

Abstract
Allergens, viral, and bacterial infections are responsible for asthma exacerbations that occur with progression of airway inflammation. cPLA2 α and sPLA2X are responsible for delivery of arachidonic acid for production of eicosanoids—one of the key mediators of airway inflammation. However, cPLA2 α and sPLA2X role in allergic inflammation has not been fully elucidated. The aim of this study was to analyze the influence of rDer p1 and rFel d1 and lipopolysaccharide (LPS) on cPLA2 α expression and sPLA2X secretion in PBMC of asthmatics and in A549 cell line. PBMC isolated from 14 subjects, as well as A549 cells, were stimulated with rDer p1, rFel d1, and LPS. Immunoblotting technique was used to study the changes in cPLA2 α protein expression and ELISA was used to analyze the release of sPLA2X. PBMC of asthmatics released more sPLA2X than those from healthy controls in the steady state. rDer p1 induced more sPLA2X secretion than cPLA2 α protein expression. rFel d1 caused decrease in cPLA2 α relative expression in PBMC of asthmatics and in A549 cells. Summarizing, Der p1 and Fel d1 involve phospholipase A2 enzymes in their action. sPLA2X seems to be one of important PLA2 isoform in allergic inflammation, especially caused by house dust mite allergens.
1. Introduction
Despite intensive studies on asthma pathogenesis and seeking the effective treatment, the number of new cases is increasing globally. Asthma is a very heterogeneous disease whose etiology has not been fully understood. Allergens, drugs, viral and bacterial infections, and stress are the most common factors that initiate and exacerbate asthma. About 70% of asthmatics are atopic [1]. The proposed mechanism of allergic asthma development suggests that allergen exposure causes sensitization, and continued exposure leads to airway hyperresponsiveness and inflammation. Airway inflammation primarily initiated as a defense process aiming to eliminate the damaging factor, which evolves to chronic state causing airway remodeling and impaired lung functions. In general population of Lodz province (Poland) the most common sensitizing indoor allergens are house dust mites and cat [2]. Sensitivity to house dust mite and cat dander are risk factors associated with the development of asthma [3]. Many studies indicate that allergen exposure causes the exacerbation of asthma that occurs with impaired lung function and increases the need for hospitalization [4, 5].
Group X secretory phospholipase A2 (sPLA2X) has recently been investigated as one of the most important members of secretory PLA2 in the inflammatory process [6]. Except its enzymatic activity sPLA2 can act through the membrane receptors causing cell degranulation and initiating chemokines and cytokines production [7, 8]. Moreover, sPLA2 can influence cytosolic PLA2 (cPLA2) action [9, 10]. In human airways a lot of resident cells (mast cells, macrophages, endothelial cells, epithelial cells, and bronchial smooth muscle cells (SMC)) and haematopoetic cells (basophils, eosinophils, neurophils, lymphocytes, and monocytes) are potential source of secretory phospholipases [11]. In asthmatics expression of sPLA2-X predominates in airway epithelium. Moreover, both sPLA2-X and sPLA2-IIA are the main phospholipases detected in BAL fluid [6, 12]. sPLA2-X and sPLA2-XII are elevated in induced sputum cells of patients with asthma [13]. The studies with knockout mice showed that deficiency of sPLA2-X reduced allergen-induced features of airway inflammation [14].
Cytosolic phospholipase A2 group IVA (cPLA2 α) is the most potent enzyme in phospholipase A2 superfamily catalyzing liberation of arachidonic acid (AA) from membrane phospholipids [15]. Our previous studies revealed that cPLA2 α participates in asthma pathogenesis [16]. What is more, rDer p1 caused overexpression of PLA2G4A in PBMC of asthmatics (unpublished data). Whalen et al. showed that PBMC of asthmatic patients stimulated with allergens in the presence of cPLA2 inhibitor exhibited decreased production of proinflammatory cytokines [17]. cPLA2 actions are mainly regulated by Ca2+ concentration and serine residue phosphorylation [18, 19]. LPS can modulate activity of cPLA2 by phosphorylation [20]. Der p1 can activate MAPKs in different types of cells [21, 22]. Despite the abovementioned facts that prove the Der p1-cPLA2 α interactions, other mechanisms of allergens impact on lipid mediators remains not fully understood. Thus, we investigated whether allergens or LPS can directly stimulate the expression and/or phosphorylation of cPLA2 protein in PBMC of severe asthmatics with atopic origin.
2. Material
2.1. Patients
Patients (n = 7) with severe asthma, who were allergic to house dust mite (Der p1) and cat (Fel d1) allergens, and healthy controls (n = 7) were enrolled to the study. The project was approved by the local ethics committee and an informed consent was obtained from every subject prior to the study. Patients were recruited from the Department of Internal Diseases, Asthma and Allergy of Medical University of Lodz. Asthma was recognized at least 6 months prior to the study and met the criteria of GINA Guidelines [23]. The severity of the disease was assessed according to the American Thoracic Society Workshop on Refractory Asthma 2000 Report [24]. All patients were classified as severe asthmatics. Patients were asked to not administer antihistamine drugs, oral gluccocorticoids and leukotriene receptors antagonists 24 hours, and inhaled glucocorticoids and long-acting beta agonists 12 hours before blood drawing. Healthy volunteers had no known history of asthma or seasonal allergies. Table 1 presents characteristics of subjects enrolled to the study.
Table 1.
Characteristics of patients with bronchial asthma and healthy subjects.
| Asthmatic patients | Healthy volunteers | |
|---|---|---|
| N | 7 | 7 |
| Age (years) | 48 (39–72) | 36 (24–41) |
| Women/men | 5/2 | 5/2 |
| Smokers/nonsmokers | 1/7 | 0/7 |
| Inhaled GCS (μg/day)a | 1508 (960–1600) | 0 |
| Systemic GCS (mg/day)b | 18,8 (7–20) | 0 |
| FEV1 (%) | 74.6 | N/A |
| Allergic to Der p1 |
6/7 | No |
| Allergic to Fel d1 |
3/7 | No |
aDoses were calculated as budesonide equivalents. bDoses were calculated as prednisone equivalents.
2.2. PBMC
Peripheral blood mononuclear cells (PBMC) were isolated using Histopaque 1077 (Sigma-Aldrich, St. Louis, MO), the density gradient cell separation medium according to the producer's instructions. Cells were cultured in RPMI1640 (Sigma-Aldrich, St. Louis, MO) with 10% heat-inactivated FBS (Sigma-Aldrich, St. Louis, MO) and antibiotics. 10 ng/mL of polymyxin B was added to medium used in allergen stimulation and 100 U/mL penicillin and 100 μg/mL streptomycin (Sigma-Aldrich, St. Louis, MO) for LPS incubation. 2 × 106/mL PBMC were stimulated in vitro with LoTox deglycosylated recombinant Dermatophagoides pteronyssinus allergen 1 (rDer p1), LoTox deglycosylated recombinant Felis domesticus allergen 1 (rFel d1) (Indoor Biotechnologies, Cardiff, UK) or LPS from E. coli, serotype R515 (Enzo Life Sciences, NY). In dose-response systems three concentrations of allergens: 1 μg/mL, 5 μg/mL, and 10 μg/mL and LPS: 50 ng/mL, 100 ng/mL, and 500 ng/mL were tested (at 24 h). In time-course system 5 μg/mL of each allergen and 100 ng/mL of LPS were used and cells were collected in various time points: 0.5 h, 1 h, 2 h, 6 h, and 24 h.
2.3. A549 Culture
A549 cells, a human adenocarcinoma cell line, were obtained from the European Collection of Cell Cultures, Heath Protection Agency (Salisbury, UK) and were grown in Ham's F-12 K medium (Sigma-Aldrich, St. Louis, MO) with 10% fetal bovine serum (Sigma-Aldrich, St. Louis, MO), 2 mM of L-glutamine (Sigma-Aldrich, St. Louis, MO), 100 unit/mL penicillin, and 100 μg/mL streptomycin (Sigma-Aldrich, St. Louis, MO). All experiments were performed when cells were 80% to 90% confluent. Cells were stimulated in vitro with LoTox deglycosylated recombinant Dermatophagoides pteronyssinus allergen 1 (rDer p1), LoTox deglycosylated recombinant Felis domesticus allergen 1 (rFel d1) (Indoor Biotechnologies, Cardiff, UK), or LPS from E. coli, serotype R515 (Enzo Life Sciences, NY).
3. Methods
3.1. Immunoblotting
Total protein from PBMC of patients with asthma, healthy subjects, and A549 cells was extracted in RIPA protein extraction buffer (Sigma-Aldrich, St. Louis, MO), supplemented with protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO). The lysate was centrifuged at 14,000 RPM and 4°C for 20 min, and the pellet discarded. Protein concentrations were determined by the BCA Protein Assay Kit (Pierce Thermo Scientific, Rockford, IL) according to the manufacturer's protocol and using bovine serum albumin as a standard. 20 μg of total protein was mixed with NuPAGE LDS Sample Buffer (Life Technologies, Carlsbad, CA) and in a 1 : 10 ratio with NuPAGE Reducing Agent (10x), heated for 10 min at 70°C. Protein samples were subjected to electrophoresis in 4–12% SDS-NuPAGE Gels (Life Technologies, Carlsbad, CA) at 200 V and electrophoretically transferred to a nitrocellulose membrane at 30 V for one hour. The membrane was blocked in 5% nonfat milk in TBST (20 mM Tris-HCL, 500 mM NaCl, 0.05% Tween 20, and pH 7.5) for 1 hour at room temperature. Then, the membranes were incubated for 12 h at 4°C with the one of the following antibody: polyclonal rabbit anti-cPLA2 and anti-phospho-cPLA2 (Ser505) and anti-β-actin antibodies (Cell Signaling, Danvers, MA). At the end of the overnight incubation, the membrane was washed with TBST and incubated for one hour in TBST containing the goat anti-rabbit IgG secondary antibodies conjugated with alkaline phosphatase (Sigma-Aldrich, St. Louis, MO). After incubation with secondary antibodies, the membrane was washed three times (3 × 5 mins) in TBST buffer. The band was developed using BCIP/NBT alkaline phosphatase substrate (Merck Millipore, Darmstadt, Germany). Densitometric analysis of bands was performed with Image J 1.34s software (Wayne Rasband, National Institutes of Health, Bethesda, MD) and the results are presented as fold change of optical density (OD).
3.2. ELISA
The sPLA2X protein in supernatants form PBMC and A549 cells was measured by enzyme-linked immunoabsorbent assay (ELISA) using commercially available kit (Cloud-Clone Corp., Houston, TX) according to the manufacturer's protocol. The limit of detection for sPLA2X protein was 7.813 pg/mL.
3.3. Statistical Analysis
The data were analyzed using Statistica (v. 10.0; StatSoft, Tulsa, OK). Comparisons between groups were performed by using Mann-Whitney U tests or ANOVA followed by the Tukey's post hoc test when appropriate. Values of P < 0.05 were considered statistically significant.
4. Results
4.1. PBMC of Asthmatics Overproduced sPLA2X in the Steady State
Hallstrand et al. [13] reported that sputum cells from asthmatics contained more sPLA2X mRNA (qPCR) and protein (immunostaining) in comparison to controls, so we hypothesized that similar observation in PBMC is very possible. Taking into account the fact that sPLA2X is produced as a zymogen and its cellular amount may not be relevant to its biological function we concluded that secretion of sPLA2X will be the best approach to discover its potential involvement in asthma pathogenesis. The levels of sPLA2X released by PBMC were compared in asthmatics and controls. The steady state concentration of sPLA2X was significantly higher in asthmatics (411.09 pg/mL ± 129.2) than in healthy subjects (91.96 pg/mL ± 16.37) (Figure 1).
Figure 1.
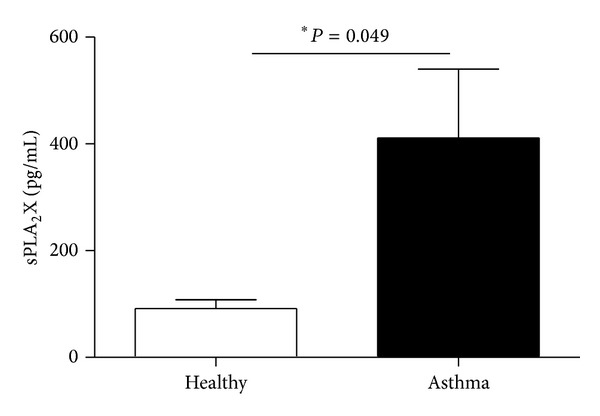
Concentration of sPLA2X released by PBMC of asthmatics and healthy controls. 2 × 106 PBMC were incubated in medium containing RPMI1640 with 10% heat-inactivated FBS and antibiotics (37°C, 5% CO2). Supernatants were collected after 24 hours of incubation in 37°C. The secretion of sPLA2X was measured in indirect ELISA. Data are presented as mean ± SEM; ∗P < 0.05 as comparison between studied groups.
4.2. rDer p1 Stimulation Results in Different sPLA2X and cPLA2 Production Patterns in Asthmatics and Controls
PBMC of asthmatics and healthy subjects were stimulated with rDer p1 for 24 hours in three different concentrations: 1, 5, and 10 μg/mL. There were no differences in sPLA2X secretion between asthmatics and controls in any dose of rDer p1 whereas dust mite allergen in concentration of 10 μg/mL significantly induced release of sPLA2X (1.54 ± 0.24) when compared with relative protein expression of cPLA2 α (0.79 ± 0.17) in asthmatics (Figure 2(a)). On the contrary, in healthy subjects, we did not observe similar fluctuation (Figure 2(b)).
Figure 2.
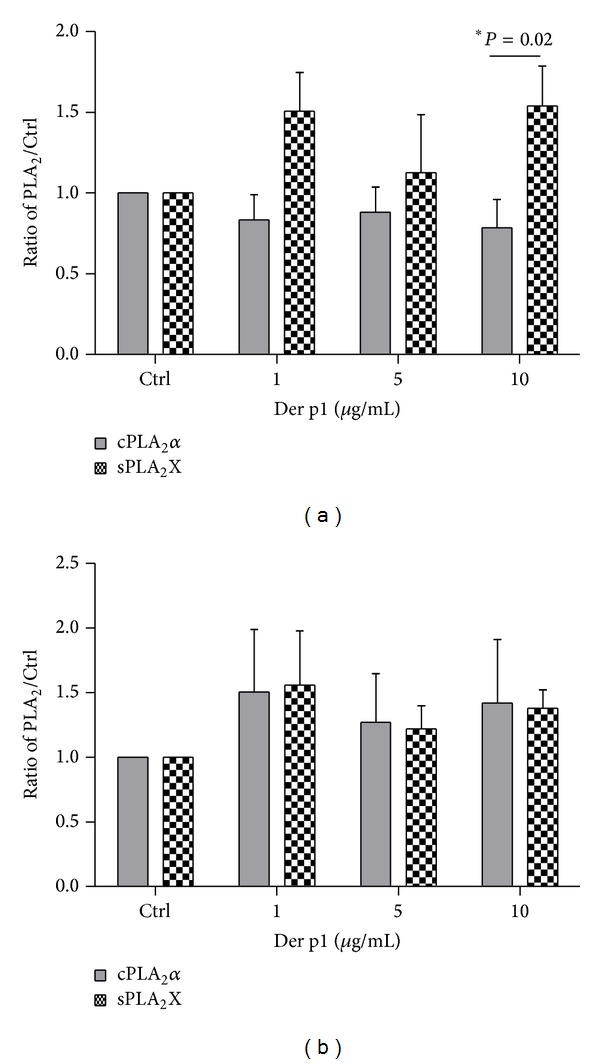
sPLA2X secretion and relative expression of cPLAα in PBMC of asthmatics (a) and healthy subjects (b) in response to rDer p1. PBMC (2 × 106) were stimulated with indicated doses of rDer p1 for 24 hours. Control represents cells treated with the vehicle. The bar graph shows the densitometry results for cPLA2 (immunoblotting results) and ELISA results for sPLA2X secretion. Data are presented as the fold change compared with the vehicle-treated cells. Data represent the mean ± SE from at least six independent experiments. ∗P < 0.05 shows comparison between relative protein expression of cPLA2 and secretion of sPLA2X.
4.3. Regulation of Relative cPLA2 Protein Expression by LPS, rFel d1, and rDer p1 in PBMC in Time-Dependent Manner
Relative expression of cPLA2 α protein was compared between healthy and asthmatic patients after stimulation with rDer p1 (5 μg/mL), rFel d1 (5 μg/mL), and LPS (100 ng/mL). cPLA2 α basal expression was significantly lower in asthmatics as compared to healthy subjects. While we did not observe differences in cPLA2 α protein synthesis between patients and controls after stimulation, there was a statistically significant increase of cPLA2 α protein expression in PBMC of severe asthmatics in all tested time points after stimulation with rDer p1 when compared to steady state level. Also rFel d1 induced expression of cPLA2 α protein after 0.5 h and 6 h of stimulation in patients. 24-hour incubation with LPS results in induction of cPLA2 in asthmatics when compared with level before stimulation. In healthy subjects cPLA2 α protein expression did not change significantly over time (Figures 3 and 4—blot).
Figure 3.
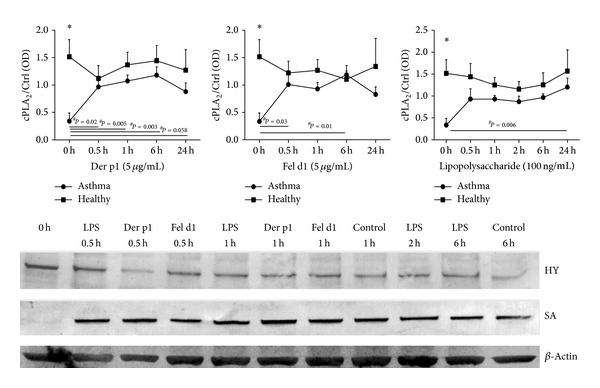
Relative cPLA2 α protein expression in PBMC from healthy subjects and asthmatic patients in vitro stimulated with rDer p1, rFel d1, and LPS in time-dependent manner. PBMC (2 × 106) were stimulated with 100 ng/mL LPS or rDer p1 (5 μg/mL) or rFel d1 (5 μg/mL) at the indicated time. Control represents cells treated with the vehicle. The immunoblot is representative of experiments in PBMC from at least six donors, each showing similar results. The line graph shows the densitometry results. Data are presented as the fold change compared with the vehicle-treated cells. The point “0” indicates cPLA2 content in PBMC freshly isolated from blood, without culturing (untreated PBMC). Data represent the mean ± SE from at least six independent experiments. # P < 0.05 shows comparison with untreated cells; ∗P < 0.05 shows comparison between asthmatics and healthy subjects. HY: healthy subjects. SA: severe asthmatics.
Figure 4.
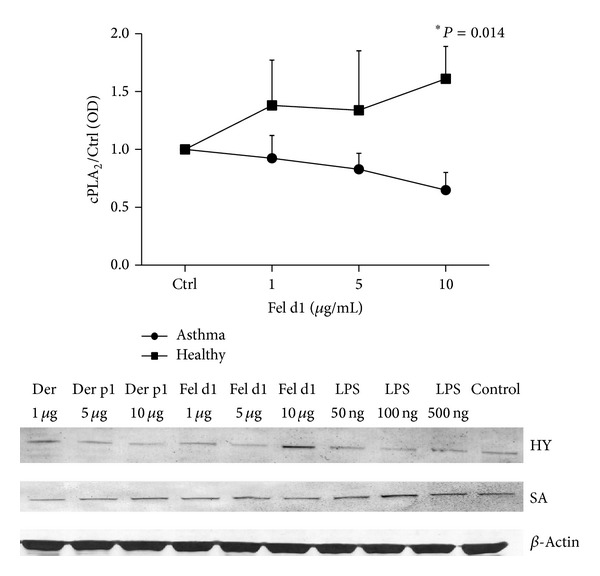
Relative cPLA2 α protein expression in PBMC from healthy subjects and asthmatic patients in vitro stimulated with rDer p1, rFel d1, and LPS in dose-dependent manner. PBMC (2 × 106) were stimulated with indicated doses of LPS or rDer p1 or rFel d1 for 24 hours. Control represents cells treated with the vehicle. The immunoblot is representative of experiments in PBMC from at least six donors, each showing similar results. The line graph shows the densitometry results obtained from cells stimulated with rFel d1. Data are presented as the fold change compared with the vehicle-treated cells (control). Data represent the mean ± SE from at least six independent experiments. ∗P < 0.05shows comparison between studied groups. HY: healthy subjects. SA: severe asthmatics.
4.4. cPLA2 Protein Synthesis Is Diminished by rFel d1 in PBMC of Asthmatics
PBMC from asthmatics and healthy subjects were stimulated with rDer p1, rFel d1, and LPS in three different concentrations for 24 hours. While being not significant (as compared to control), there was a trend of increased relative expression of cPLA2 in healthy subjects and decreased protein content in asthmatics after stimulation with rFel d1. However PBMC from asthmatics produced significantly less cPLA2 α (0.65 ± 0.15) than those from healthy subjects (1.61 ± 0.28) after stimulation with rFel d1 in concentration of 10 μg/mL (Figure 4).
4.5. LPS Induced cPLA2 Phosphorylation in PBMC of Healthy Subjects
Phosphorylation of cPLA2 α was analyzed in PBMC stimulated with rDer p1 (5 μg/mL), rFel d1 (5 μg/mL), and LPS (100 ng/mL). The cells from controls (1.38 ± 0.22) contained more phosphorylated form of cPLA2 α than patients' PBMC (0.87 ± 0.1) after 2 h of incubation with LPS. The rapid change in phosphorylation of cPLA2 α was observed after 6 h stimulation with LPS (Figure 5). The allergens did not change the phosphorylation of cPLA2.
Figure 5.
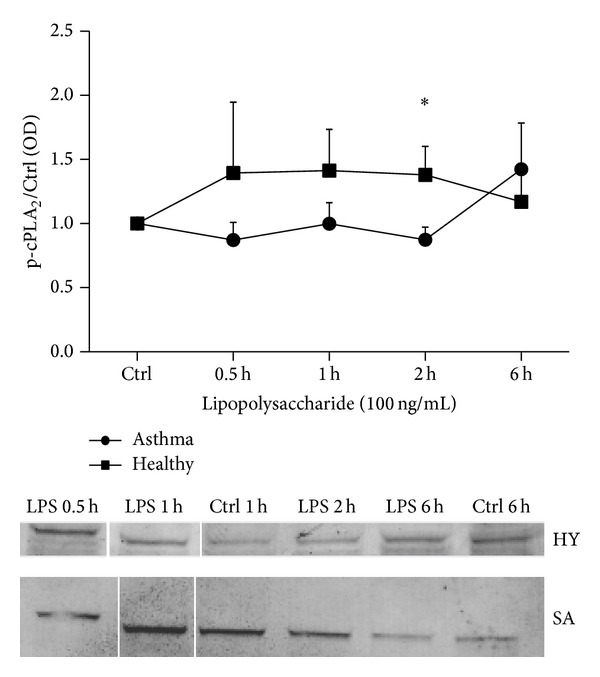
Phosphorylation of cPLA2 α protein in PBMC from healthy subjects and asthmatic patients in vitro stimulated with LPS. PBMC (2 × 106) were stimulated with 100 ng/mL of LPS at indicated timepoints. Control represents cells treated with the vehicle. The immunoblot is representative of experiments in PBMC from at least six donors, each showing similar results. The line graph shows the densitometry results obtained from PBMC stimulated with LPS. Dataare presented as the fold change compared with the vehicle-treated cells. Data represent the mean ± SE from at least six independent experiments. ∗P < 0.05 shows comparison between studied groups.
4.6. Regulation of cPLA2 Protein Synthesis by LPS, rFel d1, and rDer p1 in A549 Cells
In A549 culture the rFel d1 in concentration 10 μg/mL significantly decreased synthesis of cPLA2 (0.61 ± 0.01) (Figure 6). Any other stimulators did not change cPLA2 expression in short time of incubation.
Figure 6.
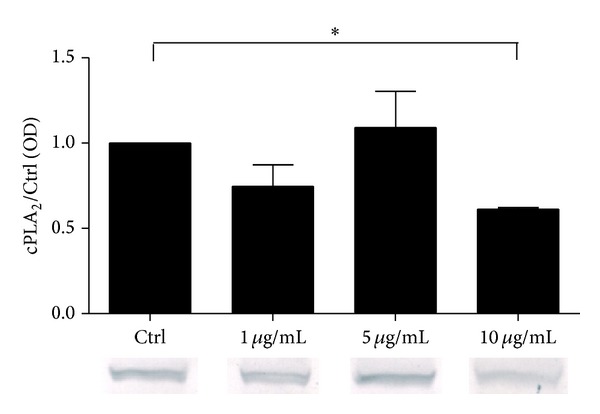
Relative cPLA2 α protein expression in A549 cells stimulated with rFel d1. A549 were stimulated with rFel d1 (5 μg/mL) for 24 hours. Control represents cells treated with the vehicle. The immunoblot is representative of three independent experiments in A549 cells, each showing similar results. The bar graph shows the densitometry results. Data are presented as the fold change compared with the vehicle-treated cells. Data represent the mean ± SE from at least three independent experiments. ∗P < 0.05 shows comparison with untreated cells.
4.7. Recombinant Der p1 Induces Morphological Changes in A549 Cells
rDer p1 dose- and time-dependently caused morphological changes in A549 cells. Low concentrations and short incubation did not induce visible changes whereas higher concentrations and longer incubations led to cells shrinking and desquamation (Figure 7).
Figure 7.
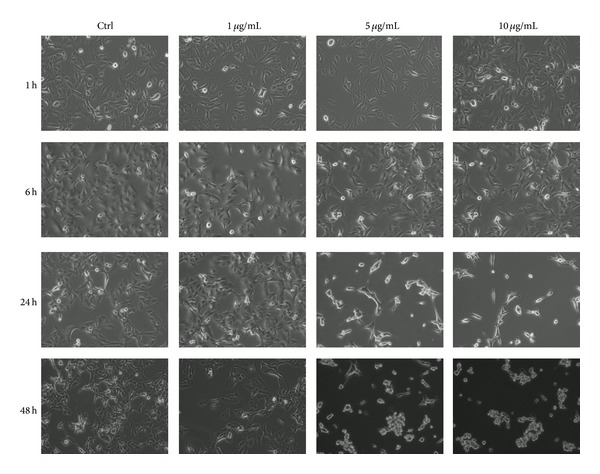
Morphological changes of A549 cells caused by rDerp1.
5. Discussion
We observed that asthmatics' PBMC released more sPLA2X than control cells in the steady state. This observation is supported by previous reports, showing that asthmatics have increased content of sPLA2X in airway epithelium, BALF, and sputum. The novelty of our study relates to regulation of different PLA2 isoforms in response to rDer p1 stimulation. Interestingly rDer p1 in highest dose (10 μg/mL) significantly upregulated release of sPLA2X protein when compared to relative cPLA2 α protein expression in asthmatics whereas in healthy subjects we did not observe this tendency. This observation suggests that sPLA2X may be one of the important isoforms of PLA2 in allergic response. Despite evidence that both enzymes cooperate in liberation of AA, sPLA2X is also able to release AA independently to cPLA2 α [25] and therefore may alone promote allergen-induced inflammation. Misso et al. suggested that increased activity of sPLA2 may be associated with atopic status [26]. Some data related to the role of sPLA2X in airway inflammation come also from animal studies. Knock-in of human sPLA2X to msPLA2X−/− mice restored allergen-induced inflammatory cell recruitment into airways as well as hyperresponsiveness to methacholine [27].
PBMC stimulated with rDer p1, rFel d1, or LPS showed increased production of cPLA2 α in comparison to steady state level in asthmatics but not in healthy subjects. The most rapid changes were observed after rDer p1 action, whereas 24-hour incubation with LPS was needed to induce significant increase of cPLA2 α content. The mechanism of Der p1 action is still not fully determined. Der p1 acts by PAR-2 receptor as well as in PAR-2-independent manner through activation of NF-κB and ERK1/2 [22, 28]. Activation of NF-κB pathway is prerequisite for cPLA2 expression in many cell types [29–31], so this pathway can partially be involved in the induction of cPLA2 α expression. In our study we did not observe the changes in production of cPLA2 α between asthmatics and controls after stimulation with allergens and LPS (time-response scheme).
Dose-response scheme of our experiment showed that rFel d1 in highest dose (10 μg/mL) significantly decreased expression of cPLA2 α in asthmatics when compared to healthy subjects. The similar significant decrease of cPLA2 synthesis was observed in A549 cells after stimulation with rFel d1. Fel d1 has been shown to may have enzymatic activity [32]. The structural analysis of Fel d1 revealed homology of the allergen with α-subunit of mouse salivary androgen binding protein [33], uteroglobin and with the related Clara cell phospholipid binding protein, CC10 [34]. Uteroglobin is anti-inflammatory protein and can inhibit PLA2 activity [35]. The similarity between rFel d1 and uteroglobin may suggest that the allergen has cytokine-like properties; thus it may be capable of inflecting the immune response [34].
Although Der p1 and Fel d1 are able to increase activity and phosphorylation of cPLA2 in eosinophils [36], in our study we did not observe the significant changes in cPLA2 α phosphorylation after allergen stimulation. Only stimulation with LPS resulted in elevated level of phosphorylated cPLA2 α form in healthy subjects. PBMC of healthy volunteers respond better to LPS treatment. The diverse effects of LPS action have been observed earlier [37, 38]. Moreover it has been proved that LPS-induced cPLA2 activity is TLR-4-dependent [39]. Different experiment systems and doses of LPS used in experiments seem to condition the results of LPS stimulation. In U937 cell line and macrophages LPS significantly increased the expression of cPLA2 protein after 8 hours but not after 24 hours of stimulation whereas in tracheal smooth muscle cells the effective time points were 16 and 24 hours [30, 40].
rDer p1 in higher doses induces the desquamation of A549 cells. This effect was observed earlier and is result of enzymatic activity of the allergen [28]. Der p1 is a cysteine protease able to degrade the occludin protein and ZO-1 protein in tight junction between epithelial cells [28, 41].
6. Conclusions
Results of the study showed that Der p1 and Fel d1 involve phospholipase A2 enzymes in their action. sPLA2X seems to be the more important PLA2 isoform in airway inflammation, especially caused by house dust mite allergens. Der p1 has protease activity and can actively degrade and penetrate the epithelium barrier in airways. Moreover stimulation of sPLA2X production whose activity is connected with further cysteinyl leukotrienes synthesis may sustain inflammatory process. This phenomenon might be supported additionally by decreased synthesis of cPLA2 and subsequent diminished PGE2 synthesis, which in respiratory track may also play protective role. Fel d1 seems to act rather by decreasing the cPLA2 expression than induction of sPLA2X. Further studies focusing on expression of different PLA2 isoforms in different timepoints after inflammatory stimulus exposition should be analyzed to better understand the molecular mechanism of allergic inflammation.
Acknowledgments
The paper was supported from science budget for years 2010–2013 as research project (N N402 516939) and Medical University of Lodz Grants 502-03/0-149-03/502-04-004, 502-03/0-149-03/502-04-021, and 503/0-149-03/503-01. This paper was partially funded by Polish National Science Centre Grant no. DEC-2012/05/N/NZ5/02630.
Conflict of Interests
The authors declare that they have no conflict of interests.
References
- 1.WHO. Global surveillance, prevention and control of chronic respiratory diseases: a comprehensive approach. WHO, 2007.
- 2.Kupryś-Liphińka I, Elgalal A, Kuna P. Skin prick test with inhaled allergens in the general population of Lodz province. Pneumonologia i Alergologia Polska. 2009;77(3):229–234. [PubMed] [Google Scholar]
- 3.Sears MR, Herbison GP, Holdaway MD, Hewitt CJ, Flannery EM, Silva PA. The relative risks of sensitivity to grass pollen, house dust mite and cat dander in the development of childhood asthma. Clinical and Experimental Allergy. 1989;19(4):419–424. doi: 10.1111/j.1365-2222.1989.tb02408.x. [DOI] [PubMed] [Google Scholar]
- 4.Green RM, Custovic A, Sanderson G, Hunter J, Johnston SL, Woodcock A. Synergism between allergens and viruses and risk of hospital admission with asthma: case-control study. British Medical Journal. 2002;324(7340):763–766. doi: 10.1136/bmj.324.7340.763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Murray CS, Poletti G, Kebadze T, et al. Study of modifiable risk factors for asthma exacerbations: virus infection and allergen exposure increase the risk of asthma hospital admissions in children. Thorax. 2006;61(5):376–382. doi: 10.1136/thx.2005.042523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hallstrand TS, Lai Y, Ni Z, et al. Relationship between levels of secreted phospholipase A2 groups IIA and X in the airways and asthma severity. Clinical and Experimental Allergy. 2011;41(6):801–810. doi: 10.1111/j.1365-2222.2010.03676.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Granata F, Nardicchi V, Loffredo S, et al. Secreted phospholipases A2: a proinflammatory connection between macrophages and mast cells in the human lung. Immunobiology. 2009;214(9-10):811–821. doi: 10.1016/j.imbio.2009.06.006. [DOI] [PubMed] [Google Scholar]
- 8.Granata F, Frattini A, Loffredo S, et al. Signaling events involved in cytokine and chemokine production induced by secretory phospholipase A2 in human lung macrophages. European Journal of Immunology. 2006;36(7):1938–1950. doi: 10.1002/eji.200535567. [DOI] [PubMed] [Google Scholar]
- 9.Fonteh AN, Atsumi G, LaPorte T, Chilton FH. Secretory phospholipase A2 receptor-mediated activation of cytosolic phospholipase A2 in murine bone marrow-derived mast cells. The Journal of Immunology. 2000;165(5):2773–2782. doi: 10.4049/jimmunol.165.5.2773. [DOI] [PubMed] [Google Scholar]
- 10.Offer S, Yedgar S, Schwob O, et al. Negative feedback between secretory and cytosolic phospholipase A2 and their opposing roles in ovalbumin-induced bronchoconstriction in rats. American Journal of Physiology—Lung Cellular and Molecular Physiology. 2005;288(3):L523–L529. doi: 10.1152/ajplung.00199.2004. [DOI] [PubMed] [Google Scholar]
- 11.Triggiani M, Granata F, Giannattasio G, Marone G. Secretory phospholipases A2 in inflammatory and allergic diseases: not just enzymes. Journal of Allergy and Clinical Immunology. 2005;116(5):1000–1006. doi: 10.1016/j.jaci.2005.08.011. [DOI] [PubMed] [Google Scholar]
- 12.Lai Y, Oslund RC, Bollinger JG, et al. Eosinophil cysteinyl leukotriene synthesis mediated by exogenous secreted phospholipase A2 group X. The Journal of Biological Chemistry. 2010;285(53):41491–41500. doi: 10.1074/jbc.M110.153338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hallstrand TS, Chi EY, Singer AG, Gelb MH, Henderson WR., Jr. Secreted phospholipase A2 group X overexpression in asthma and bronchial hyperresponsiveness. The American Journal of Respiratory and Critical Care Medicine. 2007;176(11):1072–1078. doi: 10.1164/rccm.200707-1088OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Henderson WR, Jr., Chi EY, Bollinger JG, et al. Importance of group X-secreted phospholipase A2 in allergen-induced airway inflammation and remodeling in a mouse asthma model. Journal of Experimental Medicine. 2007;204(4):865–877. doi: 10.1084/jem.20070029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Leslie CC. Properties and regulation of cytosolic phospholipase A2 . The Journal of Biological Chemistry. 1997;272(27):16709–16712. doi: 10.1074/jbc.272.27.16709. [DOI] [PubMed] [Google Scholar]
- 16.Sokolowska M, Stefanska J, Wodz-Naskiewicz K, Cieslak M, Pawliczak R. Cytosolic phospholipase A2 group IVA is overexpressed in patients with persistent asthma and regulated by the promoter microsatellites. Journal of Allergy and Clinical Immunology. 2010;125(6):1393–1395. doi: 10.1016/j.jaci.2010.02.016. [DOI] [PubMed] [Google Scholar]
- 17.Whalen KA, Legault H, Hang C, et al. In vitro allergen challenge of peripheral blood induces differential gene expression in mononuclear cells of asthmatic patients: inhibition of cytosolic phospholipase A2 α overcomes the asthma-associated response. Clinical and Experimental Allergy. 2008;38(10):1590–1605. doi: 10.1111/j.1365-2222.2008.03059.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lin L, Wartmann M, Lin AY, Knopf JL, Seth A, Davis RJ. CPLA2 is phosphorylated and activated by MAP kinase. Cell. 1993;72(2):269–278. doi: 10.1016/0092-8674(93)90666-e. [DOI] [PubMed] [Google Scholar]
- 19.Evans JH, Gerber SH, Murray D, Leslie CC. The calcium binding loops of the cytosolic phospholipase A2 C2 domain specify targeting to Golgi and ER in live cells. Molecular Biology of the Cell. 2004;15(1):371–383. doi: 10.1091/mbc.E03-05-0338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tian W, Wijewickrama GT, Jung HK, et al. Mechanism of regulation of group IVA phospholipase A2 activity by Ser727 phosphorylation. The Journal of Biological Chemistry. 2008;283(7):3960–3971. doi: 10.1074/jbc.M707345200. [DOI] [PubMed] [Google Scholar]
- 21.Wong CK, Wang CB, Li MLY, Ip WK, Tian YP, Lam CWK. Induction of adhesion molecules upon the interaction between eosinophils and bronchial epithelial cells: Involvement of p38 MAPK and NF-κB. International Immunopharmacology. 2006;6(12):1859–1871. doi: 10.1016/j.intimp.2006.08.003. [DOI] [PubMed] [Google Scholar]
- 22.Adam E, Hansen KK, Fernandez OA, et al. Erratum: The house dust mite allergen der p 1, unlike der p 3, stimulates the expression of interleukin-8 in human airway epithelial cells via a proteinase-activated receptor-2-independent mechanism. Journal of Biological Chemistry. 2007;282(7):p. 5100. doi: 10.1074/jbc.M507140200. [DOI] [PubMed] [Google Scholar]
- 23. Global Strategy for Asthma Management and Prevention, Global Initiative for Asthma. In; 2011.
- 24.Wenzel SE, Fahy JV, Irvin C, et al. Proceedings of the ATS workshop on refractory asthma: current understanding, recommendations, and unanswered questions. American Journal of Respiratory and Critical Care Medicine. 2000;162(6):2341–2351. doi: 10.1164/ajrccm.162.6.ats9-00. [DOI] [PubMed] [Google Scholar]
- 25.Saiga A, Uozumi N, Ono T, et al. Group X secretory phospholipase A2 can induce arachidonic acid release and eicosanoid production without activation of cytosolic phospholipase A2 alpha. Prostaglandins and Other Lipid Mediators. 2005;75(1–4):79–89. doi: 10.1016/j.prostaglandins.2004.10.001. [DOI] [PubMed] [Google Scholar]
- 26.Misso NLA, Petrovic N, Grove C, Celenza A, Brooks-Wildhaber J, Thompson PJ. Plasma phospholipase A2 activity in patients with asthma: association with body mass index and cholesterol concentration. Thorax. 2008;63(1):21–26. doi: 10.1136/thx.2006.074112. [DOI] [PubMed] [Google Scholar]
- 27.Henderson WR, Jr., Oslund RC, Bollinger JG, et al. Blockade of human group X secreted phospholipase A2 (GX-sPLA 2)-induced airway inflammation and hyperresponsiveness in a mouse asthma model by a selective GX-sPLA 2 inhibitor. The Journal of Biological Chemistry. 2011;286(32):28049–28055. doi: 10.1074/jbc.M111.235812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kauffman HF, Tamm M, Timmerman JAB, Borger P. House dust mite major allergens Der p 1 and Der p 5 activate human airway-derived epithelial cells by protease-dependent and protease-independent mechanisms. Clinical and Molecular Allergy. 2006;4, article 5 doi: 10.1186/1476-7961-4-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hsieh H, Wu C, Hwang T, Yen M, Parker P, Yang C. BK-induced cytosolic phospholipase A2 expression via sequential PKC-δ, p42/p44 MARK, and NF-κB activation in rat brain astrocytes. Journal of Cellular Physiology. 2006;206(1):246–254. doi: 10.1002/jcp.20457. [DOI] [PubMed] [Google Scholar]
- 30.Luo SF, Lin WN, Yang CM, et al. Induction of cytosolic phospholipase A2 by lipopolysaccharide in canine tracheal smooth muscle cells: involvement of MAPKs and NF-κB pathways. Cellular Signalling. 2006;18(8):1201–1211. doi: 10.1016/j.cellsig.2005.09.011. [DOI] [PubMed] [Google Scholar]
- 31.Luo S, Lin C, Chen H, et al. Involvement of MAPKs, NF-kappaB and p300 co-activator in IL-1beta-induced cytosolic phospholipase A2 expression in canine tracheal smooth muscle cells. Toxicology and Applied Pharmacology. 2008;232(3):396–407. doi: 10.1016/j.taap.2008.07.019. [DOI] [PubMed] [Google Scholar]
- 32.Ring PC, Wan H, Schou C, Kristensen AK, Roepstorff P, Robinson C. The 18-kDa form of cat allergen Felis domesticus 1 (Fel d 1) is associated with gelatin- and fibronectin-degrading activity. Clinical and Experimental Allergy. 2000;30(8):1085–1096. doi: 10.1046/j.1365-2222.2000.00805.x. [DOI] [PubMed] [Google Scholar]
- 33.Karn RC. The mouse salivary androgen-binding protein (ABP) alpha subunit closely resembles chain 1 of the cat allergen Fel dI. Biochemical Genetics. 1994;32(7-8):271–277. doi: 10.1007/BF00555830. [DOI] [PubMed] [Google Scholar]
- 34.Kaiser L, Grönlund H, Sandalova T, et al. The crystal structure of the major cat allergen Fel d 1, a member of the secretoglobin family. Journal of Biological Chemistry. 2003;278(39):37730–37735. doi: 10.1074/jbc.M304740200. [DOI] [PubMed] [Google Scholar]
- 35.Chowdhury B, Mantile-Selvaggi G, Miele L, Cordella-Miele E, Zhang Z, Mukherjee AB. Lys 43 and Asp 46 in α-helix 3 of uteroglobin are essential for its phospholipase A2-inhibitory activity. Biochemical and Biophysical Research Communications. 2002;295(4):877–883. doi: 10.1016/s0006-291x(02)00767-2. [DOI] [PubMed] [Google Scholar]
- 36.Seeds MC, Peachman KK, Bowton DL, Sivertson KL, Chilton FH. Regulation of arachidonate remodeling enzymes impacts eosinophil survival during allergic asthma. The American Journal of Respiratory Cell and Molecular Biology. 2009;41(3):358–366. doi: 10.1165/rcmb.2008-0192OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hernandez ML, Herbst M, Lay JC, et al. Atopic asthmatic patients have reduced airway inflammatory cell recruitment after inhaled endotoxin challenge compared with healthy volunteers. Journal of Allergy and Clinical Immunology. 2012;130(4):869.e2–876.e2. doi: 10.1016/j.jaci.2012.05.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chun E, Lee S, Shim E, et al. Toll-like receptor expression on peripheral blood mononuclear cells in asthmatics; implications for asthma management. Journal of Clinical Immunology. 2010;30(3):459–464. doi: 10.1007/s10875-009-9363-z. [DOI] [PubMed] [Google Scholar]
- 39.Qi H, Shelhamer JH. Toll-like receptor 4 signaling regulates cytosolic phospholipase A2 activation and lipid generation in lipopolysaccharide-stimulated macrophages. Journal of Biological Chemistry. 2005;280(47):38969–38975. doi: 10.1074/jbc.M509352200. [DOI] [PubMed] [Google Scholar]
- 40.Jiang YJ, Lu B, Choy PC, Hatch GM. Regulation of cytosolic phospholipase A2, cyclooxygenase-1 and -2 expression by PMA, TNFα, LPS and M-CSF in human monocytes and macrophages. Molecular and Cellular Biochemistry. 2003;246(1-2):31–38. [PubMed] [Google Scholar]
- 41.Wan H, Winton HL, Soeller C, et al. Quantitative structural and biochemical analyses of tight junction dynamics following exposure of epithelial cells to house dust mite allergen Der p 1. Clinical and Experimental Allergy. 2000;30(5):685–698. doi: 10.1046/j.1365-2222.2000.00820.x. [DOI] [PubMed] [Google Scholar]