

Abstract
Drug-resistance and imbalance of apoptotic regulation limit chemotherapy clinical application for the human hepatocellular carcinoma (HCC) treatment. The reactivation of p53 is an attractive therapeutic strategy in cancer with disrupted-p53 function. Nutlin-3, a MDM2 antagonist, has antitumor activity in various cancers. The post-translational modifications of p53 are a hot topic, but there are some controversy ideas about the function of phospho-Ser392-p53 protein in cancer cell lines in response to Nutlin-3. Therefore, we investigated the relationship between Nutlin-3 and phospho-Ser392-p53 protein expression levels in SMMC-7721 (wild-type TP53) and HuH-7 cells (mutant TP53). We demonstrated that Nutlin-3 induced apoptosis through down-regulation phospho-Ser392-p53 in two HCC cells. The result suggests that inhibition of p53 phosphorylation on Ser392 presents an alternative for HCC chemotherapy. [BMB Reports 2014; 47(4): 221-226]
Keywords: Apoptosis, Human hepatocellular carcinoma, Nutlin-3, p53, Phospho-Ser392-p53
INTRODUCTION
Liver cancer is the fifth most commonly diagnosed cancer in men and the seventh in women (1), and the fourth leading cause of cancer deaths worldwide (2), but it lacks effective therapies. The chemotherapy remains the viable option for the hepatocellular carcinoma (HCC) treatment, but the dose-dependent toxicities and drug-resistance limit its clinical applications. Therefore, the chemotherapy efficacy against HCC is still unsatisfactory.
The tumor suppressor p53, a “genome gatekeeper,” is usually inactivated or lost in human cancer cells (3). Even in cancers with wild-type TP53, p53 transcriptional activity is primarily repressed by murine double minute 2 (MDM2), which directly binds to and blocks the N-terminal transactivation domain of p53 (4), leading to p53 ubiquitination, nuclear export or cytoplasmic degradation in proteasome (5,6). Therefore, blocking the MDM2-p53 interaction is a promising therapeutic strategy for cancer. The function of p53 is also highly modulated by post-translational modifications (7). For instance, in response to genotoxic stress, p53 is stabilized via post-translational modifications which prevent MDM2 binding (8). Phosphorylation on Ser392 of p53 is a common and integral event by diverse stimuli (9). However, there are some controversy ideas about the function of phospho-Ser392-p53. Some studies showed that dephosphorylation on Ser392 of both wild-type and mutant p53 correlated with the cellular resistance to DNA-damaging agents such as UVB radiation and Cisplatin (10). Accordingly, phosphorylation on Ser392 of p53 plays an important role in its stabilization and tetramer formation (11). In agreement, hypoxia decreased phosphorylation on Ser392 and inhibited p53-dependent apoptosis (12). Others reported that high level of phospho-Ser392-p53 has been linked with high level of lymphatic invasion and poorer prognosis (13,14). Additionally, phosphorylation on Ser392 has other negative roles on p53, such as promotion of p53 export from the nucleus (15), MDM2-mediated degradation (16), and reduced p53 transforming activity (16). Casein kinase 2 (CK2) and cyclin dependent kinase 9 (CDK9) phosphorylate Ser392 of p53 in response to UV radiation (17). In the last decade, several inhibitors of CK2 have been discovered as potential anticancer drugs (18).
Nutlin-3, an MDM2 antagonist, was used to activate p53 (19). Although a number of studies have attempted to elucidate the mechanism of Nutlin-3 function, in hepatoma cells the effects of Nutlin-3 on phospho-Ser392-p53 protein still remain to be elucidated. In the present study, we investigated the relationship between Nutlin-3 and phospho-Ser392-p53 protein expression levels in two HCC cell lines with different TP53 type. The current study revealed that Nutlin-3 induced apoptosis through down-regulated phospho-Ser392-p53 protein in HCC cells. We identified phosphorylation on p53-Ser392 as a mechanism to regulate HCC cell survival. Our results indicate that inhibition of phosphorylation on Ser392 of p53 could be a promising chemotherapeutic approach to the HCC treatment.
RESULTS
Nutlin-3 inhibits cell proliferation in SMMC-7721 and HuH-7 cells
To test the effect of Nutlin-3 on cell growth, SMMC-7721 (wild-type TP53) and Huh7 (mutant TP53) were exposed to Nutlin-3 (1.25, 2.5, 5, 10, and 20 μM respectively) for 24 h, 48 h, and 72 h and performed CCK8 analysis. Meanwhile, we selected SMMC-7721 cells for the p53-inhibited model, which were pretreated with PFT-α, a p53 inhibitor (20), for 12 h to inhibit p53 activity, and then exposed to Nutlin-3 together with PFT-α for 12 h, 36 h, and 60 h at the same conditions. We observed that the number of Nutlin-3-treated cells decreased with increased concentration at 48 h treatment (Fig. 1A). Meanwhile, SMMC-7721 cells were more sensitive to Nutlin-3 than HuH-7 cells (Fig. 1B). These results showed that Nutlin-3 can inhibit two HCC cell growths.
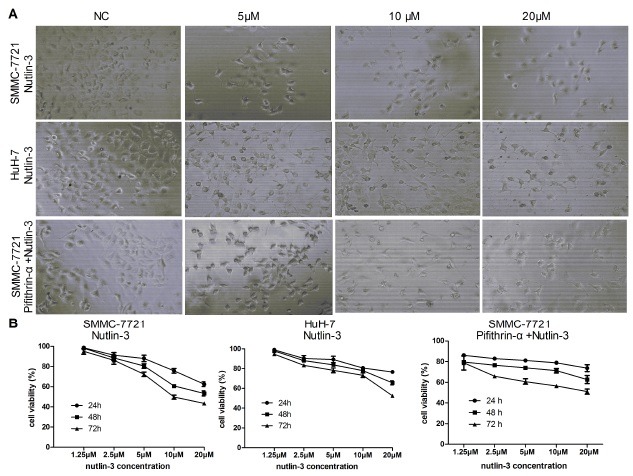
Fig. 1. Nutlin-3 inhibits cell proliferation in SMMC-7721 and HuH-7 cells. (A) The decreased number cells were detected at 48 h treatment with different concentration of Nutlin- 3 as indicated (100×). (B) Inhibitory effect of Nutlin-3 on the cell proliferation. Two HCC cell lines were treated with Nutlin-3 as indicated. Data are shown by means ± S.D. (n = 3).
Nutlin-3 induces apoptosis in SMMC-7721 and HuH-7 cells
To investigate whether Nutlin-3 induces apoptosis in the HCC cells, Hoechst 33258 staining showed that Nutlin-3-treated SMMC-7721 (with or without PFT-α) and HuH-7 cells had marked morphological changes including chromatin condensation or fragmentation, and apoptotic bodies, while most control cells had regular nuclei (Fig. 2). Interestingly, SMMC-7721 cells with inhibited p53 activation and p53-mutant HuH-7 cells were observed the apoptotic morphology treated by Nutlin-3. These results demonstrated that Nutlin-3 induced morphological characteristic of cell apoptosis.
Fig. 2. Nutlin-3 induced apoptotic nuclear changes. Nuclear staining of cells with Hoechst 33258 treated as indicated in method. The apoptotic nuclear changes with smaller and brilliant staining (white arrows) were examined by fluorescence microscopy (200×).
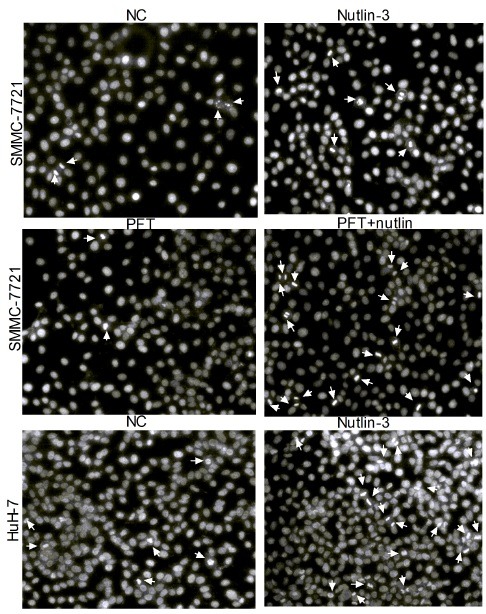
Additional evidence for the occurrence of apoptosis was quantified by Flow Cytometry under Annexin V-FITC/PI double staining. FACS analysis showed that compared with the control (4.13% ± 0.40%), the apoptotic percentage of Nutlin group (10.37% ± 1.51%) and PFT + Nutlin group (10.73% ± 0.49%) significantly up-regulated in SMMC-7721 cells (P < 0.05; Fig. 3A and B). No significant differences were observed between the two groups (P > 0.05; Fig. 3B). Interestingly, the apoptotic percentage of Nutlin-3-treated HuH-7 cells significantly increased at 11.10% ± 2.55%, while the control was 1.03% ± 0.15% (P < 0.05; Fig. 3A and B). These data demonstrated that Nutlin-3 induces p53-independent apoptosis in SMMC- 7721 and HuH-7 cells.
Fig. 3. Nutlin-3 induced apoptosis in two HCC cell lines. (A) Representative Flow Cytometric graphs of Annexin V-FITC/PI double staining treated with or without nutlin-3 (10 μM) as indicated in method. (B) The apoptotic percentage was quantified via Flow Cytometry under Annexin V-FITC/PI staining. Data are shown by means ± S.D. (n = 3), *P < 0.05 vs. NC group. (C) Western blots were performed for protein expression of Bax, Bcl-2, and caspase-3. β-actin was used as the loading control.
Mechanistically, in Nutlin-3-treated SMMC-7721 (with or without PFT-α) and HuH-7 cells, the reduced expression of anti-apoptotic Bcl-2 protein and increased pro-apoptotic Bax protein were further showed the effect of Nutlin-3 on cell apoptosis compared to control cells by Western blot analysis (Fig. 3C). Furthermore, the increased caspase-3 expression level confirms execution of apoptosis in Nutlin-3 treated HCC cells (Fig. 3C). Together, these findings provided strong evidence that Nutlin-3 has an apoptotic effect on HCC cells. Additionally, the SMMC-7721 cells only incubated with PFT-α had reduced Bax, increased Bcl-2, and reduced apoptotic percentage (3.87% ± 0.21%) compared to the control (Fig. 3B and C). Thus, PFT-α had inhibited SMMC-7721 cell death.
Nutlin-3 down-regulates the protein expression level of phospho-Ser392-p53
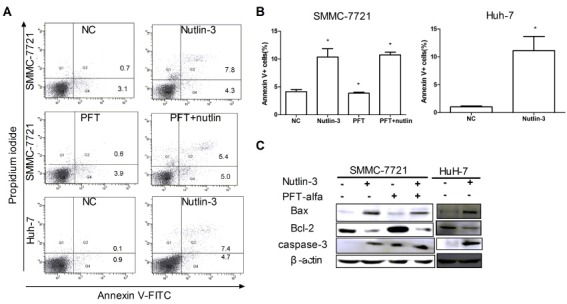
It has been reported that the status of p53 phosphorylation on Ser392 correlates with cell apoptosis (15). Therefore, we investigated whether Nutlin-3 regulates phos-p53-Ser392 in HCC cells. Western blot analysis showed that PFT-α inhibited p53 expression in SMMC-7721 cells (Fig. 4A). The result suggested that PFT-α acted as a wild-type p53 inhibitor in SMMC-7721 cells. Moreover, Nutlin-3 up-regulated p53 level in SMCC-7721 cells (Fig. 4A). Similarly, we inhibited p53 activity by PFT-α, and then incubated them together with Nutlin-3 for 36 h. The p53 level in the PFT + Nutlin group increased compared with the control (Fig. 4A). These observations confirmed the results obtained in Fig. 3C and indicated that Nutlin-3 up-regulated p53 levels whether p53 activity was inhibited or not in SMCC-7721 cells. However, the protein expression level of p53 in Nutlin-3-treated HuH-7 cells down-regulated (Fig. 4A).
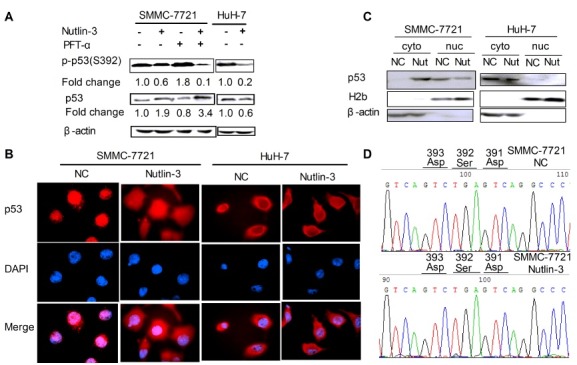
Fig. 4. Nutlin-3 down-regulates the protein expression levels of phospho-Ser392-p53. (A) The protein expression of phospho-Ser392-p53 and p53. The cells were treated as indicated. β-actin served as the loading control. (B) The subcellular localization of p53 was detected under immunofluorescence microscope (1,000×). The cells were treated as indicated and stained for p53 (red). Nuclei were counterstained with DAPI (blue). Images were merged using Image-Pro plus 6.0. (C) The subcellular localization of p53 was detected by western blot analysis. Anti-H2b and β-actin antibodies were used as a loading control of nuclear and cytoplasmic proteins, respectively. (cyto) cytoplasmic proteins; (nuc) nuclear proteins. (D) Representative sequencing data identified p53- Ser392 mutant from PCR products of SMMC- 7721 cells treated with or without Nutlin-3.
Next, an antibody specific for phospho-Ser392-p53 showed that this modification was in fact occurring. The decreased phospho-Ser392-p53 was observed under the up-regulated p53 level in SMMC-7721 cells treated with Nutlin-3 alone or together with PFT-α (Fig. 4A). Meanwhile, the level of phospho- Ser392-p53 protein was decreased in Nutlin-3-treated HuH-7 cells under down-regulated expression p53 condition (Fig. 4A). Surprisingly, we found that the increased phospho-Ser392-p53 protein under down-regulated p53 expression level in SMMC- 7721 cells treated with PFT-α alone (Fig. 4A). These results suggested that Nutlin-3 down-regulates the expression levels of phospho-Ser392-p53 regardless of p53 status.
Effects of Nutlin-3 on the subcellar localization of p53 in HuH-7 and SMMC-7721 cells
The p53 localization also has an important role in HCC apoptosis (21). To test whether Nutlin-3 could affect the subcellar localization of p53, we performed the immunostaining. The assay showed that wild-type p53 concentrated mainly in the nucleus of untreated SMMC-7721 cells (Fig. 4B). Interestingly, after Nutlin-3 treatment, p53 was throughout nucleus and cytoplasm of SMMC-7721 cells (Fig. 4B). The result demonstrated that Nutlin-3 promoted wild-type p53 export from the nucleus in SMMC-7721 cells. To further confirm the result, we performed Western blot analysis after cytoplasm and nucleus extraction. We also demonstrated that p53 remains in the nucleus of SMMC-7721 cells while partly localizes to the cytoplasm treated by Nutlin-3 (Fig. 4C). These observations confirmed and correlated with the results obtained in Fig. 4B. However, mutant p53 was predominantly present in the cytoplasm of HuH-7 cells treated with or without Nutlin-3 (Fig. 4B and C). Together, these results showed that Nutlin-3 could affect the subcellar localization of wild-type p53 in SMMC-7721 cells, but not mutant p53 in HuH-7 cells.
Mutation of Ser392 resulted in decreased phosphorylation and relocalization of wild-type p53 (15,22). To test whether Nutlin-3-treated SMMC-7721 cells express a mutant form of p53-Ser392, we sequenced the TP53 DNA including Ser392 site amplified by PCR. However, we have not detected any nucleotide mutations in the Ser392 site of DNA from SMMC-7721 cells treated with or without Nutlin-3 (Fig. 4D). We concluded that down-regulation of phospho-Ser392-p53 and re-localization of wild-type p53 might not induced by p53-Ser392 mutation in Nutlin-3-treated SMMC-7721 cells.
DISCUSSION
In this study, we investigated the anti-tumor activity of Nutlin-3 in two different HCC cells with a particular emphasis on the mechanism of cell apoptosis. Here, we provide the evidence that Nutlin-3 induced apoptosis and down-regulated the expression level of phospho-Ser392-p53 protein in HCC cells.
The reactivation of p53 is an attractive therapeutic strategy in a tumor with disrupted p53 function (23). In this study, we showed that Nutlin-3 particularly enriched wild-type p53 in PFT-α-treated SMMC-7721 cells with inhibited-p53 activation, consistent with the published finding that Nutlin-3 induces apoptosis with wild-type p53 accumulation (24). However, p53 is frequently mutated in HCC (25), and mutant p53 expressed level is elevated in HCC cells and tumor tissues (16). In HuH-7 cells, the study showed down-regulated level of mutant p53 protein accords with the oncogenic functions of mutant p53 including enhanced chemo-resistance (16) and genomic instability (26) in a range of cell lines (3,27). In this study, the down-regulation of mutant p53 protein may lead to decreased expression level of phospho-Ser392-p53 protein.
Mechanistically, down-regulation of wild-type p53 phosphorylation on Ser392 leads to decreased MDM2 and induces cell apoptosis in HCC, an observation closely resembles our results (28). In the present study, Nutlin-3 strongly decreased phospho-Ser392-p53 level in HuH-7 cells compared to that of Nutlin-3-treated SMMC-7721 cells, but apoptotic population by Nutlin-3 in both cells does not significantly different, which consistent with our finding that SMMC-7721 cells were more sensitive to Nutlin-3 than HuH-7 cells (Fig. 1B). Consistently, dephosphorylation of p53-Ser392 by TGF-β1 in HuH-7 cells was associated with increased caspase 3 activation and apoptosis (29). Moreover, tumors with mutant p53 usually have high levels of phospho-Ser392-p53 and correlated with malignancy, radio-resistance, and chemo-resistance (10,30). Therefore, we identified p53-Ser392 phosphorylation as a mechanism to regulate cell survival in HCC with wild-type or mutant TP53.
Lower level of MDM2 causes mono-ubiquitination and nuclear export of p53 (6). Meanwhile, we also found that Nutlin-3 promoted wild-type p53 export from the nucleus of SMMC-7721 cells, consistent with the published finding that Nutlin-3 acts as an MDM2 antagonist and induces p53 activation but no stabilization (31). In addition, the p53-Ser392 mutation can lead to a decreased level of phosphorylation (22), which an alanine mutant of p53 (p53-392A) is localized mainly in the nucleus, whereas a glutamic acid mutant, p53-392E, are distributed in the cytoplasm and nucleus (15). However, we did not find the relationship between the Ser392 mutation and phosphorylation/ localization of p53. Therefore, more studies are needed to decipher the mechanisms of p53 re-localization in Nutlin-3-treated SMMC-7721 cells.
In this study, we identified that Nutlin-3 can decrease p53-Ser392 phosphorylation lead to cell apoptosis in SMMC-7721 and HuH-7 cells. Based on the results of this study, we propose that inhibition of p53-Ser392 phosphorylation may be beneficial in treating liver tumors expressing wild-type or mutant p53, although the precise mechanism requires further studies.
MATERIALS AND METHODS
Cell lines and Nutlin-3 treatment
HuH-7 (mutant TP53) and SMMC-7721 (wild-type TP53) cell lines were generous gifts from Prof. Liu Cong of the West China Second University Hospital/West China Women’s and Children’s Hospital. The cultivation medium contains DMEM (Gibco, USA) supplemented with 10% fetal bovine serum (Gibco, USA), and 100 U/ml penicillin and 100 μg/ml streptomycin. These cells were cultured at 37℃ under a 5% CO2 atmosphere.
Nutlin-3 (Sigma, USA) and Pifithrin-α (PFT-α, Sigma, USA) were dissolved in DMSO (Sigma, USA) and stored as 10 mM and 20 mM stock solutions at -20℃, respectively. The cultivation medium containing 0.1% DMSO was used as the control culture.
The Nutlin group of HuH-7 and SMMC-7721 cell lines were cultured with 10 μM Nutlin-3 for 48 h (32). The PFT group of SMMC-7721 cells was treated with PFT-α (20 μM) for 48 h. The PFT + Nutlin group of SMMC-7721 cells were pretreated with PFT-α (20 μM) for 12 h and then exposed to Nutlin-3 (10 μM) for 36 h together with PFT-α (33).
Cell viability assay
Two cell lines were seeded in 96-well plates (1 × 104 cells/ well) and treated with Nutlin-3 at different concentration (1.25, 2.5, 5, 10, and 20 μM) for different time (24, 48, and 72 h). Then, the cells were treated using a Cell Counting Kit-8 (CCK-8, Dojindo Molecular Technology, Japan) according to manufacturer's protocol. Finally, optical density (OD) was monitored at 450 nm with 650 nm as a reference wavelength by Multiskan Spectrum Microplate Reader (Thermo, USA). The cell viability values were calculated as previously described (34).
Hoechst 33258 staining
Two HCC cell lines were cultured in 24-well plates (2 × 105 cells/well) for 48 h on glass cover slips with or without Nutlin-3 treatment. Then the cells were fixed and measured using the Hoechst 33258 Staining Kit (Boster, China) under the manufacturer’s guidance. After the cells were washed with PBS, morphological changes, including nuclear chromatin condensation and apoptotic bodies, were observed and photographed at 200× magnification under a fluorescence microscope (Olympus BX51).
Annexin-V/PI double staining assay
Briefly, two HCC cell lines were cultured in 6-well plates (3 × 105 cells/well) for 48 h with or without Nutlin-3 treatment. Then, the cells were trypsinized, collected and stained with an Annexin V-FITC Apoptosis Kit (Becton Dickinson Biosciences, USA) according to the manufacturer’s protocol. Finally, apoptotic cells were determined on a FACS Calibur Flow Cytometer (BD, USA).
Western blot analysis
Cytoplasm and nucleus extracts were prepared using a Nuclear and Cytoplasmic Protein Extraction Kit (PP2201, Bioteke, China) according to manufacturer's protocol. Whole-cell extract of HCC cells were prepared and subjected to immunoblot analysis. The primary antibodies were rabbit anti-Bcl-2 polyclonal antibody (#2876, CST, diluted 1:1,000), rabbit anti-Bax polyclonal antibody (#2772, CST, diluted 1:1,000), rabbit anti-p53 polyclonal antibody (BA0521, Boster, diluted 1:500), rabbit anti-p53 phospho (pS392) monoclonal antibody (#1644-1, Epitomics, diluted 1:1,000), rabbit anti-Caspase-3 monoclonal antibody (BA2142, Boster, diluted 1:500), rabbit anti-H2b polyclonal antibody (BS1657, Bioworld, diluted 1:500), and β-actin antibody (BA2305, Boster, diluted 1:500). The secondary antibodies, goat anti-mouse IgG-HRP (sc-2005, diluted 1:5,000) and goat anti-rabbit IgG-HRP (sc-2004, diluted 1:5,000), were purchased from Santa Cruz Biotechnology. β-actin served as the loading controls.
Immunofluorescence
HCC cells were cultured for 48 h on glass coverslips in 24-well plates (2 × 105 cells /well) with or without Nutlin-3 treatment, fixed, permeabilized, and then blocked. The samples were incubated with rabbit anti-p53 polyclonal antibody (BA0521, Boster, diluted 1:500), and then with sheep anti-rabbit Cy3-conjugated secondary antibody (C2306, Sigma, diluted 1:100). Cells were counterstained with 4', 6-diamidino-2-phenylindole dihydrochloride (DAPI) (10 μg/ml) (Sigma, USA). Images were captured under immunofluorescence microscope (Olympus BX51).
RT-PCR and sequencing of the TP53 gene at 392 site coding sequence
Total RNA was isolated with TRIzol (Invitrogen, China) and reverse transcribed using a Prime ScriptTM Kit (TaKaRa, China). Specific human wild-type TP53 primers were used as follows (forward and reverse): 5'-AGT CTA CCT CCC GCC ATA AAA-3' and 5'-AAG TCC TGG GTG CTT CTG AC-3'. The RT-PCR was performed as follows: 95℃ for 3 min, followed by 40 cycles of 95℃ for 10 s and 56℃ for 30 s. Subsequently, the products were sequenced with reverse TP53 primer by Sangon Biotech (Shanghai) Co., Ltd.
Statistical analysis
All statistical tests were performed using SPSS 18.0 (SPSS, Chicago, IL). The data are presented as mean ± SD. The means were analyzed using one-way ANOVA to compare the groups, followed by a post hoc Tukey’s test. Differences with P < 0.05 were considered statistically significant.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (Grant no. 81071362), the Scientific Research Foundation of China Higher Education (Grant no. 2010SCU21004), and the Science and Technology Project of Hebei Province of China (Grant no. 132777189). The authors thank Jingci Yang (the Second Hospital of Hebei Medical University), Ping Zhang (State Key Laboratory of Oral Diseases, Sichuan University), Lin Wei, Shuxia Song, Xiaolin Yin, Xuesong Qian and Wenjian Li (Hebei Medical University) for technical guidance and valuable comments. X.S. and M.L. designed research; X.S. performed experiments; X.S. wrote the manuscript with contributions from all authors.
References
- 1.Jemal A., Bray F., Center M. M., Ferlay J., Ward E., Forman D. Global cancer statistics. CA- Cancer J. Clin. (2011);61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Farazi P. A., DePinho R. A. Hepatocellular carcinoma pathogenesis: from genes to environment. Nat. Rev. Cancer. (2006);6:674–687. doi: 10.1038/nrc1934. [DOI] [PubMed] [Google Scholar]
- 3.Petitjean A., Achatz M., Borresen-Dale A., Hainaut P., Olivier M. TP53 mutations in human cancers: functional selection and impact on cancer prognosis and outcomes. Oncogene. (2007);26:2157–2165. doi: 10.1038/sj.onc.1210302. [DOI] [PubMed] [Google Scholar]
- 4.Lee S. H. Structural Origin for the Transcriptional Activity of Human p 53. BMB Rep. (2001);34:73–79. [Google Scholar]
- 5.Haupt Y., Maya R., Kazaz A., Oren M. Mdm2 promotes the rapid degradation of p53. Nature. (1997);387:296–299. doi: 10.1038/387296a0. [DOI] [PubMed] [Google Scholar]
- 6.Li M., Brooks C. L., Wu-Baer F., Chen D., Baer R., Gu W. Mono-versus polyubiquitination: differential control of p53 fate by Mdm2. Science. (2003);302:1972–1975. doi: 10.1126/science.1091362. [DOI] [PubMed] [Google Scholar]
- 7.Vousden K. H., Lane D. P. p53 in health and disease. Nat. Rev. Mol. Cell Bio. (2007);8:275–283. doi: 10.1038/nrm2147. [DOI] [PubMed] [Google Scholar]
- 8.Kruse J. P., Gu W. SnapShot: p53 posttranslational modifications. Cell. (2008);133:930. doi: 10.1016/j.cell.2008.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cox M. L., Meek D. W. Phosphorylation of serine 392 in p53 is a common and integral event during p53 induction by diverse stimuli. Cell. Signal. (2010);22:564–571. doi: 10.1016/j.cellsig.2009.11.014. [DOI] [PubMed] [Google Scholar]
- 10.Yap D. B. S., Hsieh J. K., Zhong S., Heath V., Gusterson B., Crook T., Lu X. Ser392 phosphorylation regulates the oncogenic function of mutant p53. Cancer Res. (2004);64:4749–4754. doi: 10.1158/0008-5472.CAN-1305-2. [DOI] [PubMed] [Google Scholar]
- 11.Sakaguchi K., Sakamoto H., Lewis M. S., Anderson C. W., Erickson J. W., Appella E., Xie D. Phosphorylation of serine 392 stabilizes the tetramer formation of tumor suppressor protein p53. Biochemistry. (1997);36:10117–10124. doi: 10.1021/bi970759w. [DOI] [PubMed] [Google Scholar]
- 12.Achison M., Hupp T. R. Hypoxia attenuates the p53 response to cellular damage. Oncogene. (2003);22:3431–3440. doi: 10.1038/sj.onc.1206434. [DOI] [PubMed] [Google Scholar]
- 13.Matsumoto M., Furihata M., Kurabayashi A., Sasaguri S., Araki K., Hayashi H., Ohtsuki Y. Prognostic significance of serine 392 phosphorylation in overexpressed p53 protein in human esophageal squamous cell carcinoma. Oncology. (2004);67:143–150. doi: 10.1159/000081001. [DOI] [PubMed] [Google Scholar]
- 14.Bar J. K., Slomska I., Rabczynki J., Noga L., Grybos M. Expression of p53 protein phosphorylated at serine 20 and serine 392 in malignant and benign ovarian neoplasms: correlation with clinicopathological parameters of tumors. Int. J. Gynecol. Cancer. (2009);19:1322–1328. doi: 10.1111/IGC.0b013e3181b70465. [DOI] [PubMed] [Google Scholar]
- 15.Kim Y. Y., Park B. J., Kim D. J., Kim W. H., Kim S., Oh K. S., Lim J. Y., Kim J., Park C., Park S. I. Modification of serine 392 is a critical event in the regulation of p53 nuclear export and stability. FEBS Lett. (2004);572:92–98. doi: 10.1016/j.febslet.2004.07.014. [DOI] [PubMed] [Google Scholar]
- 16.Muller P. A., Vousden K. H. p53 mutations in cancer. Nat. Cell Biol. (2013);15:2–8. doi: 10.1038/ncb2641. [DOI] [PubMed] [Google Scholar]
- 17.Radhakrishnan S. K., Gartel A. L. CDK9 phosphorylates p53 on serine residues 33, 315 and 392. Cell cycle. (2006);5:519–521. doi: 10.4161/cc.5.5.2514. [DOI] [PubMed] [Google Scholar]
- 18.Cozza G., A Pinna L., Moro S. Kinase CK2 Inhibition: An Update. Curr. Med. Chem. (2013);20:671–693. doi: 10.2174/092986713804999312. [DOI] [PubMed] [Google Scholar]
- 19.Shangary S., Wang S. Small-molecule inhibitors of the MDM2-p53 protein-protein interaction to reactivate p53 function: a novel approach for cancer therapy. Annu. Rev. Pharmacol. (2009);49:223–241. doi: 10.1146/annurev.pharmtox.48.113006.094723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Komarov P. G., Komarova E. A., Kondratov R. V., Christov-Tselkov K., Coon J. S., Chernov M. V., Gudkov A. V. A chemical inhibitor of p53 that protects mice from the side effects of cancer therapy. Science. (1999);285:1733–1737. doi: 10.1126/science.285.5434.1733. [DOI] [PubMed] [Google Scholar]
- 21.D Amaral J., M Xavier J., J Steer C., MP Rodrigues C. Targeting the p53 pathway of apoptosis. Curr. Pharm. Design. (2010);16:2493–2503. doi: 10.2174/138161210791959818. [DOI] [PubMed] [Google Scholar]
- 22.Ashcroft M., Kubbutat M. H., Vousden K. H. Regulation of p53 function and stability by phosphorylation. Mol. Cell. Biol. (1999);19:1751–1758. doi: 10.1128/mcb.19.3.1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sullivan K. D., Gallant-Behm C. L., Henry R. E., Fraikin J.-L., Espinosa J. M. The p53 circuit board. BBA-Rev. Cancer. (2012);1825:229–244. doi: 10.1016/j.bbcan.2012.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vassilev L. T., Vu B. T., Graves B., Carvajal D., Podlaski F., Filipovic Z., Kong N., Kammlott U., Lukacs C., Klein C. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science. (2004);303:844–848. doi: 10.1126/science.1092472. [DOI] [PubMed] [Google Scholar]
- 25.Guan Y.-S., La Z., Yang L., He Q., Li P. p53 gene in treatment of hepatic carcinoma: status quo. World J. Gastroentero. (2007);13:985–992. doi: 10.3748/wjg.v13.i2.228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hanel W., Moll U. M. Links between mutant p53 and genomic instability. J. Cell. Biochem. (2012);113:433–439. doi: 10.1002/jcb.23400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gillotin S., Yap D., Lu X. Mutation at Ser392 specifically sensitizes mutant p53H175 to mdm2-mediated degradation. Cell Cycle. (2010);9:1390–1398. doi: 10.4161/cc.9.7.11253. [DOI] [PubMed] [Google Scholar]
- 28.Furlan A., Stagni V., Hussain A., Richelme S., Conti F., Prodosmo A., Destro A., Roncalli M., Barila D., Maina F. Abl interconnects oncogenic Met and p53 core pathways in cancer cells. Cell Death Differ. (2011);18:1608–1616. doi: 10.1038/cdd.2011.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fan G., Ma X., Wong P., Rodrigues C., Steer C. p53 dephosphorylation and p21Cip1/Waf1 translocation correlate with caspase-3 activation in TGF-β1-induced apoptosis of HuH-7 cells. Apoptosis. (2004);9:211–221. doi: 10.1023/B:APPT.0000018803.11260.02. [DOI] [PubMed] [Google Scholar]
- 30.Matsumoto M., Furihata M., Ohtsuki Y. Posttranslational phosphorylation of mutant p53 protein in tumor development. Med. Mol. Morphol. (2006);39:79–87. doi: 10.1007/s00795-006-0320-0. [DOI] [PubMed] [Google Scholar]
- 31.Wallace M., Worrall E., Pettersson S., Hupp T. R., Ball K. L. Dual-site regulation of MDM2 E3-ubiquitin ligase activity. Mol. cell. (2006);23:251–263. doi: 10.1016/j.molcel.2006.05.029. [DOI] [PubMed] [Google Scholar]
- 32.Wang J., Zheng T., Chen X., Song X., Meng X., Bhatta N., Pan S., Jiang H., Liu L. MDM2 antagonist can inhibit tumor growth in hepatocellular carcinoma with different types of p53 in vitro. J. Gastroen. Hepatol. (2011);26:371–377. doi: 10.1111/j.1440-1746.2010.06440.x. [DOI] [PubMed] [Google Scholar]
- 33.Yu Y., Gu Z., Yin J., Chou W., Kwok C., Qin Z., LIANG Z. Ursolic acid induces human hepatoma cell line SMMC-7721 apoptosis via p53-dependent pathway. Chinese Med. J. (2010);123:1915–1923. [PubMed] [Google Scholar]
- 34.Cai X., Ye T., Liu C., Lu W., Lu M., Zhang J., Wang M., Cao P. Luteolin induced G2 phase cell cycle arrest and apoptosis on non-small cell lung cancer cells. Toxicol. In Vitro. (2011);25:1385–1391. doi: 10.1016/j.tiv.2011.05.009. [DOI] [PubMed] [Google Scholar]