

Abstract
Purpose
The objective of this study was to investigate the effects of feeding a high-concentrate corn straw (HCS) diet (65% concentrate+35% corn straw) on the epigenetic changes in the mammary tissue of dairy cows in comparison with a low-concentrate corn straw (LCS) diet (46% concentrate+54% corn straw) and with a low-concentrate mixed forage (LMF) diet (46% concentrate+54% mixed forage).
Experimental Design
Multiparous mid-lactation Chinese Holstein cows were fed one of these three diets for 6 weeks, at which time blood samples and mammary tissue samples were collected. Mammary arterial and venous blood samples were analyzed for lipopolysaccharide (LPS) concentrations while mammary tissue samples were assayed for histone H3 acetylation and the methylation of specific genes associated with fat and protein synthesis.
Results
Extraction of histones and quantification of histone H3 acetylation revealed that acetylation was significantly reduced in cows fed the HCS diet, as compared with cows fed the LCS diet. Cows fed the HCS diet had significantly higher LPS concentrations in the mammary arterial blood, as compared with cows fed the LCS diet. We found that the extent of histone H3 acetylation was negatively correlated with LPS concentrations. The methylation of the stearoyl-coenzyme A desaturase gene associated with milk fat synthesis was increased in cows fed the HCS diet. By contrast, methylation of the gene encoding the signal transducer and activator of transcription 5A was reduced in cows fed the HCS diet, suggesting that feeding a high-concentrate corn straw diet may alter the methylation of specific genes involved in fat and protein synthesis in the mammary tissue of dairy cows.
Conclusions
Feeding the high-concentrate diet induced epigenetic changes in the mammary tissues of dairy cows, possibly through effecting the release of differing amounts of LPS into the mammary blood.
Introduction
Epigenetics is defined as the study of heritable changes in gene expression that are independent of DNA sequence [1]. Major epigenetic events include acetylation and methylation of histones, DNA methylation, and gene regulation by non-coding RNAs. Histone binding to DNA in the eukaryotic nucleus modifies the properties of DNA. Various histone modifications include acetylation, methylation, phosphorylation, and ubiquitination [2]. The acetylation and methylation of histones H3 and H4, which occur at lysine or arginine residues, are the most common and well-studied histone modifications. DNA methylation refers to the addition of a methyl group at cytosine residues of the DNA template, which occurs predominantly in cytosine-phosphate-guanine (CpG) dinucleotides in mammals [3]. Epigenetic modifications of histones and DNA occur in response to environmental factors. Diet, toxins, pollutants, disease and other environmental factors may have profound impacts on gene expression through epigenetic regulation. The potential of epigenetic regulation to impact mammary function in dairy cows has received attention in recent years [4] [5].
In dairy production systems, cattle are often fed high-concentrate diets due to a seasonal or regional lack of quality forage material. The feeding of dairy cows diets that contain high proportions of concentrate to support high milk production has been associated with a high incidence of subacute ruminal acidosis (SARA) [6] [7] [8]. SARA is a digestive disorder that poses a health threat to lactating dairy cows and has become the most important metabolic disease in dairy production in the world. The high rumen digestibility of most grains in concentrate mixtures increases the production of volatile fatty acids in the rumen and causes a corresponding drop in rumen pH [6] [9] [10], which can result in alterations in the rumen environment, leading to changes in the composition of rumen microbiota [11] [12] and the accumulation of endotoxin (lipopolysaccharide; LPS), a potentially harmful cell-wall component of gram-negative bacteria [7] [13] [14]. LPS can translocate into the bloodstream across the epithelial barrier of the gastrointestinal tract [15] [16] [17], triggering inflammatory responses in cows [15] [18] [19].
LPS may also induce harmful epigenetic changes in the mammary tissue of dairy cows. However, the association between LPS and epigenetic changes in the mammary tissue of dairy cows during the feeding of diets high in concentrate has not yet been documented. In addition, epigenetic controls of milk gene expression through histone modification and DNA methylation during the feeding of high-concentrate diets to dairy cows are unknown. In many countries, corn straw is frequently used in the diets of cows due to a lack of quality roughage, requiring an increase in the proportion of concentrate in the diet to meet the nutritional requirements for lactation. We hypothesized that increasing the concentrate proportion in a corn straw-based diet will result in increased entry of endotoxin into the mammary gland and thus elicit epigenetic alterations in the udder, which may exert an adverse effect on milk gene expression. Therefore, the objective of this study was to evaluate the effects of feeding a high-concentrate corn straw (HCS) diet on the epigenetic changes in the mammary tissue of dairy cows in comparison with a low-concentrate corn straw (LCS) diet and a low-concentrate mixed quality forage (LMF) diet.
Materials and Methods
Ethics statement
The study was approved by Animal Ethics Committee of Southwest University, and all experimental procedures and the care of the animals were in strict accordance with the “Guidelines on Ethical Treatment of Experimental Animals (2006, No. 398)” issued by the Ministry of Science and Technology of China. Blood and mammary tissue sampling described in detail below was performed under either general or regional anesthesia to minimize suffering and pain in the animals.
Animals, diets and experimental procedure
Thirty second-parity Chinese Holstein cows in mid-lactation, averaging 543±57 kg of body weight and producing 24.32±3.86 kg milk per day at the onset of the experiment, were randomly assigned to 1 of 3 diets (n = 10 per treatment; Table 1). These diets included: 1) the LMF diet with a concentrate to roughage ratio of 46∶54, containing Chinese wildrye (Aneurolepidium chinense), alfalfa hay, and corn silage. This diet is commonly regarded as an excellent diet for lactating cows and served as a control in the study. 2) the HCS diet with a concentrate to roughage ratio of 65∶35, containing corn straw as the only roughage but otherwise identical (except fiber content) to the LMF diet. 3) the LCS diet with the same concentrate to roughage ratio (46∶54) as the LMF diet, and the LCS diet also contained corn straw as the only roughage material. Diets (the LMF and the HCS diets) were formulated based on the nutrient requirements of dairy cows, as recommended by NRC [20]. The LCS diet was formulated by simply replacing the forages of the LMF diet with corn straw. Diets were mixed and offered as total mixed ration twice daily (0830 and 1730 h). Orts were discarded before the next feeding each day and the amount of feed was adjusted to ensure a 5% feed residual. Cows were milked twice daily at 0800 and 1900 h. The experiment lasted 6 weeks. Cows were housed with free access to water. Cows in the experiment were observed daily for feed ingestion, and the rectal temperature and respiratory rates were also examined each day. The cows showed no clinical signs of infectious disease throughout the entire experimental period. This experiment was conducted at the Inner Mongolia Dairy United Technology Co., Ltd., Hohhot, China.
Table 1. Diet compositions.
| Diet1 | |||
| Item | LMF | HCS | LCS |
| Ingredient (% dry matter) | |||
| Alfalfa hay | 23.4 | 0.0 | 0.0 |
| Corn silage | 26.7 | 0.0 | 0.0 |
| Chinese wildrye | 3.7 | 0.0 | 0.0 |
| Corn straw | 0.0 | 35.0 | 53.8 |
| Corn | 24.6 | 35.3 | 24.6 |
| Soybean meal | 14.8 | 20.8 | 14.8 |
| Whole cottonseed | 5.1 | 7.2 | 5.1 |
| Dicalcium phosphate | 0.6 | 0.8 | 0.6 |
| Salt | 0.5 | 0.5 | 0.5 |
| Premix2 | 0.6 | 0.6 | 0.6 |
| Concentrate∶Roughage | 46∶54 | 65∶35 | 46∶54 |
| Nutrient composition (%) of dry matter) | |||
| Net Energy (Mcal/kg) | 1.50 | 1.54 | 1.40 |
| Crude protein | 16.8 | 16.9 | 13.9 |
| Neutral detergent fiber | 37.6 | 34.4 | 46.0 |
| Acid detergent fiber | 23.9 | 19.9 | 26.7 |
| Non-fiber carbohydrate3 | 37.0 | 41.3 | 32.6 |
| Ether extract | 3.4 | 3.2 | 2.5 |
| Calcium | 0.5 | 0.5 | 0.4 |
| Phosphorus | 0.4 | 0.4 | 0.4 |
LMF, low-concentrate mixed forage diet; HCS, high-concentrate corn straw diet; LCS, low-concentrate corn straw diet.
Premix contains: 2,142 mg/kg Cu (as sulfate); 15,428 mg/kg Mn (as sulfate); 15,428 mg/kg Zn (as sulfate); 28 mg/kg Co (as chloride); 231 mg/kg I (as iodate); 57 mg/kg Se (as selenite); 2,285,000 IU/kg vitamin A; 457,000 IU/kg vitamin D; and 11,400 mg/kg vitamin E.
Non-fiber carbohydrate = 100−(% Neutral detergent fiber+% Crude protein+% Ether extract+% Ash).
Blood sampling and analysis
Blood samples were obtained shortly before the morning feeding from the mammary (external pudendal) artery and the mammary vein, respectively, on day 6 and 7 of the last week of the animal trial. Cows were restrained in a standing posture during blood sampling. To keep cows sedated, 20 mL procaine hydrochloride was injected into the muscles near the artery blood collection point, located between the femoral and hip joints. The operator put one hand into the cow's rectum against the external pudendal artery and used the other hand to hold a 18#, 20 cm long sterile needle which was vertically penetrated into the fossa formed through convergence of the internal abdominal oblique muscle, the vastus lateralis muscle and the gluteus medius muscle, after shearing and disinfection. Blood samples were collected in 10-mL vacuum tubes containing heparin sodium anticoagulant and were stored on ice and centrifuged (Feige TDL-40B, Shanghai Anting Scientific Instrument Factory) within 10 min at 3,000× g and 4°C to harvest plasma. The plasma was immediately stored at −20°C until being analyzed for LPS concentrations. The concentrations of LPS in plasma were determined by chromogenic end-point assay using the limulus amebocyte lysate (LAL) test reagent kit (QCL-1000, Lonza Group Ltd., Basel, Switzerland) with a minimum detection limit of 0.1 EU/mL. Samples were initially treated as described by Khafipour et al. [15] to inactivate inhibitory factors in plasma. Then a metallo-modified polyanionic dispersant named Pyrosperse (Lonza Group Ltd.) was added to the samples at a ratio of 1/200 (vol/vol) before LAL testing. The LAL assay was performed by dispensing 50 µL of sample into a pyrogen-free glass tube in a 37°C water bath and mixing with 50 µL LAL reagent. After 10 min, 100 µL substrate solution prewarmed to 37°C was added. After 16 min, stop reagent and azoic reagent (Xiamen Limulus Experiment Factory, Xiamen, China) were added. Optical density at 545 nm was measured using a microplate reader (Synergy H4, BioTek, Winooski, VT).
Mammary tissue sampling and analysis
Mammary tissue samples were obtained by a punch biopsy according to Baumgard et al. [21] with modifications. Cows were restrained from movement under general anesthesia. The mammary gland was exposed and 20 mL procaine hydrochloride was administered in a circular pattern surrounding the incision site. A 5–6 cm incision in the skin on the midpoint of the rear quarter of the right mammary gland was made after shearing and disinfection. Connective tissue was dissected away to reveal the gland capsule and mammary tissue biopsy (∼500 mg) was obtained and divided in half. One half of the sample was used in histone H3 acetylation assays while the other was used in DNA methylation assays. All samples were immediately stored in liquid nitrogen. After mammary tissue sampling was performed, the gland capsule, connective tissue and skin were sutured, and the animals were given intramuscular penicillin to prevent infections.
Total histones were extracted using the EpiQuik™ Total Histone Extraction Kit (Epigentek Group Inc.) according to the manufacturer's protocols. Histone H3 acetylation was quantified using the EpiSeeker Histone H3 Acetylation Assay Kit (Abcam). Histone H3 acetylation sites were identified by resolving histone H3 using sodium dodecyl sulfate-polyacrylamide gel electrophoresis and staining with Coomassie brilliant blue. The histone H3 band was excised and destained with 50% acetonitrile+25 mM ammonium bicarbonate. Histone H3 was digested with trypsin and peptides were analyzed using liquid chromatography-mass spectrometry (MS)×MS (Thermo Scientific Inc.). Peptide mass spectra were measured and analyzed to determine acetylation sites using Mascot software, with special attention paid to the detection of ions with a mass-to-charge ratio (m/z) of 126.1, which is diagnostic for lysine acetylation [22]. Lysine-acetylated peptides were also confirmed by detecting mass differences of 170 Da between adjacent b-ions or y-ions.
Total DNA from the mammary tissue was extracted using a reagent kit (Tiangen, Beijing, China) and used in DNA methylation assays of specific genes via bisulfite sequencing PCR (BSP). DNA was first treated with bisulfate to convert unmethylated cytosines to uracils through deamination while methylated cytosines remained unchanged during the treatment. Uracils were converted to thymines during PCR. PCR primers were designed using Methyl Primer Express v1.0 (Table 2). PCR products were cloned and sequenced to determine the DNA methylation levels of specific genes using BiQ Analyzer software.
Table 2. Primers used for gene methylation studies.
| Primer name1 | Sequence (5′-3′) |
| ACSL1-f | TAGGTATTGTGGTGAAATYGTA |
| ACSL1-r | CCTAAATTTCACAAAAAATCC |
| FASN-f | GTGGTTTTAGGAGATAGTAAGGGT |
| FASN-r | TAACTTAAACAAAAAAATCTCCCT |
| SCD-f | GTTGGTATGGGTATAGGGATA |
| SCD-r | CAACCCAAAACTAACCCC |
| S6K1-f | TAAGGAAATYGAGGTTTTGA |
| S6K1-r | AAAAAAAAAACCACAACAAATCT |
| STAT5A-f | TTTAGGGTTTGAAATATTTGATT |
| STAT5A-r | TCAATTTTCTCATCTATCAAAAA |
ACSL1, acyl-CoA synthetase long-chain family member 1; FASN, fatty acid synthase; SCD, stearoyl-coenzyme A desaturase; S6K1, ribosomal protein S6 kinase 1; STAT5A, signal transducer and activator of transcription 5A.
Statistical analyses
The general linear model (GLM) of SPSS (v.18) was used in the data analysis. The GLM included the random cow effect and the fixed effect of diets, and mean differences for all variables were separated and compared using Duncan's multiple comparison procedure. Data are presented as means ± standard deviation. Significance was declared at p<0.05. Statistical correlation was performed using GraphPad PRISM 5.0, and standard error, p-value, and R2 were computed and used to evaluate the goodness of fit.
Results
We were interested in determining the impact of feeding the HCS diet on the epigenetic changes in the mammary tissue of dairy cows in comparison with the LCS diet and the LMF diet. We first analyzed the effect of these diets on the amount of histone H3 acetylation in mammary tissues. Extraction of histones and quantification of histone H3 acetylation revealed that acetylation was significantly reduced in cows fed the HCS diet, as compared with cows fed the LCS diet ( figure 1 ). These data suggest that the high-concentrate diet may induce epigenetic changes in the mammary tissues.
Figure 1. Effect of different diets on histone H3 acetylation levels in the mammary gland of dairy cows.
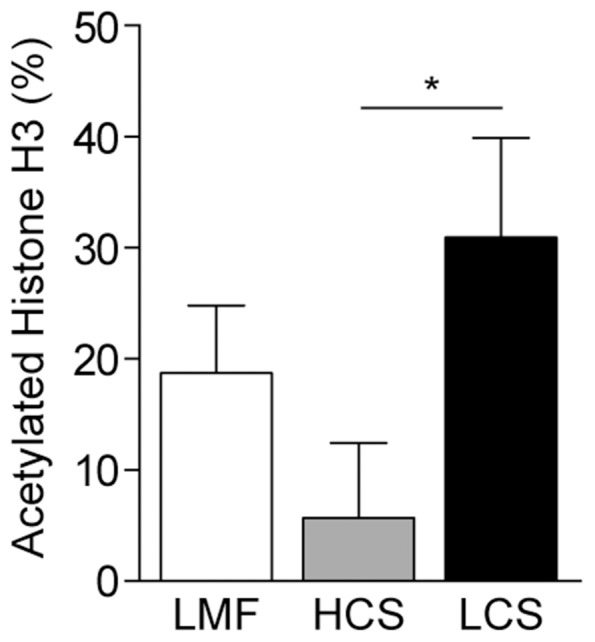
The extent of histone H3 acetylation in the mammary glands of dairy cows was quantified using the EpiSeeker Histone H3 Acetylation Assay Kit. Data represent the mean and standard deviation (n = 10/group) and the asterisk indicates statistical difference (p<0.05) between the indicated columns. LMF, low-concentrate mixed forage diet; HCS, high-concentrate corn straw diet; LCS, low-concentrate corn straw diet.
To validate these data, we further examined histone H3 acetylation by identifying specific acetylated peptides using mass spectrometry. Specific peaks diagnostic of lysine acetylation were identified ( figure 2 ). Histone H3 acetylation occurred on lysines 5 and 7 (Table 3).
Figure 2. Mass spectra of histone H3 lysine-acetylated peptides identified using mass spectrometry.
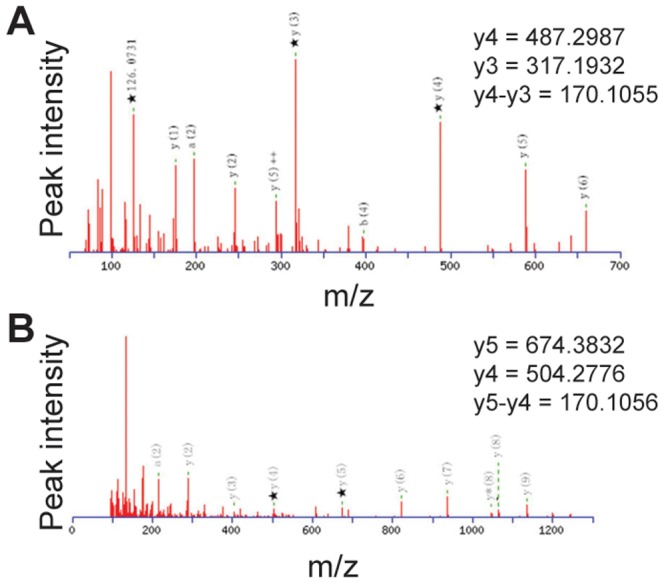
A. MS/MS spectra from m/z ion 882.4923. B. MS/MS spectra from m/z ion 1376.6936.
Table 3. Histone H3 acetylation sites identified in the mammary tissue of dairy cows.
| Diet1 | Molecular weight | Peptide site | Peptide sequence2 | Acetylated site3 |
| LMF | 882.4923 | 20-27 | K.QLATKA*AR.K | K5 |
| 882.4923 | 20-27 | K.QLATKA*AR.K | K5 | |
| 1376.6936 | 74-84 | R.EIAQDFKT*DLR.F | K7 | |
| HCS | 882.4923 | 20-27 | K.QLATKA*AR.K | K5 |
| 1376.6936 | 74-84 | R.EIAQDFKT*DLR.F | K7 | |
| LCS | 882.4923 | 20-27 | K.QLATKA*AR.K | K5 |
| 882.4923 | 20-27 | K.QLATKA*AR.K | K5 | |
| 899.5188 | 20-27 | K.QLATKA*AR.K | K5 | |
| 1376.6936 | 74-84 | R.EIAQDFKT*DLR.F | K7 |
LMF, low-concentrate mixed forage diet; HCS, high-concentrate corn straw diet; LCS, low-concentrate corn straw diet.
The asterisk denotes the acetylated site.
K represents lysine.
A hallmark of feeding high-concentrate diets to cattle is the development of SARA [8] [16] [17], often resulting in the accumulation of LPS in the bloodstream, with subsequent inflammation [15]. To determine if the different diets affected LPS release, we quantified LPS in mammary artery and vein plasma samples. Cows fed the HCS diet had significantly higher LPS concentrations in the blood, as compared with cows fed the LCS diet ( figure 3 ).
Figure 3. Effect of different diets on LPS concentrations in the mammary artery and vein plasma of dairy cows.

LPS concentrations were quantified by chromogenic end-point assay using the limulus amebocyte lysate (LAL) test reagent kit. Data represent the mean and standard deviation (n = 10/group) and the asterisk indicates statistical difference (p<0.05) between the indicated columns. LMF, low-concentrate mixed forage diet; HCS, high-concentrate corn straw diet; LCS, low-concentrate corn straw diet.
We next determined whether LPS concentrations in the mammary blood were correlated with differing levels of histone H3 acetylation in the mammary tissue. We found that the extent of histone H3 acetylation was negatively correlated with LPS concentrations ( figure 4 ).
Figure 4. Relationship between mammary artery plasma LPS contents and histone H3 acetylation levels.

The extent of histone H3 acetylation in mammary tissue samples was plotted vs. the LPS concentrations of mammary artery plasma samples.
The extent of DNA methylation of specific genes involved in fat and protein metabolism in the mammary gland was assessed using BSP. The methylation of the SCD gene encoding a stearoyl-coenzyme A desaturase associated with milk fat synthesis was increased in cows fed the HCS diet, as compared with cows fed either the LCS or LMF diets ( figure 5 ). By contrast, methylation of STAT5A, encoding the signal transducer and activator of transcription 5A involved in protein synthesis, was reduced in cows fed the HCS diet ( figure 5 ).
Figure 5. Effects of different diets on gene methylation levels in the mammary tissue of dairy cows.

Gene methylation was analyzed using bisulfite sequencing PCR. Data represent the mean and standard deviation (n = 10/group) and the asterisk indicates statistical difference (p<0.05) between the indicated columns. ACSL1, acyl-CoA synthetase long-chain family member 1; FASN, fatty acid synthase; SCD, stearoyl-coenzyme A desaturase; S6K1, ribosomal protein S6 kinase 1; STAT5A, signal transducer and activator of transcription 5A. LMF, low-concentrate mixed forage diet; HCS, high-concentrate corn straw diet; LCS, low-concentrate corn straw diet.
Discussion
Histone H3 acetylation regulates various cellular physiological processes, including transcription, chromatin assembly, DNA replication and repair, and cell proliferation. Histone acetylation is associated with an “open” chromatin conformation that facilitates transcription [23] [24] [25]. The acetylation of histones can induce chromatin to adopt a more relaxed structure, thereby modulating the accessibility of DNA [26] [27]. We found that feeding cows the HCS diet decreased histone H3 acetylation as compared to feeding either the LCS diet or the LMF diet. In an earlier study of ours [28], we found that cows fed the HCS diet had identical dietary nutritional levels as compared with cows fed the LMF diet, but that various milk production parameters were negatively impacted by the HCS diet. Feeding HCS diets may affect milk gene expression so as to affect the milk component synthesis.
Our study showed feeding the HCS diet increased LPS concentrations in both mammary artery and vein plasma compared with cows fed the LCS diet. We also found that histone H3 acetylation levels were negatively correlated with LPS concentrations in mammary blood. It was shown previously that the LPS concentrations in both ruminal contents and blood increased after 21% dry matter in the control diet with a concentrate to roughage ratio of 50∶50 was replaced with a grain mixture comprised of 50% ground wheat and 50% ground barley [15]. The presence of LPS in the blood of dairy cows and goats fed diets containing a high proportion of concentrate was also reported in several studies [29] [30] [31]. Therefore, the lower histone H3 acetylation in the mammary tissue of cows fed the HCS diet can be attributed to the increased entry of LPS into the mammary tissue. Since there are few reports about the relationship of LPS and histone H3 acetylation in the mammary tissue of dairy cows to date, further studies are warranted to confirm the LPS-mediated histone hypoacetylation in the mammary tissue of dairy cows. Also unknown is the relationship between histone H3 acetylation and the physiological function of the mammary tissue in dairy cows.
DNA methylation is the oldest epigenetic mechanism that is known to correlate with gene repression [32]. The methylation status of DNA in promoter regions of specific genes efficiently regulates the transcription of the corresponding gene. High methylation tends to decrease the access of specific transcriptional factors to the promoter region of genes, whereas low or absent methylation results in an increased accessibility and transcriptional activity [33] [34]. For instance, a lower level of DNA methylation, measured as a percentage of 5′ methyldeoxycytidine, in the mammary tissue of Holstein cows was correlated with an increase in β-casein mRNA [35]. We found that feeding the HCS diet increased the methylation of genes (acyl-CoA synthetase long-chain family member 1, ACSL1; fatty acid synthase, FASN; and stearoyl-coenzyme A desaturase, SCD) associated with milk fat synthesis but decreased the methylation of genes (ribosomal protein S6 kinase 1, S6K1; and signal transducer and activator of transcription 5A, STAT5A) involved in protein synthesis as compared with feeding the LCS and LMF diets. We speculate that feeding the HCS diet could reduce milk fat synthesis but enhance protein synthesis in the udder. In support of this idea, the milk fat content for cows fed the HCS in this study was 3.58%, whereas the milk fat contents of cows fed the LCS and LMF diets were 3.95 and 4.23%, respectively. It has also been shown that when dairy cows were fed diets containing different proportions of barley grain, milk fat content was negatively correlated with barley content [14]. Several studies also demonstrated that both milk fat percentage and milk fat yield decreased during grain-induced SARA [15] [36] [37]. By contrast, SARA induction increases milk protein content [15] [38]. However, this increase in milk protein content may not necessarily reflect the rise of casein concentrations in milk, and in fact, casein content decreased after intramammary challenge with LPS [39]. The increase in milk protein is instead thought to be due to the rise of immune proteins such as acute phase proteins, albumin, β-lactoglobulin, lactoferrin, and antimicrobial peptides [40] [41] [42] [43].
Collectively, feeding a high-concentrate corn straw diet decreased histone H3 acetylation in the mammary tissue of dairy cows, primarily on K5 residues. Histone H3 acetylation levels were negatively correlated with LPS contents in the mammary plasma. LPS may play an epigenetic regulatory role in the mammary tissue of dairy cows. Feeding a high-concentrate corn straw diet tended to increase the methylation of genes associated with fat synthesis but decrease the methylation of genes associated with protein synthesis.
Acknowledgments
We gratefully thank the teachers and students of the “Dairy 973” team of Inner Mongolia Agricultural University for their assistance in conducting this experiment. We also acknowledge Professor Yongju Zhang and Dr. Caode Jiang of the animal genetics and breeding group of Southwest University for their advice in designing this study and writing this paper.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
The study was supported by funds from the National Key Basic Research Program of China (No. 2011CB100803). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Feinberg AP (2007) Phenotypic plasticity and the epigenetics of human disease. Nature 447: 433–440. [DOI] [PubMed] [Google Scholar]
- 2. Shilatifard A (2006) Chromatin modifications by methylation and ubiquitination: implications in the regulation of gene expression. Annu Rev Biochem 75: 243–269. [DOI] [PubMed] [Google Scholar]
- 3.Li E, Bird A (2007) DNA methylation in mammals. In: Allis CD, Jenuwein T, Reinberg D, Caparros, ML, editors. Epigenetics. New York: Cold Spring Harbor Laboratory Press. p. 341–356.
- 4. Singh K, Erdman RA, Swanson KM, Molenaar AJ, Maqbool NJ, et al. (2010) Epigenetic regulation of milk production in dairy cows. J Mammary Gland Biol Neoplasia 15: 101–112. [DOI] [PubMed] [Google Scholar]
- 5. Singh K, Molenaar AJ, Swanson KM, Gudex B, Arias JA, et al. (2002) Epigenetics: a possible role in acute and transgenerational regulation of dairy cow milk production. Animal 6: 375–381. [DOI] [PubMed] [Google Scholar]
- 6. Nocek JE (1997) Bovine acidosis: implications on laminitis. J Dairy Sci 80: 1005–1028. [DOI] [PubMed] [Google Scholar]
- 7. Ametaj BN, Emmanuel DGV, Zebeli Q, Dunn SM (2009) Feeding high proportions of barley grain in a total mixed ration perturbs diurnal patterns of plasma metabolites in lactating dairy cows. J Dairy Sci 92: 1084–1091. [DOI] [PubMed] [Google Scholar]
- 8. Dong GZ, Liu SM, Wu YX, Lei CL, Zhou J, et al. (2011) Diet-induced bacterial immunogens in the gastrointestinal tract of dairy cows: Impacts on immunity and metabolism. Acta Vet Scand 53: 48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Emmanuel DGV, Madsen KL, Churchill TA, Dunn SM, Ametaj BN (2007) Acidosis and lipopolysaccharide from Escherichia coli B:055 cause hyperpermeability of rumen and colon tissues. J Dairy Sci 90: 5552–5557. [DOI] [PubMed] [Google Scholar]
- 10. Iqbal S, Zebeli Q, Mazzolari A, Bertoni G, Dunn SM, et al. (2009) Feeding barley grain steeped in lactic acid modulates rumen fermentation patterns and increases milk fat content in dairy cows. J Dairy Sci 92: 6023–6032. [DOI] [PubMed] [Google Scholar]
- 11. Keunen JE, Plaizier JC, Kyriazakis L, Duffield TF, Widowski TM, et al. (2002) Effects of a subacute ruminal acidosis model on the diet selection of dairy cows. J Dairy Sci 85: 3304–3313. [DOI] [PubMed] [Google Scholar]
- 12. Zebeli Q, Dunn SM, Ametaj BN (2011) Perturbations of plasma metabolites correlated with the rise of rumen endotoxin in dairy cows fed diets rich in easily degradable carbohydrates. J Dairy Sci 94: 2374–2382. [DOI] [PubMed] [Google Scholar]
- 13. Ametaj BN, Bradford BJ, Bobe G, Nafikov RA, Lu Y, et al. (2005) Strong relationships between mediators of the acute phase response and fatty liver in dairy cows. Can J Anim Sci 85: 165–175. [Google Scholar]
- 14. Zebeli Q, Ametaj BN (2009) Relationships between rumen lipopolysaccharide and mediators of inflammatory response with milk fat production and efficiency in dairy cows. J Dairy Sci 92: 3800–3809. [DOI] [PubMed] [Google Scholar]
- 15. Khafipour E, Krause DO, Plaizier JC (2009) A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J Dairy Sci 92: 1060–1070. [DOI] [PubMed] [Google Scholar]
- 16. Plaizier JC, Khafipour E, Li S, Gozho GN, Krause DO (2012) Subacute ruminal acidosis (SARA), endotoxins and health consequences. Anim Feed Sci Tech 72: 9–21. [Google Scholar]
- 17. Plaizier JC, Krause DO, Gozho GN, McBride BW (2009) Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet J 76: 21–31. [DOI] [PubMed] [Google Scholar]
- 18. Emmanuel DG, Dunn SM, Ametaj BN (2008) Feeding high proportions of barley grain stimulates an inflammatory response in dairy cows. J Dairy Sci 91: 606–614. [DOI] [PubMed] [Google Scholar]
- 19. Ametaj BN, Koenig KM, Dunn SM, Yang WZ, Zebeli Q, et al. (2009) Backgrounding and finishing diets are associated with inflammatory responses in feedlot steers. J Dairy Sci 87: 1314–1320. [DOI] [PubMed] [Google Scholar]
- 20.NRC (2001) Nutrient Requirements of Dairy Cattle. Washington D.C.: National Academy Press.
- 21. Baumgard LH, Matitashvili E, Corl BA, Dwyer DA, Bauman DE (2002) trans-10, cis-12 Conjugated linoleic acid decreases lipogenic rates and expression of genes Involved in milk lipid synthesis in dairy cows. J Dairy Sci 85: 2155–2163. [DOI] [PubMed] [Google Scholar]
- 22. Kim JY, Kim KW, Kwon HJ, Lee DW, Yoo JS (2002) Probing lysine acetylation with a modification-specific marker ion using high-performance liquid chromatography/electrospray-mass spectrometry with collision-induced dissociation. Anal Chem 74: 5443–5449. [DOI] [PubMed] [Google Scholar]
- 23. Campos EI, Reinberg D (2009) Histones: annotating chromatin. Annu Rev Genet 43: 559–599. [DOI] [PubMed] [Google Scholar]
- 24. Cheng X, Blumenthal RM (2010) Coordinated chromatin control: structural and functional linkage of DNA and histone methylation. Biochemistry 49: 2999–3008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Ramsey SA, Knijnenburg TA, Kennedy KA, Zak DE, Gilchrist M, et al. (2010) Genome-wide histone acetylation data improve prediction of mammalian transcription factor binding sites. Bioinformatics 26: 2071–2075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Horn PJ, Peterson CL (2002) Molecular biology. Chromatin higher order folding–wrapping up transcription. Science 297: 1824–1827. [DOI] [PubMed] [Google Scholar]
- 27. Tse C, Sera T, Wolffe AP, Hansen JC (1998) Disruption of higher-order folding by core histone acetylation dramatically enhances transcription of nucleosomal arrays by RNA polymerase III. Mol Cell Biol 18: 4629–4638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Zhou J, Dong GZ, Ao CJ, Zhang S, Qiu M, et al. (2014) Feeding a high-concentrate corn straw diet increased the release of endotoxin in the rumen and pro-inflammatory cytokines in the mammary gland of dairy cows. BMC Vet Res 10: 172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Dong H, Wang S, Jia Y, Ni Y, Zhang Y, et al. (2013) Long-term effects of subacute ruminal acidosis (SARA) on milk quality and hepatic gene expression in lactating goats fed a high-concentrate diet. PLoS One 8: e82850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Huo W, Zhu W, Mao S (2013) Effects of Feeding increasing proportions of corn grain on concentration of lipopolysaccharide in the rumen fluid and the subsequent alterations in immune responses in goats. Asian Australas J Anim Sci 10: 1437–1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Zhang RY, Yoon I, Zhu WY, Mao SY (2013) Effect of Saccharomyces cerevisiae fermentation product on lactation performance and lipopolysaccharide concentration of dairy cows. Asian Australas J Anim Sci 26: 1137–1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Raizin A, Riggs AD (1980) DNA methylation and gene function. Science 210: 604–610. [DOI] [PubMed] [Google Scholar]
- 33. Ollikainen M, Smith KR, Joo EJ, Ng HK, Andronikos R, et al. (2010) DNA methylation analysis of multiple tissues from newborn twins reveals both genetic and intrauterine components to variation in the human neonatal epigenome. Hum Mol Genet 19: 4176–4188. [DOI] [PubMed] [Google Scholar]
- 34. McKay JA, Mathers JC (2011) Diet induced epigenetic changes and their implications for health. Acta Physiol (Oxf) 202: 103–118. [DOI] [PubMed] [Google Scholar]
- 35. Choi YJ, Jang K, Yim DS, Baik MG, Myung KH, et al. (1998) Effects of compensatory growth on the expression of milk protein gene and biochemical changes of the mammary gland in Holstein cows. J Nutr Biochem 9: 380–387. [Google Scholar]
- 36. Gozho GN, Krause DO, Plaizier JC (2007) Rumen lipopolysaccharide concentration and inflammatory responses during grain-induced subacute ruminal acidosis in dairy cows. J Dairy Sci 90: 856–866. [DOI] [PubMed] [Google Scholar]
- 37. Li S, Gozho GN, Gakhar N, Khafipour E, Krause DO, et al. (2012) Evaluation of diagnostic measures for subacute ruminal acidosis in dairy cows. Can J Anim Sci 92: 353–364. [Google Scholar]
- 38. Colman E, Khafipour E, Vlaeminck B, De Baets B, Plaizier JC, et al. (2013) Grain-based versus alfalfa-based subacute ruminal acidosis induction experiments: Similarities and differences between changes in milk fatty acids. J Dairy Sci 96: 4100–4111. [DOI] [PubMed] [Google Scholar]
- 39. Hinz K, Larsen LB, Wellnitz O, Bruckmaier RM, Kelly AL (2012) Proteolytic and proteomic changes in milk at quarter level following infusion with Escherichia coli lipopolysaccharide. J Dairy Sci 95: 1655–1666. [DOI] [PubMed] [Google Scholar]
- 40. Hiss S, Mielenz M, Bruckmaier M, Sauerwein H (2004) Haptoglobin concentrations in blood and milk after endotoxin challenge and quantification of mammary Hp mRNA expression. J Dairy Sci 87: 3778–3784. [DOI] [PubMed] [Google Scholar]
- 41. Schmitz S, Pfaffl MW, Meyer HHD, Bruckmaier RM (2004) Short-term changes of mRNA expression of various inflammatory factors and milk proteins in mammary tissue during LPS-induced mastitis. Domest Anim Endocrin 26: 111–126. [DOI] [PubMed] [Google Scholar]
- 42. Shamay A, Homan R, Fuerman Y, Levin I, Barash H, et al. (2005) Expression of albumin in nonhepatic tissues and its synthesis by the bovine mammary gland. J Dairy Sci 88: 569–576. [DOI] [PubMed] [Google Scholar]
- 43. Isobe N, Morimoto K, Nakamura J, Yamasaki A, Yoshimura Y (2009) Intramammary challenge of lipopolysaccharide stimulates secretion of lingual antimicrobial peptide into milk of dairy cows. J Dairy Sci 92: 6046–6051. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.