

Abstract
Purpose
Hyperosmolarity has been recognized as a proinflammatory stress in the pathogenesis of dry eye disease. This study investigated the suppressive effect of osmoprotectants (L-carnitine, erythritol, and betaine) on the production and activity of matrix metalloproteinases (MMPs) in primary human corneal epithelial cells (HCECs) exposed to hyperosmotic stress.
Methods
Primary HCECs were established from fresh donor limbal tissue explants. The cultures in iso-osmolar medium (312 mOsM) were switched to hyperosmotic media with or without prior incubation with different concentrations of L-carnitine, erythritol, or betaine (2, 10, or 20 mM). The mRNA expression of the MMPs was determined with reverse transcription and quantitative real-time PCR (RT-qPCR). Protein production and activity were evaluated with immunofluorescent staining and gelatin zymography.
Results
Hyperosmotic media (400, 450, or 500 mOsM) significantly stimulated mRNA expression of collagenase MMP-13, gelatinases MMP-9 and MMP-2, stromelysin MMP-3, and matrilysin MMP-7, mostly in an osmolarity-dependent fashion. The stimulated mRNA expression and protein production of these MMPs were significantly but differentially suppressed by L-carnitine, erythritol, or betaine, as evaluated with RT-qPCR and immunofluorescent staining. Interestingly, these osmoprotectants not only suppressed production but also inhibited activation of MMP-9 and MMP-2, as evaluated with gelatin zymography.
Conclusions
Our findings for the first time demonstrate that osmoprotectants, L-carnitine, erythritol, and betaine, suppress the gene expression, protein production, and enzymatic activity of MMPs in HCECs exposed to hyperosmotic stress. L-carnitine appears to have the broadest and strongest suppressive effect on these MMPs. These osmoprotectants may have potential effects in protecting ocular surface epithelia from MMP-mediated disorders in dry eye disease.
Introduction
Dry eye is often accompanied by increased osmolarity of the tear film and inflammation of the ocular surface. Reduced aqueous tear flow and/or increased evaporation of the aqueous tear phase leads to tear hyperosmolarity, a key step in the vicious circle of dry eye disease pathology. Previous studies including ours have shown that hyperosmolarity is a potent proinflammatory stress, which induces inflammation [1,2], promotes apoptosis [3-6], and stimulates the expression and activity of matrix metalloproteinases (MMPs) [7,8] in ocular surface epithelium.
To date, 24 genes of MMPs, including a gene duplication, have been identified as expressed in humans, which encode 23 unique MMP proteins [9]. According to their substrate specificity, they can be categorized into six subclasses: collagenases (MMP-1, MMP-8, MMP-13), gelatinases (MMP-2, MMP-9), stromelysins (MMP-3, MMP-10, MMP-11), matrilysins (MMP-7, MMP-26), membrane-type MMPs, and others. MMPs are a family of neutral zinc proteases that degrade extracellular components including the triple-helical structure of collagen. They are synthesized and secreted by multiple cell types, including corneal epithelial cells and fibroblasts. Generally, MMPs are secreted as inactive zymogens (pro-MMPs) requiring extracellular activation, and their activity is tightly regulated by specific tissue inhibitors of MMPs (TIMPs). The MMP family has been implicated in a wide range of physiologic and pathological processes [10]. MMP has a pivotal role in maintaining the homeostasis of the extracellular matrix (ECM) and tissue remodeling during development, wound healing, and pathogenesis, e.g., tumorigenesis. However, in pathological conditions, MMPs can aid tissue destruction, tumor infiltration and metastasis, angiogenesis, and inflammatory reactions [11]. Our previous studies have demonstrated that increased production and activity of MMPs play an important role in patients with dry eye [12] and the murine dry eye model [2,13-15].
Traditional approaches to reduce tear hyperosmolarity in dry eye include use of hypotonic tear substitutes, which have relatively short persistence in the eye. Recent attempts to counteract tear hyperosmolarity include topical application of osmoprotectants, which are small organic molecules used in many cell types to restore cell volume, stabilize protein function, and protect cells from hyperosmolarity stress [16]. Osmoprotectants, also known as “organic osmolytes” or “compatible solutes,” can be classified as amino acids (e.g., glycine, betaine, proline, taurine), polyols (e.g., glycerol, erythritol, inositols, sorbitol), small carbohydrates (e.g., trehalose), methylamines/methylsulfonium solutes (e.g., L-carnitine), or urea [16-20]. Their uptake is accompanied by a decreasing concentration of intracellular inorganic salts [21].
We have previously demonstrated that L-carnitine and erythritol protected human corneal epithelial cells against mitogen-activated protein (MAP) kinase activation by hyperosmotic stress [22]. Reduced levels of tear L-carnitine in patients with dry eye compared with healthy subjects have been reported [23]. In the murine dry eye model, topical application of osmoprotectants, L-carnitine, betaine, or erythritol, was observed to reduce the severity of dry eye conditions induced by the desiccated environment [24]. However, there is no report whether these osmoprotectants affect the expression of MMPs that are involved in the pathogenesis of inflammatory dry eye disease. The present study explored the suppressive effects of osmoprotectants, L-carnitine, erythritol, or betaine on the expression, production, and activity of MMPs stimulated by hyperosmolar media in primary human corneal epithelial cells (HCECs).
Methods
Materials and reagents
Cell culture dishes, plates, centrifuge tubes, and other plastic ware were purchased from BD Biosciences (Lincoln Park, NJ). Dulbecco modified Eagle medium (DMEM), Ham F-12, amphotericin B, and gentamicin were from Invitrogen (Grand Island, NY). Fetal bovine serum (FBS) was from Hyclone (Logan, UT). L-carnitine, erythritol, and betaine were from Sigma-Aldrich (St. Louis, MO). The RNeasy Plus Mini RNA extraction kit from Qiagen (Valencia, CA). The Ready-To-Go-Primer First-Strand Beads were from GE Healthcare (Piscataway, NJ). The TaqMan gene expression assays and real-time PCR master mix were from Applied Biosystems (Foster City, CA). The MMP antibodies were from Sigma-Aldrich (St. Louis, MO). The ready zymogram gels were from Bio-Rad (Hercules, CA).
Primary cultures of human corneal epithelial cells and cell treatment
Fresh human corneoscleral tissues (<72 h after death) not suitable for clinical use, from donors aged 19 to 67 years, were obtained from the Lions Eye Bank of Texas (Houston, TX). This study was approved by Baylor College of Medicine Institutional Review Board and adhered to the tenets of the Declaration of Helsinki and the ARVO statement on human tissue specimens. Primary HCECs were cultured in 12-well plates using explants from corneal limbal rims in a supplemented hormonal epidermal medium (SHEM) containing 5% FBS using our previous methods [25]. Confluent primary corneal epithelial cultures in 14 days were switched to an equal volume (1.0 ml/well) of serum-free medium (SHEM without FBS) for 24 h or passaged to eight-chamber slides, and then treated for 4 or 24 h with different osmolar media (312, 400, 450, and 500 mOsM), with or without 1 h prior incubation with different concentrations (2, 10, or 20 mM) of L-carnitine, erythritol, or betaine. The osmolarity of the culture media was measured with a vapor pressure osmometer in the Body Fluid Chemistry Clinical Laboratory of the Methodist Hospital (Houston, TX). The SHEM is an isotonic medium with 312 mOsM osmolarity. To increase the osmolarity to 400, 450, or 500 mOsM, 22, 34.5, or 47 µl of sterile solution of 2N sodium chloride were added into 1 ml SHEM of each well of the 12-well culture plates, which generated additional 44, 69, or 94 mM sodium chloride, corresponding to an increase of 88, 138, or 188 mOsM, and thus made the final osmolarity of these media to 400, 450, or 500 mOsM, respectively. The cells treated for 4 h were lysed in RLT buffer from the Qiagen RNeasy Plus Mini kit for RNA extraction. The cells treated for 24 h were used for zymography or immunofluorescent staining. The conditioned media were collected and centrifuged, and the supernatants were stored at −80 °C before being used for zymography.
RNA extraction, reverse transcription, and quantitative real-time PCR
Total RNA was extracted with a RNeasy Plus Mini Kit (Qiagen, Valencia, CA) according to the manufacturer’s instructions, quantified with a spectrophotometer (NanoDrop ND-1000; Thermo Scientific, Wilmington, DE), and stored at −80 °C before use. The first strand cDNA was synthesized using Ready-To-Go You-Prime First-Strand Beads as previously described [13,26]. In brief, the first strand cDNA was synthesized at 37 °C for 60 min in 33 μl reaction volume mixed with 1.0 μg of total RNA, 1 μl of random hexamers, and 2 beads containing dATP, dCTP, dGTP, dTTP, and M-MuLV reverse transcriptase. Quantitative real-time PCR (RT-qPCR) was performed in a Mx3005P QPCR System (Stratagene, La Jolla, CA) with 20 µl reaction volume containing 5 µl of cDNA, 1 µl gene expression assay, and 10 µl gene expression master mix (TaqMan; ABI). The TaqMan gene expression assays used for this study were glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Hs99999905_m1), MMP-13 (Hs00942589_m1), MMP-9 (Hs00234579_m1), MMP-2 (Hs01548724_m1), MMP-3 (Hs00233962_m1), and MMP-7 (Hs00159163_ml). The thermocycler parameters were 50 °C for 2 min and 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. A nontemplate control was included to evaluate DNA contamination. The results were analyzed with the comparative threshold cycle (Ct) method and normalized with GAPDH as an internal control [27].
Immunofluorescent staining
The human corneal epithelial cells on eight-chamber slides were fixed with 100% methanol at 4 °C for 10 min and permeabilized with 0.2% Triton X-100 in PBS (1X; 150 mM sodium chloride, 10 mM sodium phosphate, pH 7.3) at room temperature for 10 min. Primary goat polyclonal antibody (Ab) against human MMP-13, MMP-3, or MMP-7 (dilution 1:50) were used for immunofluorescent staining. Alexa-Fluor 488 conjugated secondary antibodies were applied, and propidium iodide (PI) was used for nuclear counterstaining. Secondary antibody alone without primary antibody was used as negative control. The results were photographed with epifluorescence microscopy (Eclipes 400; Nikon, Garden City, NY) using a digital camera (DMX1200, Nikon).
Gelatin zymography
To determine the relative concentrations of gelatinases in the conditioned media from the corneal epithelial cultures that received various treatments, sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) gelatin zymography was performed using a previously reported method [28]. Briefly, 10 μl of each conditioned medium was used. The media supernatants were treated with SDS sample buffer without boiling or reduction. Samples were fractionated in a 10% polyacrylamide gel with electrophoresis at 100 V for 90 min at 4 °C. The gels were soaked in 0.25% Triton X-100 for 30 min at room temperature to remove the SDS, and incubated in a digestion buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 10 mM CaCl2, 2 μM ZnSO4, and 0.01% Brij-35) containing 5 mM phenylmethylsulfonyl fluoride (PMSF), a serine protease inhibitor, at 37 °C overnight to allow proteinase digestion of its substrate. Gels were rinsed again in distilled water, stained with 0.25% Coomassie brilliant blue R-250 in 40% isopropanol for 2 h, and destained with 7% acetic acid. Gelatinolytic activities appeared as clear bands of digested gelatin against a dark blue background of stained gelatin.
Statistical analysis
The Student t test was used to compare differences between two groups. One-way ANOVA test was used to make comparisons among three or more groups, followed by Dunnett’s post-hoc test. p<0.05 was considered statistically significant.
Results
The expression of collagenase MMP-13, gelatinases MMP-9 and MMP-2, stromelysin MMP-3, and matrilysin MMP-7 was significantly increased in HCECs exposed to hyperosmotic media
Primary HCECs cultured in SHEM with iso-osmolarity (312 mOsM) were switched to hyperosmotic media (400, 450, or 500 mOsM) for 4 h for evaluation of mRNA expression of MMPs. As shown in Figure 1, the mRNA levels of MMP-13 (Figure 1A), MMP-9 (Figure 1B), MMP-2 (Figure 1C), MMP-3 (Figure 1D), and MMP-7 (Figure 1E) were significantly increased in primary HCECs exposed to hyperosmotic media, mostly in a osmolarity-dependent manner from 312 to 400 and 450 mOsM. Compared with the iso-osmolar medium as control, the MMP-13 mRNA expression of HCECs significantly increased to 3.06±1.98, 6.99±2.92, and 7.48±3.10 fold, respectively, when exposed to increasing hyperosmolarity, 400, 450, or 500 mOsM (p<0.01, p<0.001, p<0.001, respectively). MMP-9 mRNA increased to 1.65±0.86, 2.54±1.52, and 2.30±1.25 fold, and MMP-2 expression increased to 1.68±0.46, 2.90±1.14, and 2.94±1.19 fold, respectively, in response to hyperosmolarity at 400, 450, or 500 mOsM (p<0.05, p<0.01, p<0.01, respectively). The mRNA levels of MMP-3 increased to 2.18±1.14, 4.08±1.70, and 2.56±1.24 fold, and MMP-7 increased to 2.36±1.28, 4.53±1.41, and 2.49±1.47-fold, respectively, by hyperosmotic media (400, 450, or 500 mOsM), all p<0.01 or p<0.001, respectively.
Figure 1.
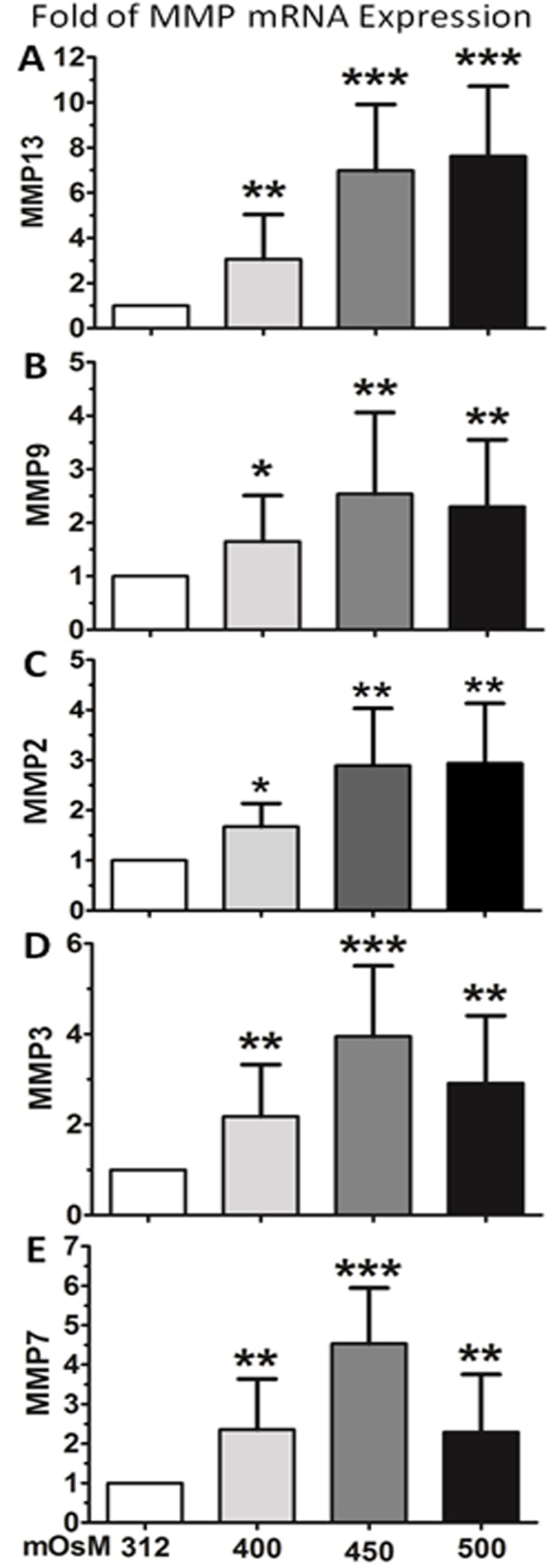
Expression of MMP mRNA in primary HCECs. The increased mRNA expression of MMP-13 (A), MMP-9 (B), MMP-2 (C), MMP-3 (D) and MMP-7 (E) was observed in HCECs exposed to hyperosmotic media. Each bar represents mean±SD of 4 independent experiments. *P<0.05, **P<0.01, ***P<0.001, compared with normal control medium with 312 mOsM.
L-carnitine, erythritol, or betaine suppressed the mRNA expression of MMP-13, MMP-9, MMP-2, MMP-3, and MMP-7 stimulated by hyperosmolarity in HCECs
Primary HCECs in iso-osmolar SHEM were switched to SHEM with 450 mOsM osmolar media, without or with 1 h prior incubation of L-carnitine, erythritol, or betaine at different concentrations (2, 10, or 20 mM). The differentially suppressive effects of L-carnitine, erythritol, or betaine on MMP expression were observed in HCECs exposed to hyperosmotic media.
Taking the stimulated mRNA levels (7.0 fold) by 450 mOsM hyperosmolarity as a positive control, MMP-13 expression significantly decreased to 4.06±0.80 (p<0.05), 3.26±0.98 (p<0.01), 3.09±0.47 (p<0.01) fold by L-carnitine at 2, 10, and 20 mM, respectively (Figure 2A). L-carnitine at 2, 10, and 20 mM showed a similar suppressive effect on MMP-9 expression that decreased from stimulated 2.74±1.52 fold to 1.34±0.61 (p<0.05), 1.22±0.24 (p<0.01), and 1.18±0.46 (p<0.01; Figure 2B). Stronger inhibition of L-carnitine (2–20 mM) on MMP-3 (Figure 2D) and MMP-7 (Figure 2E) was also observed with statistical significance reached at p<0.001.
Figure 2.
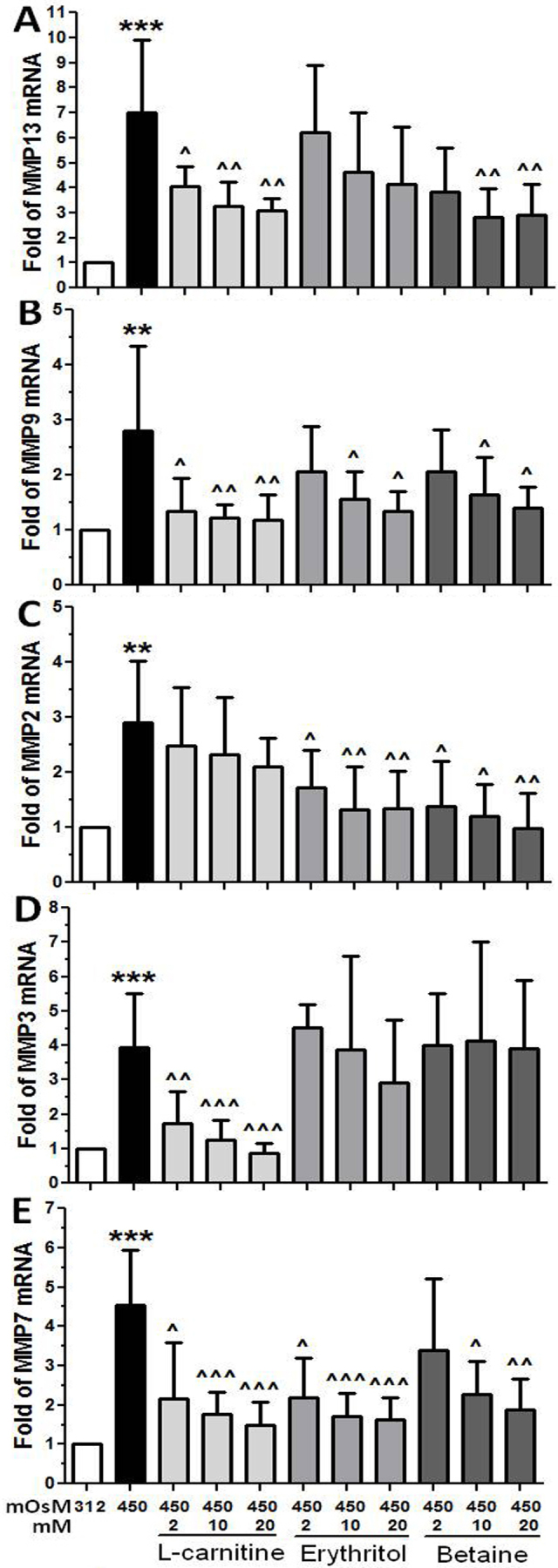
Effects of osmoprotectants on MMP expression. Suppressive effects of L-carnitine, erythritol and betaine on the mRNA expression of MMP-13 (A), MMP-9 (B) , MMP-2 (C), MMP-3 (D) and MMP-7 (E) were observed in primary HCECs exposed to hyperosmotic medium (450 mOsM). Each bar represents mean±SD of 4 independent experiments. *P<0.05, **P<0.01, ***P<0.001, compared with 312 mOsM; ^^P<0.01, ^^^P<0.001, compared with 450 mOsM.
Erythritol at 10–20 mM suppressed the mRNA expression of MMP-9, MMP-2, and MMP-7, but not MMP-13 and MMP-3. The expression of MMP-9 (Figure 2B) andMMP -2 (Figure 2C) was induced to 2.74±1.52 and 2.90±1.14 fold by 450 mOsM respectively, but decreased to 1.57±0.48 and 1.35±0.35 or 1.33±0.78 and 1.35±0.67 fold (p<0.05, p<0.01) by 10 or 20 mM of erythritol respectively. The MMP-7 mRNA levels were also from induced 4.53±1.41 fold decreased to 2.17±1.01 (p<0.05), 1.70±0.60 (p<0.001), and 1.60±0.54 (p<0.001), respectively, by erythritol at 2, 10, or 20 mM (Figure 2E).
Suppressed mRNA levels of MMP-13, MMP-9, MMP-2, and MMP-7 but not MMP-3 were observed when HCECs in 450 mOsM medium treated with betaine. MMP-13 expression was induced to 6.99±2.92 fold by 450 mOsM, but decreased to 2.82±1.17 and 2.92±1.23 (both p<0.01) by 10 or 20 mM of betaine (Figure 2A), MMP-9 expression decreased from 2.74±1.52 fold to 1.60±0.48 and 1.40±0.39 (Figure 2B, both p<0.05); and MMP-2 mRNA decreased from 2.90±1.14 fold to near normal levels (Figure 2C, p<0.01) by betaine at 10 or 20 mM. The MMP-7 mRNA levels were also from induced 4.53±1.41 fold decreased to 2.26±0.86 (p<0.05) and 1.89±0.77 (p<0.01), respectively, by betaine at 10 or 20 mM (Figure 2E).
L-carnitine, erythritol, or betaine inhibited the protein production of MMP-13, MMP-3, and/or MMP-7 stimulated by hyperosmolarity in HCECs
The protein production of MMP-13, MMP-3, and MMP-7 was evaluated with immunofluorescent staining with specific antibodies (Figure 3). It appeared that the immunoreactivity of normal HCECs to these three MMP antibodies was weak. The staining intensity was markedly increased when the cells were treated with hyperosmotic media at 400 and 450 mOsM for 24 h. Obviously, prior incubation with 20 mM of L-carnitine blocked the stimulated reactivity as the decreased intensity of MMP13, MMP-3, and MMP-7 shown in Figure 3. Erythritol at 20 mM displayed a suppressive effect on MMP-7 immunofluorescent staining but weak or no inhibition of MMP-13 or MMP-3 staining. Betaine (20 mM) showed suppressive effects on MMP-13 and MMP-7 but not MMP-3, similar to the effect pattern on the mRNA expression of MMPs.
Figure 3.

Suppressive effects of L-carnitine, erythritol, or betaine at 20 mM on MMP protein production in primary HCECs exposed to hyperosmotic media (400–450 mOsM), evaluated with immunofluorescent staining for MMP-13, MMP-3, and MMP-7.
L-carnitine, erythritol, or betaine inhibited the protein production and enzymatic activity of MMP-9 stimulated by hyperosmolarity in HCECs
MMP-9 has been proven to be a key MMP highly involved in dry eye disease [2,12-14]. Gelatin zymography was performed to further evaluate the effects of osmoprotectants on the protein production and enzymatic activity of gelatinases, MMP-9 and MMP-2. As shown in Figure 4, mainly in the proforms, 72 kDa MMP-2 and 92 kDa MMP-9 were secreted in the culture media by normal primary HCECs, with barely or weak detected bands of the activated forms, 62 kDa MMP-2 and 82 kDa MMP-9. The total production and activity of 92 kDa MMP-9, and especially the 82 kDa active MMP-9, were significantly stimulated in an osmolarity-dependent fashion in the media from HCECs exposed to media with higher osmolarity (400–450 mOsM). The stimulated protein production and enzymatic activity was dramatically suppressed by L-carnitine, erythritol, or betaine in a concentration-dependent manner. MMP-2, known as a constitutively produced gelatinase, was also stimulated by hyperosmolar media. Interestingly, erythritol and betaine but not L-carnitine showed suppressive effects on MMP-2, especially on the active form of the 62 kDa MMP-2 protein (Figure 4).
Figure 4.

Representative images of zymography. Suppressive effects of L-carnitine (A), erythritol (B) or betaine (C) at 2-20 mM on protein production and enzymatic activity of gelatinases MMP-9 and MMP-2 were observed in primary HCECs exposed to hyperosmotic media (400-450 mOsM), evaluated by gelatin zymography.
Discussion
In the present study, we evaluated the effects of L-carnitine, betaine, and erythritol on mRNA expression, protein production, and enzymatic activity of MMPs in HCECs exposed to media with higher osmolarity (400–500 mOsM), as measured with RT-qPCR, immunofluorescent staining, and zymography. MMPs have been implicated in the pathogenesis of several ocular surface diseases, including dry eye, wound healing, sterile corneal ulceration, recurrent epithelial erosion, corneal neovascularization, pterygium, and conjunctivochalasis [29-34]. Increased levels of MMPs, such as MMP-3 and MMP-9, have been observed in the tear fluid of patients with dry eye [28,35]. Since elevated tear fluid osmolarity has been recognized as a common feature of dry eye for decades [36], it is not surprising that the MMPs were stimulated by hyperosmolarity. A previous study showed that hyperosmotic stress stimulates the expression and production of collagenases (MMP-1 and MMP-13), gelatinase (MMP-9), and stromelysin (MMP-3) by HCECs [8]. In the present study, we consistently observed increased MMP13, MMP-9, and MMP-3, as well as a matrilysins MMP-7 in HCECs exposed to the hyperosmotic medium.
L-carnitine acts as an osmoprotectant against the hyperosmotic stress of corneal epithelial cells in vitro [22,37,38]. The tear L-carnitine levels in patients with dry eye were lower than that in healthy subjects [23]. The topical use of L-carnitine has been demonstrated to result in rapid and consistent improvements in the signs and symptoms of dry eye [39]. The structure of the osmolyte erythritol resembles mannitol, a well-known antioxidant [18], and oxidative damage has been implicated in the pathogenesis and development of dry eye [40,41]. Betaine, a metabolite of plant and animal tissues, is a protectant against salt and temperature stress in plants [17]. Betaine also stabilizes corneal epithelial cell volume under hyperosmotic stress and limited hyperosmotic stress-induced apoptosis of human corneal epithelial cells [42]. L-carnitine, erythritol, and/or betaine has been reported to serve as an osmoprotectant against hyperosmotic stress [17,19,43], and inflammatory conditions [43,44]. However, it is not clear whether these small molecules of osmoprotectants affect the expression and activity of MMPs.
In the present study, for the first time, we demonstrated that these osmoprotectants differentially suppress gene expression, protein production, and enzymatic activity of collagenase MMP-13, gelatinases MMP-9 and MMP-2, stromelysin MMP-3, and matrilysin MMP-7 in primary HCECs exposed in hyperosmotic stress. Among the three molecules, L-carnitine appears to have a broader and stronger suppressive effect on these MMPs in four groups. Erythritol mainly suppresses MMP-9, MMP-2, and MMP-7; betaine inhibited the four MMPs but not MMP-3. More interestingly, these osmoprotectants not only suppress gene expression at transcriptional levels and protein production at the translational level of MMPs but also inhibit the activation of pro-MMPs at post-translational levels, as shown with zymography for MMP-9 and MMP-2 as representative examples.
Most MMPs are synthesized as inactive zymogens (pro-MMP) that undergo subsequent activation in the extracellular milieu. Pro-MMPs can be activated by various factors such as organomercurials, serine proteinases, hypochlorous acid, and acid exposure [45]. However, intermolecular activation of pro-MMP by active MMP may be the physiologic mechanism of activation in vivo. The stromelysins MMP-3 and MMP-10 play a central role in the activation of various pro-MMPs, including pro-MMP-9 [46,47], pro-MMP-1 [48], pro-MMP-8 [49], and pro-MMP-7 [47]. Pro-MMP-9 (92 kDa) is processed to its full activity as a low molecular weight species of 82 kDa and 65 kDa by MMP-3 or -10. Membrane-type metalloproteinase 1 (MT1-MMP, MMP-14) can activate pro-MMP-2 in vitro [45].
Gelatin zymography (Figure 4) clearly showed that the production and activity of the 92 kDa MMP-9, and especially the 82 kDa active-MMP-9, were markedly stimulated by hyperosmolarity in the media of HCECs, but concentration-dependently suppressed by L-carnitine, erythritol, or betaine. Twenty mM seems to be the best dose to suppress the activity of MMP-9. Zymography also showed that the 62 kDa active-MMP-2 protein was stimulated by hyperosmolar media, and inhibited by erythritol and betaine, but not L-carnitine.
In conclusion, our findings demonstrate that hyperosmotic stress stimulates the production and activity of MMPs in HCECs. Serving as osmoprotectants, L-carnitine, erythritol, and betaine differentially suppress the expression, production, and activation of MMPs in primary HCECs exposed to hyperosmotic stress. These osmoprotectants may have potential effects in protecting ocular surface epithelia from MMP-mediated disorders in dry eye disease. MAP kinases are involved in directing cellular responses to a diverse array of stimuli, including mitogens, osmotic stress, and proinflammatory cytokines. Our previous studies have showed that these osmoprotectants protect corneal epithelia against hyperosmotic stress by inhibiting the activation of MAP kinases [22]. However, further studies are necessary to explore the mechanisms by which these osmoprotectants protect ocular surface epithelia against hyperosmotic stress by suppressing MMP production and activity.
Acknowledgments
This study was supported by National Eye Institute, National Institutes of Health grants R01EY11915 (SCP) and Core Grant for Vision Research P30EY002520, Allergan Inc., Eye Bank Association of America (DQL), an unrestricted grant from Research to Prevent Blindness, the Oshman Foundation and the William Stamps Farish Fund.
References
- 1.Li D-Q, Luo L, Chen Z, Kim HS, Song XJ, Pflugfelder SC. JNK and ERK MAP kinases mediate induction of IL-1beta, TNF-alpha and IL-8 following hyperosmolar stress in human limbal epithelial cells. Exp Eye Res. 2006;82:588–96. doi: 10.1016/j.exer.2005.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.De Paiva CS, Corrales RM, Villarreal AL, Farley WJ, Li D-Q, Stern ME, Pflugfelder SC. Corticosteroid and doxycycline suppress MMP-9 and inflammatory cytokine expression, MAPK activation in the corneal epithelium in experimental dry eye. Exp Eye Res. 2006;83:526–35. doi: 10.1016/j.exer.2006.02.004. [DOI] [PubMed] [Google Scholar]
- 3.Luo L, Li DQ, Pflugfelder SC. Hyperosmolarity-induced apoptosis in human corneal epithelial cells is mediated by cytochrome c and MAPK pathways. Cornea. 2007;26:452–60. doi: 10.1097/ICO.0b013e318030d259. [DOI] [PubMed] [Google Scholar]
- 4.Chen W, Zhang X, Liu M, Zhang J, Ye Y, Lin Y, Luyckx J, Qu J. Trehalose protects against ocular surface disorders in experimental murine dry eye through suppression of apoptosis. Exp Eye Res. 2009;89:311–8. doi: 10.1016/j.exer.2009.03.015. [DOI] [PubMed] [Google Scholar]
- 5.Clouzeau C, Godefroy D, Riancho L, Rostene W, Baudouin C, Brignole-Baudouin F. Hyperosmolarity potentiates toxic effects of benzalkonium chloride on conjunctival epithelial cells in vitro. Mol Vis. 2012;18:851–63. [PMC free article] [PubMed] [Google Scholar]
- 6.Png E, Samivelu GK, Yeo SH, Chew J, Chaurasia SS, Tong L. Hyperosmolarity-mediated mitochondrial dysfunction requires Transglutaminase-2 in human corneal epithelial cells. J Cell Physiol. 2011;226:693–9. doi: 10.1002/jcp.22389. [DOI] [PubMed] [Google Scholar]
- 7.Li D-Q, Pflugfelder SC. Matrix metalloproteinases in corneal inflammation. Ocul Surf. 2005;3:S198–202. doi: 10.1016/s1542-0124(12)70255-0. [DOI] [PubMed] [Google Scholar]
- 8.Li D-Q, Chen Z, Song XJ, Luo L, Pflugfelder SC. Stimulation of matrix metalloproteinases by hyperosmolarity via a JNK pathway in human corneal epithelial cells. Invest Ophthalmol Vis Sci. 2004;45:4302–11. doi: 10.1167/iovs.04-0299. [DOI] [PubMed] [Google Scholar]
- 9.Khokha R, Murthy A, Weiss A. Metalloproteinases and their natural inhibitors in inflammation and immunity. Nat Rev Immunol. 2013;13:649–65. doi: 10.1038/nri3499. [DOI] [PubMed] [Google Scholar]
- 10.Sorokin L. The impact of the extracellular matrix on inflammation. Nat Rev Immunol. 2010;10:712–23. doi: 10.1038/nri2852. [DOI] [PubMed] [Google Scholar]
- 11.Sakimoto T, Sawa M. Metalloproteinases in corneal diseases: degradation and processing. Cornea. 2012;31(Suppl 1):S50–6. doi: 10.1097/ICO.0b013e318269ccd0. [DOI] [PubMed] [Google Scholar]
- 12.Chotikavanich S, de Paiva CS, Li DQ, Chen JJ, Bian F, Farley WJ, Pflugfelder SC. Production and Activity of Matrix Metalloproteinase-9 on the Ocular Surface Increase in Dysfunctional Tear Syndrome. Invest Ophthalmol Vis Sci. 2009;50:3203–9. doi: 10.1167/iovs.08-2476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Luo L, Li D-Q, Doshi A, Farley W, Corrales RM, Pflugfelder SC. Experimental dry eye stimulates production of inflammatory cytokines and MMP-9 and activates MAPK signaling pathways on the ocular surface. Invest Ophthalmol Vis Sci. 2004;45:4293–301. doi: 10.1167/iovs.03-1145. [DOI] [PubMed] [Google Scholar]
- 14.Pflugfelder SC, Farley W, Luo L, Chen LZ, de Paiva CS, Olmos LC, Li D-Q, Fini ME. Matrix metalloproteinase-9 knockout confers resistance to corneal epithelial barrier disruption in experimental dry eye. Am J Pathol. 2005;166:61–71. doi: 10.1016/S0002-9440(10)62232-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Luo L, Li D-Q, Corrales RM, Pflugfelder SC. Hyperosmolar saline is a proinflammatory stress on the mouse ocular surface. Eye Contact Lens. 2005;31:186–93. doi: 10.1097/01.icl.0000162759.79740.46. [DOI] [PubMed] [Google Scholar]
- 16.Baudouin C, Aragona P, Messmer EM, Tomlinson A, Calonge M, Boboridis KG, Akova YA, Geerling G, Labetoulle M, Rolando M. Role of hyperosmolarity in the pathogenesis and management of dry eye disease: proceedings of the OCEAN group meeting. Ocul Surf. 2013;11:246–58. doi: 10.1016/j.jtos.2013.07.003. [DOI] [PubMed] [Google Scholar]
- 17.Eklund M, Bauer E, Wamatu J, Mosenthin R. Potential nutritional and physiological functions of betaine in livestock. Nutr Res Rev. 2005;18:31–48. doi: 10.1079/NRR200493. [DOI] [PubMed] [Google Scholar]
- 18.den Hartog GJ, Boots AW, Adam-Perrot A, Brouns F, Verkooijen IW, Weseler AR, Haenen GR, Bast A. Erythritol is a sweet antioxidant. Nutrition. 2010;26:449–58. doi: 10.1016/j.nut.2009.05.004. [DOI] [PubMed] [Google Scholar]
- 19.Peluso G, Barbarisi A, Savica V, Reda E, Nicolai R, Benatti P, Calvani M. Carnitine: an osmolyte that plays a metabolic role. J Cell Biochem. 2000;80:1–10. doi: 10.1002/1097-4644(20010101)80:1<1::aid-jcb10>3.0.co;2-w. [DOI] [PubMed] [Google Scholar]
- 20.Verheul A, Wouters JA, Rombouts FM, Abee T. A possible role of ProP, ProU and CaiT in osmoprotection of Escherichia coli by carnitine. J Appl Microbiol. 1998;85:1036–46. doi: 10.1111/j.1365-2672.1998.tb05269.x. [DOI] [PubMed] [Google Scholar]
- 21.Yancey PH. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J Exp Biol. 2005;208:2819–30. doi: 10.1242/jeb.01730. [DOI] [PubMed] [Google Scholar]
- 22.Corrales RM, Luo L, Chang EY, Pflugfelder SC. Effects of osmoprotectants on hyperosmolar stress in cultured human corneal epithelial cells. Cornea. 2008;27:574–9. doi: 10.1097/ICO.0b013e318165b19e. [DOI] [PubMed] [Google Scholar]
- 23.Pescosolido N, Imperatrice B, Koverech A, Messano M. L-carnitine and short chain ester in tears from patients with dry eye. Optom Vis Sci. 2009;86:E132–8. doi: 10.1097/OPX.0b013e318194e767. [DOI] [PubMed] [Google Scholar]
- 24.Chen W, Zhang X, Li J, Wang Y, Chen Q, Hou C, Garrett Q. Efficacy of osmoprotectants on prevention and treatment of murine dry eye. Invest Ophthalmol Vis Sci. 2013;54:6287–97. doi: 10.1167/iovs.13-12081. [DOI] [PubMed] [Google Scholar]
- 25.Kim HS, Jun S. X, de Paiva CS, Chen Z, Pflugfelder SC, Li D-Q. Phenotypic characterization of human corneal epithelial cells expanded ex vivo from limbal explant and single cell cultures. Exp Eye Res. 2004;79:41–9. doi: 10.1016/j.exer.2004.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yoon KC, de Paiva CS, Qi H, Chen Z, Farley WJ, Li DQ, Pflugfelder SC. Expression of Th-1 chemokines and chemokine receptors on the ocular surface of C57BL/6 mice: effects of desiccating stress. Invest Ophthalmol Vis Sci. 2007;48:2561–9. doi: 10.1167/iovs.07-0002. [DOI] [PubMed] [Google Scholar]
- 27.Ma P, Bian F, Wang Z, Zheng X, Chotikavanich S, Pflugfelder SC, Li DQ. Human corneal epithelium-derived thymic stromal lymphopoietin links the innate and adaptive immune responses via TLRs and Th2 cytokines. Invest Ophthalmol Vis Sci. 2009;50:2702–9. doi: 10.1167/iovs.08-3074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sobrin L, Liu Z, Monroy DC, Solomon A, Selzer MG, Lokeshwar BL, Pflugfelder SC. Regulation of MMP-9 activity in human tear fluid and corneal epithelial culture supernatant. Invest Ophthalmol Vis Sci. 2000;41:1703–9. [PubMed] [Google Scholar]
- 29.Kvanta A, Sarman S, Fagerholm P, Seregard S, Steen B. Expression of matrix metalloproteinase-2 (MMP-2) and vascular endothelial growth factor (VEGF) in inflammation-associated corneal neovascularization. Exp Eye Res. 2000;70:419–28. doi: 10.1006/exer.1999.0790. [DOI] [PubMed] [Google Scholar]
- 30.Li D-Q, Lee SB, Gunja-Smith Z, Liu Y, Solomon A, Meller D, Tseng SC. Overexpression of collagenase (MMP-1) and stromelysin (MMP-3) by pterygium head fibroblasts. Arch Ophthalmol. 2001;119:71–80. [PubMed] [Google Scholar]
- 31.Fini ME, Cook JR, Mohan R. Proteolytic mechanisms in corneal ulceration and repair. Arch Dermatol Res. 1998;290(Suppl):S12–23. doi: 10.1007/pl00007449. [DOI] [PubMed] [Google Scholar]
- 32.Dursun D, Kim MC, Solomon A, Pflugfelder SC. Treatment of recalcitrant recurrent corneal erosions with inhibitors of matrix metalloproteinase-9, doxycycline and corticosteroids. Am J Ophthalmol. 2001;132:8–13. doi: 10.1016/s0002-9394(01)00913-8. [DOI] [PubMed] [Google Scholar]
- 33.Di Girolamo N, Wakefield D, Coroneo MT. Differential expression of matrix metalloproteinases and their tissue inhibitors at the advancing pterygium head. Invest Ophthalmol Vis Sci. 2000;41:4142–9. [PubMed] [Google Scholar]
- 34.Li D-Q, Meller D, Liu Y, Tseng SC. Overexpression of MMP-1 and MMP-3 by cultured conjunctivochalasis fibroblasts. Invest Ophthalmol Vis Sci. 2000;41:404–10. [PubMed] [Google Scholar]
- 35.Pflugfelder SC, Solomon A, Stern ME. The diagnosis and management of dry eye: a twenty-five-year review. Cornea. 2000;19:644–9. doi: 10.1097/00003226-200009000-00009. [DOI] [PubMed] [Google Scholar]
- 36.Farris RL. Contact lenses and the dry eye. Int Ophthalmol Clin. 1994;34:129–36. doi: 10.1097/00004397-199403410-00012. [DOI] [PubMed] [Google Scholar]
- 37.Khandekar N, Willcox MD, Shih S, Simmons P, Vehige J, Garrett Q. Decrease in hyperosmotic stress-induced corneal epithelial cell apoptosis by L-carnitine. Mol Vis. 2013;19:1945–56. [PMC free article] [PubMed] [Google Scholar]
- 38.Garrett Q, Min-Hsuan Shih S, Simmons PA, Vehige J, Willcox M. Carnitine and the potential osmoprotectants protect corneal epithelial cells from hyperosmolar solution induced damage. Exp Eye Res. 2010;108:33–41. [Google Scholar]
- 39.Simmons P, Vehige J, Sasai A, Carlisle-Wilcox C, Beard B, Blue M, Lai F. Clinical Performance of an Osmoprotective Formulation for Dry Eye Therapy. In 8th Int Symp Ocular Pharm Ther, Proceedings, Bologna: Medimond, 2009, pp 169-173 [Google Scholar]
- 40.Uchino Y, Kawakita T, Miyazawa M, Ishii T, Onouchi H, Yasuda K, Ogawa Y, Shimmura S, Ishii N, Tsubota K. Oxidative stress induced inflammation initiates functional decline of tear production. PLoS ONE. 2012;7:e45805. doi: 10.1371/journal.pone.0045805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pescosolido N, Imperatrice B, Karavitis P. The aging eye and the role of L-carnitine and its derivatives. Drugs R D. 2008;9(Suppl 1):3–14. doi: 10.2165/0126839-200809001-00002. [DOI] [PubMed] [Google Scholar]
- 42.Garrett Q, Khandekar N, Shih S, Flanagan JL, Simmons P, Vehige J, Willcox MD. Betaine stabilizes cell volume and protects against apoptosis in human corneal epithelial cells under hyperosmotic stress. Exp Eye Res. 2013;108:33–41. doi: 10.1016/j.exer.2012.12.001. [DOI] [PubMed] [Google Scholar]
- 43.Flanagan JL, Simmons PA, Vehige J, Willcox MD, Garrett Q. Role of carnitine in disease. Nutr Metab (Lond) 2010;7:30. doi: 10.1186/1743-7075-7-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Morita Y, Tadokoro S, Sasai M, Kitamoto D, Hirashima N. Biosurfactant mannosyl-erythritol lipid inhibits secretion of inflammatory mediators from RBL-2H3 cells. Biochim Biophys Acta. 2011;1810:1302–8. doi: 10.1016/j.bbagen.2011.07.002. [DOI] [PubMed] [Google Scholar]
- 45.Nagase H. Activation mechanisms of matrix metalloproteinases. Biol Chem. 1997;378:151–60. [PubMed] [Google Scholar]
- 46.Ramos-DeSimone N, Hahn-Dantona E, Sipley J, Nagase H, French DL, Quigley JP. Activation of matrix metalloproteinase-9 (MMP-9) via a converging plasmin/stromelysin-1 cascade enhances tumor cell invasion. J Biol Chem. 1999;274:13066–76. doi: 10.1074/jbc.274.19.13066. [DOI] [PubMed] [Google Scholar]
- 47.Nakamura H, Fujii Y, Ohuchi E, Yamamoto E, Okada Y. Activation of the precursor of human stromelysin 2 and its interactions with other matrix metalloproteinases. Eur J Biochem. 1998;253:67–75. doi: 10.1046/j.1432-1327.1998.2530067.x. [DOI] [PubMed] [Google Scholar]
- 48.Suzuki K, Enghild JJ, Morodomi T, Salvesen G, Nagase H. Mechanisms of activation of tissue procollagenase by matrix metalloproteinase 3 (stromelysin). Biochemistry. 1990;29:10261–70. doi: 10.1021/bi00496a016. [DOI] [PubMed] [Google Scholar]
- 49.Knäuper V, Murphy G, Tschesche H. Activation of human neutrophil procollagenase by stromelysin 2. Eur J Biochem. 1996;235:187–91. doi: 10.1111/j.1432-1033.1996.00187.x. [DOI] [PubMed] [Google Scholar]