

Abstract
Increasing data shows miR-29a is a key regulator of oncogenic processes. It is significantly down-regulated in some kind of human tumors and possibly functionally linked to cellular proliferation, survival and migration. However, the mechanism remains unclear. In this study, we report miR-29a is significantly under-expressed in gastric cancer compared to the healthy donor. The microvessel density is negatively related to miR-29a expression in gastric cancer tissues. The ectopic expression of miR-29a significantly inhibits proliferation and invasion of gastric cancer cells. Furthermore, western blot combined with the luciferase reporter assays demonstrate that vascular endothelial growth factor A (VEGF-A) is direct target of miR-29a. This is the first time miR-29a was found to suppress the tumor microvessel density in gastric cancer by targeting VEGF-A. Taken together, these results suggest that miR-29a is a tumor suppressor in gastric cancer. Restoration of miR-29a in gastric cancer may be a promising therapeutic approach. [BMB Reports 2014; 47(1):39-44]
Keywords: Gastric cancer, Hsa-miR-29a, MicroRNAs, Post-transcriptional regulation, VEGF-A
INTRODUCTION
As a new class of regulatory molecules, miRNAs are found to play critical roles in a broad range of biological events (1,2). Based on computer-aided predictions, miRNAs are estimated to regulate about one-third of the human genes (3), including many of the tumor-related genes (4). Many miRNAs are located at fragile sites or cancer-associated regions (5). Maybe it explains why miRNAs could play a key role in human malignancy. miRNAs can act either as oncogenes (6,7) or as tumor suppressors (8-10), as well as cancer biomarkers for diagnosis or prognosis predication (11). Moreover, miR-29a is a highly conserved miRNA among different species, being expressed on the transcript from a locus at chromosome 7q32.3 which coincides with the common fragile site FRA7H (12). For example, miR-29a has been shown to be down-regulated in anaplastic large-cell lymphomas (13), which contributes to apoptosis blockade through MCL-1 overexpression. However the expression and function of miR-29a in gastric cancer has seldom been reported yet. In this study, we report that the expression level of miR-29a is related to vascularization in gastric cancer tissues, and suppresses proliferation and invasion in SGC-7901 cells by targeting VEGF-A.
RESULTS
Down-regulated expression of miR-29a in gastric cancer
To explore the potential role of miR-29a in gastric cancer, we performed quantitative real-time PCR analysis of miR-29a expression in a panel of fresh frozen gastric mucosa tissues (26 samples of gastric cancer without metastasis, 20 samples of gastric cancers with metastasis, and 15 samples from healthy donors) in a double-blinded fashion. We found that miR-29a expression level was significantly lower in tissues from gastric cancer with metastasis, compared with that from gastric cancer without metastasis (0.97 ± 0.05 vs. 1.26 ± 0.04, P < 0.01, Fig. 1A), and miR-29a expression level was significantly lower in gastric cancer without metastasis compared with that in gastric mucosa from healthy donor (1.26 ± 0.04 vs. 1.58 ± 0.10, P < 0.01, Fig. 1A).
Fig. 1. Expression of miR-29a in gastric cancer and healthy donors. (A) HD: healthy donor; GC-NM: gastric cancer without metastasis; GC-M: gastric cancer with metastasis. (B) Microvessel density in gastric cancer without metastasis and with metastasis. (C) Relation between miR-29a and microvessel density (MVD) in gastric cancer. Each sample was analyzed in triplicate.
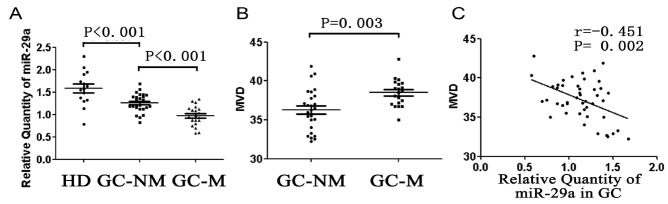
Microvessel density is negatively related to miR-29a expression in gastric cancer
Our findings revealed that microvessel density (MVD) was significantly higher in gastric cancer with metastasis than that in gastric cancer without metastasis (38.49 ± 0.39 vs. 36.29 ± 0.53, P < 0.01, Fig. 1B). What’s more, MVD was inversely related to miR-29a expression level in gastric cancer (Fig. 1C).
Growth inhibition of gastric cancer cell lines by miR-29a
The above results suggest that miR-29a may function as a tumor suppressor in gastric cancer. To test this hypothesis, we performed proliferation assays in human gastric cancer cell line SGC-7901 and MKN-45 with miR-29a over expression. Gastric cancer cells were infected with either miR-29a expression lentivirus (Lv-miR29a) or non-specific cel-mir-67 expression lentivirus (Lv-cel, as negative control). The quantitative real-time PCR confirmed that the gastric cancer cells infected with Lv-miR29a expressed two times higher of exogenous miR29a than that infected with Lv-cel (SGC-7901: 2.20 ± 0.12 vs. 1.10 ± 1.15, P < 0.01; MKN-45: 2.79 ± 0.16 vs. 1.17 ± 0.14, P < 0.01, Fig. 2A). MTT assays indicated that the growth of gastric cancer cells infected with Lv-miR29a was slower than that infected with Lv-cel (Fig. 2B). At the sixth day, the absorbance at 490 nm of gastric cancer cells infected with Lv-miR29a was significantly lower than that infected with Lv-cel (SGC-7901: 0.52 ± 0.06 vs. 0.77 ± 0.10, P < 0.01; MKN-45: 0.68 ± 0.04 vs. 0.88 ± 0.05, P < 0.01), while the gastric cancer cells infected neither of the lentivirus showed no obviously difference from the gastric cancer cells infected with Lv-cel (Fig. 2B).
Fig. 2. miR-29a suppresses cell growth and invasion of gastric cancer cells in vitro. (A) miR-29a expression in GES-1, SGC-7901 and MKN-45 cells. Lv-miR29a: lentivirus carrying pre-miR-29a; Lv-CEL: lentivirus carrying non-specific cel-mir-67. (B) Cell growth evaluated by MTT assays. The viabilities of SCG-7901-Lv-miR29a and MKN45-Lv-miR29a cells were both significantly decreased and the highest inhibitory rates were 30.85 ± 6.44 and 22.08 ± 2.67% on day 6, respectively. (C) miR-29a inhibits cell invasion in vitro. Gastric cancer cells were first infected with lentivirus carrying pre-miR-29a or cel-mir-67 alone and then were tested for invasion ability in matrigel chambers as described in Materials and Methods.
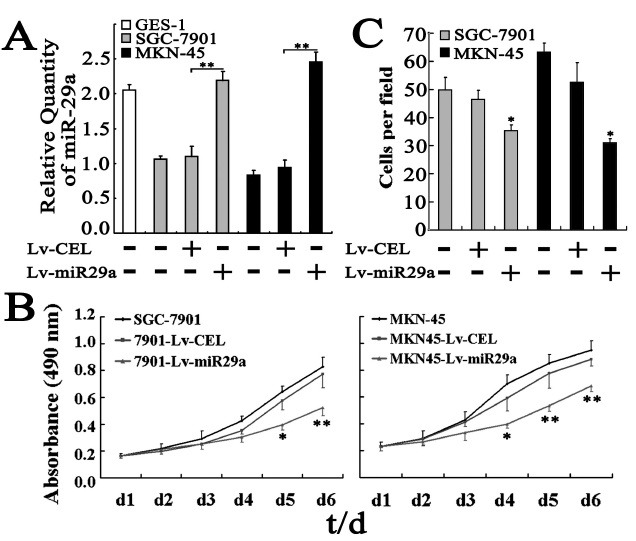
miR-29a suppresses invasiveness in gastric cancer cells
The above result of significantly higher miR-29a expression in gastric cancers without metastasis than that with metastasis suggests that miR-29a may be involved in the invasion of cancer cells. To test this hypothesis, we chose human gastric cancer cell line SGC-7901 and MKN-45 because both cells also expressed lower miR-29a compared to human gastric epithelial cell line GES-1 (Fig. 2A). Matrigel chamber assays indicated that the invasion ability of gastric cancer cells was obviously reduced by exogenous expression of miR-29a (SGC-7901: 35.40 ± 1.94 vs. 46.40 ± 3.26, P < 0.05; MKN-45: 31.00 ± 1.52 vs. 52.60 ± 7.00, P < 0.05, Fig. 2C). The suppressed invasion by miR-29a in SGC-7901 cells suggests that miR-29a may also affect gastric cancer metastasis.
Target gene prediction of miR-29a
To understand the molecular mechanisms by which miR-29a inhibits tumor cell growth and cell invasion, we searched for putative miR-29a targets as predicted by the commonly cited programs including TargetScan (14), DIANA-microT 3.0 (15,16) and miRanda (17) online target gene prediction software and finally VEGF-A was predicted to be a potential target gene of miR-29a. Further analysis by RNAhybrid software (17) showed miR-29a binding VEGF-A 3’UTR with a minimum free energy of −20.2 kcal/mol, predicting the generation of a highly stable duplex (Fig. 3A). These results suggest VEGF-A is most likely a target gene of miR-29a since VEGF-A is a key regulator in angiogenesis and tumor proliferation and metastasis (18).
Fig. 3. Identification of VEGF-A as direct targets for miR-29a. (A) miR-29a binding to VEGF-A 3’UTR. (B) Alignment of miR-29a with VEGF-A at the 3’-UTR and the mutation site of seed area. (C) Relative expression level of miR-29a in HEK-293 cells transfected with different oligo. SC: Scramble oligo as control to miR-29a; miR29a: miR-29a mimic; anti-NC: negative control oligo to anti-miR-29a; anti-29a: anti-miR-29a oligo. (D) Luciferase activity in HEK-293 cells.
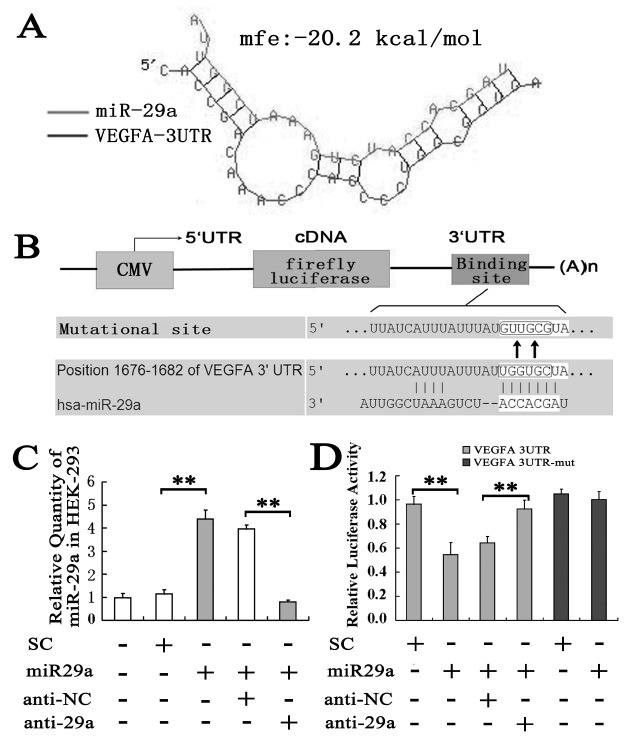
miR-29a directly targets VEGF-A expression
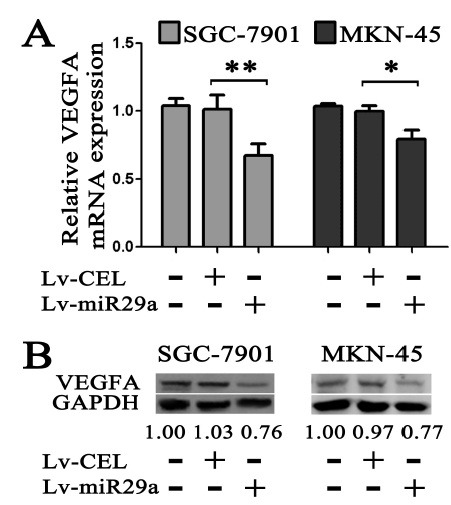
Therefore we constructed luciferase reporters carrying the VEGF-A 3'-UTR (Fig. 3B). The luciferase assay showed that the exogenous miR-29a gave a significant reduction of the luciferase activity (0.55 ± 0.10 vs. 0.97 ± 0.06, P < 0.01, Fig. 3D). But the luciferase activity restored when the anti-miR-29a oligo was transfected along with the miR-29a mimic (Fig. 3D). To further confirm that miR-29a-mediated reduction of the luciferase activity is due to direct interaction between miR-29a and its putative binding site, we mutated the miR-29a binding site by site-directed mutagenesis (Fig. 3B). As expected, the miR-29a-mediated suppression of the luciferase activity was abolished in mutation type compared to scrambled oligo (1.00 ± 0.07 vs. 1.05 ± 0.04, P > 0.05, Fig. 3D), suggesting that the miR-29a binding site is critical for miR-29a-mediated suppression. We also examined the VEGF-A expression in mRNA and protein level by using real-time PCR and western blot. As shown in Fig. 4A, exogenous expression of miR-29a down-regulated the VEGF-A mRNA level significantly, as well as the VEGF-A protein (Fig. 4B).
Fig. 4. mRNA and protein expression of VEGF-A in gastric cancer cells infected with Lv-miR29a. (A) VEGF-A mRNA expression. (B) VEGF-A protein expression. Gastric cancer cell line SGC-7901 or MKN-45 cells were infected with Lv-miR29a and then subjected to mRNA expression assay by real-time PCR or protein expression assay by Western blotting respectively. GAPDH and U6 were used as an internal loading control. A reproducible result was obtained in three independent experiments.
DISCUSSION
Growing evidence has indicated that aberrant expression of certain miRNAs is directly associated with some cancers (19). In addition to the role in cancer, miR-29a is also involved in diabetes (20,21), myogenesis, apoptosis, rhabdomyosarcoma, osteoblastic differentiation, sclerosis (22), Alzheimer's disease (23,24), HIV-1 replication (25), HCV eplication (26), cardiac or liver (27) fibrosis and tumorigenesis (13). Results of this study show the down-regulated expression of miR-29a in gastric cancer with/without metastasis, and its negative relationship with MVD. These findings suggest miR-29a may be a potential suppressor in gastric cancer. To confirm this notion, we demonstrate that ectopic expression of miR-29a suppresses proliferation and invasiveness in gastric cancer cells. Moreover, our results show the expression level of miR-29a inversely correlates to the microvessel density in gastric cancer, suggesting miR-29a takes a role in the inhibition of angiogenesis. Angiogenesis is an early event in tumorigenesis and can facilitates tumor progression and metastasis. To understand the molecular mechanisms of miR-29a suppressing the gastric cancer cells, we searched for putative miR-29a targets by the combination of online target gene prediction softwares and VEGF-A was predicted to be a potential target gene of miR-29a. Further luciferase assays and western blot results confirmed that miR-29a can suppress the endogenous expression level of VEGF-A by binding to the 3’-untranslated region of VEGF-A. Since VEGF-A is an important invasion and metastasis factor in angiogenesis and tumor metastasis, it is easy to understand that the reduced level of VEGF-A could contribute to the observed suppression of cell invasion in vitro and in vivo (28,29). Taken together, we find miR-29a can suppress the angiogenesis in gastric cancer by inhibiting the target gene VEGF-A, which consequently represses the growth and invasion of gastric cancer cells. What’s more, higher expression of VEGF-A and higher microvessel density is a poor prognostic indicator (30,31). Therefore, identification of VEGF-A as a direct target for miR-29a may imply that miR-29a is a novel target for gastric cancer therapy and prognostic indicator.
Since a single miRNA can post-transcriptionally suppress multiple targets, it is probably that miR-29a may also target other tumor-promoting genes simultaneously to inhibit gastric cancer growth in addition to the VEGF-A. For example, a recent report indicates that miR-29a represses Ppm1d phosphatase, which in turn enhances p53 activity and suppresses the growth of liver cancer cells (32). Cui et al. (33) found miR-29a inhibits cell proliferation and induces cell cycle arrest through the down-regulation of p42.3 in human gastric cancer. Therefore the observed miR-29a-mediated inhibition of gastric cancer growth and metastasis is likely due to simultaneous targeting of multiple targets, which may explains slightly expression level change of miR-29a can induce great effects. Further efforts are needed to identify the other possible target genes of miR-29a in suppressing gastric cancer progress.
MATERIALS AND METHODS
Ethics statement
All experimental procedures were approved by the Institutional Review Board of the 324th Hospital of PLA. Written informed consent was obtained for all patient and healthy donor samples.
Clinical samples
Forty-six patients (34 males and 12 females) who had undergone gastrectomy with lymph node dissection for gastric carcinoma and 15 healthy donors at 324th Hospital in 2010 were included in the study. None of the patients received preoperative chemotherapy.
Cell culture
The following cell lines were from the American Type Culture Collection and cultured according to the vendor's instructions: GES-1, SGC-7901, MKN45, and HEK-293 cells.
Real-time RT-PCR
Total RNA from the frozen tissues or cultured cells was isolated with mirVanaTM PARISTM Kit (Ambion, USA) according to the manufacturer’s instructions. RNA was first reversely transcribed into cDNA by using RT reagent Kit (TOYOBO, Japan). Then the cDNA was subjected to real-time PCR with an SYBRⓇ Green Realtime PCR Master Mix kit (TOYOBO, Japan) in an ABI PRISM 7500 system (Applied Biosystems, USA) by using the miR-29a primers set and U6 primers set (Ribobio, China). Human VEGF-A primers for RT-PCR were produced by the Sangon Inc., Shanghai, China. The sequence of VEGFA reverse primer is (34) : 5’-ATGATTCTGCCCTCCTCCTT-3’; and the forward primer is: 5’-CCTTGCTGCTCTACC TCCAC-3’ (74 bp). The relative quantification of RNA expression was calculated using the 2-ΔΔCt method (35).
Microvascular density assessment
Microvascular density (MVD) assessment by CD34 immunohistochemical staining was performed as described by Weidner et al (36). The entire section was scanned at low power (100×) to identify the hot spots. An average count in five hot spots was calculated as MVD at power 400× magnification. Single endothelial cell, endothelial cell clusters and microvessels in the tumor were counted. Peritumoral vascularity, vascularity in areas of necrosis and vessels with thick smooth muscle or in a diameter larger than eight erythrocytes was not scored. All counts were made by three pathologists who had no knowledge of the corresponding clinicopathologic data.
miRNA oligo and lentiviral constructs design and cell transfection
The miR-29a mimic oligo and anti-miR-29a, along with the negative control oligo, were purchased from RiboBio (Guangzhou, China). The LipofectamineⓇ 2000 (Invitrogen, USA) was used for miRNA oligo transfection according to the manufacturer’s manual. Lentiviral constructs containing pre-miR-29a (Lv-miR29a) was purchased from GeneChem Inc., Shanghai, China. A construct including the nonspecific cel-mir-67 (99 bp, MI0000038) was used as a negative control (Lv-CEL) (37). These viruses were used to infect gastric cancer cells at multiplicity of infection (MOI) of 10 and the infection efficiency was 100%.
Cell proliferation assay
The cell viability of gastric cancer cells was measured by a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT, Sigma, USA) assay as described previously (38). Optical densities were determined on a Versamax microplate reader (Molecular Devices, Sunnyvale, CA) at 490 nm.
Invasion assay
The invasion ability of gastric cancer cells was determined using matrigel invasion chambers (BD Biosciences, USA). Gastric cancer cells were seeded into inserts at 5 × 104 per insert in serum-free medium and then transferred to wells filled with the culture medium containing 10% FBS. After 24 h of incubation, non-invading cells on the top of the membrane were removed by scraping. Invaded cells on the bottom of the membrane were fixed, followed by staining with 0.05% crystal violet. The number of invaded cells on the membrane was then counted under a microscope.
Luc-VEGFA vector
We amplified a 343-bp VEGFA 3'-UTR from SGC-7901 cells cDNA using the following PCR primers: VEGFA3-UTR-5, 5'-ATCGGTGACAGTCACTAG-3' and VEGFA3-UTR-3, 5'-TACGGATAAACAGTAGCA-3'. The amplified fragment was first cloned into pCR2.1-TOPO vector (Invitrogen, USA) and subsequently cloned into the SpeI and HindIII sites of the pMIR-REPORT miRNA expression reporter vector (Applied Biosystems, USA). The first six nucleotides complementary to the miR-29a seed-region were mutated from the mutant constructs using the QuikChange Site-Directed Mutagenesis Kit (Stratagene, USA).
Luciferase assay
HEK-293 cells were seeded in 24-well plates 24 h before transfection. Luciferase reporter plasmid (Luc-VEGFA) and pRL-TK control plasmid (Promega) were cotransfected into HEK-293 cells along with scrambled RNA oligo negative control, or miR-29a mimic, or anti-miR29a oligo (RiboBio, Guangzhou, China) by using the LipofectamineⓇ 2000. Cells were harvested 48 h after transfection. Luciferase activity was determined using Dual-luciferase assay system (Promega) according to the manufacturer’s instruction.
Western blot
For western blot analysis, total protein was extracted from gastric cancer cells by using RIPA buffer (50 mM TrisC1, 50 mM NaCl, 10% glycerol, 1% Nonidet P-40, 0.5% deoxycholate, 0.1% SDS, 10 mM NaF, 0.4 mM EDTA, pH 8.0, with leupeptin and aprotinin) and centrifuged at 10,000 × g for 10 min. Samples with 25 μg of total protein were resolved by SDS-PAGE on a 10% gel and transferred onto an NC membrane. The membrane was probed with 1:500 goat primary antibodies against human VEGF-A (sc-152, Sant-Cruz, US), followed the secondary HRP-conjugated anti-goat antibody (Sant-Cruz). Signals were detected using the ECL western blotting analysis system (Amersham Biosciences, USA). GAPDH was used as an endogenous protein for normalization.
Statistical analysis
All data are presented as means ± SE and were analyzed using Prism 5.0 software (GraphPad). The significance of the observed differences was determined with the Student’s t-test. The relationships between the miR-29a and microvessel density were performed Pearson correlation analysis. P < 0.05 was considered statistically significant (*P < 0.05; **P < 0.01).
Acknowledgments
This study was supported by grants from the National Nature Science Foundation of China (No. 30900679) and the Natural Science Foundation Project of CQ CSTC (cstc2010BB5158, cstc2011jjA10111, cstc2011jjA10114, cstc2011AC5024).
References
- 1.Kloosterman W. P., Plasterk R. H. The diverse functions of microRNAs in animal development and disease. Dev. Cell. (2006);11:441–450. doi: 10.1016/j.devcel.2006.09.009. [DOI] [PubMed] [Google Scholar]
- 2.Esquela-Kerscher A., Slack F. J. Oncomirs - microRNAs with a role in cancer. Nat. Rev. Cancer. (2006);6:259–269. doi: 10.1038/nrc1840. [DOI] [PubMed] [Google Scholar]
- 3.Lewis B. P., Burge C. B., Bartel D. P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. (2005);120:15–20. doi: 10.1016/j.cell.2004.12.035. [DOI] [PubMed] [Google Scholar]
- 4.Wicha M. S. Stemming a tumor with a little miR. Nat. Med. (2011);17:162–164. doi: 10.1038/nm0211-162. [DOI] [PubMed] [Google Scholar]
- 5.Calin G. A., Sevignani C., Dumitru C. D., Hyslop T., Noch E., Yendamuri S., Shimizu M., Rattan S., Bullrich F., Negrini M., Croce C. M. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. U. S. A. (2004);101:2999–3004. doi: 10.1073/pnas.0307323101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhu S., Wu H., Wu F., Nie D., Sheng S., Mo Y. Y. MicroRNA-21 targets tumor suppressor genes in invasion and metastasis. Cell Res. (2008);18:350–359. doi: 10.1038/cr.2008.24. [DOI] [PubMed] [Google Scholar]
- 7.Hiyoshi Y., Kamohara H., Karashima R., Sato N., Imamura Y., Nagai Y., Yoshida N., Toyama E., Hayashi N., Watanabe M., Baba H. MicroRNA-21 regulates the proliferation and invasion in esophageal squamous cell carcinoma. Clin. Cancer Res. (2009);15:1915–1922. doi: 10.1158/1078-0432.CCR-08-2545. [DOI] [PubMed] [Google Scholar]
- 8.Huang L., Luo J., Cai Q., Pan Q., Zeng H., Guo Z., Dong W., Huang J., Lin T. MicroRNA-125b suppresses the development of bladder cancer by targeting E2F3. Int. J. Cancer. (2011);128:1758–1769. doi: 10.1002/ijc.25509. [DOI] [PubMed] [Google Scholar]
- 9.Lo W. L., Yu C. C., Chiou G. Y., Chen Y. W., Huang P. I., Chien C. S., Tseng L. M., Chu P. Y., Lu K. H., Chang K. W., Kao S. Y., Chiou S. H. MicroRNA-200c attenuates tumour growth and metastasis of presumptive head and neck squamous cell carcinoma stem cells. J. Pathol. (2011);223:482–495. doi: 10.1002/path.2826. [DOI] [PubMed] [Google Scholar]
- 10.Li N., Fu H., Tie Y., Hu Z., Kong W., Wu Y., Zheng X. miR-34a inhibits migration and invasion by down-regulation of c-Met expression in human hepatocellular carcinoma cells. Cancer Lett. (2009);275:44–53. doi: 10.1016/j.canlet.2008.09.035. [DOI] [PubMed] [Google Scholar]
- 11.Ward B. P., Tsongalis G. J., Kaur P. MicroRNAs in chronic lymphocytic leukemia. Exp. Mol. Pathol. (2011);90:173–178. doi: 10.1016/j.yexmp.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 12.Calin G. A., Sevignani C., Dumitru C. D., Hyslop T., Noch E., Yendamuri S., Shimizu M., Rattan S., Bullrich F., Negrini M., Croce C. M. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. U. S. A. (2004);101:2999–3004. doi: 10.1073/pnas.0307323101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Desjobert C., Renalier M. H., Bergalet J., Dejean E., Joseph N., Kruczynski A., Soulier J., Espinos E., Meggetto F., Cavaille J., Delsol G., Lamant L. MiR-29a downregulation in ALK-positive anaplastic large-cell lymphomas contributes to apoptosis blockade through MCL-1 overexpression. Blood. (2011);117:6627–6637. doi: 10.1182/blood-2010-09-301994. [DOI] [PubMed] [Google Scholar]
- 14.Grimson A., Farh K. K., Johnston W. K., Garrett-Engele P., Lim L. P., Bartel D. P. MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol. Cell. (2007);27:91–105. doi: 10.1016/j.molcel.2007.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Maragkakis M., Alexiou P., Papadopoulos G. L., Reczko M., Dalamagas T., Giannopoulos G., Goumas G., Koukis E., Kourtis K., Simossis V. A., Sethupathy P., Vergoulis T., Koziris N., Sellis T., Tsanakas P., Hatzigeorgiou A. G. Accurate microRNA target prediction correlates with protein repression levels. BMC Bioinformatics. (2009);10:295. doi: 10.1186/1471-2105-10-295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Maragkakis M., Reczko M., Simossis V. A., Alexiou P., Papadopoulos G. L., Dalamagas T., Giannopoulos G., Goumas G., Koukis E., Kourtis K., Vergoulis T., Koziris N., Sellis T., Tsanakas P., Hatzigeorgiou A. G. DIANA-microT web server: elucidating microRNA functions through target prediction. Nucleic Acids Res. (2009);37:W273–276. doi: 10.1093/nar/gkp292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rehmsmeier M., Steffen P., Hochsmann M., Giegerich R. Fast and effective prediction of microRNA/target duplexes. RNA. (2004);10:1507–1517. doi: 10.1261/rna.5248604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hirakawa S., Kodama S., Kunstfeld R., Kajiya K., Brown L. F., Detmar M. VEGF-A induces tumor and sentinel lymph node lymphangiogenesis and promotes lymphatic metastasis. J. Exp. Med. (2005);201:1089–1099. doi: 10.1084/jem.20041896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nicoloso M. S., Spizzo R., Shimizu M., Rossi S., Calin G. A. MicroRNAs--the micro steering wheel of tumour metastases. Nat. Rev. Cancer. (2009);9:293–302. doi: 10.1038/nrc2619. [DOI] [PubMed] [Google Scholar]
- 20.Pandey A. K., Verma G., Vig S., Srivastava S., Srivastava A. K., Datta M. miR-29a levels are elevated in the db/db mice liver and its overexpression leads to attenuation of insulin action on PEPCK gene expression in HepG2 cells. Mol. Cell. Endocrinol. (2011);332:125–133. doi: 10.1016/j.mce.2010.10.004. [DOI] [PubMed] [Google Scholar]
- 21.Du B., Ma L. M., Huang M. B., Zhou H., Huang H. L., Shao P., Chen Y. Q., Qu L. H. High glucose down-regulates miR-29a to increase collagen IV production in HK-2 cells. FEBS Lett. (2010);584:811–816. doi: 10.1016/j.febslet.2009.12.053. [DOI] [PubMed] [Google Scholar]
- 22.Kawashita Y., Jinnin M., Makino T., Kajihara I., Makino K., Honda N., Masuguchi S., Fukushima S., Inoue Y., Ihn H. Circulating miR-29a levels in patients with scleroderma spectrum disorder. J. Dermatol. Sci. (2011);61:67–69. doi: 10.1016/j.jdermsci.2010.11.007. [DOI] [PubMed] [Google Scholar]
- 23.Hebert S. S., Horre K., Nicolai L., Papadopoulou A. S., Mandemakers W., Silahtaroglu A. N., Kauppinen S., Delacourte A., De Strooper B. Loss of microRNA cluster miR-29a/b-1 in sporadic Alzheimer's disease correlates with increased BACE1/beta-secretase expression. Proc. Natl. Acad. Sci. U. S .A. (2008);105:6415–6420. doi: 10.1073/pnas.0710263105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shioya M., Obayashi S., Tabunoki H., Arima K., Saito Y., Ishida T., Satoh J. Aberrant microRNA expression in the brains of neurodegenerative diseases: miR-29a decreased in Alzheimer disease brains targets neurone navigator 3. Neuropathol. Appl. Neurobiol. (2010);36:320–330. doi: 10.1111/j.1365-2990.2010.01076.x. [DOI] [PubMed] [Google Scholar]
- 25.Ahluwalia J. K., Khan S. Z., Soni K., Rawat P., Gupta A., Hariharan M., Scaria V., Lalwani M., Pillai B., Mitra D., Brahmachari S. K. Human cellular microRNA hsa-miR-29a interferes with viral nef protein expression and HIV-1 replication. Retrovirology. (2008);5:117. doi: 10.1186/1742-4690-5-117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bandyopadhyay S., Friedman R. C., Marquez R. T., Keck K., Kong B., Icardi M. S., Brown K. E., Burge C. B., Schmidt W. N., Wang Y., McCaffrey A. P. Hepatitis C virus infection and hepatic stellate cell activation downregulate miR-29: miR-29 overexpression reduces hepatitis C viral abundance in culture. J. Infect. Dis. (2011);203:1753–1762. doi: 10.1093/infdis/jir186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li J., Zhang Y., Kuruba R., Gao X., Gandhi C. R., Xie W., Li S. Roles of miR-29a in the antifibrotic effect of FXR in hepatic stellate cells. Mol. Pharmacol. (2011);89:191–200. doi: 10.1124/mol.110.068247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mulkeen A. L., Silva T., Yoo P. S., Schmitz J. C., Uchio E., Chu E., Cha C. Short interfering RNA-mediated gene silencing of vascular endothelial growth factor: effects on cellular proliferation in colon cancer cells. Arch. Surg. (2006);141:367–374. doi: 10.1001/archsurg.141.4.367. [DOI] [PubMed] [Google Scholar]
- 29.Petit A. M., Rak J., Hung M. C., Rockwell P., Goldstein N., Fendly B., Kerbel R. S. Neutralizing antibodies against epidermal growth factor and ErbB-2/neu receptor tyrosine kinases down-regulate vascular endothelial growth factor production by tumor cells in vitro and in vivo: angiogenic implications for signal transduction therapy of solid tumors. Am. J. Pathol. (1997);151:1523–1530. [PMC free article] [PubMed] [Google Scholar]
- 30.Des Guetz G., Uzzan B., Nicolas P., Cucherat M., Morere J. F., Benamouzig R., Breau J. L., Perret G. Y. Microvessel density and VEGF expression are prognostic factors in colorectal cancer. Meta-analysis of the literature. Br. J. Cancer. (2006);94:1823–1832. doi: 10.1038/sj.bjc.6603176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kolev Y., Uetake H., Iida S., Ishikawa T., Kawano T., Sugihara K. Prognostic significance of VEGF expression in correlation with COX-2, microvessel density, and clinicopathological characteristics in human gastric carcinoma. Ann. Surg. Oncol. (2007);14:2738–2747. doi: 10.1245/s10434-007-9484-7. [DOI] [PubMed] [Google Scholar]
- 32.Ugalde A. P., Ramsay A. J., de la Rosa J., Varela I., Marino G., Cadinanos J., Lu J., Freije J. M., Lopez-Otin C. Aging and chronic DNA damage response activate a regulatory pathway involving miR-29 and p53. EMBO J. (2011);30:2219–2232. doi: 10.1038/emboj.2011.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cui Y., Su W. Y., Xing J., Wang Y. C., Wang P., Chen X. Y., Shen Z. Y., Cao H., Lu Y. Y., Fang J. Y. MiR-29a inhibits cell proliferation and induces cell cycle arrest through the downregulation of p42.3 in human gastric cancer. PLoS One. (2011);6:e25872. doi: 10.1371/journal.pone.0025872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ghosh R., Lipson K. L., Sargent K. E., Mercurio A. M., Hunt J. S., Ron D., Urano F. Transcriptional regulation of VEGF-A by the unfolded protein response pathway. PLoS One. (2010);5:e9575. doi: 10.1371/journal.pone.0009575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Livak K. J., Schmittgen T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. (2001);25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 36.Weidner N., Folkman J., Pozza F., Bevilacqua P., Allred E. N., Moore D. H., Meli S., Gasparini G. Tumor angiogenesis: a new significant and independent prognostic indicator in early-stage breast carcinoma. J. Natl. Cancer Inst. (1992);84:1875–1887. doi: 10.1093/jnci/84.24.1875. [DOI] [PubMed] [Google Scholar]
- 37.Tie J., Pan Y., Zhao L., Wu K., Liu J., Sun S., Guo X., Wang B., Gang Y., Zhang Y., Li Q., Qiao T., Zhao Q., Nie Y., Fan D. MiR-218 inhibits invasion and metastasis of gastric cancer by targeting the Robo1 receptor. PLoS Genet. (2010);6:e1000879. doi: 10.1371/journal.pgen.1000879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhu S., Si M. L., Wu H., Mo Y. Y. MicroRNA-21 targets the tumor suppressor gene tropomyosin 1 (TPM1). J. Biol. Chem. (2007);282:14328–14336. doi: 10.1074/jbc.M611393200. [DOI] [PubMed] [Google Scholar]