

Abstract
Adenylyl cyclase (AC) is a key enzyme that synthesizes cyclic AMP (cAMP) at the onset of the signaling pathway to activate sperm motility. Here, we showed that both transmembrane AC (tmAC) and soluble AC (sAC) are distinctly involved in the regulation of sperm motility in the ascidian Ciona intestinalis. A tmAC inhibitor blocked both cAMP synthesis and the activation of sperm motility induced by the egg factor sperm activating and attracting factor (SAAF), as well as those induced by theophylline, an inhibitor of phoshodiesterase. It also significantly inhibited cAMP-dependent phosphorylation of a set of proteins at motility activation. On the other hand, a sAC inhibitor does not affect on SAAF-induced transient increase of cAMP, motility activation or protein phosphorylation, but it reduced swimming velocity to half in theophylline-induced sperm. A sAC inhibitor KH-7 induced circular swimming trajectory with smaller diameter and significantly suppressed chemotaxis of sperm to SAAF. These results suggest that tmAC is involved in the basic mechanism for motility activation through cAMP-dependent protein phosphorylation, whereas sAC plays distinct roles in increase of flagellar beat frequency and in the Ca2+-dependent chemotactic movement of sperm.
Keywords: calcium, sperm chemotaxis, fertilization, protein kinase, protein phosphorylation
1. Introduction
In most animal species, sperm are immotile in testis or sperm duct. Flagellar motility is activated and regulated by external cues such as specific ions, osmotic change or factors from the egg or female genital tract at fertilization [1]. In the ascidian Ciona intestinalis and C. savignyi, a sulfated steroid called sperm activating and attracting factor (SAAF) induces both sperm motility activation and chemotaxis [2]. Cyclic AMP (cAMP) is one of the most important intracellular factors in the signaling pathway for SAAF-induced sperm activation. A 21 kDa light chain of outer arm dynein (LC2) and a 26 kDa axonemal protein are phosphorylated, and dynein intermediate chains IC2 and IC116 are dephosphorylated in a cAMP-dependent manner, resulting in activation of flagellar motility [3,4].
Adenylyl cyclases (ACs) are the key enzyme that synthesizes cAMP at the onset of the signaling pathway to activate axonemal dyneins through protein phosphorylation. In sea urchin, frog and mammalian sperm, both soluble adenylyl cyclase (sAC) and transmembrane adenylyl cyclases (tmACs) are present and play important roles in sperm function [5,6,7,8,9,10]. However, the roles of two types of ACs are controversial. tmACs are suggested to be involved in the regulation of sperm motility, in particular during chemotactic behavior [11,12]. However, tmACs are shown localized at the acrosome [9,13]. sAC-deficient mice are infertile, lacking sperm motility [8]. In fact, sAC is localized in sperm flagella in mouse [8] and sea urchin [14]. In this study we first characterized sperm ACs in the ascidian C. intestinalis. Using inhibitors specific to each type of AC, we demonstrate that tmAC or sAC critically plays a distinct role in the basic activation of flagellar motility or increase in flagellar beat frequency and Ca2+-dependent regulation of flagellar waveform during chemotactic movement, respectively.
2. Results and Discussion
2.1. Expression of Adenylyl Cyclase (AC) Genes in Ciona Testis
Ten isoforms of AC have been cloned and characterized in mammals; nine are tmACs and the other is sAC [5,6]. Their distributions and regulations differ among isoforms. To clarify the functions of AC in Ciona sperm, we first examined the expression of AC genes in Ciona tesits. A search for AC genes in the genome of Ciona intestinalis revealed the presence of three tmAC genes and one sAC gene. Phylogenetic analysis suggested that three tmAC genes are grouped with mammalian AC5/6, AC8 and AC9 (Figure 1). Next, we examined the expression of AC genes by reverse-transcriptase PCR (RT-PCR) using PCR primers for these four AC genes. Recently, AC5/6 was reported to be expressed in the intestine of Ciona juvenile [15]. From the analysis in the present study, it turned out that all AC isoforms were expressed in testis but the expression of AC8 is only testis-specific. sAC was expressed in testis at a high level but was also significantly expressed in ovary (Figure 2).
Figure 1.

Phylogenetic analysis of adenylyl cyclases. Ciona tetrameric potassium-selective cyclic nucleotide-gated channel (Ci-KCNG) was used as the outgroup. The value on each branch represents the number of times that a node was supported in 1000 bootstrap pseudo replications. BLASTP search of adenylyl cyclase (AC) against Ciona genome revealed four AC gene models. Phylogenetic analysis suggests that ACs encoded by these four genes are grouped with mammalian AC5/6, AC8, AC9 and soluble AC (sAC) respectively. Accession numbers of proteins are: Homo sapiens: Hs-AC1 (NP_066939), Hs-AC2 (NP_065433), Hs-AC3 (NP_004027), Hs-AC4 (NP_640340), Hs-AC5 (NP_899200), Hs-AC6 (NP_056085), Hs-AC7 (NP_001105), Hs-AC8 (NP_001106), Hs-AC9 (NP_001107), Hs-sAC (NP_060887); Mus musculus: Mm-AC1 (NP_033752), Mm-AC2 (NP_705762), Mm-AC3 (NP_001153008), Mm-AC4 (NP_536683), Mm-AC5 (NP_001012783), Mm-AC6 (EDL04180), Mm-AC7 (NP_001032813), Mm-AC8 (NP_033753), Mm-AC9 (NP_033754), Mm-sAC (NP_766617). Sequences of Ciona proteins are obtained from the Ghost database (http://ghost.zool.kyoto-u.ac.jp). Proteins encoded by Ciona gene models are indicated in red. The bars in green or in red represents tmAC or sAC, respectively.
Figure 2.
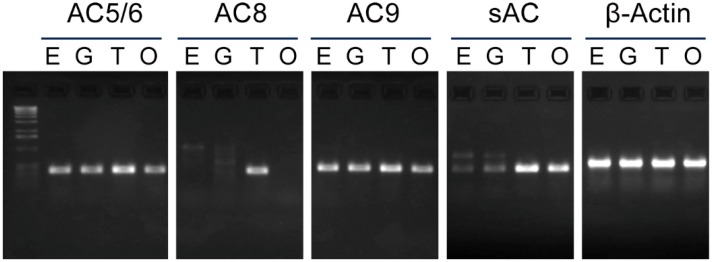
Tissue expression patterns of the adenylyl cyclase (AC) genes in Ciona tissues. RT-PCR analysis of mRNA from several adult Ciona tissues shows that genes encoding all AC isoforms are expressed in testis but the expression of AC8 is testis-specific. Expression of sAC gene was found in testis and ovary. β-actin was used as an internal control. E, endostyle; G, gill; T, testis; and O, ovary.
2.2. Effects of AC Inhibitors on the Motility Activation of Ciona Sperm
2.2.1. Sperm Activating and Attracting Factor (SAAF)-Induced Motility Activation
Ciona sperm show less motility in seawater but become activated by an egg-derived factor, sperm activating and attracting factor (SAAF) [16]. SAAF induces Ca2+ influx, membrane hyperpolarization and activation of adenylyl cyclase to produce cAMP [16]. Transient increase of intracellular cAMP and subsequent cAMP-dependent phosphorylation of dynein subunits are observed in response to the addition of SAAF [3,17]. Thus, cAMP is one of important factors for SAAF-induced sperm motility activation. To specify an AC isoform that is involved in SAAF-induced motility activation, we examined the effect of AC inhibitors specific to tmAC or sAC on motility activation and cAMP synthesis (Figure 3). The tmAC inhibitor, MDL12330A, completely blocked SAAF-induced motility activation and transient increase of cAMP. MDL12339A did not affect any changes on the motility Triton X-100-demembranated sperm (data not shown). In contrast, the sAC inhibitor, KH-7, did not inhibit SAAF-induced motility activation or transient increase of cAMP synthesis. However it reduced swimming velocity to half of the control and brought basal cAMP level to almost zero in 5 min. Swimming velocity is closely related to the flagellar beat frequency [18]. These results indicate that tmAC has a major role in the signaling pathway in SAAF-induced motility activation and that sAC is involved in the increase in beat frequency and in the maintenance of basal level of cAMP. This agrees with the observation in mouse sperm that a sAC activator HCO3− induces the increase of flagellar beat frequency through cAMP-dependent protein kinase (protein kinase A; PKA) [19].
Figure 3.

Effects of AC inhibitors on sperm activating and attracting factor (SAAF)-induced activation of Ciona sperm motility and cAMP synthesis. The percentage of motile sperm (A); curvilinear velocity (B) and intracellular cAMP level (C) were measured before, 1 min and 5 min after the addition of 100 nM SAAF. Sperm were treated with 0.5% dimethyl sulfoxide (DMSO, control), 100 μM MDL12330A or 10 μM KH-7 respectively for 3 min before SAAF addition. Values are means ± standard error (SE) of the results from three experiments.
2.2.2. Valinomycin- and Theophylline-Induced Motility Activation
Valinomycin is a potassium ion-selective ionophore and induces motility activation of Ciona sperm by causing membrane hyperpolarization [17]. In contrast, an increase in intracellular cAMP levels was induced by a phosphodiesterase inhibitor theophylline, resulting in the motility activation [16]. To elucidate if sAC participates in SAAF-induced signaling pathway in the motility activation, we examined the effect of AC inhibitors on valinomycin- and theophylline-induced motility activation in the presence or absence of Ca2+ (Figure 4). Both valinomycin and theophylline activated sperm motility in ASW but valinomycin-treated sperm swam with lower velocity (Figure 4A,B). The tmAC inhibitor MDL12330A completely suppressed both motility and swimming velocity in artificial seawater (ASW) (Figure 4A,B). These results suggest the involvement of tmAC in the motility of valinomycin-treated sperm. Valinomycin-treated sperm were immotile in Ca2+-free seawater (CaFSW), as also previously reported [20], suggesting that Ca2+-influx is necessary for tmAC and motility activation (Figure 4C).
Figure 4.

Effects of AC inhibitors on valinomycin (Val)- or theophylline (Theo)-induced activation of Ciona sperm motility. The percentage of motile sperm and curvilinear velocity were measured before, 1 and 5 min after the addition of 10 nM valinomycin or 1 mM theophylline in artificial sea water (ASW; A,B) or Ca2+-free sea water (CaFSW; C,D). Sperm were treated with 0.5% DMSO (control), 100 μM MDL12330A or 10 μM KH-7 respectively for 3 min before the addition of valinomycin or theophylline. Values are means ± SE of the results from three experiments.
Theophylline-treated sperm showed activated in both ASW and CaFSW. The sAC inhibitor KH-7 showed ~40% suppression of the motility at 1 min but the suppression became not significant at 5 min (Figure 4B). The velocity of theophylline-treated sperm decreased to half of the control by KH-7 in ASW (Figure 4B), supporting the idea that sAC is involved in the increase of flagellar beat frequency. In contrast, theophylline-treated sperm showed motility activation even in CaFSW. The motility was suppressed by KH-7 (Figure 4D), suggesting that sAC is involved in the sperm motility in Ca2+-free condition. The sAC in human sperm shows synthesis of cAMP in the absence of Ca2+ but is activated by Ca2+ in a concentration-dependent manner [21]. Therefore, it is likely that the activation of sperm motility in CaFSW by theophylline is caused by basal activity of sAC in the absence of Ca2+. Ca2+-activated sAC could be involved in other process, such as regulation of intracellular pH (see 2.2.3 in the Results and Discussion Section) and sperm chemotaxis (see 2.3 in the Results and Discussion Section).
2.2.3. Sperm Motility Is Activated by HCO3− but not by Forskolin
To further test the roles of two types of ACs in the regulation of sperm motility, we examined the effects of two activators for ACs, forskolin and HCO3−. Forskolin is a diterpene that effectively activates tmAC1 to tmAC8 [22,23] and is shown to increase cAMP and activate motility in mammalian sperm [13,24]. In Ciona sperm, forskolin showed no effect on the activation of sperm motility even at 100 μM (Figure 5A), suggesting that either Ciona tmAC5/6 or tmAC8 does not participate in SAAF-dependent activation of sperm motility.
Figure 5.

Effects of AC activators on activation of Ciona sperm motility. The percentage of motile sperm was measured 3 min (A) or 1 min (B,C) after the addition of sperm to the solutions containing forskolin (A), NaHCO3 (B,C) with or without sAC inhibitors. The concentration of KH-7 or nigericin was 10 or 5 μM, respectively. Values are means ± SE of the results from three experiments.
sAC is directly activated by HCO3− and Ca2+ [25,26], both of which are two important factors for capacitation of mammalian sperm [27]. Although it is known that CO2 inhibits motility of sperm from marine animals showing external fertilization, such as sea urchin [28,29] and flatfish [30], activation of sperm motility by HCO3− has not been reported. Here, we observed that extracellular HCO3− activated the motility in Ciona sperm (Figure 5B). Because insoluble CaCO3 formed by the addition of high concentration of HCO3− into sea water disturbed the observation of sperm motility, we used the artificial sea water containing 1 mM CaCl2 (low calcium sea water; LCSW). The activation was observed in LCSW but not in CaFSW (Figure 5B), indicating that extracellular Ca2+ is required for HCO3−mediated motility activation. To see if the action of HCO3− is due to the change in intracellular pH, we used an antiporter of H+ and K+, nigericin, to bring intracellular pH to the same pH of extracellular solution. Nigericin itself activated sperm motility in ASW at pH 7.5, but the level of activation was lower than of sperm activated by HCO3− (Figure 5C). The motility of HCO3−-activated sperm was reduced by nigericin, suggesting that intracellular pH of HCO3−-activated sperm was higher than 7.5. KH-7 inhibited both nigericin-activated sperm and HCO3−-activated sperm. These results suggest that the action of HCO3− is mediated by sAC in a mechanism closely coupled with the increase in intracellular pH. It is likely that the activity of Ciona sAC is highly dependent on pH like the case in sea urchin sperm [14] and that cAMP produced by sAC activates a Na+/H+ exchanger (sNHE) through cyclic nucleotide binding site to raise intracellular pH [31,32] (see [33] for review).
2.3. Effects of sAC Inhibitors on Sperm Chemotaxis and Swimming Trajectory
Sperm chemotaxis in Ciona is controlled by transient increase in intracellular Ca2+ concentration, followed by calaxin-mediated propagation of asymmetric flagella waveforms [34,35]. Next, we examined the effect of KH-7 on the chemotactic behavior (Figure 6). The movement with quick “turn” toward the attractant was significantly suppressed by KH-7 (Figure 6A,B). The calculated chemotactic index (LECI, [2]) demonstrates that sperm chemotaxis is significantly inhibited by KH-7 (Figure 6C). We found that diameter of the circle of sperm trajectory became smaller in the presence of KH-7. An analysis of flagellar waveform asymmetry indicates that flagellar waveform become more asymmetric in the presence of KH-7 (Figure 7). This suggests that intracellular Ca2+ concentration increases by KH-7, most likely by inhibiting Ca2+-efflux system, such as Na+/Ca2+ exchanger [36]. It was also reported in mammalian sperm that HCO3−-mediated sAC activity is involved in conversion into symmetrical flagellar waveform [27]. Alternatively, sAC could participate in the mechanism for the resumption from calaxin-mediated asymmetric waveform to symmetric waveform [35]. In either case, inhibition of sAC by KH-7 results in loss of efficient turn movement, which would cause suppression of chemotactic movement of sperm. It is still possible that the loss of efficient turn movement is secondarily resulted from KH-7-induced decrease in swimming velocity.
Figure 6.

Effects of sAC inhibitor KH-7 on chemotaxis of Ciona sperm. (A) Swimming trajectories during 2 s of sperm treated with 0.5% DMSO (control, left) and sperm treated with 10 μM KH-7 (right). The photo was integrated from 200 images taken at 100 fps. The red asterisks represent the tip of glass capillary filled with SAAF; (B) Sperm path trajectories of three representative sperm are shown; and (C) Comparison of linear equation chemotaxis indices (LECI) showing significant inhibition of chemotaxis by KH-7. Values are means ± SE. n = 29 (control) and 34 (KH-7). *** Significant at p < 0.001 (Student’s t-test) as compared with the control.
Figure 7.

Effects of sAC inhibitor KH-7 on the swimming trajectory and flagellar wave asymmetry of Ciona sperm. (A) Swimming trajectories during 1 sec of sperm treated with 0.5% DMSO (control, left) and sperm treated with 10 μM KH-7 (right). Sperm were activated by 1 mM theophylline. The photo was integrated from 200 images taken at 200 fps; (B) Flagellar bending pattern of control or KH-7 treated sperm. Data from 20 waveforms are overwritten. Red dots indicate head position; and (C) Diameter of the circle of sperm trajectory in the presence of DMSO (control) or KH-7. Values are means ± SD. n = 120 (control) and 128 (KH-7). *** Significant at p < 0.001 (Student’s t-test) as compared with the control.
2.4. Effects of AC Inhibitors on cAMP-Dependent Protein Phosphorylation in Ciona Sperm
cAMP-dependent protein phosphorylation is prerequisite for activation of sperm flagellar motility [3,37,38,39]. Next we examined the effects of AC inhibitors on the PKA-dependent protein phosphorylation in Ciona sperm (Figure 8). SAAF induced PKA-mediated phosphorylation of six proteins, including a high molecular mass (HMM) protein (possibly dynein heavy chain), 105 kDa protein, 80 kDa protein, 65 kDa protein, 48 kDa protein (possibly PKA regulatory subunit), and 26 kDa protein (Figure 8, asterisks). Phosphorylation of these proteins, except for the 48 kDa protein, was greatly diminished in sperm treated with MDL12330A in both the presence and absence of Ca2+. Phosphorylation of both the 105 and 80 kDa proteins were suppressed by KH-7 in ASW, suggesting that these are closely related to the function of sAC. In contrast, HMM, 65 and 26 kDa proteins were significantly phosphorylated in KH-7-treated sperm, suggesting that these proteins are involved more in basic activation of sperm motility than in the increase of beat frequency or in chemotactic behavior (see motility experiments in Figure 4, Figure 5, Figure 6 and Figure 7). In CaFSW, only theophylline-treated sperm without AC inhibitors exhibit motility. The phosphorylation pattern well coincided with the motility data (Figure 4); phosphorylation of all the six major proteins of theophylline-treated sperm in CaFSW was more prominent than the others (Figure 8).
Figure 8.

Effects of AC inhibitors on SAAF-, valinomycin (Val)- or theophylline (Theo)-induced protein phosphorylation by protein kinase (PKA). CBB-stained pattern and corresponding western blot with anti-PKA-substrate antibody were shown for sperm treated by artificial seawater (ASW), 100 nM SAAF, 10 nM valinomycin or 1 mM theophylline with 0.5% DMSO (control), 100 μM MDL12330A or 10 μM KH-7. Data obtained in ASW (top) and in CaFSW (bottom) are shown. The percentage of polyacrylamide in the separating gel is 10%. Proteins with significant increase in phosphorylation by SAAF-activation are indicated by asterisks in the lane of sperm sample activated by SAAF in ASW. Those showing decreased phospholylation by KH-7 in ASW are marked in green and others are in red. Data show a typical result from three independent experiments.
In the present study, we showed distinct roles of tmAC and sAC in the regulation of sperm motility in Ciona. tmAC is thought to be involved in the basal pathway for activation of motility; it transiently synthesizes high level of cAMP in response to SAAF-induced Ca2+ influx and membrane hyperpolarization, resulting in the activation of PKA that phosphorylates axonemal proteins for motility. In contrast, sAC is thought to mainly participate in the regulation of the asymmetry of flagellar waveforms and increase of beat frequency. The swimming velocity of SAAF-activated, KH-7-treated sperm showed half of the control sperm. This is also the case in sperm activated by valinomycin as well as theophylline-activated, KH-7-treated sperm. Thus, it is likely that flagellar motility is regulated by at least two pathways: SAAF-dependent tmAC pathway for basic activation for motility and SAAF-independent sAC pathway for full motility with high beat frequency and waveform conversion leading to sperm chemotaxis. Proteins that are differently phosphorylated with or without AC inhibitors may be the key to prove this model. Several studies demonstrate distinct localization of AC isoforms in mammalian [40,41,42] and sea urchin sperm [14]. Studies on immunolocalization of ACs in Ciona sperm would surely provide further evidence for the roles of ACs in the regulation of sperm motility.
3. Experimental Section
3.1. Ciona Sperm
The ascidian Ciona intestinalis was supplied by the Education and Research Center of Marine Bio-Resources, Tohoku University, Onagawa, Japan, and the National Bio-Resource Project of the Ministry of Education, Culture, Sports, Science and Technology (MEXT). Animals were kept in aquaria under constant light for accumulation of gametes without spontaneous spawning. Semen samples were collected by dissecting the sperm duct and kept on ice until use.
3.2. Chemicals and Solutions
Artificial seawater (ASW) contained 462.01 mM NaCl, 9.39 mM KCl, 10.81 mM CaCl2, 48.27 mM MgCl2, and 10 mM Hepes-NaOH (pH 7.6). Ca2+-free sea water (CaFSW) was as ASW without CaCl2 but with 478.2 mM NaCl and 10 mM EGTA. Low Ca2+ sea water (LCSW) was as ASW with 1 mM CaCl2 and 476.7 mM NaCl. MDL-12,330A from Calbiochem (San Diego, CA, USA), KH-7 from Chemical Diversity Research Institute (Khimki, Russia) and valinomycine and forskolin from Sigma-Aldrich (St. Louis, MO, USA) were dissolved in dimethyl sulfoxide (DMSO). Theophylline from Sigma-Aldrich was dissolved in ASW. Other reagents were of analytical grades. SAAF was partially purified from the egg-seawater of C. intestinalis according to Yoshida et al. [43].
3.3. Sequence Analysis
BLAST searches of mammalian AC sequences in the C. intestinalis genome database (http://ghost.zool.kyoto-u.ac.jp) identified four AC gene models (Ci-sAC: KH.C10.393, Ci-AC5/6: KH.C14.32, Ci-AC8: KyotoGrail2006.694.2.1, Ci-AC9: KH.C14.534). Phylogenetic tree was constructed by ClustalW2 (http://www.ebi.ac.uk/Tools/msa/ clustalw2/) with the aid of MEGA5 [44].
3.4. Reverse-Transcriptase (RT) PCR
Total RNA was extracted from the endostyle, gill, testis and ovary of C. intestinalis by using Isogen (Nippon gene, Toyama, Japan), separated by chloroform:isoamyl alcohol 24:1 (Sigma-Aldrich) and precipitated by isopropanol. cDNA was synthesized from the total RNA of each tissue by using SuperScript III reverse transcriptase (Invitrogen, Carlsbad, CA, USA). PCR was performed with Elongase® Enzyme Mix (Invitrogen, Carlsbad, CA, USA) and specific primers as follows: Ci-sAC, 5'-GATATTGGGGATGTTCCACG-3' (forward) and 5'-GTGCGGGCCATCTTAATAGA-3' (reverse); Ci-AC5/6, 5'-TGAATGTGTCGCCGTTATGT-3' (forward) and 5'-CAACGTTTACCGTATTGCCC-3' (reverse); Ci-AC8, 5'-ATGCTAGCTCACCTTGCGTT-3' (forward) and 5'-ATCAGTGTGCCATCTTTCCC-3' (reverse); Ci-AC9, 5'-AACCCCACCGACAATTACAA-3' (forward) and 5'-ACAACGGTTTCGAACAGACC-3' (reverse). Amplifications were carried out for 35 cycles (94 °C for 1 min, 42 °C for 2 min and 68 °C for 2 min).
3.5. Assessment of Sperm Motility
Semen was suspended in 2000 volumes of ASW containing DMSO or inhibitors and incubated for 3 min. The sperm suspension was immediately placed on a glass slide coated with 1% BSA to avoid adhesion of sperm to the glass. The sperm movement was recorded at RT under a phase contrast microscope (BX51, Olympus, Tokyo, Japan) with a 10× objective (UPlan FLN, Olympus, Tokyo, Japan) and analyzed using Sperm Motility Analysis System (SMAS, DITECT Corporation, Tokyo, Japan). The percentage of motile sperm and curvilinear velocity were analyzed before, 1 and 5 min after the addition of SAAF, theophylline and valinomycin (final concentration; 100 nM, 1 mM and 10 nM respectively). Analysis of chemotactic behavior and flagellar waveform of sperm was performed by the method previously described [35]. Images were recorded with a high-speed CCD camera (HAS-D3, DITECT Corporation, Tokyo, Japan) and analyzed using Bohboh software (Bohboh Soft, Tokyo, Japan).
3.6. Measurement of cAMP
Levels of intracellular cAMP were assayed by using Amersham cAMP Biotrak Enzyme immunoassay (EIA) System (GE Healthcare, Buckinghamshire, UK). Semen was suspended in 2000 volumes of ASW containing DMSO (WAKO, Osaka, Japan) or inhibitors and incubated for 3 min, and SAAF (final concentration 100 nM) was added to the suspension. The sperm suspension before, 10 s, 1 min and 5 min after the addition of SAAF were transferred to a lysis buffer to stop the reaction. The lysate was centrifuged at 12,000× g for 10 min at RT and the supernatant was used for the measurement of cAMP.
3.7. Immunoblotting by Anti-Phospho-PKA Substrate Antibody
Semen was suspended in 100 volumes of ASW or CaFSW containing DMSO or inhibitors and incubated for 3 min, and SAAF, theophylline or valinomycin (final concentration; 100 nM, 1 mM or 10 nM respectively) were added to the suspension. After 1 min cells were solubilized in a sample buffer; 62.4 mM Tris-HCl, 2% SDS, 4% glycerol, 0.004% bromophenol-blue, pH 6.8 and boiled at 95 °C for 2 min. Proteins separated by SDS-PAGE were transferred to a polyvinylidene difluoride (PVDF) membrane and subjected to Western blotting with anti-phospho-cAMP-dependent protein kinase (PKA) substrate monoclonal antibody (#9624, Cell Signaling Technology, Beverly, MA, USA) as the primary antibody and horseradish peroxidase (HRP)-conjugated anti-rabbit IgG (Invitrogen, Carlsbad, CA, USA) as the secondary antibody. The immunoreactive bands were detected using ECL-prime (GE Healthcare, Buckinghamshire, UK).
4. Conclusions
Three tmAC (AC5/6, AC8 and AC9) and one sAC are encoded in the genome of the ascidian Ciona intestinalis. All of these AC genes are expressed in the testis. From the analyses of sperm motility using inhibitors for tmAC and sAC, we show that tmAC and sAC play distinct roles of in the regulation of sperm motility. tmAC transiently synthesizes high level of cAMP in response to SAAF-induced Ca2+ influx and membrane hyperpolarization, resulting in the activation of PKA that phosphorylates axonemal proteins for basal activation of sperm motility. In contrast, sAC mainly participates in the regulation of the asymmetry of flagellar waveforms and increase in beat frequency in a HCO3−/pH-dependent manner. Phosphorylation of 105 and 80 kDa proteins appears closely related to this process. Thus, flagellar motility is likely to be regulated by two pathways: (1) SAAF-dependent tmAC pathway for basic activation of motility and (2) SAAF-independent sAC pathway for full motility with high beat frequency and waveform conversion.
Acknowledgments
We are grateful to all the staff members of Education and Research Center of Marine Bio-Resources, Tohoku University and National Bio Resource Project (NBRP), and International Coastal Research Center, University of Tokyo for supplying Ciona intestinalis. This work was supported in part by a grant from MEXT (Ministry of Education, Culture, Sports, Science and Technology), Japan to K.I. and K.S. and by JST-BIRD (Japan Science and Technology Agency-Institute for Bioinformatics Research and Development), Japan to K.I.
Author Contributions
K.S. and K.I. designed the study, performed experiments and data acquisition, performed statistical analysis and wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Inaba K. Molecular architecture of the sperm flagella: Molecules for motility and signaling. Zool. Sci. 2003;20:1043–1056. doi: 10.2108/zsj.20.1043. [DOI] [PubMed] [Google Scholar]
- 2.Yoshida M., Murata M., Inaba K., Morisawa M. A chemoattractant for ascidian spermatozoa is a sulfated steroid. Proc. Natl. Acad. Sci. USA. 2002;99:14831–14836. doi: 10.1073/pnas.242470599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nomura M., Inaba K., Morisawa M. Cyclic AMP- and calmodulin-dependent phosphorylation of 21 and 26 kDa proteins in axoneme is a prerequisite for SAAF-induced motile activation in ascidian spermatozoa. Dev. Growth Differ. 2000;42:129–138. doi: 10.1046/j.1440-169x.2000.00489.x. [DOI] [PubMed] [Google Scholar]
- 4.Hozumi A., Padma P., Toda T., Ide H., Inaba K. Molecular characterization of axonemal proteins and signaling molecules responsible for chemoattractant-induced sperm activation in Ciona intestinalis. Cell Motil. Cytoskelet. 2008;65:249–267. doi: 10.1002/cm.20258. [DOI] [PubMed] [Google Scholar]
- 5.Hanoune J., Pouille Y., Tzavara E., Shen T., Lipskaya L., Miyamoto N., Suzuki Y., Defer N. Adenylyl cyclases: Structure, regulation and function in an enzyme superfamily. Mol. Cell Endocrinol. 1997;128:179–194. doi: 10.1016/S0303-7207(97)04013-6. [DOI] [PubMed] [Google Scholar]
- 6.Buck J., Sinclair M.L., Schapal L., Cann M.J., Levin L.R. Cytosolic adenylyl cyclase defines a unique signaling molecule in mammals. Proc. Natl. Acad. Sci. USA. 1999;96:79–84. doi: 10.1073/pnas.96.1.79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Baxendale R.W., Fraser L.R. Evidence for multiple distinctly localized adenylyl cyclase isoforms in mammalian spermatozoa. Mol. Reprod. Dev. 2003;66:181–189. doi: 10.1002/mrd.10344. [DOI] [PubMed] [Google Scholar]
- 8.Hess K.C., Jones B.H., Marquez B., Chen Y., Ord T.S., Kamenetsky M., Miyamoto C., Zippin J.H., Kopf G.S., Suarez S.S., et al. The “soluble” adenylyl cyclase in sperm mediates multiple signaling events required for fertilization. Dev. Cell. 2005;9:249–259. doi: 10.1016/j.devcel.2005.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Beltran C., Zapata O., Darszon A. Membrane potential regulates sea urchin sperm adenylylcyclase. Biochemistry. 1996;35:7591–7598. doi: 10.1021/bi952806v. [DOI] [PubMed] [Google Scholar]
- 10.O’Brien E.D., Krapf D., Cabada M.O., Visconti P.E., Arranz S.E. Transmembrane adenylyl cyclase regulates amphibian sperm motility through protein kinase A activation. Dev. Biol. 2011;350:80–88. doi: 10.1016/j.ydbio.2010.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Spehr M., Schwane K., Riffell J.A., Barbour J., Zimmer R.K., Neuhaus E.M., Hatt H. Particulate adenylate cyclase plays a key role in human sperm olfactory receptor-mediated chemotaxis. J. Biol. Chem. 2004;279:40194–40203. doi: 10.1074/jbc.M403913200. [DOI] [PubMed] [Google Scholar]
- 12.Livera G., Xie F., Garcia M.A., Jaiswal B., Chen J., Law E., Storm D.R., Conti M. Inactivation of the mouse adenylyl cyclase 3 gene disrupts male fertility and spermatozoon function. Mol. Endocrinol. 2005;19:1277–1290. doi: 10.1210/me.2004-0318. [DOI] [PubMed] [Google Scholar]
- 13.Wertheimer E., Krapf D., de la Vega-Beltran J.L., Sanchez-Cardenas C., Navarrete F., Haddad D., Escoffier J., Salicioni A.M., Levin L.R., Buck J., et al. Compartmentalization of distinct cAMP signaling pathways in mammalian sperm. J. Biol. Chem. 2013;288:35307–35320. doi: 10.1074/jbc.M113.489476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nomura M., Beltran C., Darszon A., Vacquier V.D. A soluble adenylyl cyclase from sea urchin spermatozoa. Gene. 2005;353:231–238. doi: 10.1016/j.gene.2005.04.034. [DOI] [PubMed] [Google Scholar]
- 15.Yoshida R., Sasakura Y. Establishment of enhancer detection lines expressing GFP in the gut of the ascidian Ciona intestinalis. Zool. Sci. 2012;29:11–20. doi: 10.2108/zsj.29.11. [DOI] [PubMed] [Google Scholar]
- 16.Yoshida M., Inaba K., Morisawa M. Sperm chemotaxis during the process of fertilization in the ascidians Ciona-savignyi and Ciona-intestinalis. Dev. Biol. 1993;157:497–506. doi: 10.1006/dbio.1993.1152. [DOI] [PubMed] [Google Scholar]
- 17.Izumi H., Marian T., Inaba K., Oka Y., Morisawa M. Membrane hyperpolarization by sperm-activating and -attracting factor increases cAMP level and activates sperm motility in the ascidian Ciona intestinalis. Dev. Biol. 1999;213:246–256. doi: 10.1006/dbio.1999.9367. [DOI] [PubMed] [Google Scholar]
- 18.Hancock G.J., Gray J. The propulsion of sea-urchin spermatozoa. J. Exp. Biol. 1955;32:802–814. [Google Scholar]
- 19.Nolan M.A., Babcock D.F., Wennemuth G., Brown W., Burton K.A., McKnight G.S. Sperm-specific protein kinase A catalytic subunit Calpha2 orchestrates cAMP signaling for male fertility. Proc. Natl. Acad. Sci. USA. 2004;101:13483–13488. doi: 10.1073/pnas.0405580101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhu L., Inaba K. Lipid rafts function in Ca2+ signaling responsible for activation of sperm motility and chemotaxis in the ascidian Ciona intestinalis. Mol. Reprod. Dev. 2011;78:920–929. doi: 10.1002/mrd.21382. [DOI] [PubMed] [Google Scholar]
- 21.Jaiswal B.S., Conti M. Calcium regulation of the soluble adenylyl cyclase expressed in mammalian spermatozoa. Proc. Natl. Acad. Sci. USA. 2003;100:10676–10681. doi: 10.1073/pnas.1831008100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Seamon K.B., Daly J.W. Forskolin: Its biological and chemical properties. Adv. Cyclic. Nucleotide Protein Phosphorylation Res. 1986;20:1–150. [PubMed] [Google Scholar]
- 23.Tang W.J., Hurley J.H. Catalytic mechanism and regulation of mammalian adenylyl cyclases. Mol. Pharmacol. 1998;54:231–240. doi: 10.1124/mol.54.2.231. [DOI] [PubMed] [Google Scholar]
- 24.Vijayaraghavan S., Hoskins D.D. Forskolin stimulates bovine epididymal sperm motility and cyclic AMP levels. J. Cycl. Nucleotide Protein Phosphorylation Res. 1985;10:499–510. [PubMed] [Google Scholar]
- 25.Chen Y., Cann M.J., Litvin T.N., Iourgenko V., Sinclair M.L., Levin L.R., Buck J. Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor. Science. 2000;289:625–628. doi: 10.1126/science.289.5479.625. [DOI] [PubMed] [Google Scholar]
- 26.Kleinboelting S., Diaz A., Moniot S., van den Heuvel J., Weyand M., Levin L.R., Buck J., Steegborn C. Crystal structures of human soluble adenylyl cyclase reveal mechanisms of catalysis and of its activation through bicarbonate. Proc. Natl. Acad. Sci. USA. 2014;111:3727–3732. doi: 10.1073/pnas.1322778111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wennemuth G., Carlson A.E., Harper A.J., Babcock D.F. Bicarbonate actions on flagellar and Ca2+-channel responses: Initial events in sperm activation. Development. 2003;130:1317–1326. doi: 10.1242/dev.00353. [DOI] [PubMed] [Google Scholar]
- 28.Brokaw C.J. CO2-inhibition of the amplitude of bending of triton-demembranated sea urcin sperm flagella. J. Exp. Biol. 1977;71:229–240. doi: 10.1242/jeb.71.1.229. [DOI] [PubMed] [Google Scholar]
- 29.Cohn E.J. Studies in the physiology of spermatozoa. Biol. Bull. 1918;34:167–218. doi: 10.2307/1536264. [DOI] [Google Scholar]
- 30.Inaba K., Dreanno C., Cosson J. Control of flatfish sperm motility by CO2 and carbonic anhydrase. Cell Motil. Cytoskelet. 2003;55:174–187. doi: 10.1002/cm.10119. [DOI] [PubMed] [Google Scholar]
- 31.Wang D., King S.M., Quill T.A., Doolittle L.K., Garbers D.L. A new sperm-specific Na+/H+ exchanger required for sperm motility and fertility. Nat. Cell Biol. 2003;5:1117–1122. doi: 10.1038/ncb1072. [DOI] [PubMed] [Google Scholar]
- 32.Wang D., Hu J., Bobulescu I.A., Quill T.A., McLeroy P., Moe O.W., Garbers D.L. A sperm-specific Na+/H+ exchanger (sNHE) is critical for expression and in vivo bicarbonate regulation of the soluble adenylyl cyclase (sAC) Proc. Natl. Acad. Sci. USA. 2007;104:9325–9330. doi: 10.1073/pnas.0611296104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Darszon A., Beltran C., Felix R., Nishigaki T., Trevino C.L. Ion transport in sperm signaling. Dev. Biol. 2001;240:1–14. doi: 10.1006/dbio.2001.0387. [DOI] [PubMed] [Google Scholar]
- 34.Shiba K., Baba S.A., Inoue T., Yoshida M. Ca2+ bursts occur around a local minimal concentration of attractant and trigger sperm chemotactic response. Proc. Natl. Acad. Sci. USA. 2008;105:19312–19317. doi: 10.1073/pnas.0808580105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mizuno K., Shiba K., Okai M., Takahashi Y., Shitaka Y., Oiwa K., Tanokura M., Inaba K. Calaxin drives sperm chemotaxis by Ca2+-mediated direct modulation of a dynein motor. Proc. Nat. Acad. Sci. USA. 2012;109:20497–20502. doi: 10.1073/pnas.1217018109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shiba K., Marian T., Krasznai Z., Baba S.A., Morisawa M., Yoshida M. Na+/Ca2+ exchanger modulates the flagellar wave pattern for the regulation of motility activation and chemotaxis in the ascidian spermatozoa. Cell Motil. Cytoskelet. 2006;63:623–632. doi: 10.1002/cm.20149. [DOI] [PubMed] [Google Scholar]
- 37.Inaba K., Morisawa S., Morisawa M. Proteasomes regulate the motility of salmonid fish sperm through modulation of cAMP-dependent phosphorylation of an outer arm dynein light chain. J. Cell Sci. 1998;111:1105–1115. doi: 10.1242/jcs.111.8.1105. [DOI] [PubMed] [Google Scholar]
- 38.Inaba K., Kagami O., Ogawa K. Tctex2-related outer arm dynein light chain is phosphorylated at activation of sperm motility. Biochem. Biophys. Res. Commun. 1999;256:177–183. doi: 10.1006/bbrc.1999.0309. [DOI] [PubMed] [Google Scholar]
- 39.Hayashi H., Yamamoto K., Yonekawa H., Morisawa M. Involvement of tyrosine protein kinase in the initiation of flagellar movement in rainbow trout spermatozoa. J. Biol. Chem. 1987;262:16692–16698. [PubMed] [Google Scholar]
- 40.Baxendale R.W., Fraser L.R. Immunolocalization of multiple Ga subunits in mammalian spermatozoa and additional evidence for Galphas. Mol. Reprod. Dev. 2003;65:104–113. doi: 10.1002/mrd.10295. [DOI] [PubMed] [Google Scholar]
- 41.Beltran C., Vacquier V.D., Moy G., Chen Y., Buck J., Levin L.R., Darszon A. Particulate and soluble adenylyl cyclases participate in the sperm acrosome reaction. Biochem. Biophys. Res. Commun. 2007;358:1128–1135. doi: 10.1016/j.bbrc.2007.05.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wade M.A., Roman S.D., Jones R.C., Aitken R.J. Adenylyl cyclase isoforms in rat testis and spermatozoa from the cauda epididymidis. Cell Tissue Res. 2003;314:411–419. doi: 10.1007/s00441-003-0801-6. [DOI] [PubMed] [Google Scholar]
- 43.Yoshida M., Inaba K., Ishida K., Morisawa M. Calcium and cyclic-AMP mediate sperm activation, but Ca2+ alone contributes sperm chemotaxis in the ascidian, Ciona savignyi. Dev. Growth Differ. 1994;36:589–595. doi: 10.1111/j.1440-169X.1994.00589.x. [DOI] [PubMed] [Google Scholar]
- 44.Tamura K., Peterson D., Peterson N., Stecher G., Nei M., Kumar S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011;28:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]