

Abstract
Reactivating the p53 pathway in tumors is an important strategy for anticancer therapy. In response to diverse cellular stresses, the tumor suppressor p53 mediates apoptosis in a transcriptionindependent and transcription-dependent manner. Although extensive studies have focused on the transcription-dependent apoptotic pathway of p53, the transcription-independent apoptotic pathway of p53 has only recently been discovered. Molecular interactions between p53 and Bcl-2 family proteins in the mitochondria play an essential role in the transcriptionindependent apoptosis of p53. This review describes the structural basis for the transcription-independent apoptotic pathway of p53 and discusses its potential application to anticancer therapy. [BMB Reports 2014; 47(3): 167-172]
Keywords: Anticancer therapy, Apoptosis, Bcl-2 family proteins, p53, Structure
INTRODUCTION
p53 is a tumor suppressor protein that triggers cell cycle arrest, apoptosis, and senescence in response to diverse stresses such as DNA damage, hypoxia, ultraviolet light, and oncogene activation (1-4). In many human cancers, p53 function is abrogated by p53 gene mutation (5). However, approximately 50% of human cancers retain wild type p53, but impairment in p53 signaling or regulation inactivates its function and thus serves a critical role in cancer progression by protecting cancer cells from p53-dependent responses. Thus, restoring the function of inactivated p53 in tumors retaining wild type p53 is an important strategy for cancer treatment. Previously, several mouse model studies supported this therapeutic concept by showing that the reactivation of wild type p53 function induces cancer regression in vivo through apoptosis and/or senescence (6-8). Using this therapeutic strategy, pharmaceutical and academic research has identified many lead compounds for cancer drug discovery. Here, this review provides structural insights into targeting the transcription-independent p53 apoptotic pathway for anticancer therapy.
TRANSCRIPTION-DEPENDENT APOPTOTIC PATHWAY OF P53
Within the nucleus, p53 induces transcription-dependent apoptosis via the transactivation of target genes such as Noxa, Bax, and Puma (Fig. 1). Transcription-dependent apoptosis induced by p53 is mediated via interactions of the p53 transactivation domain (p53TAD) with the components of the basal transcriptional machinery. Under normal conditions, the p53 level is precisely controlled by mouse double minute 2 (MDM2). MDM2 is an E3 ubiquitin ligase that negatively regulates p53 stability through ubiquitination and inhibits the transactivation ability of p53 by interacting with the p53 transactivation domain (p53TAD). In many cancers, p53 function is impaired by overexpression of MDM2. Interaction of the p53TAD with transcriptional machinery components such as the transcriptional coactivator p300/CBP is neutralized by MDM2 in cancer cells. This neutralization by MDM2 could be inhibited by p53-mimetic compounds for cancer treatment. Thus, blocking the interaction of p53 with MDM2 is a promising anticancer strategy to reactivate the p53 pathway. The α-helical structure of the 15-residue p53TAD peptide fragment (residues 15-29) in complex with an N-terminal domain of MDM2 provides a useful template for the structure-based rational design of MDM2-inhibiting anticancer drugs (9) (Fig. 2A and B).
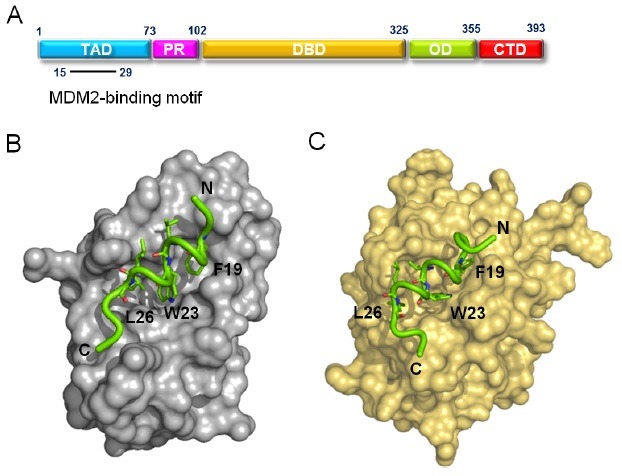
Fig. 1. Dual role of p53 in different cellular compartments to promote apoptosis. Interaction between p53 and the transcriptional coactivator CBP/p300 induces transcription-dependent apoptosis within the nucleus. This interaction can be neutralized by the interaction of p53 with MDM2. At the same time, binding of p53 to anti-apoptotic Bcl-2 and Bcl-XL can occur in the mitochondria, releasing proapoptotic Bak/Bax from complexes and triggering transcription-independent apoptosis.
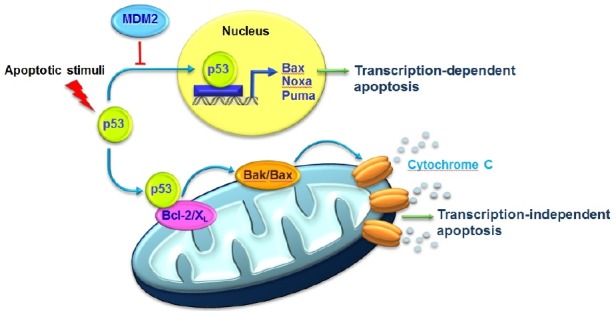
Fig. 2. Structural similarity between MDM2/p53TAD and Bcl-2/p53TAD complexes. (A) Domain organization of p53. p53 consists of a transactivation domain (TAD), proline-rich domain (PR), DNA-binding domain (DBD), oligomerization domain (OD), and C-terminal domain (CTD). The residues 15-29 of p53TAD are indicated as MDM2-binding motif. (B) Crystal structure of the MDM2/p53TAD peptide (residues 15-29) complex (PDB code: 1YCR) (9). (C) A refined structural model for the Bcl-2/p53TAD peptide (residues 15-29) complex generated from an NMR data-driven structure calculation (21). The p53TAD peptide is shown in green.
TRANSCRIPTION-INDEPENDENT APOPTOTIC PATHWAY OF P53
Recent studies have provided evidences supporting the pro-apoptotic effects of cytoplasmic p53 independent of its transcription ability (10-14). This non-transcriptional apoptosis of p53 is mainly achieved by the interaction between p53 and anti-apoptotic as well as pro-apoptotic B-cell lymphoma 2 (Bcl-2) family proteins. In an immediate response to apoptotic stresses, the cytoplasmic p53 moves rapidly to the mitochondria (14). In the mitochondria, p53 binds to anti-apoptotic Bcl-2 family proteins Bcl-XL and Bcl-2 and, releasing the pro-apoptotic effectors Bak/Bax from the complex with the anti-apoptotic proteins. Subsequently, the released Bak and Bax induce the lipid pore formation in the outer mitochondrial membrane, which elicits cytochrome c release (15). In addition, p53 can directly activate Bak and/or Bax through a “hit and run” mechanism to trigger the permeabilization of outer mitochondrial membrane (11).
A DUAL ROLE OF P53 IN APOPTOSIS PATHWAY
Based on results from recent studies, it is proposed that, under pro-apoptotic conditions, p53 can play a dual role in apoptosis in different cellular compartments (Fig. 1). Within the nucleus, p53 acts as a transcriptional activator and induces target gene expression through its interaction with the basic transcriptional machinery components, such as transcriptional coactivator CBP/p300, eventually leading to transcription-dependent apoptosis. This transcription-dependent interaction of p53 can be inhibited by binding with MDM2. In the mitochondria, p53 interacts with Bcl-2 and Bcl-XL, thereby triggering transcription-independent apoptosis. It should be noted that these binding events with different partners, occurring in distinct cellular compartments, are governed by the same binding motif of p53TAD (residues 15-29).
Previous mutational studies have shown that Leu22/Trp23 and Trp53/Phe54 mutations in the p53TAD render p53 completely incapable of mediating transcription-dependent apoptosis (16) because they are involved in interactions with the transcription machinery components such as TATA box-binding protein (TBP) (17), CREB-binding protein (CBP)/p300 (18), and hTAFII31 (19). This suggests that the p53TAD is crucial for transcription-dependent apoptosis induced by p53. Mutational and structural researches showed that the p53TAD also serves a critical role in the transcription-independent interaction of p53 with the anti-apoptotic Bcl-2 family proteins in the mitochondria (20,21). Thus, the p53TAD may be important in both the transcription-dependent and transcription-independent activation of apoptotic pathways by p53. Furthermore, a noticeable similarity was identified in the binding mode and binding site of the p53TAD with CBP/p300, MDM2, and Bcl-2/Bcl-XL (20). This finding revealed a highly conserved molecular mechanism underlying the interaction of the p53TAD with CBP/p300 and MDM2 in transcription-dependent p53 apoptotic pathway, and Bcl-XL and Bcl-2 and in transcription-independent p53 apoptosis pathway.
BCL-2 PROTEIN FAMILY
The Bcl-2 family proteins regulate mitochondrial apoptosis by modulating outer mitochondrial membrane permeability and cytochrome c release (11,13,14,22-26). According to the structure of Bcl-2 homology (BH) domains, the Bcl-2 protein family is classified into anti-apoptotic and pro-apoptotic subfamilies. Pro-apoptotic Bcl-2 family proteins have three BH domains called BH1, BH2, and BH3 (e.g. Bax and Bak) or a singular BH3 domain (BH3-only proteins), while anti-apoptotic Bcl-2 family proteins (e.g. Bcl-XL, Bcl-2, Mcl-1, A1, and Bcl-w,) have four BH domains called BH1, BH2, BH3, and BH4. The Bcl-2 protein family maintains the balance between cell survival and death via sophisticated interactions between pro-apoptotic and anti-apoptotic subfamily members (26). The pro-apoptotic Bcl-2 family effectors, Bax and Bak, undergo oligomerization to form lipid pores, which induces the outer mitochondrial membrane permeabilization and cytochrome c release. Antiapoptotic Bcl-2 family proteins perform their pro-survival function by binding and sequestering the pro-apoptotic effector proteins Bax and Bak and inhibiting membrane pore formation.
STRUCTURAL BASIS FOR THE INTERACTION BETWEEN P53 AND BCL-2 FAMILY PROTEINS
Interactions of p53 with Bcl-XL and Bcl-2 have been observed within the cell (14). Previously, the p53 DNA-binding domain (p53DBD) was suggested to mediate the interactions with Bcl-XL and Bcl-2 (15,28-30). Recently, it was shown that the binding of the p53DBD induces an allosteric conformational change in the BH3 peptide-binding region of Bcl-XL to facilitate binding with pro-apoptotic BH3-only proteins (29,31). However, previous cell biology studies showed that the p53 N-terminal domain (p53NTD), encompassing the p53TAD and proline-rich (PR) domain, is sufficiently able to trigger mitochondrial apoptosis in a transcription-independent manner, while the p53DBD is dispensable (32). In addition, the caspasecleaved p53 N-terminal fragment corresponding to residues 1-186, which includes the p53NTD and the truncated p53DBD, triggered transcription-independent apoptosis (33). These accumulated evidences indicated that the p53NTD alone, without the p53DBD, can mediate transcription- independent apoptosis by binding to anti-apoptotic Bcl-2 family proteins.
Recently, the biochemical and structural studies showed that the MDM2-binding motif (ΦXXΦΦ sequence motif, Φ; bulky hydrophobic residue, X; any amino acid) in the p53TAD binds to the BH3 peptide-binding groove in diverse anti-apoptotic Bcl-2 family proteins with a similar mode of binding as to MDM2 (20,21,34-37). These results revealed that the MDM2-binding peptide motif in the p53TAD can act as an universal site for binding diverse anti-apoptotic Bcl-2 family proteins. This is in conflict with the previous observation that anti-apoptotic Bcl-2 family proteins have unique specificity for pro-apoptotic Bcl-2 family proteins. For instance, Bcl-2, Bcl-XL, and Bcl-w bind to Bax whereas Mcl-1 and A1 bind to Noxa. However, the p53TAD was shown to bind to various anti-apoptotic members of Bcl-2 protein family such as Bcl-2, Bcl-XL, Bcl-w, and Mcl-1. Indiscriminate binders of Bcl-2 family proteins such as Bim, tBid, and Puma are known to be more potent inducers of apoptotic cell death.
Refined structural models of Bcl-XL/p53TAD and Bcl-2/p53TAD complexes unveiled a molecular basis for the mechanism underlying the interaction between the p53TAD and anti-apoptotic Bcl-2 family proteins (Fig. 2C), which explains how the p53TAD could block the function of the anti-apoptotic Bcl-2 family proteins. The sites filled by the p53TAD peptide in Bcl-2 and Bcl-XL are significantly overlapped with those occupied by the BH3 peptide of the pro-apoptotic effectors Bak and Bax, indicating that binding of both peptides to Bcl-2 and Bcl-XL is mutually exclusive. Hence, the p53TAD would competitively block the binding of Bak and Bax to Bcl-2 and Bcl-XL, which consequently liberates Bak and Bax (Fig. 1). Based on these results, it was proposed that the p53TAD can lead to mitochondrial apoptosis through competitive inhibition of the interaction between pro-apoptotic Bcl-2 family proteins and anti-apoptotic Bcl-2 family proteins. This multifunctional role of p53 might provide an alternative mechanism of regulation for Bcl-2 family proteins.
APPLICATION TO ANTICANCER THERAPY
Since the interaction of p53 with MDM2 is a promising target in anticancer therapy, various peptidomimetic small molecules have been developed as protein-protein interaction blockers using the structure of the p53TAD peptide bound to MDM2 (9,38,39). Nutlin-3 is an imidazoline-based MDM2 antagonist that potently inhibits the interaction between p53TAD and MDM2 (40,41) (Fig. 2A). Currently, Nutlin derivatives are in early clinical development as anticancer therapeutics. Previously, anticancer apoptotic pathway activation by Nutlin was thought to stem only from its inhibition of the p53-MDM2 interaction. However, recent studies showed that Nutlin exerts its pharmacological action via transcription-independent as well as transcription-dependent apoptosis program of p53 (42). Recently, it was also shown that the transcription-independent p53 apoptosis largely contributes to Nutlin-triggered apoptosis in cancer cells (43,44).
Anti-apoptotic Bcl-2 family proteins are anticancer therapeutic targets since they are overexpressed in many tumor cells and are associated with cancer progression and resistance against anticancer chemotherapy (45,46). Although the Bcl-2 family protein inhibitor ABT-737 induces the regression of solid tumors (47), it can bind only to Bcl-2, Bcl-w, and Bcl-XL, but not to Mcl-1, which results in resistance to apoptosis induced by ABT-737 (48,49). Therefore, concomitant inhibition of various anti-apoptotic Bcl-2 family proteins is more effective in anticancer therapy.
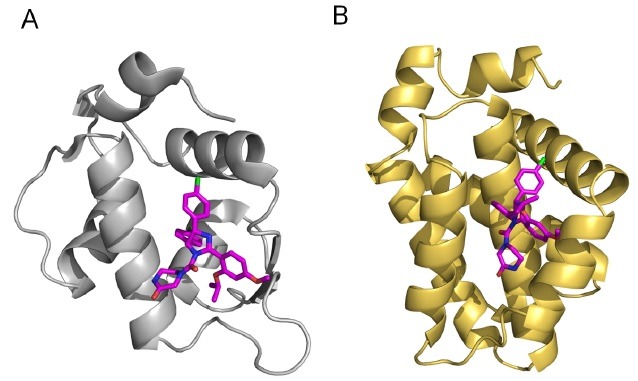
A recent study provided a structural model of the Bcl-XL/Nutlin-3 complex (Fig. 3B), which suggests the multi-target-based mechanism of Nutlin-3 binding to distinct target proteins, MDM2 and Bcl-XL (50). MDM2 and Bcl-XL display high similarity in their binding pockets, although their three-dimensional structures are quite different. Structural comparison between the Bcl-XL/Nutlin-3 and the MDM2/Nutlin-3 complexes showed a highly conserved binding mechanism of Nutlin-3 to MDM2 and Bcl-XL (Fig. 3). The imidazoline-based scaffold structure of Nutlin-3 resembles the α-helical structure of both the pro-apoptotic BH3 and p53TAD peptide (residues 15-29) backbones (41,51). In addition, the aromatic rings of Nutlin-3 bind to the binding sites surrounded by the bulky hydrophobic side-chains of MDM2 and Bcl-XL. A chlorophenyl ring of Nutlin-3 makes the most important contribution to complex formation with both MDM2 and Bcl-XL by fitting snugly into the hydrophobic grooves. Therefore, the overall mechanism underlying the binding of Nutlin-3 with Bcl-XL is analogous to that of Nutlin-3 with MDM2.
Fig. 3. Comparison between MDM2 and Bcl-XL in the binding mode of Nutlin-3. (A) Crystal structure of Nutlin-3 in complex with MDM2 (PDB code: 4HG7). (B) A refined structural model of Nutlin-3 in complex with Bcl-XL generated from an NMR data-driven structure calculation (50). Nutlin-3 is shown in purple.
MDM2 and Bcl-2 are promising anticancer target proteins that are related to cell cycle arrest and mitochondrial apoptosis, respectively (52). Concurrent suppression of MDM2 and Bcl-2 has a synergistic apoptotic effect in acute myeloid leukemia (AML) (53). In addition, co-treatment with Nutlin-3 and ABT-737 greatly enhanced the sensitivity of cells with high MDM2 levels to apoptosis (54). Thus, the combined inhibition of MDM2 and Bcl-2 could be a multi-target-based anticancer strategy to trigger tumor cell death. In combination with the previously reported results that the Bcl-2 inhibitor BH3I blocked the interaction between p53 and MDM2 (55), the Bcl-XL/Nutlin-3 complex structure sheds lights on structure-based design of a multi-targeting anticancer agent that can simultaneously inhibit MDM2 and Bcl-XL proteins. The three-dimensional structural information could also be utilized to optimize multi-target anticancer therapeutics.
CONCLUSION
To summarize, the molecular interaction of p53 with anti-apoptotic Bcl-2 family proteins serves a critical role in transcription-independent apoptotic pathway of p53. Recent studies showed that MDM2 and anti-apoptotic Bcl-2 family proteins have a highly conserved binding mechanism with the p53TAD. Based on this similarity, it was revealed that a potent MDM2 inhibitor Nutlin-3 can target the transcription-independent apoptotic pathway of p53 by binding anti-apoptotic Bcl-2 family proteins. These results provided structural insight into a multi-target-based therapeutic strategy, which have significant implications in the fields of cancer drug discovery.
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (NRF-2011-0016011).
References
- 1.Harris S. L., Levine A. J. The p53 pathway: positive and negative feedback loops. Oncogene. (2005);24:2899–2908. doi: 10.1038/sj.onc.1208615. [DOI] [PubMed] [Google Scholar]
- 2.Sherr C. J. Principles of tumor suppression. Cell. (2004);116:235–246. doi: 10.1016/S0092-8674(03)01075-4. [DOI] [PubMed] [Google Scholar]
- 3.Vogelstein B., Lane D., Levine A. J. Surfing the p53 network. Nature. (2000);408:307–310. doi: 10.1038/35042675. [DOI] [PubMed] [Google Scholar]
- 4.Vousden K. H., Lu X. Live or let die: the cell's response to p53. Nature Rev. (2002);2:594–604. doi: 10.1038/nrc864. [DOI] [PubMed] [Google Scholar]
- 5.Hollstein M., Sidransky D., Vogelstein B., Harris C. C. p53 mutations in human cancers. Science. (1991);253:49–53. doi: 10.1126/science.1905840. [DOI] [PubMed] [Google Scholar]
- 6.Ventura A., Kirsch D. G., McLaughlin M. E., Tuveson D. A., Grimm J., Lintault L., Newman J., Reczek E. E., Weissleder R., Jacks T. Restoration of p53 function leads to tumour regression in vivo. Nature. (2007);445:661–665. doi: 10.1038/nature05541. [DOI] [PubMed] [Google Scholar]
- 7.Xue W., Zender L., Miething C., Dickins R. A., Hernando E., Krizhanovsky V., Cordon-Cardo C., Lowe S. W. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature. (2007);445:656–660. doi: 10.1038/nature05529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Martins C. P., Brown-Swigart L., Evan G. I. Modeling the therapeutic efficacy of p53 restoration in tumors. Cell. (2006);127:1323–1334. doi: 10.1016/j.cell.2006.12.007. [DOI] [PubMed] [Google Scholar]
- 9.Kussie P. H., Gorina S., Marechal V., Elenbaas B., Moreau J., Levine A. J., Pavletich N. P. Structure of the MDM2 oncoprotein bound to the p53 tumor suppressor transactivation domain. Science. (1996);274:948–953. doi: 10.1126/science.274.5289.948. [DOI] [PubMed] [Google Scholar]
- 10.Chipuk J. E., Bouchier-Hayes L., Kuwana T., Newmeyer D. D., Green D. R. PUMA couples the nuclear and cytoplasmic proapoptotic function of p53. Science. (2005);309:1732–1735. doi: 10.1126/science.1114297. [DOI] [PubMed] [Google Scholar]
- 11.Chipuk J. E., Kuwana T., Bouchier-Hayes L., Droin N. M., Newmeyer D. D., Schuler M., Green D. R. Direct activation of Bax by p53 mediates mitochondrial membrane permeabilization and apoptosis. Science. (2004);303:1010–1014. doi: 10.1126/science.1092734. [DOI] [PubMed] [Google Scholar]
- 12.Jiang P., Du W., Heese K., Wu M. The Bad guy cooperates with good cop p53: Bad is transcriptionally up-regulated by p53 and forms a Bad/p53 complex at the mitochondria to induce apoptosis. Mol. Cell Biol. (2006);26:9071–9082. doi: 10.1128/MCB.01025-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Leu J. I., Dumont P., Hafey M., Murphy M. E., George D. L. Mitochondrial p53 activates Bak and causes disruption of a Bak-Mcl1 complex. Nat. Cell Biol. (2004);6:443–450. doi: 10.1038/ncb1123. [DOI] [PubMed] [Google Scholar]
- 14.Mihara M., Erster S., Zaika A., Petrenko O., Chittenden T., Pancoska P., Moll U. M. p53 has a direct apoptogenic role at the mitochondria. Mol. Cell. (2003);11:577–590. doi: 10.1016/S1097-2765(03)00050-9. [DOI] [PubMed] [Google Scholar]
- 15.Tomita Y., Marchenko N., Erster S., Nemajerova A., Dehner A., Klein C., Pan H., Kessler H., Pancoska P., Moll U. M. WT p53, but not tumor-derived mutants, bind to Bcl2 via the DNA binding domain and induce mitochondrial permeabilization. J. Biol. Chem. (2006);281:8600–8606. doi: 10.1074/jbc.M507611200. [DOI] [PubMed] [Google Scholar]
- 16.Zhu J., Zhou W., Jiang J., Chen X. Identification of a novel p53 functional domain that is necessary for mediating apoptosis. J. Biol. Chem. (1998);273:13030–13036. doi: 10.1074/jbc.273.21.13030. [DOI] [PubMed] [Google Scholar]
- 17.Chang J., Kim D. H., Lee S. W., Choi K. Y., Sung Y. C. Transactivation ability of p53 transcriptional activation domain is directly related to the binding affinity to TATA-binding protein. J. Biol. Chem. (1995);270:25014–25019. doi: 10.1074/jbc.270.42.25014. [DOI] [PubMed] [Google Scholar]
- 18.Gu W., Shi X. L., Roeder R. G. Synergistic activation of transcription by CBP and p53. Nature. (1997);387:819–823. doi: 10.1038/42972. [DOI] [PubMed] [Google Scholar]
- 19.Uesugi M., Verdine G. L. The alpha-helical FXXPhiPhi motif in p53: TAF interaction and discrimination by MDM2. Proc. Natl. Aca. Sci. U.S.A. (1999);96:14801–14806. doi: 10.1073/pnas.96.26.14801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ha J. H., Shin J. S., Yoon M. K., Lee M. S., He F., Bae K. H., Yoon H. S., Lee C. K., Park S. G., Muto Y., Chi S. W. Dual-site interactions of p53 protein transactivation domain with anti-apoptotic Bcl-2 family proteins reveal a highly convergent mechanism of divergent p53 pathways. J. Biol. Chem. (2013);288:7387–7398. doi: 10.1074/jbc.M112.400754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ha J. H., Won E. Y., Shin J. S., Jang M., Ryu K. S., Bae K. H., Park S. G., Park B. C., Yoon H. S., Chi S. W. Molecular mimicry-based repositioning of nutlin-3 to anti-apoptotic Bcl-2 family proteins. J. Am. Chem. Soc. (2011);133:1244–1247. doi: 10.1021/ja109521f. [DOI] [PubMed] [Google Scholar]
- 22.Bullock A. N., Henckel J., Fersht A. R. Quantitative analysis of residual folding and DNA binding in mutant p53 core domain: definition of mutant states for rescue in cancer therapy. Oncogene. (2000);19:1245–1256. doi: 10.1038/sj.onc.1203434. [DOI] [PubMed] [Google Scholar]
- 23.Kim H. R., Heo Y. M., Jeong K. I., Kim Y. M., Jang H. L., Lee K. Y., Yeo C. Y., Kim S. H., Lee H. K., Kim S. R., Kim E. G., Choi J. K. FGF-2 inhibits TNF-alpha mediated apoptosis through upregulation of Bcl2-A1 and Bcl-xL in ATDC5 cells. BMB Rep. (2012);45:287–292. doi: 10.5483/BMBRep.2012.45.5.287. [DOI] [PubMed] [Google Scholar]
- 24.Qin B., Xiao B., Liang D., Li Y., Jiang T., Yang H. MicroRNA let-7c inhibits Bcl-xl expression and regulates ox-LDL-induced endothelial apoptosis. BMB Rep. (2012);45:464–469. doi: 10.5483/BMBRep.2012.45.8.033. [DOI] [PubMed] [Google Scholar]
- 25.Kelekar A., Thompson C. B. Bcl-2-family proteins: the role of the BH3 domain in apoptosis. Trends Cell Biol. (1998);8:324–330. doi: 10.1016/S0962-8924(98)01321-X. [DOI] [PubMed] [Google Scholar]
- 26.Youle R. J., Strasser A. The BCL-2 protein family: opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. (2008);9:47–59. doi: 10.1038/nrm2308. [DOI] [PubMed] [Google Scholar]
- 27.Lee D. H., Ha J. H., Kim Y., Bae K. H., Park J. Y., Choi W. S., Yoon H. S., Park S. G., Park B. C., Yi G. S., Chi S. W. Interaction of a putative BH3 domain of clusterin with anti-apoptotic Bcl-2 family proteins as revealed by NMR spectroscopy. Biochem. Biophys. Res. Commun. (2011);408:541–547. doi: 10.1016/j.bbrc.2011.04.054. [DOI] [PubMed] [Google Scholar]
- 28.Sot B., Freund S. M., Fersht A. R. Comparative biophysical characterization of p53 with the pro-apoptotic BAK and the anti-apoptotic BCL-xL. J. Biol. Chem. (2007);282:29193–29200. doi: 10.1074/jbc.M705544200. [DOI] [PubMed] [Google Scholar]
- 29.Hagn F., Klein C., Demmer O., Marchenko N., Vaseva A., Moll U. M., Kessler H. BclxL changes conformation upon binding to wild-type but not mutant p53 DNA binding domain. J. Biol. Chem. (2010);285:3439–3450. doi: 10.1074/jbc.M109.065391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Petros A. M., Gunasekera A., Xu N., Olejniczak E. T., Fesik S. W. Defining the p53 DNA-binding domain/Bcl-x(L)-binding interface using NMR. FEBS Lett. (2004);559:171–174. doi: 10.1016/S0014-5793(04)00059-6. [DOI] [PubMed] [Google Scholar]
- 31.Lee D. H., Ha J. H., Kim Y., Jang M., Park S. J., Yoon H. S., Kim E. H., Bae K. H., Park B. C., Park S. G., Yi G. S., Chi S. W. A conserved mechanism for binding of p53 DNA-binding domain and anti-apoptotic Bcl-2 family proteins. Mol. Cells. (2014);37:264–269. doi: 10.14348/molcells.2014.0001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chipuk J. E., Maurer U., Green D. R., Schuler M. Pharmacologic activation of p53 elicits Bax-dependent apoptosis in the absence of transcription. Cancer Cell. (2003);4:371–381. doi: 10.1016/S1535-6108(03)00272-1. [DOI] [PubMed] [Google Scholar]
- 33.Sayan B. S., Sayan A. E., Knight R. A., Melino G., Cohen G. M. p53 is cleaved by caspases generating fragments localizing to mitochondria. J. Biol. Chem. (2006);281:13566–13573. doi: 10.1074/jbc.M512467200. [DOI] [PubMed] [Google Scholar]
- 34.Xu H., Tai J., Ye H., Kang C. B., Yoon H. S. The N-terminal domain of tumor suppressor p53 is involved in the molecular interaction with the anti-apoptotic protein Bcl-Xl. Biochem. Biophys. Res. Commun. (2006);341:938–944. doi: 10.1016/j.bbrc.2005.12.227. [DOI] [PubMed] [Google Scholar]
- 35.Xu H., Ye H., Osman N. E., Sadler K., Won E. Y., Chi S. W., Yoon H. S. The MDM2-binding region in the transactivation domain of p53 also acts as a Bcl-X(L)-binding motif. Biochemistry. (2009);48:12159–12168. doi: 10.1021/bi901188s. [DOI] [PubMed] [Google Scholar]
- 36.Bharatham N., Chi S. W., Yoon H. S. Molecular basis of Bcl-X(L)-p53 interaction: insights from molecular dynamics simulations. PLoS One. (2011);6:e26014. doi: 10.1371/journal.pone.0026014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ha J., Won E., Yoon H., Kang S., Bae K., Park S., Park B., Choi B., Lee J., Chi S. Molecular Interaction between a Bcl-2 Homolog from Kaposi Sarcoma Virus and p53. Bull. Korean Chem. Soc. (2009);30:1655. doi: 10.5012/bkcs.2009.30.7.1655. [DOI] [Google Scholar]
- 38.Lee M. S., Ha J. H., Yoon H. S., Lee C. K., Chi S. W. Structural basis for the conserved binding mechanism of MDM2-inhibiting peptides and anti-apoptotic Bcl-2 family proteins. Biochem. Biophys. Res. Commun. (2014);445:120–125. doi: 10.1016/j.bbrc.2014.01.130. [DOI] [PubMed] [Google Scholar]
- 39.Shin J. S., Ha J. H., Chi S. W. Targeting of p53 peptide analogues to anti-apoptotic Bcl-2 family proteins as revealed by NMR spectroscopy. Biochem. Biophys. Res. Commun. (2014);443:882–887. doi: 10.1016/j.bbrc.2013.12.054. [DOI] [PubMed] [Google Scholar]
- 40.Fesik S. W. Promoting apoptosis as a strategy for cancer drug discovery. Nat. Rev. Cancer. (2005);5:876–885. doi: 10.1038/nrc1736. [DOI] [PubMed] [Google Scholar]
- 41.Vassilev L. T., Vu B. T., Graves B., Carvajal D., Podlaski F., Filipovic Z., Kong N., Kammlott U., Lukacs C., Klein C., Fotouhi N., Liu E. A. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science. (2004);303:844–848. doi: 10.1126/science.1092472. [DOI] [PubMed] [Google Scholar]
- 42.Kojima K., Konopleva M., McQueen T., O S., Plunkett W., Andreeff M. Mdm2 inhibitor Nutlin-3a induces p53-mediated apoptosis by transcription-dependent and transcription-independent mechanisms and may overcome Atm-mediated resistance to fludarabine in chronic lymphocytic leukemia. Blood. (2006);108:993–1000. doi: 10.1182/blood-2005-12-5148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Vaseva A. V., Marchenko N. D., Moll U. M. The transcription-independent mitochondrial p53 program is a major contributor to nutlin-induced apoptosis in tumor cells. Cell Cycle. (2009);8:1711–1719. doi: 10.4161/cc.8.11.8596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Saha M. N., Jiang H., Chang H. Molecular mechanisms of nutlin-induced apoptosis in multiple myeloma: evidence for p53-transcription-dependent and -independent pathways. Cancer Biol. Ther. (2010);10:567–578. doi: 10.4161/cbt.10.6.12535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kang M. H., Reynolds C. P. Bcl-2 inhibitors: targeting mitochondrial apoptotic pathways in cancer therapy. Clin. Cancer Res. (2009);15:1126–1132. doi: 10.1158/1078-0432.CCR-08-0144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lessene G., Czabotar P. E., Colman P. M. BCL-2 family antagonists for cancer therapy. Nat. Rev. Drug Discov. (2008);7:989–1000. doi: 10.1038/nrd2658. [DOI] [PubMed] [Google Scholar]
- 47.Oltersdorf T., Elmore S. W., Shoemaker A. R., Armstrong R. C., Augeri D. J., Belli B. A., Bruncko M., Deckwerth T. L., Dinges J., Hajduk P. J., Joseph M. K., Kitada S., Korsmeyer S. J., Kunzer A. R., Letai A., Li C., Mitten M. J., Nettesheim D. G., Ng S., Nimmer P. M., O'Connor J. M., Oleksijew A., Petros A. M., Reed J. C., Shen W., Tahir S. K., Thompson C. B., Tomaselli K. J., Wang B., Wendt M. D., Zhang H., Fesik S. W., Rosenberg S. H. An inhibitor of Bcl-2 family proteins induces regression of solid tumours. Nature. (2005);435:677–681. doi: 10.1038/nature03579. [DOI] [PubMed] [Google Scholar]
- 48.Konopleva M., Contractor R., Tsao T., Samudio I., Ruvolo P. P., Kitada S., Deng X., Zhai D., Shi Y. X., Sneed T., Verhaegen M., Soengas M., Ruvolo V. R., McQueen T., Schober W. D., Watt J. C., Jiffar T., Ling X., Marini F. C., Harris D., Dietrich M., Estrov Z., McCubrey J., May W. S., Reed J. C., Andreeff M. Mechanisms of apoptosis sensitivity and resistance to the BH3 mimetic ABT-737 in acute myeloid leukemia. Cancer Cell. (2006);10:375–388. doi: 10.1016/j.ccr.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 49.van Delft M. F., Wei A. H., Mason K. D., Vandenberg C. J., Chen L., Czabotar P. E., Willis S. N., Scott C. L., Day C. L., Cory S., Adams J. M., Roberts A. W., Huang D. C. The BH3 mimetic ABT-737 targets selective Bcl-2 proteins and efficiently induces apoptosis via Bak/Bax if Mcl-1 is neutralized. Cancer Cell. (2006);10:389–399. doi: 10.1016/j.ccr.2006.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Shin J. S., Ha J. H., He F., Muto Y., Ryu K. S., Yoon H. S., Kang S., Park S. G., Park B. C., Choi S. U., Chi S. W. Structural insights into the dual-targeting mechanism of Nutlin-3. Biochem. Biophys. Res. Commun. (2012);420:48–53. doi: 10.1016/j.bbrc.2012.02.113. [DOI] [PubMed] [Google Scholar]
- 51.Lee E. F., Sadowsky J. D., Smith B. J., Czabotar P. E., Peterson-Kaufman K. J., Colman P. M., Gellman S. H., Fairlie W. D. High-resolution structural characterization of a helical alpha/beta-peptide foldamer bound to the anti-apoptotic protein Bcl-xL. Angew. Chem. Int. Ed. Engl. (2009);48:4318–4322. doi: 10.1002/anie.200805761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Vassilev L. T. MDM2 inhibitors for cancer therapy. Trends Mol. Med. (2007);13:23–31. doi: 10.1016/j.molmed.2006.11.002. [DOI] [PubMed] [Google Scholar]
- 53.Kojima K., Konopleva M., Samudio I. J., Schober W. D., Bornmann W. G., Andreeff M. Concomitant inhibition of MDM2 and Bcl-2 protein function synergistically induce mitochondrial apoptosis in AML. Cell Cycle. (2006);5:2778–2786. doi: 10.4161/cc.5.23.3520. [DOI] [PubMed] [Google Scholar]
- 54.Wade M., Rodewald L. W., Espinosa J. M., Wahl G. M. BH3 activation blocks Hdmx suppression of apoptosis and cooperates with Nutlin to induce cell death. Cell Cycle. (2008);7:1973–1982. doi: 10.4161/cc.7.13.6072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Porter J. R., Helmers M. R., Wang P., Furman J. L., Joy S. T., Arora P. S., Ghosh I. Profiling small molecule inhibitors against helix-receptor interactions: the Bcl-2 family inhibitor BH3I-1 potently inhibits p53/hDM2. Chem. Commun. (2010);46:8020–8022. doi: 10.1039/c0cc02969f. [DOI] [PMC free article] [PubMed] [Google Scholar]