

Abstract
CD1 molecules belong to non-polymorphic MHC class I-like proteins and present lipid antigens to T cells. Five different CD1 genes (CD1a-e) have been identified and classified into two groups. Group 1 include CD1a-c and present pathogenic lipid antigens to αβ T cells reminiscence of peptide antigen presentation by MHC-I molecules. CD1d is the only member of Group 2 and presents foreign and self lipid antigens to a specialized subset of αβ T cells, NKT cells. NKT cells are involved in diverse immune responses through prompt and massive production of cytokines. CD1d-dependent NKT cells are categorized upon the usage of their T cell receptors. A major subtype of NKT cells (type I) is invariant NKT cells which utilize invariant Vα14-Jα18 TCR alpha chain in mouse. The remaining NKT cells (type II) utilize diverse TCR alpha chains. Engineered CD1d molecules with modified intracellular trafficking produce either type I or type II NKT cell-defects suggesting the lipid antigens for each subtypes of NKT cells are processed/generated in different intracellular compartments. Since the usage of TCR by a T cell is the result of antigen-driven selection, the intracellular metabolic pathways of lipid antigen are a key in forming the functional NKT cell repertoire. [BMB Reports 2014; 47(5): 241-248]
Keywords: Antigen presentation, CD1d, Endosome, Endolysosomal compartment, Glycolipid, Lysosome, NKT cells
INTRODUCTION
Major histocompatibility complex class I (MHC-I) and MHC-II molecules present peptide antigens to CD8+ T cells and CD4+ T cells, respectively. Pathogen derived protein antigens synthesized in the intracellular compartment are loaded as a short peptide on the MHC-I molecules in the endoplasmic reticulum (ER) before MHC-I presents the peptide antigens to CD8+ αβ T cells (1). Extracellular-derived pathogenic peptides are loaded on MHC-II molecules in the MHC-II compartment (MIIC) such as lysosomes before they are presented to CD4+ αβ T cells (2).
Mammalian immune systems can recognize not only protein antigens but also lipid antigens presented by CD1 molecules. CD1 molecules are MHC-I-like proteins that are non-covalently associated with β2 microglobulin (β2m) (3). Based on sequence analysis, CD1 proteins can be classified into two groups in human. Group 1 compromises CD1a, CD1b, CD1c, and CD1e, group 2 CD1d (4). CD1e not expressed on the cell surface also can be independently classified as group 3 (5). Each CD1 molecule presents specific lipid antigens derived from foreign microorganism and/or self lipid antigens. For example, CD1a presents didehydroxymycobactin of mycobacteria (6). CD1b presents mycolic acids, phosphatidylinositol mannosides of mycobacteria and self antigen monosialotetrahexosylganglioside (GM1), the prototype ganglioside (7-9). CD1c presents mannosyl-β1-phosphomycoketides of mycobacteria (10,11). Lastly, CD1d present marine sponge-derived α-galactosylceramide (α-GalCer) and self lipid antigen, disialoganglioside GD3 (12,13).
CD1a-c proteins are recognized by diverse conventional αβ T cells. By contrast, CD1d proteins are recognized by a specialized subset of αβ T cells, NKT cells (14). Unlike human, mouse CD1 proteins only contain group 2 (CD1d) (5). Due to the lack of group 1 CD1 genes in mouse, the biology of CD1 molecules including antigen presentation mechanism is less well studied in group 1 CD1 molecules. Here, we review recent progresses on lipid antigen presentation by CD1d.
CD1D STRUCTURE
CD1d is highly expressed in antigen presenting cells (APCs) such as dendritic cells (DCs), macrophages and B cells. APCs engulf exogenous pathogens such as mycobacterium and CD1d encounters glycolipid antigens of engulfed pathogens at endocytic vesicles such as late endosome and lysosome and loads glycolipid antigens through lysosomal machinery (15, 16).
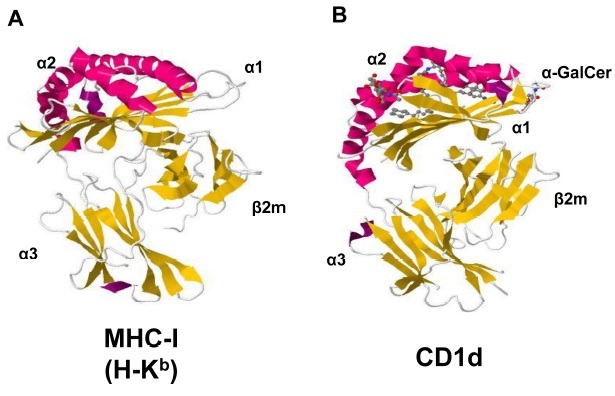
The overall structure of CD1 molecules (isoform a-e) is similar to MHC-I molecules. As the case of MHC-I, CD1 molecules consist of α1, α2 and α3 domains. Each α1 and α2 domain contains an α-helical structure positioned antiparallelly each on top of a β-pleated sheet and form an antigen binding region. The α3 domain connects antigen binding region to the transmembrane segment which is followed by a short cytoplasmic tail region. The α3 domain is also associated noncovalently with β2m (Fig. 1) (17). Although the overall structures of CD1 molecules are quite similar to that of MHC-I molecules, the antigen binding grooves of two molecules shows substantial differences. While MHC-I molecules have relatively shallow and long antigen binding groove along with horizontal axis, CD1 molecules have more deeper and voluminous antigen binding groove (17). Although the antigen binding grooves of CD1 isoforms have similar hydrophobic nature, their fine architectures are slightly different in accommodating distinct glycolipid antigens (17). The antigen binding groove of CD1d consists of two pockets: the A’ pocket and the F’ pocket with a narrow entrance at the junction between these two pockets. The deep and hydrophobic pockets of CD1d can accommodate alkyl chains of glycolipid antigens and the hydrophilic sugar in the head group protrudes toward approaching TCRs of NKT cells (17). The crystal structure of the TCR-α-GalCer-CD1d ternary complex shows the contact of the head group of α-GalCer and the CD1 α-helices with the TCR of NKT cells as predicted (18). While TCRs of conventional T cells in most of the structure so far solved, approach diagonally or orthogonally to the central top of peptide-MHC plane and produce substantial area of footprint, the TCRs of NKT cells approach parallel to the long axis of the antigen binding groove of CD1d and towards one end of CD1d molecule (18).
Fig. 1. The structures of MHC class I and CD1d. The side views of MHC-I (A) and CD1d (B) molecules. α1 and α2 domains consisting two alpha helixes on top of the molecules (magenta) and a beta-plated sheet (yellow) beneath the alpha helixes form an antigen binding region, which is facing toward T cell receptors. β2m-associated α3 domain is attached on the cell surface. PDB accession numbers are 2VAA (MHC I) and 1ZT4 (CD1d).
NKT CELLS
CD1d-restricted T cells express T cell markers along with canonical NK cell markers on the cell surface, thus they are called as NKT cells. In mouse, NKT cells represent ∼1% of the T cell population in most lymphatic tissues including peripheral blood, spleen, thymus and lymph nodes but NKT cells represent up to 30% of T cells in the liver (15,16). The expression of LFA-1 and CXCR6 on NKT cells seems important for the striking levels of accumulation of NKT cells in the liver (19-21).
NKT cells can produce a large quantity of cytokines promptly upon recognition of glycolipid antigens presented by CD1d. The major cytokines, including IFN-γ, IL-4, IL-10, IL-13, IL-21, GM-CSF and IL-17 secreted by NKT cells have been identified so far (16). Cytokines secreted by NKT cells can transactivate other lymphocytes such as DCs, NK cells, T cells and B cells. Thus NKT cells can bridge innate immunity and adaptive immunity (16).
CD1d-restricted NKT cells consist of type I and type II NKT cells depending on the usage of TCR α chains. Type I NKT cells utilize an invariant Vα14-Jα18 TCR alpha chain in mouse (Vα24-Jα18 in human) coupled mostly with Vβ2, 7 and 8 (Vβ11 in human), hence they are called invariant NKT (iNKT) cells (16). While type I NKT cells occupy about 80% of NKT cells, the remaining 20% is type II NKT cells which utilize diverse TCR Vα and TCR Vβ chains. Both type I and type II NKT cells show autoreactivity against CD1d expressing cells, especially thymocytes (16). Although both types of NKT cells show autoreactivity, their antigen specificities are non-overlapping due to their different TCR usage. Type I NKT cells but not type II NKT cells recognize a well known prototype NKT cell antigen, α-GalCer and a self antigen isoglobotrihexosylceramide (iGb3) (15). Instead, type II NKT cells recognize sulfatide which is not recognized by type I NKT cells (22). Both type I and type II NKT cells have been known to cross-regulate with each other (23-25).
CD1D ANTIGEN
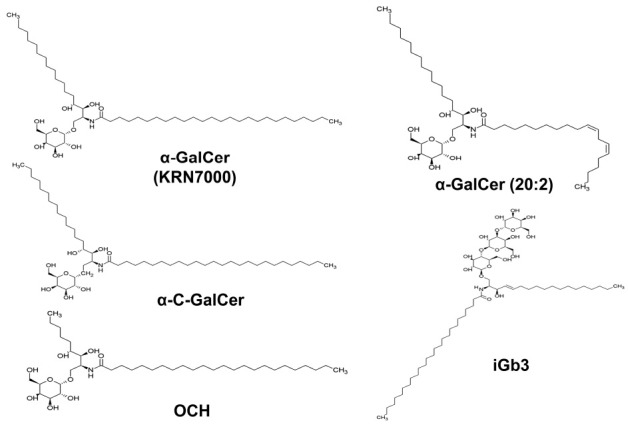
α-GalCer, a typical glycolipid antigen of type I NKT cells was derived from a marine sponge Agelas mauritanius in an anticancer drug screening (13). The galactose residue is connected to sphingosine base through α-annomeric glycosidic linkage. Various derivatives of α-GalCer have been synthesized through the modification of acyl carbon chain and some of those derivatives induced either Th1 or Th2-biased responses of type I NKT cells. For example, α-C-galactosylceramide (α-C-GalCer) induces Th1-biased responses from type I NKT cells (26-29), while OCH and α-GalCer(C20:2) produce Th2-biased responses from type I NKT cells (30,31) (Fig. 2). Although α-GalCer induces strong responses of type I NKT cells and has been a reference antigen for NKT cell research, it may not be considered as the natural antigen for NKT cells as it is not found in the body of mammals. In addition to the synthetic glycolipids, antigens derived from microbes have been shown to be presented by CD1d. Phosphatidylinositol mannosides derived from Mycobacterium tuberculosis, α-Galactosyldiacylgycerol derived from Borrelia burgdorferi, lipophosphoglycan derived from Leishmania donovani and α-glucuronosylceramide derived from Sphingomonas spp. are presented by CD1d (32-35). An enormous effort has been invested to date in trying to find self antigens of NKT cells as NKT cells clearly showed autoreactivity against syngenic cells, especially thymocytes without the addition of exogenous antigens (36-39). iGb3 was the first identified self glycolipid antigen and was considered to be involved in the development of NKT cells (40). The frequency of NKT cells in the lysosomal glycosphingolipid degrading enzyme β-hexosaminidase b subunit knock-out (Hexb-/-) mice, deficient of β-hexosaminidases involved in the biosynthesis of iGb3, was dramatically reduced in the thymus and spleen (40). However, iGb3 synthase-deficient mice still maintained normal development of NKT cells (41). Regardless of such controversy, it is clear that iGb3 can be loaded on CD1d of antigen presenting cells and can stimulate type I NKT cells in vitro (33,40).
Fig. 2. The chemical structures of CD1dspecific glycolipid antigens. The structures of α-galactosylceramide (α-GalCer), α-C-galactosylceramide (α-C-GalCer), an α-GalCer with truncated sphigosine chain (OCH), an α-GalCer with unsaturated carbons in acyl chain (α-GalCer (C20:2)), and isoglobotrihexosylceramide (iGb3).
Other self glycolipid antigens for NKT cells such as phosphatidylinositol, phosphatidylgycerol, and phosphatidylehanolamine have been eluted from endoplasmic reticulum (ER) using recombinant CD1d (42). Ganglioside GD3, a major antigen for autoantibodies was isolated from human melanoma. Human melanoma cell or GD3-pulsed dendritic cells stimulated NKT cells in a CD1d-dependent manner (12). According to a recent research, blocking of a biosynthetic pathway for peroxisome-derived self glycolipids affected the normal development of NKT cells (43). An analogue of peroxisome-derived self glycolipid, synthetic ether-bonded plasmalogen C16-lysophosphatidylethanolamine (pLPE) has been shown to be presented by CD1d and stimulate type I NKT cells (43). Sulfatide, a component of myelin, is a glycosphingolipid that has sulfated β-linked galactose headgroup. Initially, sulfatide was known to be presented by CD1a, b, and c (44) but it has been evident that sufatide is presented by CD1d and stimulate type II NKT cells (22,45,46).
CD1D PRESENTATION PATHWAY
CD1d assembly
The biosynthesis and surface expression pathways of CD1d are mostly common with those of conventional MHC-I molecules. Newly synthesized CD1d molecule has signal sequence directing the molecule to the lumen of ER and thus, CD1d is immediately glycosylated in the ER (47). For proper folding, ER-directed nascent CD1d is retained in the ER by the lectin-like ER chaperone calnexin, which assists in protein folding and promotes assembly with β2m. Calreticulin, another lectin-like ER chaperone, replaces calnexin and make complex with a thiol oxidoreductase ERp57 (47-49). In case of conventional MHC-I molecules, calreticulin and ERp57 complexed with tapasin and are connected to transporters associated with antigen processing (TAP) 1and TAP2 to form peptide loading complex (PLC). In case of CD1d, however, an ER-resident lipid transfer protein (MTP) is involved in the presentation of lipid antigen to CD1d (50). Although the detailed action mechanism is not clear, MTP, involved in proper assembly and secretion of very low density lipoprotein (VLDL), probably mediate the loading of phospholipids to CD1d in the ER. Stabilized-CD1d/β2m/lipid complex moves to the cell surface through trans-Golgi network (TGN) (14).
CD1d endocytic pathway
CD1d exit to the cell surface through TGN after being loaded with self lipid antigen in the ER. The NKT cells which recognize ER-loaded self lipid antigens do not respond against α-GalCer although they have TCR Vα and Vβ usage similar with those of type I NKT cells (42). The presentation of self lipid antigens on CD1d seems to be mediated by MTP as APCs with defective MTP function has a defect in CD1d antigen presentation ability (50). Also the treatment of MTP inhibitor during fetal thymic organ culture (FTOC) almost completely reduced the number of developing NKT cells (51).
CD1d molecules which arrived on the cell surface are internalized into the early endosome by interacting with adaptor protein-2 (AP-2) and move to late endosome or lysosome by the mediation of AP-3 (52-54). There, the lipid antigen loaded in the ER is replaced by lysosomal-derived lipid antigens. Here, saposin is known to be involved in the replacement process (55-58). Lysosomal antigen-loaded CD1d molecules are recycled to the cell surface and present the antigen to type I NKT cells. Tyrosine motif on the cytoplamic tail of CD1d is important in the interaction with AP-2 and AP-3, which mediate the endocytic pathway and lysosomal trafficking of CD1d (52,53,59). APCs with cytoplamic tail-deleted CD1d (CD1d-TD) were reported to have defective antigen presentation to type I NKT cells (60-63). CD1d-TD showed a defect in the presentation of disaccharide glycolipid antigens and also a defect in optimal presentation of α-GalCer (63-65). CD1d knock-out mouse reconstituted with CD1d-TD also showed dramatically reduced number of type I NKT cells (62). However, the removal of cytoplasmic tail of CD1d did not affect the development of type II NKT cells (62). Type I NKT cell-specific defect in antigen presentation of CD1d-TD was considered to be a problem originated from the inability of its trafficking to the endolysosomal compartment where it replaces ER antigens with type NKT cell-specific lysosomal antigens.
CD1d molecules have several positive charged amino acids in between transmembrane region and short cytoplamic tail. Recently, it has been shown that when three consecutive arginines of this motif in murine CD1d were replaced with non-charged alanines (CD1d-RA), antigen presentation ability of CD1d-RA to type I NKT cells was severely reduced either for endogenous self lipid antigen and exogenous lipid antigens even though CD1d molecules were highly accumulated in the endolysosomal (lamp-1+) compartments. Moreover, CD1d-RA also had a defect in antigen presentation to type II NKT cells (66).
Strikingly however, the removal of both arginine motif and cytoplasmic tail of CD1d (CD1d-RATD) showed normal trafficking of CD1d molecules to the lamp-1+ endolysosomal compartment (Fig. 3). Since CD1d-TD obviously has a defect in recycling CD1d from cell surface to endosomal compartment, the finding of CD1d-RATD in the endolysomal compartment suggest the existence of a yet unidentified direct trafficking route of CD1d from ER to lysosomal compartment other than the internalization from cell surface. In that sense, this direct trafficking seems reminiscence of that of MHC-II, where it is a direct traffic from Golgi apparatus to endosomal compartment by the guidance of invariant chain (Ii). Indeed, similar trafficking was observed when MHC-II or Ii was overexpressed with CD1d-TD (60,67). However, CD1d-RATD traffic to lamp-1+ compartment regardless of the existence of MHC-II or Ii (66). Moreover, CD1d-TD trafficked to lamp-1+ compartment by overexpressed MHC-II or Ii showed recovered antigen presentation to type I NKT cells compared to CD1d-TD (60,67). However, the antigen presentation ability of CD1d-RATD to type I NKT cells was not recovered even though CD1d molecules were highly accumulated in lamp-1+ compartment (66).
Fig. 3. Model for the assembly, trafficking and antigen presentation pathway of CD1d. CD1d molecules assembled in the ER bind with ER glycolipid and traffic to the extracellular surface through TGN and then they are internalized into type I lysosomal compartments (LY-I) where they replace antigens with type I NKT cell-specific lysosomal antigens. Some CD1d molecules may traffic directly from ER/Golgi to a distinct lysosomal compartment (LY-II) where they are loaded with type II NKT cell-specific glycolipid antigens. The tri-argine motif may facilitate the exit of antigen-loaded CD1d molecules from lysosomal compartment to the extracellular surface for the interaction with NKT cells.
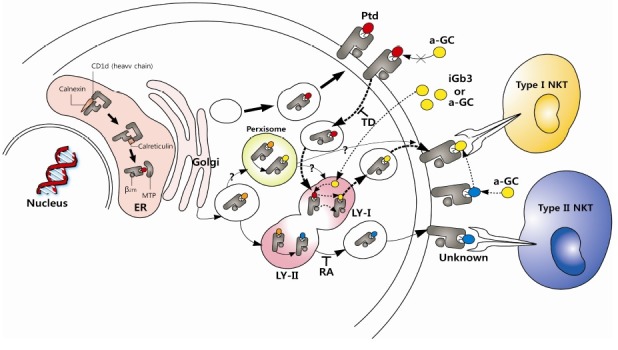
Although lamp-1+ compartment is considered as late endosome and lysosome, there is a possibility that the lamp-1+ compartment where CD1d internalized from cell surface (type I lysosome) and the lamp-1+ compartment where CD1d traffic directly from ER or TGN regardless (type II lysosome) are not within the same compartment. Interestingly, the presence of NKT cell antigens not only on the ER or lysosome but also in peroxsome has been reported (43). Although there is no direct evidence showing the trafficking of CD1d directly to peroxisome, it is possible that CD1d acquires lipid antigens directly or indirectly from other compartment not belong to ER or lysosome. Considering both, it is possible that CD1d in the type I lysosomal compartment recycled from the cell surface pick up glycolipid antigens for type I NKT cells while CD1d trafficked to type II lysosomal compartment directly from ER/Golgi apparatus pick up antigens for type II NKT cells. The arginine motif of CD1d probably then functions to modulate upward recycling of CD1d from lysosomal compartments to the cell surface. The finding of these additional trafficking routes of CD1d may open the basis to understand how CD1d can differentially present lipid antigens to the different types of NKT cells (Fig. 3).
CONCLUSION
Two decades since the discovery of NKT cells, NKT cells have drawn extensive interest because they were believed to involve in many immunological situations as immuno-regulatory cells. NKT cells can promptly produce a large amount of key cytokines such as IFN-γ and IL-4 upon recognition of glicolipid antigens. The cytokines produced by NKT cells can modulate various ongoing immune responses such as microbe infection, autoimmune diseases and cancer rejection. The involvement of NKT cells in these immunological conditions can produce either beneficial or detrimental effects depending on the involved subtypes of NKT cells. For example, there are evidences that the activation of NKT cells alleviate autoimmune disease diabetes while worsening another autoimmune disease, rheumatoid arthritis, (68-70). Although rich evidences support antitumor reactivity of type I NKT cells, type II NKT cells seem to suppress tumor-specific cytotoxic T cell responses (71). These conflicting activities of NKT cells are probably dependent on the lipid antigens they recognize at the site of inflammation as shown by the α-GalCer and a derivative of α-GalCer, OCH drive NKT cells into Th1 and Th2 cytokine production, respectively. By these reasons, the effect of NKT cell-mediated immunotherapies will be greatly affected by the type of different glycolipid antigens. Based on the facts that several different intracellular antigen-loading compartments for CD1d such as ER, TGN, endosome and lysosome and obvious differentiation of the antigen loading compartments for type I and type II NKT cells, the revealing of intracellular trafficking routes of CD1d and the antigen loading compartments for different antigens is essential for immunotherapies using NKT cells.
Acknowledgments
This research was supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF) (NRF-2012R1A2A4A01007183, S.-H.P.), (NRF-2013R1A1A20 64561, J.H.S.) and a Korea University Grants (S.-H.P.)
References
- 1.Rock K. L., York I. A., Saric T., Goldberg A. L. Protein degradation and the generation of MHC class I-presented peptides. Adv. Immunol. (2002);80:1–70. doi: 10.1016/s0065-2776(02)80012-8. [DOI] [PubMed] [Google Scholar]
- 2.Hiltbold E. M., Roche P. A. Trafficking of MHC class II molecules in the late secretory pathway. Curr. Opin. Immunol. (2002);14:30–35. doi: 10.1016/S0952-7915(01)00295-3. [DOI] [PubMed] [Google Scholar]
- 3.Porcelli S. A. The CD1 family: a third lineage of antigen-presenting molecules. Adv. Immunol. (1995);59:1–98. doi: 10.1016/s0065-2776(08)60629-x. [DOI] [PubMed] [Google Scholar]
- 4.Calabi F., Jarvis J. M., Martin L., Milstein C. Two classes of CD1 genes. Eur. J. Immunol. (1989);19:285–292. doi: 10.1002/eji.1830190211. [DOI] [PubMed] [Google Scholar]
- 5.de la Salle H., Mariotti S., Angenieux C., Gilleron M., Garcia-Alles L. F., Malm D., Berg T., Paoletti S., Maitre B., Mourey L., Salamero J., Cazenave J. P., Hanau D., Mori L., Puzo G., De Libero G. Assistance of microbial glycolipid antigen processing by CD1e. Science. (2005);310:1321–1324. doi: 10.1126/science.1115301. [DOI] [PubMed] [Google Scholar]
- 6.Moody D. B., Young D. C., Cheng T. Y., Rosat J. P., Roura-Mir C., O'Connor P. B., Zajonc D. M., Walz A., Miller M. J., Levery S. B., Wilson I. A., Costello C. E., Brenner M. B. T cell activation by lipopeptide antigens. Science. (2004);303:527–531. doi: 10.1126/science.1089353. [DOI] [PubMed] [Google Scholar]
- 7.Beckman E. M., Porcelli S. A., Morita C. T., Behar S. M., Furlong S. T., Brenner M. B. Recognition of a lipid antigen by CD1-restricted alpha beta+ T cells. Nature. (1994);372:691–694. doi: 10.1038/372691a0. [DOI] [PubMed] [Google Scholar]
- 8.Sieling P. A., Chatterjee D., Porcelli S. A., Prigozy T. I., Mazzaccaro R. J., Soriano T., Bloom B. R., Brenner M. B., Kronenberg M., Brennan P. J., Modlin R. L. CD1-restricted T cell recognition of microbial lipoglycan antigens. Science. (1995);269:227–230. doi: 10.1126/science.7542404. [DOI] [PubMed] [Google Scholar]
- 9.Shamshiev A., Donda A., Prigozy T. I., Mori L., Chigorno V., Benedict C. A., Kappos L., Sonnino S., Kronenberg M., De Libero G. The alphabeta T cell response to self-glycolipids shows a novel mechanism of CD1b loading and a requirement for complex oligosaccharides. Immunity. (2000);13:255–264. doi: 10.1016/S1074-7613(00)00025-X. [DOI] [PubMed] [Google Scholar]
- 10.Moody D. B., Ulrichs T., Muhlecker W., Young D. C., Gurcha S. S., Grant E., Rosat J. P., Brenner M. B., Costello C. E., Besra G. S., Porcelli S. A. CD1c-mediated T-cell recognition of isoprenoid glycolipids in Mycobacterium tuberculosis infection. Nature. (2000);404:884–888. doi: 10.1038/35009119. [DOI] [PubMed] [Google Scholar]
- 11.Matsunaga I., Bhatt A., Young D. C., Cheng T. Y., Eyles S. J., Besra G. S., Briken V., Porcelli S. A., Costello C. E., Jacobs W. R., Jr., Moody D. B. Mycobacterium tuberculosis pks12 produces a novel polyketide presented by CD1c to T cells. J. Exp. Med. (2004);200:1559–1569. doi: 10.1084/jem.20041429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wu D. Y., Segal N. H., Sidobre S., Kronenberg M., Chapman P. B. Cross-presentation of disialoganglioside GD3 to natural killer T cells. J. Exp. Med. (2003);198:173–181. doi: 10.1084/jem.20030446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kawano T., Cui J., Koezuka Y., Toura I., Kaneko Y., Motoki K., Ueno H., Nakagawa R., Sato H., Kondo E., Koseki H., Taniguchi M. CD1d-restricted and TCR-mediated activation of valpha14 NKT cells by glycosylceramides. Science. (1997);278:1626–1629. doi: 10.1126/science.278.5343.1626. [DOI] [PubMed] [Google Scholar]
- 14.Brigl M., Brenner M. B. CD1: antigen presentation and T cell function. Annu. Rev. Immunol. (2004);22:817–890. doi: 10.1146/annurev.immunol.22.012703.104608. [DOI] [PubMed] [Google Scholar]
- 15.Barral D. C., Brenner M. B. CD1 antigen presentation: how it works. Nat. Rev. Immunol. (2007);7:929–941. doi: 10.1038/nri2191. [DOI] [PubMed] [Google Scholar]
- 16.Bendelac A., Savage P. B., Teyton L. The biology of NKT cells. Annu. Rev. Immunol. (2007);25:297–336. doi: 10.1146/annurev.immunol.25.022106.141711. [DOI] [PubMed] [Google Scholar]
- 17.Zeng Z., Castano A. R., Segelke B. W., Stura E. A., Peterson P. A., Wilson I. A. Crystal structure of mouse CD1: An MHC-like fold with a large hydrophobic binding groove. Science. (1997);277:339–345. doi: 10.1126/science.277.5324.339. [DOI] [PubMed] [Google Scholar]
- 18.Borg N. A., Wun K. S., Kjer-Nielsen L., Wilce M. C., Pellicci D. G., Koh R., Besra G. S., Bharadwaj M., Godfrey D. I., McCluskey J., Rossjohn J. CD1d-lipid-antigen recognition by the semi-invariant NKT T-cell receptor. Nature. (2007);448:44–49. doi: 10.1038/nature05907. [DOI] [PubMed] [Google Scholar]
- 19.Geissmann F., Cameron T. O., Sidobre S., Manlongat N., Kronenberg M., Briskin M. J., Dustin M. L., Littman D. R. Intravascular immune surveillance by CXCR6+ NKT cells patrolling liver sinusoids. PLoS Biol. (2005);3:e113. doi: 10.1371/journal.pbio.0030113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Thomas S. Y., Scanlon S. T., Griewank K. G., Constantinides M. G., Savage A. K., Barr K. A., Meng F., Luster A. D., Bendelac A. PLZF induces an intravascular surveillance program mediated by long-lived LFA-1-ICAM-1 interactions. J. Exp. Med. (2011);208:1179–1188. doi: 10.1084/jem.20102630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ohteki T., Maki C., Koyasu S., Mak T. W., Ohashi P. S. Cutting edge: LFA-1 is required for liver NK1.1+TCR alpha beta+ cell development: evidence that liver NK1.1+TCR alpha beta+ cells originate from multiple pathways. J. Immunol. (1999);162:3753–3756. [PubMed] [Google Scholar]
- 22.Blomqvist M., Rhost S., Teneberg S., Lofbom L., Osterbye T., Brigl M., Mansson J. E., Cardell S. L. Multiple tissue-specific isoforms of sulfatide activate CD1d-restricted type II NKT cells. Eur. J. Immunol. (2009);39:1726–1735. doi: 10.1002/eji.200839001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang G., Nie H., Yang J., Ding X., Huang Y., Yu H., Li R., Yuan Z., Hu S. Sulfatide-activated type II NKT cells prevent allergic airway inflammation by inhibiting type I NKT cell function in a mouse model of asthma. Am. J. Physiol. Lung Cell Mol. Physiol. (2011);301:L975–984. doi: 10.1152/ajplung.00114.2011. [DOI] [PubMed] [Google Scholar]
- 24.Halder R. C., Aguilera C., Maricic I., Kumar V. Type II NKT cell-mediated anergy induction in type I NKT cells prevents inflammatory liver disease. J. Clin. Invest. (2007);117:2302–2312. doi: 10.1172/JCI31602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ambrosino E., Terabe M., Halder R. C., Peng J., Takaku S., Miyake S., Yamamura T., Kumar V., Berzofsky J. A. Cross-regulation between type I and type II NKT cells in regulating tumor immunity: a new immunoregulatory axis. J. Immunol. (2007);179:5126–5136. doi: 10.4049/jimmunol.179.8.5126. [DOI] [PubMed] [Google Scholar]
- 26.Lu X., Song L., Metelitsa L. S., Bittman R. Synthesis and evaluation of an alpha-C-galactosylceramide analogue that induces Th1-biased responses in human natural killer T cells. Chembiochem. (2006);7:1750–1756. doi: 10.1002/cbic.200600197. [DOI] [PubMed] [Google Scholar]
- 27.Wipf P., Pierce J. G. Expedient synthesis of the alpha-C-glycoside analogue of the immunostimulant galactosylceramide (KRN7000). Org. Lett. (2006);8:3375–3378. doi: 10.1021/ol0613057. [DOI] [PubMed] [Google Scholar]
- 28.Yang G., Schmieg J., Tsuji M., Franck R. W. The C-glycoside analogue of the immunostimulant alpha-galactosylceramide (KRN7000): synthesis and striking enhancement of activity. Angew. Chem. Int. Ed. Engl. (2004);43:3818–3822. doi: 10.1002/anie.200454215. [DOI] [PubMed] [Google Scholar]
- 29.Schmieg J., Yang G., Franck R. W., Tsuji M. Superior protection against malaria and melanoma metastases by a C-glycoside analogue of the natural killer T cell ligand alpha-Galactosylceramide. J. Exp. Med. (2003);198:1631–1641. doi: 10.1084/jem.20031192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Oki S., Tomi C., Yamamura T., Miyake S. Preferential T(h)2 polarization by OCH is supported by incompetent NKT cell induction of CD40L and following production of inflammatory cytokines by bystander cells in vivo. Int. Immunol. (2005);17:1619–1629. doi: 10.1093/intimm/dxh342. [DOI] [PubMed] [Google Scholar]
- 31.Yu K. O., Im J. S., Molano A., Dutronc Y., Illarionov P. A., Forestier C., Fujiwara N., Arias I., Miyake S., Yamamura T., Chang Y. T., Besra G. S., Porcelli S. A. Modulation of CD1d-restricted NKT cell responses by using N-acyl variants of alpha-galactosylceramides. Proc. Natl. Acad. Sci. U. S. A. (2005);102:3383–3388. doi: 10.1073/pnas.0407488102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kinjo Y., Wu D., Kim G., Xing G. W., Poles M. A., Ho D. D., Tsuji M., Kawahara K., Wong C. H., Kronenberg M. Recognition of bacterial glycosphingolipids by natural killer T cells. Nature. (2005);434:520–525. doi: 10.1038/nature03407. [DOI] [PubMed] [Google Scholar]
- 33.Mattner J., Debord K. L., Ismail N., Goff R. D., Cantu C.,, 3rd, Zhou D., Saint-Mezard P., Wang V., Gao Y., Yin N., Hoebe K., Schneewind O., Walker D., Beutler B., Teyton L., Savage P. B., Bendelac A. Exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature. (2005);434:525–529. doi: 10.1038/nature03408. [DOI] [PubMed] [Google Scholar]
- 34.Kinjo Y., Tupin E., Wu D., Fujio M., Garcia-Navarro R., Benhnia M. R., Zajonc D. M., Ben-Menachem G., Ainge G. D., Painter G. F., Khurana A., Hoebe K., Behar S. M., Beutler B., Wilson I. A., Tsuji M., Sellati T. J., Wong C. H., Kronenberg M. Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Nat. Immunol. (2006);7:978–986. doi: 10.1038/ni1380. [DOI] [PubMed] [Google Scholar]
- 35.Amprey J. L., Im J. S., Turco S. J., Murray H. W., Illarionov P. A., Besra G. S., Porcelli S. A., Spath G. F. A subset of liver NK T cells is activated during Leishmania donovani infection by CD1d-bound lipophosphoglycan. J. Exp. Med. (2004);200:895–904. doi: 10.1084/jem.20040704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bendelac A. Positive selection of mouse NK1+ T cells by CD1-expressing cortical thymocytes. J. Exp. Med. (1995);182:2091–2096. doi: 10.1084/jem.182.6.2091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bendelac A. CD1: presenting unusual antigens to unusual T lymphocytes. Science. (1995);269:185–186. doi: 10.1126/science.7542402. [DOI] [PubMed] [Google Scholar]
- 38.Exley M., Garcia J., Balk S. P., Porcelli S. Requirements for CD1d recognition by human invariant Valpha24+ CD4-CD8- T cells. J. Exp. Med. (1997);186:109–120. doi: 10.1084/jem.186.1.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bendelac A., Killeen N., Littman D. R., Schwartz R. H. A subset of CD4+ thymocytes selected by MHC class I molecules. Science. (1994);263:1774–1778. doi: 10.1126/science.7907820. [DOI] [PubMed] [Google Scholar]
- 40.Zhou D., Mattner J., Cantu C.,, 3rd, Schrantz N., Yin N., Gao Y., Sagiv Y., Hudspeth K., Wu Y. P., Yamashita T., Teneberg S., Wang D., Proia R. L., Levery S. B., Savage P. B., Teyton L., Bendelac A. Lysosomal glycosphingolipid recognition by NKT cells. Science. (2004);306:1786–1789. doi: 10.1126/science.1103440. [DOI] [PubMed] [Google Scholar]
- 41.Porubsky S., Speak A. O., Luckow B., Cerundolo V., Platt F. M., Grone H. J. Normal development and function of invariant natural killer T cells in mice with isoglobotrihexosylceramide (iGb3) deficiency. Proc. Natl. Acad. Sci. U. S. A. (2007);104:5977–5982. doi: 10.1073/pnas.0611139104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gumperz J. E., Roy C., Makowska A., Lum D., Sugita M., Podrebarac T., Koezuka Y., Porcelli S. A., Cardell S., Brenner M. B., Behar S. M. Murine CD1d-restricted T cell recognition of cellular lipids. Immunity. (2000);12:211–221. doi: 10.1016/S1074-7613(00)80174-0. [DOI] [PubMed] [Google Scholar]
- 43.Facciotti F., Ramanjaneyulu G. S., Lepore M., Sansano S., Cavallari M., Kistowska M., Forss-Petter S., Ni G., Colone A., Singhal A., Berger J., Xia C., Mori L., De Libero G. Peroxisome-derived lipids are self antigens that stimulate invariant natural killer T cells in the thymus. Nat. Immunol. (2012);13:474–480. doi: 10.1038/ni.2245. [DOI] [PubMed] [Google Scholar]
- 44.Shamshiev A., Gober H. J., Donda A., Mazorra Z., Mori L., De Libero G. Presentation of the same glycolipid by different CD1 molecules. J. Exp. Med. (2002);195:1013–1021. doi: 10.1084/jem.20011963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jahng A., Maricic I., Aguilera C., Cardell S., Halder R. C., Kumar V. Prevention of autoimmunity by targeting a distinct, noninvariant CD1d-reactive T cell population reactive to sulfatide. J. Exp. Med. (2004);199:947–957. doi: 10.1084/jem.20031389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Arrenberg P., Halder R., Dai Y., Maricic I., Kumar V. Oligoclonality and innate-like features in the TCR repertoire of type II NKT cells reactive to a beta- linked self-glycolipid. Proc. Natl. Acad. Sci. U. S. A. (2010);107:10984–10989. doi: 10.1073/pnas.1000576107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kang S. J., Cresswell P. Calnexin, calreticulin, and ERp57 cooperate in disulfide bond formation in human CD1d heavy chain. J. Biol. Chem. (2002);277:44838–44844. doi: 10.1074/jbc.M207831200. [DOI] [PubMed] [Google Scholar]
- 48.Hughes E. A., Cresswell P. The thiol oxidoreductase ERp57 is a component of the MHC class I peptide-loading complex. Current biology : CB. (1998);8:709–712. doi: 10.1016/S0960-9822(98)70278-7. [DOI] [PubMed] [Google Scholar]
- 49.Morrice N. A., Powis S. J. A role for the thiol-dependent reductase ERp57 in the assembly of MHC class I molecules. Current biology: CB. (1998);8:713–716. doi: 10.1016/S0960-9822(98)70279-9. [DOI] [PubMed] [Google Scholar]
- 50.Dougan S. K., Salas A., Rava P., Agyemang A., Kaser A., Morrison J., Khurana A., Kronenberg M., Johnson C., Exley M., Hussain M. M., Blumberg R. S. Microsomal triglyceride transfer protein lipidation and control of CD1d on antigen-presenting cells. J. Exp. Med. (2005);202:529–539. doi: 10.1084/jem.20050183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Brozovic S., Nagaishi T., Yoshida M., Betz S., Salas A., Chen D., Kaser A., Glickman J., Kuo T., Little A., Morrison J., Corazza N., Kim J. Y., Colgan S. P., Young S. G., Exley M., Blumberg R. S. CD1d function is regulated by microsomal triglyceride transfer protein. Nat. Med. (2004);10:535–539. doi: 10.1038/nm1043. [DOI] [PubMed] [Google Scholar]
- 52.Cernadas M., Sugita M., van der Wel N., Cao X., Gumperz J. E., Maltsev S., Besra G. S., Behar S. M., Peters P. J., Brenner M. B. Lysosomal localization of murine CD1d mediated by AP-3 is necessary for NK T cell development. J. Immunol. (2003);171:4149–4155. doi: 10.4049/jimmunol.171.8.4149. [DOI] [PubMed] [Google Scholar]
- 53.Elewaut D., Lawton A. P., Nagarajan N. A., Maverakis E., Khurana A., Honing S., Benedict C. A., Sercarz E., Bakke O., Kronenberg M., Prigozy T. I. The adaptor protein AP-3 is required for CD1d-mediated antigen presentation of glycosphingolipids and development of Valpha14i NKT cells. J. Exp. Med. (2003);198:1133–1146. doi: 10.1084/jem.20030143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lawton A. P., Prigozy T. I., Brossay L., Pei B., Khurana A., Martin D., Zhu T., Spate K., Ozga M., Honing S., Bakke O., Kronenberg M. The mouse CD1d cytoplasmic tail mediates CD1d trafficking and antigen presentation by adaptor protein 3-dependent and -independent mechanisms. J. Immunol. (2005);174:3179–3186. doi: 10.4049/jimmunol.174.6.3179. [DOI] [PubMed] [Google Scholar]
- 55.Salio M., Ghadbane H., Dushek O., Shepherd D., Cypen J., Gileadi U., Aichinger M. C., Napolitani G., Qi X., van der Merwe P. A., Wojno J., Veerapen N., Cox L. R., Besra G. S., Yuan W., Cresswell P., Cerundolo V. Saposins modulate human invariant Natural Killer T cells self-reactivity and facilitate lipid exchange with CD1d molecules during antigen presentation. Proc. Natl. Acad. Sci. U. S. A. (2013);110:E4753–4761. doi: 10.1073/pnas.1310050110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kang S. J., Cresswell P. Saposins facilitate CD1d-restricted presentation of an exogenous lipid antigen to T cells. Nat. Immunol. (2004);5:175–181. doi: 10.1038/ni1034. [DOI] [PubMed] [Google Scholar]
- 57.Yuan W., Qi X., Tsang P., Kang S. J., Illarionov P. A., Besra G. S., Gumperz J., Cresswell P. Saposin B is the dominant saposin that facilitates lipid binding to human CD1d molecules. Proc. Natl. Acad. Sci. U. S. A. (2007);104:5551–5556. doi: 10.1073/pnas.0700617104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhou D., Cantu C.,, 3rd, Sagiv Y., Schrantz N., Kulkarni A. B., Qi X., Mahuran D. J., Morales C. R., Grabowski G. A., Benlagha K., Savage P., Bendelac A., Teyton L. Editing of CD1d-bound lipid antigens by endosomal lipid transfer proteins. Science. (2004);303:523–527. doi: 10.1126/science.1092009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sugita M., Cao X., Watts G. F., Rogers R. A., Bonifacino J. S., Brenner M. B. Failure of trafficking and antigen presentation by CD1 in AP-3-deficient cells. Immunity. (2002);16:697–706. doi: 10.1016/S1074-7613(02)00311-4. [DOI] [PubMed] [Google Scholar]
- 60.Jayawardena-Wolf J., Benlagha K., Chiu Y. H., Mehr R., Bendelac A. CD1d endosomal trafficking is independently regulated by an intrinsic CD1d-encoded tyrosine motif and by the invariant chain. Immunity. (2001);15:897–908. doi: 10.1016/S1074-7613(01)00240-0. [DOI] [PubMed] [Google Scholar]
- 61.Chiu Y. H., Jayawardena J., Weiss A., Lee D., Park S. H., Dautry-Varsat A., Bendelac A. Distinct subsets of CD1d-restricted T cells recognize self-antigens loaded in different cellular compartments. J. Exp. Med. (1999);189:103–110. doi: 10.1084/jem.189.1.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Chiu Y. H., Park S. H., Benlagha K., Forestier C., Jayawardena-Wolf J., Savage P. B., Teyton L., Bendelac A. Multiple defects in antigen presentation and T cell development by mice expressing cytoplasmic tail-truncated CD1d. Nat. Immunol. (2002);3:55–60. doi: 10.1038/ni740. [DOI] [PubMed] [Google Scholar]
- 63.Prigozy T. I., Naidenko O., Qasba P., Elewaut D., Brossay L., Khurana A., Natori T., Koezuka Y., Kulkarni A., Kronenberg M. Glycolipid antigen processing for presentation by CD1d molecules. Science. (2001);291:664–667. doi: 10.1126/science.291.5504.664. [DOI] [PubMed] [Google Scholar]
- 64.Bai L., Sagiv Y., Liu Y., Freigang S., Yu K. O., Teyton L., Porcelli S. A., Savage P. B., Bendelac A. Lysosomal recycling terminates CD1d-mediated presentation of short and polyunsaturated variants of the NKT cell lipid antigen alphaGalCer. Proc. Natl. Acad. Sci. U. S. A. (2009);106:10254–10259. doi: 10.1073/pnas.0901228106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Im J. S., Arora P., Bricard G., Molano A., Venkataswamy M. M., Baine I., Jerud E. S., Goldberg M. F., Baena A., Yu K. O., Ndonye R. M., Howell A. R., Yuan W., Cresswell P., Chang Y. T., Illarionov P. A., Besra G. S., Porcelli S. A. Kinetics and cellular site of glycolipid loading control the outcome of natural killer T cell activation. Immunity. (2009);30:888–898. doi: 10.1016/j.immuni.2009.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Shin J. H., Park J. Y., Shin Y. H., Lee H., Park Y. K., Jung S., Park S. H. Mutation of a positively charged cytoplasmic motif within CD1d results in multiple defects in antigen presentation to NKT cells. J. Immunol. (2012);188:2235–2243. doi: 10.4049/jimmunol.1100236. [DOI] [PubMed] [Google Scholar]
- 67.Kang S. J., Cresswell P. Regulation of intracellular trafficking of human CD1d by association with MHC class II molecules. EMBO J. (2002);21:1650–1660. doi: 10.1093/emboj/21.7.1650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hong S., Wilson M. T., Serizawa I., Wu L., Singh N., Naidenko O. V., Miura T., Haba T., Scherer D. C., Wei J., Kronenberg M., Koezuka Y., Van Kaer L. The natural killer T-cell ligand alpha-galactosylceramide prevents autoimmune diabetes in non-obese diabetic mice. Nat. Med. (2001);7:1052–1056. doi: 10.1038/nm0901-1052. [DOI] [PubMed] [Google Scholar]
- 69.Kim H. Y., Kim H. J., Min H. S., Kim S., Park W. S., Park S. H., Chung D. H. NKT cells promote antibody-induced joint inflammation by suppressing transforming growth factor beta1 production. J. Exp. Med. (2005);201:41–47. doi: 10.1084/jem.20041400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Jung S., Shin H. S., Hong C., Lee H., Park Y. K., Shin J. H., Hong S., Lee G. R., Park S. H. Natural killer T cells promote collagen-induced arthritis in DBA/1 mice. Biochem. Biophys. Res. Commun. (2009);390:399–403. doi: 10.1016/j.bbrc.2009.09.008. [DOI] [PubMed] [Google Scholar]
- 71.Hong C., Park S. H. Application of natural killer T cells in antitumor immunotherapy. Crit. Rev. Immunol. (2007);27:511–525. doi: 10.1615/CritRevImmunol.v27.i6.20. [DOI] [PubMed] [Google Scholar]