

Abstract
Background
Members of the GATA transcription factor family have been used in many transfection studies to investigate their roles in the regulation of gene expression. To correct for variations in transfection efficiency, the Renilla luciferase encoding plasmids pRL-TK and pRL-SV40 are commonly used as normalization controls.
Results
We report here that plasmids expressing GATA-4 or GATA-6 transcription factor increased Renilla luciferase gene expression by 2 to 8 fold when co-transfected with pRL-TK or pRL-SV40. This alteration of the control reporter gene activity was shown to cause erroneous normalization of transfection efficiency and thus misinterpretation of results in a transactivation assay. To circumvent the problem, we generated two mutant control plasmids from which putative GATA response elements were deleted. These deletions rendered pRL-SV40 unresponsive to GATA transcription factor stimulation and reduced the response of pRL-TK. A database search also indicates that consensus GATA binding sequences are present in other commercially available Renilla luciferase encoding plasmids; therefore, the latter can potentially be transactivated by GATA transcription factors.
Conclusion
Taken together, these findings highlight the importance of the selection of an appropriate control reporter plasmid for the normalization of transfection efficiency.
Background
In transient transfection experiments, a promoter sequence of interest coupled to a reporter gene is often co-transfected with a control plasmid whose expression is constitutive in order to normalize for transfection efficiency or cell viability. Reporter plasmids which contain cDNA encoding the Renilla (Renilla reniformis) luciferase enzyme (Rluc) have become increasingly popular as controls for such a purpose. Rluc is not intrinsically expressed in mammalian cells and can function as an enzyme immediately following translation because post-translational modification is not required for its activity. Also, like firefly (Photinus pyralis) luciferase, Renilla luciferase activity can be assayed with high sensitivity and linearity over several orders of magnitude. In the widely used Dual Luciferase Reporter (DLR) assay system (Promega, Madison, WI), activities of firefly and Renilla luciferases are measured sequentially from a single sample based on the different substrate specificities of the two luciferases, thus minimizing errors due to variability in pipetting volumes and providing a convenient means to measure both the experimental and control reporter activities.
One common assumption in the use of a control reporter plasmid is that its expression is unaffected by other co-transfected plasmids or any experimental treatment. However, this assumption is not always justified. The commercially available plasmids pRL-TK and pRL-SV40 (GenBank accession numbers AF025846 and AF025845 respectively; Promega) contain the Rluc gene driven by the constitutively active herpes simplex virus thymidine kinase (HSV-TK) promoter and SV40 early enhancer/promoter respectively (figure 1). Downstream of both promoters is a chimeric intron derived from human beta-globin intron 1 and an intron from the heavy chain variable region of an immunoglobulin gene. This chimeric intron, together with the T7 RNA polymerase promoter immediately upstream of Rluc, enhances the expression of Renilla luciferase.
Figure 1.
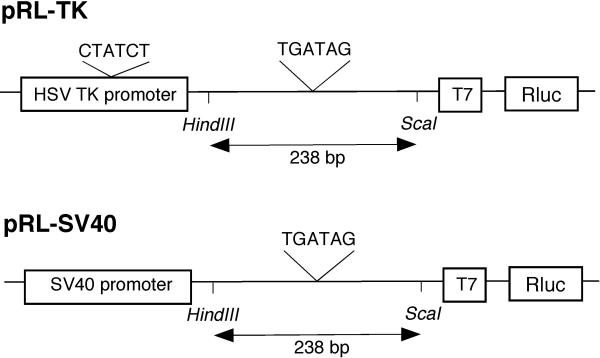
Promoter regions of pRL-TK and pRL-SV40. The Renilla luciferase gene (Rluc) is driven by the herpes simplex virus thymidine kinase (HSV-TK) promoter or SV40 enhancer/early promoter. Consensus GATA binding sequences, (A/T)GATA(A/G), are present in the intron upstream of T7 promoter and within HSV-TK but not SV40 promoter. Deletion mutants of the two control plasmids (pRL-TK-d238 and pRL-SV40-d238) were constructed by removal of 238 base pairs between the HindIII and ScaI restriction sites in the intron.
The family of GATA transcription factors are expressed in various tissues in vertebrates. They can bind to a common consensus DNA sequence, (A/T)GATA(A/G), and are implicated in the regulation of gene expression in a tissue-specific manner. To date, six members of the GATA transcription factor family have been identified and are divided into two subfamilies based on their expression patterns. GATA-1, -2 and -3 are predominantly expressed in hematopoietic stem cells and act as transcriptional regulators of genes involved in hematopoiesis [1]. GATA-4, -5 and -6, on the other hand, are found in tissues including heart, lung, gonads, gut and liver [2]. GATA-4 was shown to activate transcription of genes expressed in the gonads including steroidogenic acute regulatory (StAR) protein and aromatase [3], whereas the two isoforms of GATA-6 can transactivate a GATA-dependent firefly luciferase reporter construct [4]. In the course of our studies on gene expression regulated by several GATA transcription factors, we employed both pRL-TK and pRL-SV40 as control reporter plasmids and observed that Rluc expression by both plasmids was increased by members of the GATA transcription factor family. Here we report that these alterations in control gene expression by GATA transcription factors lead to inaccurate normalization of transfection efficiency, which can be circumvented by using a modified Renilla luciferase expression plasmid.
Results and discussion
Rluc expression by the control plasmids pRL-TK and pRL-SV40 were examined in co-transfection experiments. Transient transfection assays showed that reporter gene activities of the two control plasmids were altered by the GATA transcription factors tested (figure 2). Both isoforms of human GATA-6 transactivated Rluc expression; co-transfection of pRL-TK with hG6-MALT and hG6-MYQ increased luciferase activity by 8.4 ± 1.2 and 5.7 ± 1.7 fold (mean ± s.e.m.), respectively. In comparison, hG6-MALT augmented luciferase activity expressed by pRL-SV40 by 3.1 ± 0.3 fold. This differential transactivation potential by the two GATA-6 encoding plasmids is in agreement with previous findings that the long isoform was more effective than the short isoform in transactivating a GATA-dependent firefly luciferase reporter construct [4]. It is currently unclear whether differential functions of the two human GATA-6 isoforms exist in vivo. However, tissue-specific differential expression of the two isoforms may represent a mechanism by which GATA-6 responsive genes are regulated. Taken together, the above data highlight the importance of testing whether expression levels of a control plasmid can be altered by experimental treatments such as co-expression of a GATA transcription factor in the present study.
Figure 2.
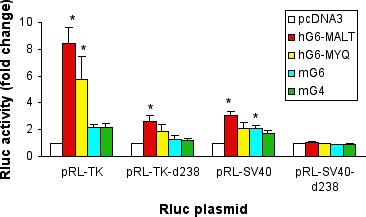
GATA transcription factors transactivate Renilla luciferase gene expression in control plasmids. Various Renilla luciferase (Rluc) reporter plasmids were co-transfected into COS-7 cells with expression plasmids encoding the two isoforms of human GATA-6 (hG6-MALT and hG6-MYQ), mouse GATA-6 (mG6), mouse GATA-4 (mG4) or the empty vector pcDNA3. 48 h after transfection, cells were harvested and lysed. Rluc activities were then measured by the Dual Luciferase Reporter assay system and expressed as fold increase compared with control cells transfected with pcDNA3 (mean ± sem of at least 4 experiments). *P < 0.05.
Given the response of pRL-TK and pRL-SV40 to GATA transcription factors reported above, we speculated that GATA response elements might be present in the two reporter plasmids. Indeed, examination of the promoter region of pRL-TK revealed two putative GATA binding sites upstream of Rluc (figure 1), one within the HSV-TK promoter and the other 91 base pairs (bp) upstream of the T7 promoter. An identical intronic sequence of 246 bp is also present in pRL-SV40. It was thus hypothesized that these putative GATA binding sites could contribute to the increased Rluc gene activities observed. To test if removal of the proximal putative GATA binding site can abrogate the Rluc transactivating effects of GATA transcription factors, 238 bp between the HindIII and ScaI restriction sites within the intronic sequence were deleted to yield mutant reporter plasmids pRL-TK-d238 and pRL-SV40-d238. Figure 2 shows that deletion of the 238 bp rendered the mutant plasmid pRL-SV40-d238 totally unresponsive to GATA-6 or GATA-4 stimulation. In contrast, despite removal of the proximal consensus GATA binding site of pRL-TK-d238, hG6-MALT could still increase its luciferase activity by 2.6 ± 0.4 fold (mean ± s.e.m.). This was possibly due to the remaining consensus GATA binding site present within the HSV-TK promoter (figure 1). Rluc activity expressed by pRL-TK-d238 was unaffected when the mutant plasmid was co-transfected with mouse GATA-6 (mG6) or GATA-4 (mG4).
Normalization of transfection efficiency by a control plasmid that is affected by experimental treatment can lead to misinterpretations of the reporter gene activity of interest. To investigate how GATA transcription factors can affect normalization of transfection efficiency, we co-transfected COS-7 cells with pGL3-CYP11A (a plasmid construct containing a CYP11A promoter region coupled to the firefly luciferase gene) and either pRL-SV40-d238 or pRL-TK. Because the four GATA-expressing plasmids did not affect Rluc activity encoded by pRL-SV40-d238, we use pGL3-CYP11A reporter activity normalized by pRL-SV40-d238 as reference values. The GATA-expressing plasmids increased pGL3-CYP11A firefly luciferase activity by 1.76 to 5.02 fold (values normalized by pRL-SV40-d238 Rluc activity, figure 3). In comparison, because pRL-TK was transactivated by the GATA-expressing plasmids, pGL3-CYP11A firefly luciferase activities became misleadingly low when normalized by this control plasmid (pRL-TK bars, figure 3). For example, an approximately fourfold stimulation of pGL3-CYP11A reporter gene activity by hG6-MALT (normalized by pRL-SV40-d238 Rluc activity) was misinterpreted as 32% inactivation when pRL-TK was used as a normalization plasmid (red bars, figure 3).
Figure 3.
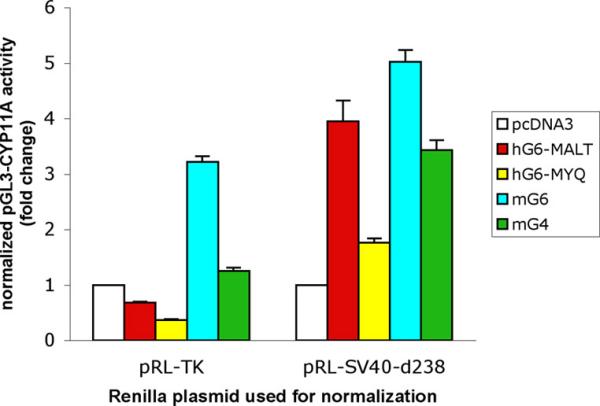
GATA-stimulated pGL3-CYP11A reporter gene activities normalized by 2 different Renilla control plasmids. COS-7 cells were transfected with pGL3-CYP11A, and combinations of GATA-expressing and Renilla luciferase-encoding plasmids. Both firefly and Renilla luciferase activities were measured using the Dual Luciferase Reporter assay system. pGL3-CYP11A firefly luciferase activity is normalized by Renilla luciferase activity and expressed as fold change (mean ± sem) compared with control cells transfected with pcDNA3 and the corresponding Renilla plasmid.
Amino acid sequence of the human full-length GATA-6 is >80% identical to that of the murine transcription factor [4]. Yet, figure 2 shows that pRL-TK was transactivated by human GATA-6 (hG6-MALT, 8.4 fold) to a larger extent than by its murine counterpart (mG6, 2.1 fold), P < 0.05. Mechanisms by which pRL-TK is differentially transactivated by human and mouse GATA-6 are currently unclear. It is possible that elements in pRL-TK other than the putative GATA binding sites or the context of the GATA response element may play a role in augmenting Rluc transactivation by human GATA-6 or diminishing the effects of mouse GATA-6.
Members of the GATA family are not the only transcription factors that can enhance expression of Renilla luciferase by pRL-TK. Other transcription factors including Sp1, Nurr1 and skNAC also induced Rluc activity expressed by pRL-TK [5-7]. Therefore, the two Rluc-encoding vectors reported here cannot be used as normalization controls in all transfection systems because of their inducibility by experimental treatment and potential misinterpretations of reporter gene activity.
To examine if other Rluc expressing plasmids can potentially be transactivated by GATA factors, we conducted a BLAST search of the NCBI database http://www.ncbi.nlm.nih.gov/BLAST and found that the same 246 bp intronic sequence between the T7 promoter and the HSV-TK or SV40 promoter is also present in the synthetic Renilla luciferase reporter plasmids phRL-TK and phRL-SV40 (Promega). Likewise, more than 85% of this sequence including the putative GATA binding site is found in other commercially available Rluc encoding plasmids including pRL-CMV, phRL-CMV, pRL-null and phRL-null (all from Promega). Given the transactivating effects of GATA transcription factors on pRL-TK and pRL-SV40, it is likely that the presence of a consensus GATA binding site close to the Rluc gene in other Rluc expressing plasmids may also render them responsive to transactivation by GATA transcription factors.
Conclusions
We report here that several GATA transcription factors stimulated gene expression by the Renilla luciferase encoding plasmids pRL-TK and pRL-SV40. Also, GATA-induced gene expression could be reduced or ablated by the removal of putative GATA response elements from these control plasmids. These data underscore the importance of monitoring variations in control gene expression due to experimental treatment in transfection studies.
Methods
Plasmid constructs
Expression plasmids encoding the mouse GATA-4 (pcDNA3-mG4) and mouse GATA-6 (pcDNA3-mG6) were generous gifts from Dr. Edward Morrisey, University of Pennsylvania [8]. Construct pMALT-ORF containing the full-length human GATA-6 cDNA was kindly provided by Dr. Christopher Gove (King's College London, UK) [4]. Expression plasmid hG6-MALT was made by subcloning the full-length human GATA-6 cDNA from pMALT-ORF to the BamHI/XhoI sites of pcDNA3 (Invitrogen). Expression plasmid encoding the short isoform of human GATA-6 (hG6-MYQ) was made as follows. Using pMALT-ORF as template, forward primer 5'-ACATTTGGATCCGGAGGAGATGTACCAGAC-3' (underlined sequence being a BamHI restriction site) and reverse primer 5'-ACTTCTCGAGGACTGCTGCACAAAAGCA-3' (underlined sequence being an XhoI restriction site), PCR was carried out to generate PCR product encoding the C-terminal 449 amino acids of the full-length human GATA-6. Using standard molecular techniques, the resultant PCR product was inserted into the BamHI/XhoI sites of pcDNA3 to yield hG6-MYQ. The reporter plasmid construct pGL3-CYP11A containing the promoter region (-1843 to +31) of the cholesterol side-chain cleavage enzyme gene (CYP11A) coupled to a firefly luciferase gene has been previously described [9].
Deletion mutant plasmids pRL-TK-d238 and pRL-SV40-d238 were generated from pRL-TK and pRL-SV40, respectively, as follows. An intronic sequence of 238 base pairs including a putative GATA binding site was deleted by partial digestion of the full-length plasmid with ScaI, followed by HindIII digestion, Klenow reaction to fill in the protruding ends and blunt-end recircularization. Both mutant plasmids were then sequenced to confirm the deletion of 238 base pairs within the intronic sequence.
Transient transfection and reporter gene assays
COS-7 cells, a monkey kidney cell line known to express no GATA-6 transcription factor [8], were maintained in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 5% fetal bovine serum and antibiotics. On day one of experiment, 45,000 cells/well were seeded into 12-well plates in the same medium. 24 h later, 40 ng Rluc-containing reporter plasmid and 80 ng GATA-encoding plasmid (or the empty vector pcDNA3), with or without 500 ng pGL3-CYP11A, were co-transfected into COS-7 cells in triplicates using FuGENE 6 transfection reagent (Roche Diagnostics, Indianapolis, IN) according to the manufacturer's protocol. Cells were lysed with 200 μl passive lysis buffer (Promega) 48 h after transfection. The cell lysates were vortexed and briefly centrifuged to sediment cell debris. A 20 μl aliquot of cell lysate was then assayed for luciferase activities using a Lumat LB9507 luminometer (Berthold) and the Dual Luciferase Reporter (DLR) assay system (Promega), according to the supplier's recommendations.
Statistical analysis
The Tukey-Kramer test was used to detect statistically significant differences (P value < 0.05) between cells transfected with GATA-expressing plasmids and pcDNA3-transfected cells (controls).
Authors' contributions
CKMH made the plasmid constructs and conducted the reporter gene assays. JFS participated in the design and coordination of the study. Both authors read and approved the final manuscript.
Acknowledgments
Acknowledgements
The authors would like to thank Drs. Christopher Gove and Edward Morrisey for their GATA plasmid constructs. We also thank Drs. Jennifer Wood and Zhibing Zhang for useful suggestions. This study was supported by NIH grant number HD34449.
Contributor Information
Clement KM Ho, Email: clement.ho@doctors.org.uk.
Jerome F Strauss, III, Email: jfs3@mail.med.upenn.edu.
References
- Orkin SH. Embryonic stem cells and transgenic mice in the study of hematopoiesis. Int J Dev Biol. 1998;42:927–934. [PubMed] [Google Scholar]
- Molkentin JD. The zinc finger-containing transcription factors GATA-4, -5, and -6. Ubiquitously expressed regulators of tissue-specific gene expression. J Biol Chem. 2000;275:38949–38952. doi: 10.1074/jbc.R000029200. [DOI] [PubMed] [Google Scholar]
- Tremblay JJ, Viger RS. GATA factors differentially activate multiple gonadal promoters through conserved GATA regulatory elements. Endocrinology. 2001;142:977–986. doi: 10.1210/en.142.3.977. [DOI] [PubMed] [Google Scholar]
- Brewer Alison, Gove Christopher, Davies Andy, McNulty Claire, Barrow Dalna, Koutsourakis Manoussos, Farzaneh Farzin, Pizzey John, Bomford Adrian, Patient Roger. The Human and Mouse GATA-6 Genes Utilize Two Promoters and Two Initiation Codons. J Biol Chem. 1999;274:38004–38016. doi: 10.1074/jbc.274.53.38004. [DOI] [PubMed] [Google Scholar]
- Osborne SA, Tonissen KF. pRL-TK induction can cause misinterpretation of gene promoter activity. Biotechniques. 2002;33:1240–1242. doi: 10.2144/02336bm12. [DOI] [PubMed] [Google Scholar]
- Matuszyk J, Ziolo E, Cebrat M, Kochel I, Strzadala L. Nurr1 affects pRL-TK but not phRG-B internal control plasmid in genetic reporter system. Biochem Biophys Res Commun. 2002;294:1036–1039. doi: 10.1016/S0006-291X(02)00601-0. [DOI] [PubMed] [Google Scholar]
- Sims R. J., 3rd, Liss AS, Gottlieb PD. Normalization of luciferase reporter assays under conditions that alter internal controls. Biotechniques. 2003;34:938–940. doi: 10.2144/03345bm07. [DOI] [PubMed] [Google Scholar]
- Morrisey EE, Ip HS, Lu MM, Parmacek MS. GATA-6: a zinc finger transcription factor that is expressed in multiple cell lineages derived from lateral mesoderm. Dev Biol. 1996;177:309–322. doi: 10.1006/dbio.1996.0165. [DOI] [PubMed] [Google Scholar]
- Wood JR, Nelson VL, Ho C, Jansen E, Wang CY, Urbanek M, McAllister JM, Mosselman S, Strauss J. F., 3rd. The molecular phenotype of polycystic ovary syndrome (PCOS) theca cells and new candidate PCOS genes defined by microarray analysis. J Biol Chem. 2003;278:26380–26390. doi: 10.1074/jbc.M300688200. [DOI] [PubMed] [Google Scholar]