DOI] [PubMed] [Google Scholar]
- 81.Cif L, Vasques X, Gonzalez V, et al. Long-term follow-up of DYT1 dystonia patients treated by deep brain stimulation: an open-label study. Mov Disord. 2010;25(3):289–299. doi: 10.1002/mds.22802. [DOI] [PubMed] [Google Scholar]
- 82.Tisch S, Limousin P, Rothwell JC, et al. Changes in blink reflex excitability after globus pallidus internus stimulation for dystonia. Mov Disord. 2006;21(10):1650–1655. doi: 10.1002/mds.20899. [DOI] [PubMed] [Google Scholar]
- 83.Yianni J, Bain PG, Gregory RP, et al. Post-operative progress of dystonia patients following globus pallidus internus deep brain stimulation. Eur J Neurol. 2003;10(3):239–247. doi: 10.1046/j.1468-1331.2003.00592.x. [DOI] [PubMed] [Google Scholar]
- 84.Vidailhet M, Vercueil L, Houeto JL, et al. Bilateral deep-brain stimulation of the globus pallidus in primary generalized dystonia. N Engl J Med. 2005;352(5):459–467. doi: 10.1056/NEJMoa042187. [DOI] [PubMed] [Google Scholar]
- 85.Ruge D, Tisch S, Hariz MI, et al. Deep brain stimulation effects in dystonia: time course of electrophysiological changes in early treatment. Mov Disord. 2011;26(10):1913–1921. doi: 10.1002/mds.23731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ruge D, Cif L, Limousin P, et al. Shaping reversibility? Long-term deep brain stimulation in dystonia: the relationship between effects on electrophysiology and clinical symptoms. Brain. 2011;134(Pt 7):2106–2115. doi: 10.1093/brain/awr122. [DOI] [PubMed] [Google Scholar]
- 87.Delmaire C, Vidailhet M, Elbaz A, et al. Structural abnormalities in the cerebellum and sensorimotor circuit in writer’s cramp. Neurology. 2007;69(4):376–380. doi: 10.1212/01.wnl.0000266591.49624.1a. [DOI] [PubMed] [Google Scholar]
- 88.Draganski B, Thun-Hohenstein C, Bogdahn U, Winkler J, May A. “Motor circuit” gray matter changes in idiopathic cervical dystonia. Neurology. 2003;61(9):1228–1231. doi: 10.1212/01.wnl.0000094240.93745.83. [DOI] [PubMed] [Google Scholar]
- 89.Teo JT, van de Warrenburg BP, Schneider SA, Rothwell JC, Bhatia KP. Neurophysiological evidence for cerebellar dysfunction in primary focal dystonia. J Neurol Neurosurg Psychiatry. 2009;80(1):80–83. doi: 10.1136/jnnp.2008.144626. [DOI] [PubMed] [Google Scholar]
- 90.Chen R, Wassermann EM, Canos M, Hallett M. Impaired inhibition in writer’s cramp during voluntary muscle activation. Neurology. 1997;49(4):1054–1059. doi: 10.1212/wnl.49.4.1054. [DOI] [PubMed] [Google Scholar]
- 91.Jinnah HA, Hess EJ. A new twist on the anatomy of dystonia: the basal ganglia and the cerebellum? Neurology. 2006;67(10):1740–1741. doi: 10.1212/01.wnl.0000246112.19504.61. [DOI] [PubMed] [Google Scholar]
- 92.Carbon M, Eidelberg D. Abnormal structure-function relationships in hereditary dystonia. Neuroscience. 2009;164(1):220–229. doi: 10.1016/j.neuroscience.2008.12.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Detante O, Vercueil L, Thobois S, et al. Globus pallidus internus stimulation in primary generalized dystonia: a H215O PET study. Brain. 2004;127(Pt 8):1899–1908. doi: 10.1093/brain/awh213. [DOI] [PubMed] [Google Scholar]
- 94.Thobois S, Ballanger B, Xie-Brustolin J, et al. Globus pallidus stimulation reduces frontal hyperactivity in tardive dystonia. J Cereb Blood Flow Metab. 2008;28(6):1127–1138. doi: 10.1038/sj.jcbfm.9600610. [DOI] [PubMed] [Google Scholar]
- 95.Neychev VK, Gross RE, Lehericy S, Hess EJ, Jinnah HA. The functional neuroanatomy of dystonia. Neurobiol Dis. 2011;42(2):185–201. doi: 10.1016/j.nbd.2011.01.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Delnooz CC, Helmich RC, Toni I, van de Warrenburg BP. Reduced parietal connectivity with a premotor writing area in writer’s cramp. Mov Disord. 2012;27(11):1425–1431. doi: 10.1002/mds.25029. [DOI] [PubMed] [Google Scholar]
- 97.Jin SH, Lin P, Auh S, Hallett M. Abnormal functional connectivity in focal hand dystonia: mutual information analysis in EEG. Mov Disord. 2011;26(7):1274–1281. doi: 10.1002/mds.23675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Candia V, Schafer T, Taub E, et al. Sensory motor retuning: a behavioral treatment for focal hand dystonia of pianists and guitarists. Arch Phys Med Rehabil. 2002;83(10):1342–1348. doi: 10.1053/apmr.2002.35094. [DOI] [PubMed] [Google Scholar]
- 99.Zeuner KE, Bara-Jimenez W, Noguchi PS, Goldstein SR, Dambrosia JM, Hallett M. Sensory training for patients with focal hand dystonia. Ann Neurol. 2002;51(5):593–598. doi: 10.1002/ana.10174. [DOI] [PubMed] [Google Scholar]
- 100.Zeuner KE, Shill HA, Sohn YH, et al. Motor training as treatment in focal hand dystonia. Mov Disord. 2005;20(3):335–341. doi: 10.1002/mds.20314. [DOI] [PubMed] [Google Scholar]
- 101.Zeuner KE, Peller M, Knutzen A, Hallett M, Deuschl G, Siebner HR. Motor re-training does not need to be task specific to improve writer’s cramp. Mov Disord. 2008;23(16):2319–2327. doi: 10.1002/mds.22222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Siebner HR, Tormos JM, Ceballos-Baumann AO, et al. Low-frequency repetitive transcranial magnetic stimulation of the motor cortex in writer’s cramp. Neurology. 1999;52(3):529–537. doi: 10.1212/wnl.52.3.529. [DOI] [PubMed] [Google Scholar]
- 103.Benninger DH, Lomarev M, Lopez G, Pal N, Luckenbaugh DA, Hallett M. Transcranial direct current stimulation for the treatment of focal hand dystonia. Mov Disord. 2011;26(9):1698–1702. doi: 10.1002/mds.23691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Huang YZ. The modulation of cortical motor circuits and spinal reflexes using theta burst stimulation in healthy and dystonic subjects. Restor Neurol Neurosci. 2010;28(4):449–457. doi: 10.3233/RNN-2010-0554. [DOI] [PubMed] [Google Scholar]
- 105.Lalli S, Piacentini S, Franzini A, et al. Epidural premotor cortical stimulation in primary focal dystonia: Clinical and (18) F-fluoro deoxyglucose positron emission tomography open study. Mov Disord. 2012;27(4):533–538. doi: 10.1002/mds.24949. [DOI] [PubMed] [Google Scholar]
- 106.Kanai R, Chaieb L, Antal A, Walsh V, Paulus W. Frequency-dependent electrical stimulation of the visual cortex. Curr Biol. 2008;18(23):1839–1843. doi: 10.1016/j.cub.2008.10.027. [DOI] [PubMed] [Google Scholar]
- 107.Kanai R, Paulus W, Walsh V. Transcranial alternating current stimulation (tACS) modulates cortical excitability as assessed by TMS-induced phosphene thresholds. Clin Neurophysiol. 2010;121(9):1551–1554. doi: 10.1016/j.clinph.2010.03.022. [DOI] [PubMed] [Google Scholar]
- 108.Pogosyan A, Gaynor LD, Eusebio A, Brown P. Boosting cortical activity at Beta-band frequencies slows movement in humans. Curr Biol. 2009;19(19):1637–1641. doi: 10.1016/j.cub.2009.07.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Thut G, Miniussi C. New insights into rhythmic brain activity from TMS-EEG studies. Trends Cogn Sci. 2009;13(4):182–189. doi: 10.1016/j.tics.2009.01.004. [DOI] [PubMed] [Google Scholar]
- 110.Paulus W. On the difficulties of separating retinal from cortical origins of phosphenes when using transcranial alternating current stimulation (tACS) Clin Neurophysiol. 2010;121(7):987–991. doi: 10.1016/j.clinph.2010.01.029. [DOI] [PubMed] [Google Scholar]