

After injury, damaged axons have the capacity to regenerate, but the regenerative capacity of the axon, particularly axons of the central nervous system, is quite limited. This is because the damaged axons tend to retract, because they encounter obstacles such as scar tissue and inhibitory molecules, and because their growth rates simply do not match those of a juvenile axon. In recent years, there has been a focus on microtubules as among the most important factors in encouraging injured adult axons to regenerate. Microtubules are hollow polymeric filaments composed of tubulin subunits that provide structural support for the axon. In addition to their structural role, microtubules are an important substrate for many of the molecular motor proteins responsible for intracellular transport. Microtubules are intrinsically polar structures, with their “plus” ends favored for assembly over their “minus” ends. Molecular motor proteins interact with cargoes such as membranous organelles that are transported in conjunction with the motor. In the axon, the microtubules are aligned into a paraxial array with the plus ends of the microtubules directed away from the cell body, thus establishing the directionality with which different motors convey their cargoes. Microtubules gather together and funnel into the hillock region of the axon and then splay apart again at sites of branch formation and within the growth cone at the tip of the elongating axon. Microtubules are relevant to axonal growth and regeneration for reasons related to all of these factors. Additionally, it appears that the dynamic properties of microtubules are critically important especially in the distal tip of the axon, for the capacity of the axon to form a viable growth cone, to turn properly in response to external cues, and to grow with the vitality typical of the developing nervous system.
Proof-of-principle on the importance of microtubules for regeneration of injured adult axons has come from studies indicating that taxol, a potent microtubule-stabilizing drug, can positively impact that capacity of injured adult CNS axons to regenerate (Hellal et al., 2011; Sengottuvel et al., 2011). However, other studies suggest that the key to axonal regeneration is transforming the rather stable microtubules in the adult axon into a more labile/dynamic population, especially in the distal area of the axon (Bradke et al., 2012). Interestingly, recent work indicates the importance of the status of post-translational tubulin modifications, as it appears that axons regenerate better in the PNS because the microtubules in the damaged region of the axon become less acetylated (Cho and Cavalli, 2012). Such a reduction in microtubule acetylation does not occur in the CNS, suggesting that tubulin modifications that accompany microtubule stability negatively impact the capacity of the axon to regenerate. Taxol treatment notably increases microtubule acetylation, suggesting that taxol's positive effects are not due to recapitulating the mechanisms of axonal growth that occur during development. Thus, these various observations all implicate microtubules, but there is confusion as to exactly what should be done in terms of treatment to best augment regeneration. Taxol's positive effects may be due more to its impact on non-neuronal cells relevant to nerve regeneration, and may transiently appear to give axonal growth a boost because stabilization of microtubules abnormally prevents axonal retraction. Stabilized microtubules may also enable the tip of the axon to act more like a battering ram to push the axon through normally inhibitory environments, but that kind of mechanism is far askew of how the normally dynamic growth cone of the axon functions during development. A better therapeutic approach, in my opinion, would exploit the normal mechanisms by which the microtubule array expands during development, with emphasis on adding more labile microtubule mass, as opposed to artificially stabilizing what is already there (Baas and Ahmad, 2013).
A plethora of microtubule-related proteins impacts the capacity of the axon to grow and to navigate. Optimally, we would like to identify molecules that can be manipulated to enable CNS axons to grow faster and to enable them to ignore/overcome inhibitory molecules associated with the CNS and the injury site. We have reported that drugs that inhibit kinesin-5, a microtubule-based motor protein, promote the capacity of axons to grow faster and to grow onto inhibitory molecules, especially if used in combination with other approaches (Lin et al., 2011). However, this is not due to the copious addition of new labile microtubule mass to the axon, but rather to a shift in the balance of forces on the existing microtubule array. Potential shortcomings with this approach would be that inhibiting kinesin-5 could be deleterious to the navigation of the regenerating axon to its appropriate target and that inhibiting kinesin-5 really does nothing to shift the status of the microtubule array toward expansion nor does it shift the balance of stable to labile microtubule mass. We think we could do much better in terms of promoting axonal regeneration if we could identify molecules that normally suppress the expansion of the labile microtubule mass, and then develop techniques for inhibiting those molecules in a controlled therapeutic fashion.
In our laboratory, we have been focusing for the past several years on a category of microtubule-related enzymes that function to cut or break microtubules. These enzymes are called microtubule-severing proteins. They are AAA proteins that form hexamers on the surface of the microtubule, and yank on a tubulin subunit such that it is extracted from the microtubule lattice, causing the microtubule to break. Microtubule severing, as this process is called, can occur at the centrosome to release the minus end of the microtubule so that it can then transit into an axon or a dendrite. Severing can also occur at the plus end, in which case, subunits are peeled off of the normally dynamic end of the microtubule in a depolymerizing manner. Perhaps most interesting is what happens when the severing protein breaks the microtubule somewhere along its length. Each microtubule in the axon consists of a stable domain, with most of them also consisting of a labile domain that assembles directly from the plus end of the stable domain (Baas and Ahmad, 2013). If the severing event occurs in the stable domain, the result would be two new microtubules, one with a stable and labile domain, and the other being exclusively a stable fragment that can then assemble a new labile domain from available free tubulin subunits. In this fashion, severing of stable microtubule domains is a mechanism within the axon to create new microtubules, which is critically important given that the centrosome is quite a distance away, within the cell body of the neuron. If the severing event occurs in the labile domain, however, the result would be very different, as there would be one microtubule with a stable domain and a shorter labile domain, but the microtubule fragment without a stable domain would presumably depolymerize into subunits (Figure 1). Thus, severing of the labile domain would not create new microtubules, but rather would work to pare away the labile domains, keeping them shorter and a less robust component of the total microtubule array than they would otherwise be. We posit that certain microtubule-severing proteins target the stable domains and other microtubule-severing proteins target the labile domains. We posit that the expression levels and activities of these severing proteins are tightly regulated to enable the rapid expansion of the microtubule array during axonal development, and then to tamp down the expansion of the microtubule array once the axon has reached its target.
Figure 1.
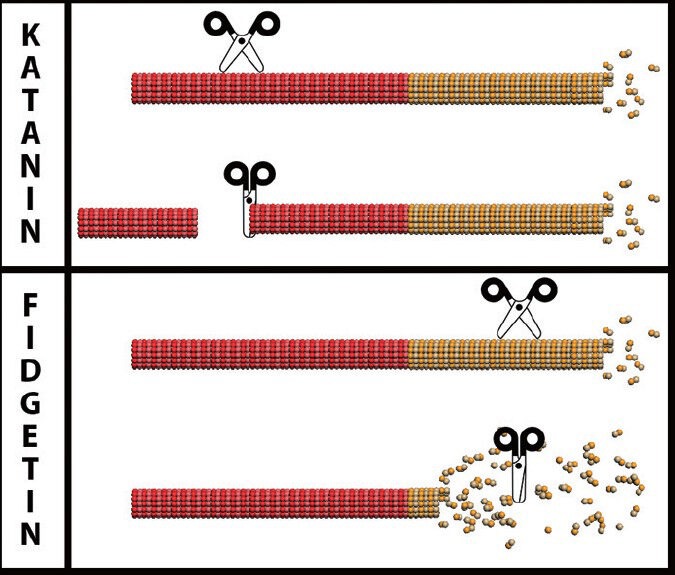
Microtubule-severing proteins have very different effects on microtubules, depending on the preference of the particular severing protein for stable or labile domains of microtubules.
Microtubules in the axon consist of a stable domain toward the minus end of the microtubule and a labile domain toward the plus end. Shown in the figure are microtubules with stable and labile domains indicated in red and yellow respectively. Severing proteins are shown as scissors. Severing in the stable domain would increase microtubule number, while severing in the labile domain would pare down the labile domain without creating more microtubules. Katanin and spastin preferentially sever stable domains whereas we posit that fidgetins preferentially sever labile domains. Figure prepared by Andrew Matamoros.
Katanin and spastin, the two best studied microtubule-severing proteins in neurons, both have a preference for severing stable domains, and appear to do so by targeting regions of microtubules that are especially rich in tubulins that have been post-translationally acetylated or polyglutamylated (Lacroix et al., 2010; Sudo and Baas, 2010). Suppressing katanin or spastin has deleterious effects on axonal growth and branching (Yu et al., 2008), as would be expected, given the important roles these proteins play in generating new microtubules via the severing of existing ones. There are two other “katanin-like” proteins expressed in vertebrates; while less is known about katanin-like-1 and katanin-like-2 than katanin, they are probably similar to katanin in having a preference for stable microtubule domains. Even less is known about a family of three proteins termed fidgetins that are also AAA proteins with putative microtubule-severing properties (Sharp and Ross, 2012). Fidgetin and fidgetin-like-2 have been shown to sever microtubules in vitro. We have speculated that these fidgetins might be specialized to sever labile domains of microtubules, potentially by targeting microtubule regions that are less rich in post-translationally modified tubulins (Baas and Ahmad, 2013). We have preliminary data that have led us to believe that this might well be the case, and that inhibition of fidgetin expression may have positive affects on injured adult neurons toward promoting the expansion of the labile microtubule mass of the axon in a manner conducive to axonal growth and regeneration (Figure 2).
Figure 2.
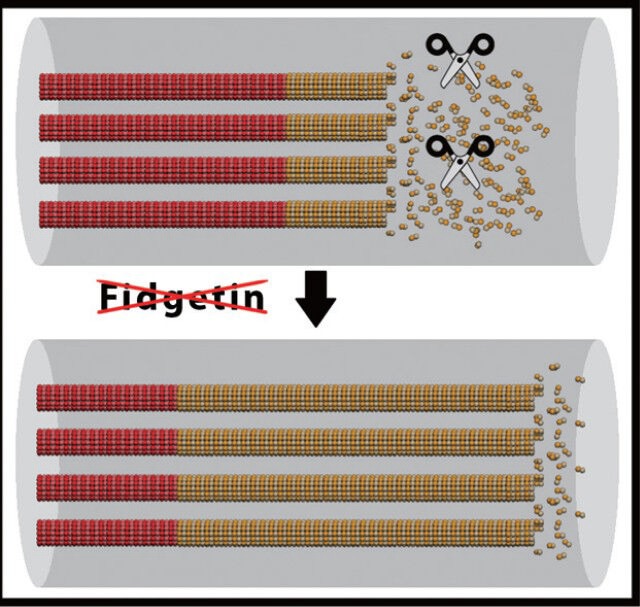
Fidgetin, a potential regulator of labile microtubule mass in the axon, may be a powerful target for therapeutic intervention to augment regeneration of injured adult axons.
If this hypothesis is correct, suppressing fidgetins would result in an expansion of labile microtubule domains, which we posit should be conducive to axonal regeneration. Shown in the figure are microtubules with stable and labile domains indicated in red and yellow respectively, with fidgetin shown as scissors. In the figure, for clarity of the hypothesis, the microtubules are shown with stable and labile domains aligned, but in actual fact, the microtubules are staggered along the length of the axon, so that stable and labile domains are not in alignment. Figure prepared by Andrew Matamoros.
References
- Baas PW, Ahmad FJ. Beyond taxol: microtubule-based treatment of disease and injury of the nervous system. Brain. 2013;136:2937–2951. doi: 10.1093/brain/awt153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradke F, Fawcett JW, Spira ME. Assembly of a new growth cone after axotomy: The precursor to axon regeneration. Nat Rev Neurosci. 2012;13:183–193. doi: 10.1038/nrn3176. [DOI] [PubMed] [Google Scholar]
- Cho Y, Cavalli V. HDAC5 is a novel injury-regulated tubulin deacetylase controlling axon regeneration. EMBO J. 2012;31:3063–3078. doi: 10.1038/emboj.2012.160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hellal F, Hurtado A, Ruschel J, Flynn KC, Laskowski CJ, Umlauf M, Kapitein LC, Strikis D, Lemmon V, Bixby J, Hoogenraad CC, Bradke F. Microtubule stabilization reduces scarring and causes axon regeneration after spinal cord injury. Science. 2011;331:928–931. doi: 10.1126/science.1201148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacroix B, van Dijk J, Gold ND, Guizetti J, Aldrian-Herrada G, Rogowski K, Gerlich DW, Janke C. Tubulin polyglutamylation stimulates spastin-mediated microtubule severing. J Cell Biol. 2010;189:945–954. doi: 10.1083/jcb.201001024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin S, Liu M, Son YJ, Timothy Himes B, Snow DM, Yu W, Baas PW. Inhibition of kinesin-5, a microtubule-based motor protein, as a strategy for enhancing regeneration of adult axons. Traffic. 2011;12:269–286. doi: 10.1111/j.1600-0854.2010.01152.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sengottuvel V, Leibinger M, Pfreimer M, Andreadaki A, Fischer D. Taxol facilitates axon regeneration in the mature CNS. J Neurosci. 2011;31:2688–2699. doi: 10.1523/JNEUROSCI.4885-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp DJ, Ross JL. Microtubule-severing enzymes at the cutting edge. J Cell Sci. 2012;125:2561–2569. doi: 10.1242/jcs.101139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudo H, Baas PW. Acetylation of microtubules influences their sensitivity to severing by katanin in neurons fibroblasts. J Neurosci. 2010;30:7215–7226. doi: 10.1523/JNEUROSCI.0048-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu W, Qiang L, Solowska JM, Karabay A, Korulu S, Baas PW. The microtubule-severing proteins spastin and katanin participate differently in the formation of axonal branches. Mol Biol Cell. 2008;19:1485–1498. doi: 10.1091/mbc.E07-09-0878. [DOI] [PMC free article] [PubMed] [Google Scholar]