

Abstract
Background
Auxin/indoleacetic acid (Aux/IAA) genes, coding a family of short-lived nuclear proteins, play key roles in wide variety of plant developmental processes, including root system regulation and responses to environmental stimulus. However, how they function in auxin signaling pathway and symbiosis with rhizobial in Medicago truncatula are largely unknown. The present study aims at gaining deeper insight on distinctive expression and function features of Aux/IAA family genes in Medicago truncatula during nodule formation.
Principal Findings
Using the latest updated draft of the full Medicago truncatula genome, a comprehensive identification and analysis of IAA genes were performed. The data indicated that MtIAA family genes are distributed in all the M. truncatula chromosomes except chromosome 6. Most of MtIAA genes are responsive to exogenous auxin and express in tissues-specific manner. To understand the biological functions of MtIAA genes involved in nodule formation, quantitative real-time polymerase chain reaction (qRT-PCR) was used to test the expression profiling of MtIAA genes during the early phase of Sinorhizobium meliloti (S. meliloti) infection. The expression patterns of most MtIAA genes were down-regulated in roots and up-regulated in shoots by S. meliloti infection. The differences in expression responses between roots and shoots caused by S. meliloti infection were alleviated by 1-NOA application.
Conclusion
The genome-wide identification, evolution and expression pattern analysis of MtIAA genes were performed in this study. The data helps us to understand the roles of MtIAA-mediated auxin signaling in nodule formation during the early phase of S. meliloti infection.
Introduction
The phytohormone auxin plays essential roles during the entire life-cycle of plants [1], [2]. Indole-3-acetic acid (IAA), the primary auxin in higher plants, has been found to effect on regulating diverse aspects of plant growth and development under environmental stimuli responses [3]. Some gene families, including Aux/IAA, GH3 (Gretchen Hagen3) and SAUR (small auxin up RNA), are responsive to auxin stimulation during early stage of auxin signaling transduction [4]. To be an important component of auxin signaling pathway, auxin/indole-3-acetic acid (Aux/IAA) proteins were well known as a direct target of the auxin transport inhibitor response 1 (TIR1) and its paralogs AUXIN RECEPTOR F-BOX/AFBs [5], [6]. Dynamic auxin concentration leads to the degradation of Aux/IAA proteins, which were involved in expression regulation of many auxin response genes by releasing ARFs (Auxin Response Factor) [7]–[9].
Most of Aux/IAA proteins contain four highly conserved domains: I, II, III and IV. Domain I, which contains a conserved leucine repeat (LXLXLX) motif, interacts with TOPLESS (TPL) protein to mediate auxin-dependent transcriptional repression during embryogenesis [10]. Domain II is responsible for the instability of Aux/IAA proteins [11]. Domain III and IV are the binding sites for homo- and hetero-dimerization among the Aux/IAA proteins and ARF proteins [12], [13]. It was reported that domain III and IV function as a complex. The crystal structure analysis revealed that Bem1p (PB1) domain in C-terminal is the special domain for the protein-protein interaction between Aux/IAA and ARF [14]. Moreover, Aux/IAA proteins contain two nuclear localization signals (NLS), which locate Aux/IAA proteins to the nucleus [15], [16].
In the past years, the functions of Aux/IAA family genes have been well studied. In Arabidopsis, loss function of IAA1/AXR5, which is a substrate of SCF (TIR1), causes a variety of auxin-related growth defects and auxin insensitivity phenotype [17]; iaa3/shy2 loss-of-function mutation affects auxin homeostasis and formation of lateral roots [18]; iaa7/axr2, iaa17/axr3, iaa19/msg2 and iaa28 are involved in the reduction of lateral root number [19]–[22]; iaa14/slr mutant even completely blocks lateral root formation [23]; a gain-of-function mutant iaa16 impedes plant growth and confers decreased response to phytohormone [20]. In monocot rice, some IAA genes also have been reported. Over-expression of OsIAA1 leads to inhibition of root elongation and shoot growth [24]; a gain-of-function in OsIAA11 results in the absent of lateral roots [25]; OsIAA23 defines postembryonic maintenance of quiescent center (QC) in rice [26]; OsIAA31 functions in lateral root development [27].
Many legume species including M. truncatula interact with nitrogen fixing bacteria (rhizobia) to form nodules, which are the symbiotic organ of legumes to host nitrogen-fixing bacteria [28], [29]. Phytohormone auxin has been reported to be a crucial element involved in nodules formation in the M. truncatula roots [30], [31]. As we already know, there is a close relationship between auxin signaling and rhizobia stimulation during the symbiosis formation [32]. As a model indeterminate legume, M. truncatula is used to reveal how auxin early responsive genes participate in nodule initiation and formation. The main role of auxin in regulation of mitosis during nodulation is controlling the inner and out cortical cell dividing [30]. After rhizobia stimulate, auxin signaling transduction and endogenesis auxin transport from shoots to roots are essential processes for root system architecture (RSA) regulation, which enhances pseudonodules formation on rhizobia inoculation roots [30], [33]. However, the underlying mechanism how auxin early responsive Aux/IAA family genes are involved in nodule formation at an early phase of S. meliloti infection of M. truncatula remains largely unknown
As an important part of auxin signaling pathway, Aux/IAAs are encoded by a multigene family and exist in most plant species. There are 29 members in Arabidopsis, 26 members in tomato (Solanum lycopersicon), 34 members in maize (Zea mays L.), and 31 members in rice (Oryza sativa L.) and 27 members in cucumber (Cucumis sativus L.) [16], [27], [34]–[38]. In this work, we provide comprehensive information on the genomic structures, chromosomal locations, sequences homology and expression patterns of seventeen IAA genes in M. truncatula. Our studies give a new insight into the complexity of M. truncatula IAA expressions at an early phase of S. meliloti infection. The distinct spatio-temporal expression patterns of M. truncatula IAA family genes and their differential responses to rhizobial symbiosis provided clues for functional characterization of these auxin responsive genes involved in formation of root nodules.
Materials and Methods
Plant material, growth conditions and hormone treatment
Medicago truncatula (Jemalong) A17 was used in this study. Seeds were dipped in sulfuric acid for 10 min to degrade seed coat, and washed with sterilized water three times. Then the seeds were germinated on plates contained 0.8% agar at 25°C until the radicals were about 1 cm. The seedling were grown in large plastic buckets containing full-strength nutrient solution and were incubated in a growth chamber at 22°C constantly during a 16 hour day and 8 hour night with a photon flux density of 100 µmolm−2s−1 for 14 days. The composition of full-strength nutrient solution is: 0.25 mM KH2PO4, 0.5 mM MgSO4·7H2O, 0.125 mM CaCl2, 1.25 mM KNO3, 0.5 mM NH4NO3, 15 µM H3BO3, 2.5 µM MnSO4·H2O, 0.5 µM ZnSO4·7H2O, 0.5 µM CuSO4·5H2O, 0.35 µM NaMoO4·2H2O, and 50 µM Fe (III) EDTA with a pH of 6.0. In auxin response experiment, the IAA concentration was 0.1 µM. Samples of shoots and roots were used to test the changes of MtIAA genes expression level at different time points (3 hr, 6 hr, 12 hr and 24 hr). The data were analyzed by five independent repeats, and standard deviations were shown with error bars. The 14-day-old seedlings were transferred to a nitrogen-free BNM medium [39] for S. meliloti infections and auxin treatment experiments. The seedlings were soaked in liquid BNM medium under different treatments. These treatments are -Sin/-NOA, +Sin/−NOA, −Sin/+NOA and +Sin/+NOA. The treatment –Sin/-NOA was used as mock treatment. (Sin = S. meliloti infection; NOA = 10 µM 1-NOA treatment; -Sin/-NOA: mock treatment). Then shoots and roots of M. truncatula seedlings were collected for RNA isolation respectively. Experiments were repeated for five biological times.
Identification of IAA genes in Medicago truncatula
The hidden Markov model (HMM) profile of the Aux/IAA protein family (Pfam: 02309 AUX/IAA family) was employed to identify the Aux/IAA genes from the M. truncatula genome. The profile was used to search the complete proteome of M. truncatula available in phytozome (http://www.phytozome.net/). All the obtained sequences were sorted as unique sequences for further Aux/IAA domains search using InterProScan Sequence Search (http://www.ebi.ac.uk/Tools/pfa/iprscan/). Linear display of synteny blocks was analyzed by the SyMAP database (http://www.symapdb.org/projects/fabaceae/)
Phylogenetic tree building, intron/exon structure, genome distribution and motif prediction
Multiple sequence alignments were performed on the MtIAA protein sequences using ClustalW with the default parameters, and the alignments were then adjusted manually. Phylogenetic tree was constructed with aligned MtIAA protein sequences using MEGA5.1 (http://www.megasoftware.net/mega5/mega.html) employing the neighbor-joining (NJ) method. Bootstrap values were calculated from 1000 iterations. The gene pairs displayed high bootstrap value (>99%) were identified as sister pair genes. The constructed tree file was visualized by TreeView1.6 (http://taxonomy.zoology.gla.ac.uk/rod/treeview.html). The software MEGA 5.1 was used for prediction four classical domains in MtIAA proteins (domain I, II, III, IV). The DNA and cDNA sequences corresponding to each predicted gene from the M. truncatula genome and the information of MtIAAs intron distribution pattern were obtained from the). To obtain the gene locations, we drew a map of the distribution of MtIAA genes throughout the M. truncatula genome using genome visualization tool CIRCOS (). M. truncatula chromosomes are arranged in a circle and the centromere of each chromosome is marked in black. Ribbon links represent the segmental duplication region retrieved from the SyMAP database. Motifs constitution of M. truncatula Aux/IAA proteins were investigated by MEME web server.
RNA isolation and qRT-PCR
The methods, including RNA extraction from various organs, reverse transcription to cDNA and qRT-PCR analysis, were performed according to Shen's publication[40]. Total RNA from cotyledons, leaves, shoots, roots and flowers was extracted using a Plant RNeasy Mini kit (Qiagen) according to the manufacturer's instructions. Then DNase I treatment was used to remove genomic DNA contamination from total RNA. The primer sequences are listed in Table S1. Mt-Actin (MTR_2g008050) was used as an internal standard to calculate the relative fold differences based on the comparative Ct method. 2−ΔΔCt refers to the fold difference in IAA expression compared with the untreated seedlings. Heat map representation was performed using the normalized Ct value with ClustalW software and Treeview to visualize the analysis data.
Bacterial strains and rhizobia infection
The rhizobia strain used for inoculating the roots of seedlings was S. meliloti strain 1021, a streptomycin-resistant derivative of wild-type. S. meliloti was grown overnight at 28°C in liquid LBMC medium (10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl, 2.6 mM MgSO4, 2.6 mM CaCl2) supplemented with 200 µg/mL streptomycin, then collected by centrifugation, and suspended in 10 mM MgSO4. The bacterial suspension was diluted with liquid BNM medium to OD600 = 0.1. For plant inoculation experiment, each seedling was put in petri dish containing nitrogen-free nutrient solution and treated with BNM medium diluted bacteria. For control (mock-inoculated), the seedlings were treated with sterilized BNM medium containing 10 mM MgSO4 without bacteria. Five different sets of plants were grown and used for the various analyses.
Analysis of AuxRE cis-elements
The promoters (−1000 to −1 bp before ATG) of MtIAA genes were scanned using Watson and Crick words for AuxRE related cis-element 1 (AUX1: TGTCTC core sequence), a less stringent variant called AUX2 (TGTVYS), three different bZIP Response Elements (ZREs) (bZIP-associated G-box Related Element (GRE): BACGTV, TGA: TGACG and AC-motif: ACTCAT) and two Myb Response Elements (MREs) (MRE1: AMCWAMC and MRE2: GGWTW). Different colors represent different cis-elements, which were added to the image manually. The promoter sequences were obtained from http://www.phytozome.net/.
Results
Genome-wide identification of IAA genes in Medicago truncatula
The new version (V3.5) of the M. truncatula genome and protein sequences was obtained from the M. truncatula genome database. Comprehensive searches of public genomic database using the hidden Markov model (HMM) profile of the IAA gene family (Pfam 02309: AUX/IAA family) was employed to identify the IAA genes selected from BLAST results. All the candidate Aux/IAA protein sequences were aligned by ClustalW and were checked manually. After protein domain and gene structure analysis, seventeen IAA genes were identified in M. truncatula. The information of these seventeen MtIAA genes, including gene names, locus IDs, intron numbers, ORF lengths, chromosome locations and basic parameters of deduced polypeptide, were listed in Table 1. The size of the deduced MtIAA proteins varies markedly ranging from 142 amino acids (MtIAA14) to 537 amino acids (MtIAA9), the corresponding molecular mass varies from 15.99 kDa to 62.45 kDa, and the predicted isoelectric point widely varies from 5.18 (MtIAA14) to 8.93 (MtIAA7). It suggested that different MtIAA proteins might function in different microenvironments.
Table 1. MtIAA gene family in Medicago truncatula.
| Gene | locus ID | ORF length | No. of introns | Chr no. | Chr location | Deduced polypeptide | ||
| Length (aa) | Mol wt (kDa) | pI | ||||||
| MtIAA1 | Medtr1g070830 | 615 | 2 | 1 | 17318116–17319328 | 204 | 22.91 | 6.42 |
| MtIAA2 | Medtr1g093240 | 711 | 4 | 1 | 26193287–26196287 | 236 | 25.72 | 8.46 |
| MtIAA3 | Medtr1g093350 | 537 | 1 | 1 | 26247246–26248612 | 178 | 19.98 | 8.00 |
| MtIAA4 | Medtr2g100780 | 810 | 4 | 2 | 32192283–32195676 | 269 | 29.96 | 8.73 |
| MtIAA5 | Medtr2g101500 | 981 | 4 | 2 | 32562344–32565049 | 326 | 35.36 | 7.96 |
| MtIAA6 | Medtr2g102490 | 597 | 4 | 2 | 33070172–33071309 | 198 | 22.5 | 8.62 |
| MtIAA7 | Medtr3g106850 | 642 | 3 | 3 | 37913558–37914702 | 213 | 24.73 | 8.94 |
| MtIAA8 | Medtr4g060470 | 1086 | 2 | 4 | 18656418–18658053 | 361 | 39.46 | 5.93 |
| MtIAA9 | Medtr4g115070 | 1614 | 4 | 4 | 39660114–39669519 | 537 | 62.45 | 6.93 |
| MtIAA10 | Medtr4g124300 | 555 | 3 | 4 | 43252133–43253003 | 184 | 20.88 | 6.83 |
| MtIAA11 | Medtr4g128070 | 522 | 1 | 4 | 44708010–44709260 | 173 | 19.22 | 7.91 |
| MtIAA12 | Medtr5g030710 | 1008 | 6 | 5 | 12694370–12699892 | 335 | 36.26 | 8.25 |
| MtIAA13 | Medtr5g067350 | 1044 | 5 | 5 | 27469789–27473618 | 335 | 36.26 | 8.25 |
| MtIAA14 | Medtr7g110790 | 429 | 2 | 7 | 35419184–35419916 | 142 | 15.99 | 5.18 |
| MtIAA15 | Medtr8g014520 | 816 | 4 | 8 | 3132788–3136332 | 271 | 30.12 | 8.19 |
| MtIAA16 | Medtr8g067530 | 1071 | 7 | 8 | 17235080–17239117 | 356 | 38.67 | 6.62 |
| MtIAA17 | Medtr8g103030 | 882 | 4 | 8 | 30131713–30134764 | 293 | 31.89 | 7.85 |
Chromosomal distribution and gene structure analysis of MtIAA genes
All seventeen MtIAA genes in M. truncatula are distributed on seven chromosomes, except for chromosome 6. The distribution of the MtIAA genes varies: there are three MtIAA genes on chromosome 1, 2 and 8; just a single MtIAA gene on chromosome 3 and 7; four MtIAA genes on chromosome 4; two MtIAA genes on chromosome 5 (Figure 1 and Table 1). The full-length cDNA sequences and genomic DNA sequences of MtIAA family genes were downloaded from phytozome 9.1 database (http://www.phytozome.net). The numbers and positions of exons-introns for each MtIAA genes were uncovered by a comparison of the full-length cDNA sequences with the corresponding genomic DNA sequences. The number of introns varied from 1 to 6 in the MtIAA gene family (Figure 2B). The MtIAA genes displayed complex distribution patterns of introns-exons even within the same phylogenetic group.
Figure 1. Chromosome mapping of MtIAA family genes.

The genome visualization tool CIRCOS was employed. Medicago truncatula chromosomes are arranged in a circle, and the centromere of each chromosome is marked in black. Ribbon links represent the segmental duplication region retrieved from the SyMAP database. MtIAA family genes are mapped by locus.
Figure 2. Phylogenetic relationships, exon-intron structure and protein domain analysis of MtIAA family genes.

(A) An unrooted phylogenetic tree was constructed using ClustalW by N-J method. The sister pair genes are indicated by red boxes. (B) Exon-intron structure analysis of MtIAA genes. The untranslated regions (UTRs) are indicated by thick red lines; the exons are indicated by blue boxes; the introns are indicated by gray lines. (C) Alignment of Medicago truncatula Aux/IAA proteins obtained with the ClustalW program and manual correction. Multiple alignments of the domains I–IV of the M. truncatula Aux/IAA proteins also were showed by red lines. Colorized shading indicates identical and conversed amino acid residues, respectively. The LXLXLX motif was also marked by thin yellow box. Two NLSs were marked by black asterisks.
Gene divergence and duplication events were the important causes for evolutionary momentum [41], [42]. Family genes duplication events including tandem and segmental duplications were used to reveal the expansion of M. truncatula IAA family genes during the evolutionary process. In this study, three sister-gene pairs have been showed in Figure 2A and these sister-gene pairs were: MtIAA12/MtIAA16, MtIAA4/MtIAA15 and MtIAA10/MtIAA14.
Protein structure and phylogenetic relationship analysis of MtIAAs
Most of the MtIAA family proteins contain four conserved domains: domain I, II, III and IV (Figure 2C). A typical LXLXLX motif was found in domain I of most MtIAA proteins, including MtIAA1-6, 9, 12, 13, 15, 16 and 17. Two different types of putative nuclear localization signals (NLS) were detected in most MtIAA proteins: a bipartite NLS and a typical NLS. The bipartite NLS contained two stretches of K/R residues. The short amino acid sequence KR is located between domain I and II and the second part of the bipartite NLS is located at the end of domain II. A typical NLS, which consisted of one cluster of largely changed amino acid residues such as lysine or arginine, located at the end of domain IV (Figure 2C) [16], [43].
To explore phylogenetic relationship among IAA proteins in different plant species, a phylogenetic tree was constructed including IAA family members from Arabidopsis and M. truncatula. The information of AtIAA gene family was listed in Table S3. The unrooted phylogenetic tree was generated by the alignment of full-length protein sequences of seventeen MtIAA proteins and twenty-nine AtIAA proteins [36], [44]. The phylogenetic distribution indicated that IAA proteins classed into five major groups named A, B, C, D and E with well supported bootstrap value. Groups A and B were further divided into several subgroups each: A1, A2, A3, A4 and A5; B1 and B2 (Figure 3A).
Figure 3. Phylogenetic relationships and motifs distribution analysis.

(A) Phylogenetic relationships among Arabidopsis and M. truncatula IAA proteins. The unrooted tree was generated by MEGA5.1 program by the N-J method. Bootstrap supports from 1000 replicates are showed at each branch. (B) The motif distribution in Arabidopsis and M. truncatula IAA proteins. Motifs of Aux/IAA proteins were analyzed by MEME web server. Four motifs representing four domains I, II, III and IV were mapped on all Aux/IAA proteins by different colors. (C) The heights of each box represent the specific amino acid conservation in each motif.
The motif distribution in Arabidopsis and M. truncatula Aux/IAA proteins were analyzed by Multiple Expectation Maximization for Motif Elicitation (MEME) tool (http://meme.nbcr.net/meme/cgi-bin/meme.cgi). Different conserved domains of Aux/IAA proteins were mapped on the Figure 3 by MEME tool. Most IAA proteins, which were belonged to Group A and B, contained four classical IAA domains. The IAA proteins with truncated domains always belonged to subfamilies C, D and E. Motif I cannot be found in MtIAA 7, 8, 10, 11 and 14. Motif II is missing in MtIAA 6, 8, 11 and 14. The domain II of MtIAA10 is modified (DWPPV). MtIAA11 does not display motif III and MtIAA14 is missing a part of domain IV (Figure 3B).
Expression patterns of MtIAA genes in different M. truncatula tissues
Analysis of transcriptional level of MtIAA genes in different M. truncatula tissues gives clues on the biological function of these auxin-responsive genes. Here, the spatio specificity expression of each member of the MtIAA family gene was examined in different organs including roots, stems, cotyledons and leaves of two weeks seedlings and flowers of two months plants using qRT-PCR. (The primer sequences of seventeen MtIAA genes were listed in Table S1). Transcript accumulations of seventeen MtIAA genes were detectable in most different organs (Figure 4). The mRNA abundances of MtIAA family genes in roots were much higher compared to other organs. Furthermore, the expression levels of MtIAA genes in flowers were lower compared to other organs. It suggested that MtIAA genes may function in root growth and development. Of particular interest, some MtIAA genes showed obviously tissue-specific expression patterns in M. truncatula. MtIAA6 and MtIAA7 exhibited root-specific expressions; MtIAA10, MtIAA11, MtIAA14 and MtIAA15 showed a higher expression level in cotyledons than that in other organs.
Figure 4. Tissues-specific expression patterns of MtIAA genes.

Based on the phylogenetic analysis, all MtIAA genes were grouped into five subfamilies (A: subfamily A; B: subfamily B; C: subfamily C; D: subfamily D; E: subfamily E). Expression patterns of the MtIAA genes in five indicated organs were analyzed by the data of qRT-PCR. The value of ACTIN (Cotyledon)/1000 defines as 1. C: cotyledon; L: leaf; R: root; S: shoot; F: flower. The data were analyzed by five independent repeats, and standard deviations were shown with error bars.
Auxin regulation expression of MtIAA genes in shoots and roots
As an essential compound of auxin signaling pathway, the expression of IAA genes showed quick responses to auxin treatment [45]. QRT-PCR was performed with total RNA isolated from shoots and roots of IAA-treated seedlings and mock seedlings. The data showed that most of MtIAA genes were responsive to exogenous IAA treatments. The MtIAA family genes showed different expression patterns under IAA treatments. The expressions of MtIAA7, MtIAA10, MtIAA11, MtIAA14, MtIAA15 and MtIAA17 were reduced by IAA treatments and the remaining ones were induced by IAA treatments in shoots (Figure 5A). On the other hand, the expression levels of MtIAA2-7, MtIAA17 were down-regulated by IAA treatments in roots (Figure 5B).
Figure 5. Real-time quantitative RT-PCR (qRT-PCR) analyses of MtIAA genes in plants under IAA treatment in both shoots (A) and roots (B).

Total RNA was extracted from the shoots and roots of M. truncatula seedlings for basal expression. The histogram shows the relative expression level of MtIAA genes under IAA treatment compared to the mock expression level. The relative mRNA level of individual genes was normalized with respect to the MtACTIN gene. The concentration of synthetic IAA was 0.1 µM. Samples of two different organs (shoots and roots) were used to test the changes of MtIAA genes expression level at different time points (3 hr, 6 hr, 12 hr and 24 hr). The data were analyzed by five independent repeats, and standard deviations were shown with error bars.
In M. truncatula, two opposite auxin-dependent expression patterns were observed under 0.1 µM IAA treatment. While the expression levels of MtIAA8, 9, 12, 13 and 16 were up-regulated, MtIAA7 and 17 were down-regulated in both roots and shoots. Many MtIAA genes displayed a quick response (3 h treatment) to exogenous auxin application, such as MtIAA8, 11, 12, 13, 14 and 16 in shoots and MtIAA1, 4, 5, 9, 10, 14 and 15 in roots during the time course (Figure 5).
In this work, five sampling time points (0, 3 hr, 6 hr, 12 hr and 24 hr) were used for test whether IAA genes in M. truncatula were auxin early response genes. The expression levels of most MtIAA family genes could be regulated by IAA under 3 hr treatment. The change folds of expression levels were almost the same from 3 hr IAA treatment to 24 hr IAA treatment. Specially, MtIAA1, MtIAA10 and MtIAA15 were induced by IAA treatment over 100 folds in roots. All the data suggested that most of MtIAA family genes were auxin early response genes.
The auxin-responsive cis-elements in the promoters of MtIAA family also were analyzed in this study. The detailed data of this promoter analysis were listed in Table S2. Totally, 11 GREs, 6 TGAs, 5 ACs, 8 AUX1s, 8 AUX2s, 17 MRE1s and 56 MRE2s were contained in all seventeen MtIAA promoters (Figure 6).
Figure 6. Motif analysis of specific cis-elements in promoters of MtIAA family genes in M. truncatula.
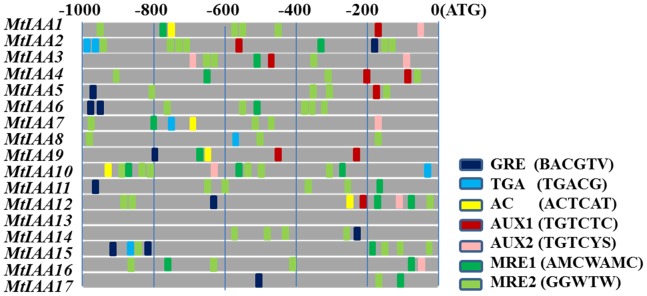
The −1000 bp promoter sequences of corresponding MtIAA genes were used to analysis of specific ZRE, AuxRE and MRE cis-elements, which are given using the presented colour code. Watson and Crick words for AUX1 is TGTCTC; AUX2 is TGTVYS, three different ZREs (GRE: BACGTV; TGA: TGACG; AC-motif: ACTCAT) and two MREs (MRE1: AMCWAMC; MRE2: GGWTW).
Expression analysis of MtIAA genes during the early phase of Sinorhizobium meliloti infection
Elevation of auxin transport between shoots and roots was the essential process for legume species to response to the rhizobial infection [31]. The expression levels of many auxin-related genes were changed by rhizobial infection during initial infection processes [46]. To reveal how auxin signaling was involved in the nodule formation after rhizobial infection, we examined the expression patterns of MtIAA genes in root and shoot of S. meliloti-inoculated M. truncatula seedlings within 72 hr post inoculation (hpi). First, we germinated the M. truncatula wild-type on filter papers, and afterwards transferred the seedlings to liquid medium. Total RNA from roots and shoots were isolated for qRT-PCR analysis. Student's t-test analysis between mock-inoculated plants and rhizobial-inoculated plants was performed to reveal the differential expression patterns of MtIAA family genes.
Differential expression patterns of MtIAA family genes were observed during the early phase of S. meliloti infection between shoots and roots in wild-type A17. Most MtIAA genes were down-regulated by S. meliloti infection in roots and were up-regulated in shoots. However, the expression levels of MtIAA6 and MtIAA17 were induced by S. meliloti infection in roots; MtIAA1, MtIAA6 and MtIAA7 were reduced by S. meliloti infection in shoots. The main expression pattern of MtIAA family genes in roots was opposite to that in shoots (Figure 7).
Figure 7. Heat map showing MtIAA genes expression pattern at the early phase of Sinorhizobium meliloti infection under different conditions.

Samples of two different organs (shoots and roots) were used to test the changes of MtIAA genes expression level at different time points (3/6/12/24/48/72 hpi) treatment. The different colors correspond to the log-transcription values of the gene change-fold ratio shown in the bar at the right of figure.
Here, we used 1-NOA (1-naphthoxyacetic acid), that binds to auxin influx transporter to block the auxin polar transport, to suppress the Aux/IAA-mediated auxin signaling in M. truncatula. The roots and shoots RNA samples of five time points (0, 3 hpi, 6 hpi, 12 hpi and 24 hpi) were separated into four independent groups: -Sin/-NOA, +Sin/-NOA, −Sin/+NOA and +Sin/+NOA. (Sin = S. meliloti infection; NOA = 10 µM 1-NOA treatment; -Sin/-NOA: mock treatment). The qRT-PCR data showed that expression levels of most MtIAA family genes displayed a drastic decline under +Sin/−NOA condition in roots and an obvious increase in shoots. Specially, the −Sin/+NOA treatment had no significant effect on expression regulation of MtIAA genes. The changes of MtIAA genes expression levels caused by S. meliloti infection were alleviated by 10 µM 1-NOA application both in roots and shoots (Figure 8).
Figure 8. Heat map showing MtIAA genes expression pattern at the early phase of Sinorhizobium meliloti infection under different conditions.

Samples of two different organs (shoots and roots) were used to test the changes of MtIAA genes expression level at different time points (3/6/12/24/48/72 hpi) and conditions (−Sin/−NOA, +Sin/−NOA, −Sin/+NOA and +Sin/+NOA). The treatment –Sin/−NOA was used as mock treatment. The different colors correspond to the log-transcription values of the gene change-fold ratio shown in the bar at the right of figure.
Discussion
As a key component of the auxin signaling pathway, Aux/IAA proteins directly bind to ARF proteins and regulate expression of down-stream auxin response genes [47], [48]. Functional characterization and expression analysis of Aux/IAA family genes help to reveal the mechanisms behind how auxin signaling is involved in plant growth and responses to environmental changes in a spatio-temporal specific manner. Phytohormone auxin is an important factor in regulation of mitosis, which is involved in nodulation [28], [49], [50]. As a model indeterminate legume, analysis of the protein structure and expression pattern of Aux/IAA in M. truncatula is a way to elucidate the relationship between auxin signaling and the symbiotic during the early phase of S. meliloti infection.
Characterization and structural analysis of the MtIAA family genes in Medicago truncatula
Aux/IAA proteins are plant specific transcriptional regulators [51]. In this study, seventeen MtIAA family genes were isolated in M. truncatula basing on the reference genome sequence (500 Mbp) [52]. The information of seventeen MtIAA genes was listed in Table 1. The number of MtIAA genes in M. truncatula is a little less compared to Arabidopsis (29 genes) or rice (31 genes) [27], [36]. Sequence analysis of the M. truncatula Aux/IAA family proteins revealed that some MtIAA proteins do not contain all classical domains of this protein family.
Most of MtIAA proteins contain conserved leucine residues in the LXLXLX motif as A. thaliana [9]. MtIAA1-6, 9, 12, 13 and 15–17 contain a typical LXLXLX motif “TELRLGLPG”. The LXLXLX motif plays an important role in repression of IAA proteins. Mutation in any of these three Leu in this motif results in total loss of repression or strongly reduced repression in the case of the mutation in the third Leu in the motif [9]. In this study, some MtIAA proteins (5 in 17) do not contain this motif and these MtIAA proteins may function differently to the classical IAA repressors. Four MtIAA proteins do not contain domain II, including MtIAA6, 8, 11 and 14 (Figure 3). There is much evidence that domain II functions as a key motif involved in Aux/IAA protein degradation, it is proposed to be functionally important due to its wide distribution in flowering plants [53]. In Arabidopsis, mutations in domain II leaded to enhanced auxin responses by blocking the degradation of IAA proteins [54]. However, in MtIAA protein family, some MtIAAs did not contain domain II. It suggested that the degradation of these MtIAA proteins were independent on domain II- mediate auxin responses. Most of the MtIAA proteins contain domain III–IV, which are the important domains for binding to ARFs. Specially, MtIAA11 does not contain domain II and MtIAA14 only contains a partial domain IV.
Auxin regulated gene expression
In the promoters of most auxin-responsive genes, auxin-responsive (AuxRE) cis-elements are characteristic structures [8]. Promoter analysis illustrated that most promoters of MtIAA genes contain AuxRE cis-elements or their variants (Figure 6) [55]. Our analysis showed that the motifs for AuxREs, ZREs and MREs related elements were significantly enriched in the promoters of MtIAA genes. All the promoters of MtIAA genes contained AuxRE cis-elements or their variants, expect for the promoter of MtIAA13, which did not contain any AuxRE cis-elements. Furthermore, take the classical AuxRE cis-elements AUX1 and AUX2 as examples. Ten of the seventeen MtIAA gene promoters contained more than one AUX1 or AUX2 within 1000 bp before ATG. It was indicating the expressions of MtIAA family genes could be regulated by auxin signaling.
In M. truncatula, six MtIAA genes were down-regulated by IAA treatment in shoot and 7 MtIAA genes were reduced by IAA treatment in root. The MtIAA family genes showed different expression patterns to response to exogenous auxin stimulation. Interestingly, the changes of expression levels of 5 MtIAA genes in shoot and 6 MtIAA genes in root were maintaining stability over time while the rest of MtIAA genes continuously increased or decreased expressions during the time course. The dynamic expressions of MtIAA family genes under IAA treatment indicated that different MtIAA genes are involved in the variability of auxin regulation.
To gain insight into the spatial pattern of the expressions of MtIAA genes, their transcript accumulations were analyzed in different plant tissues and organs. The clustering revealed five main clades: A, B, C, D and E. Most of the MtIAA family genes displayed the highest expression in roots. Interestingly, only MtIAA9 showed higher expression level in flower. No correlation was found between the clustering based on phylogenetic analysis and gene expression patterns. Some MtIAA genes displayed obvious preferential expression in a specific tissue suggesting the regulation of these genes might function essentially at the post-translational level [34]. Overall, the root-preferential expression of MtIAA family genes is indicative of their involvement in root developmental processes and responses to environmental stimulus.
MtIAAs were involved in nodule formation during the early phase of Sinorhizobium meliloti infection
Nitrogen-fixing nodule is the essential organ for symbiotic interactions between legumes and rhizobia. Formation of nodule helps soil rhizobia to convert atmospheric N2 into ammonia for M. truncatula absorption [56]. Rhizobial infection, which leads to signal exchanges between the hosts and the bacteria, is the first step for nodule organogenesis and many transcriptome and proteome changes occur in both the shoots and roots during the early stage of rhizobial infection [46], [57]. Auxin plays an important role in the initiation and development of nodules of different legumes, including white clover, Lotus japonicas and M. truncatula [57], [58]. A Lotus japonicus mutant rel3 exhibited insensitivity to auxin and produced fewer nitrogen-fixing nodules [56]. Here, we used M. truncatula as a model indeterminate legume to study how Aux/IAA-mediated auxin signaling is involved in nodule formation during the early phase of S. meliloti infection.
Auxin regulates down-stream genes transcription by promoting the degradation rate of the Aux/IAA family proteins, which function as transcriptional repressors [59]. The rhizobial-regulation expressions of MtIAA family genes trigger physiological responses in a spatio-temporal specific manner during the formation of nodules. Our qRT-PCR data showed that most MtIAA genes were involved in the inoculation of roots with the nodulating symbiont (S. meliloti). The expression profiles of MtIAA genes changed significantly during S. meliloti infection and MtIAA-mediated auxin signaling may activate or suppress the functions of many down-stream genes involved in the formation of nodules. Inoculation of M. truncatula roots reduces endogenous auxin loading from shoots to roots [60]. Most of MtIAA family genes were down-regulated in roots by S. meliloti infection and up-regulated in shoots. S. meliloti infection may suppress the MtIAA-mediated auxin signaling in roots by controlling auxin relocation between shoots and root system. Our results revealed that the changes of expression levels of MtIAA family genes occurred at early time points after S. meliloti infection (Figure 7). The expression of most MtIAA genes was found to be involved in the inoculation of roots with S. meliloti, which suggests a putative role in the formation of nitrogen-fixing nodules. In comparison to mock inoculation, MtIAA-mediated auxin signaling may activate or suppress the down-stream genes involved in the formation of nodules under S. meliloti infection.
Long distance of auxin polar transport between shoots and roots is the essential process for formation of nodules in M. truncatula [60]. Deviant nodules that lack vascular strands or with proliferating vascular tissue could be induced by auxin transport inhibitors like naphthylphthalamic acid (NPA) [33], [61]. It suggested that polar auxin transport plays a central role in vascular bundle formation in nodules. In this work, an auxin influx inhibitor 1-NOA [62], was used to interfere with MtIAA-mediated auxin signaling during the early phase of S. meliloti infection. To test how MtIAA-mediated auxin signaling plays a role in nodule formation during the early phase of S. meliloti infection, we analyze the qRT-PCR data of MtIAA genes expression levels under –Sin/−NOA, +Sin/−NOA, −Sin/+NOA and +Sin/+NOA four treatments respectively. The expression profiling of MtIAA family genes did not change too much in both shoots and roots under 1-NOA treatment only (−Sin/+NOA). 1-NOA treatment (+Sin/+NOA treatment) reduced the differences in MtIAA gene expression between shoots and roots compared to +Sin/−NOA inoculated plants. S. meliloti infection has a significant effect on the MtIAA-mediated auxin signaling pathway. However, inhibition of auxin transport relieved the differences in expressions of MtIAA genes between roots and shoots during the early phase of S. meliloti infection. S. meliloti infection first triggered the MtIAA-medicated auxin signaling to help nodule initiation and differential MtIAA-mediated auxin signaling between shoots and roots may be an essential process for expression regulation of nodule-related downstream genes.
Conclusion
M. truncatula, a close relative of alfalfa, is a model legume widely used in nitrogen fixation, symbiosis and legume genomics studies [63]. In summary, the detailed analysis of MtIAA family genes provides new insights into the structure and expression of this gene family that plays an important role in auxin signaling and gene expression regulation of M. truncatula under different conditions. The involvement of MtIAA genes during the early phase of S. meliloti infection helps us to understand the role of auxin signaling in the regulation of nodule formation.
Supporting Information
The qRT-PCR primers for MtIAA family genes.
(XLS)
The location information of AuxRE cis -elements in MtIAA family gene promoters.
(XLS)
The information of AtIAA gene family.
(XLSX)
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was funded by the Zhejiang Provincial Natural Science Foundation of China under Grant No. LQ14C060001, LQ14C020002 and LQ13C020004. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Zhao Y (2010) Auxin biosynthesis and its role in plant development. Annu Rev Plant Biol 61: 49–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Ljung K (2013) Auxin metabolism and homeostasis during plant development. Development 140: 943–950. [DOI] [PubMed] [Google Scholar]
- 3. Chung Y, Maharjan PM, Lee O, Fujioka S, Jang S, et al. (2011) Auxin stimulates DWARF4 expression and brassinosteroid biosynthesis in Arabidopsis. Plant J 66: 564–578. [DOI] [PubMed] [Google Scholar]
- 4. Abel S, Theologis A (1996) Early genes and auxin action. Plant Physiol 111: 9–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Dharmasiri N, Dharmasiri S, Estelle M (2005) The F-box protein TIR1 is an auxin receptor. Nature 435: 441–445. [DOI] [PubMed] [Google Scholar]
- 6. Tan X, Calderon-Villalobos LI, Sharon M, Zheng C, Robinson CV, et al. (2007) Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature 446: 640–645. [DOI] [PubMed] [Google Scholar]
- 7. Tiwari SB, Wang XJ, Hagen G, Guilfoyle TJ (2001) AUX/IAA proteins are active repressors, and their stability and activity are modulated by auxin. Plant Cell 13: 2809–2822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Hagen G, Guilfoyle T (2002) Auxin-responsive gene expression: genes, promoters and regulatory factors. Plant Mol Biol 49: 373–385. [PubMed] [Google Scholar]
- 9. Tiwari SB, Hagen G, Guilfoyle TJ (2004) Aux/IAA proteins contain a potent transcriptional repression domain. Plant Cell 16: 533–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Szemenyei H, Hannon M, Long JA (2008) TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 319: 1384–1386. [DOI] [PubMed] [Google Scholar]
- 11. Ouellet F, Overvoorde PJ, Theologis A (2001) IAA17/AXR3: biochemical insight into an auxin mutant phenotype. Plant Cell 13: 829–841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Kim J, Harter K, Theologis A (1997) Protein-protein interactions among the Aux/IAA proteins. Proc Natl Acad Sci U S A 94: 11786–11791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Ulmasov T, Murfett J, Hagen G, Guilfoyle TJ (1997) Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 9: 1963–1971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Korasick DA, Westfall CS, Lee SG, Nanao MH, Dumas R, et al. (2014) Molecular basis for AUXIN RESPONSE FACTOR protein interaction and the control of auxin response repression. Proc Natl Acad Sci U S A 111: 5427–5432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Abel S, Oeller PW, Theologis A (1994) Early auxin-induced genes encode short-lived nuclear proteins. Proc Natl Acad Sci U S A 91: 326–330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Wu J, Peng Z, Liu S, He Y, Cheng L, et al. (2012) Genome-wide analysis of Aux/IAA gene family in Solanaceae species using tomato as a model. Mol Genet Genomics 287: 295–211. [DOI] [PubMed] [Google Scholar]
- 17. Yang X, Lee S, So JH, Dharmasiri S, Dharmasiri N, et al. (2004) The IAA1 protein is encoded by AXR5 and is a substrate of SCF(TIR1). Plant J 40: 772–782. [DOI] [PubMed] [Google Scholar]
- 18. Uberti-Manassero NG, Lucero LE, Viola IL, Vegetti AC, Gonzalez DH (2012) The class I protein AtTCP15 modulates plant development through a pathway that overlaps with the one affected by CIN-like TCP proteins. J Exp Bot 63: 809–823. [DOI] [PubMed] [Google Scholar]
- 19. Uehara T, Okushima Y, Mimura T, Tasaka M, Fukaki H (2008) Domain II mutations in CRANE/IAA18 suppress lateral root formation and affect shoot development in Arabidopsis thaliana. Plant Cell Physiol 49: 1025–1038. [DOI] [PubMed] [Google Scholar]
- 20. Rinaldi MA, Liu J, Enders TA, Bartel B, Strader LC (2012) A gain-of-function mutation in IAA16 confers reduced responses to auxin and abscisic acid and impedes plant growth and fertility. Plant Mol Biol 79: 359–373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Tatematsu K, Kumagai S, Muto H, Sato A, Watahiki MK, et al. (2004) MASSUGU2 encodes Aux/IAA19, an auxin-regulated protein that functions together with the transcriptional activator NPH4/ARF7 to regulate differential growth responses of hypocotyl and formation of lateral roots in Arabidopsis thaliana. Plant Cell 16: 379–393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Okushima Y, Fukaki H, Onoda M, Theologis A, Tasaka M (2007) ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 19: 118–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Fukaki H, Tameda S, Masuda H, Tasaka M (2002) Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J 29: 153–168. [DOI] [PubMed] [Google Scholar]
- 24. Song Y, You J, Xiong L (2009) Characterization of OsIAA1 gene, a member of rice Aux/IAA family involved in auxin and brassinosteroid hormone responses and plant morphogenesis. Plant Mol Biol 70: 297–309. [DOI] [PubMed] [Google Scholar]
- 25. Zhu ZX, Liu Y, Liu SJ, Mao CZ, Wu YR, et al. (2012) A gain-of-function mutation in OsIAA11 affects lateral root development in rice. Mol Plant 5: 154–161. [DOI] [PubMed] [Google Scholar]
- 26. Jun N, Gaohang W, Zhenxing Z, Huanhuan Z, Yunrong W, et al. (2011) OsIAA23-mediated auxin signaling defines postembryonic maintenance of QC in rice. Plant J 68: 433–442. [DOI] [PubMed] [Google Scholar]
- 27. Nakamura A, Umemura I, Gomi K, Hasegawa Y, Kitano H, et al. (2006) Production and characterization of auxin-insensitive rice by overexpression of a mutagenized rice IAA protein. Plant J 46: 297–306. [DOI] [PubMed] [Google Scholar]
- 28. van Noorden GE, Kerim T, Goffard N, Wiblin R, Pellerone FI, et al. (2007) Overlap of proteome changes in Medicago truncatula in response to auxin and Sinorhizobium meliloti. Plant Physiol 144: 1115–1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Nallu S, Silverstein KA, Samac DA, Bucciarelli B, Vance CP, et al. (2013) Regulatory patterns of a large family of defensin-like genes expressed in nodules of Medicago truncatula. PLoS One 8: e60355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Kondorosi E, Redondo-Nieto M, Kondorosi A (2005) Ubiquitin-mediated proteolysis. To be in the right place at the right moment during nodule development. Plant Physiol 137: 1197–1204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Deinum EE, Geurts R, Bisseling T, Mulder BM (2012) Modeling a cortical auxin maximum for nodulation: different signatures of potential strategies. Front Plant Sci 3: 96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I, et al. (2005) The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433: 39–44. [DOI] [PubMed] [Google Scholar]
- 33. Rightmyer AP, Long SR (2011) Pseudonodule formation by wild-type and symbiotic mutant Medicago truncatula in response to auxin transport inhibitors. Mol Plant Microbe Interact 24: 1372–1384. [DOI] [PubMed] [Google Scholar]
- 34. Audran-Delalande C, Bassa C, Mila I, Regad F, Zouine M, et al. (2012) Genome-wide identification, functional analysis and expression profiling of the Aux/IAA gene family in tomato. Plant Cell Physiol 53: 659–672. [DOI] [PubMed] [Google Scholar]
- 35. Wang Y, Deng D, Bian Y, Lv Y, Xie Q (2010) Genome-wide analysis of primary auxin-responsive Aux/IAA gene family in maize (Zea mays. L.). Mol Biol Rep 37: 3991–4001. [DOI] [PubMed] [Google Scholar]
- 36. Jain M, Kaur N, Garg R, Thakur JK, Tyagi AK, et al. (2006) Structure and expression analysis of early auxin-responsive Aux/IAA gene family in rice (Oryza sativa). Funct Integr Genomics 6: 47–59. [DOI] [PubMed] [Google Scholar]
- 37. Wu J, Liu S, Guan X, Chen L, He Y, et al. (2014) Genome-wide identification and transcriptional profiling analysis of auxin response-related gene families in cucumber. BMC Res Notes 7: 218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Ludwig Y, Zhang Y, Hochholdinger F (2013) The maize (Zea mays L.) AUXIN/INDOLE-3-ACETIC ACID gene family: phylogeny, synteny, and unique root-type and tissue-specific expression patterns during development. PLoS One 8: e78859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Engstrom EM, Ehrhardt DW, Mitra RM, Long SR (2002) Pharmacological analysis of nod factor-induced calcium spiking in Medicago truncatula. Evidence for the requirement of type IIA calcium pumps and phosphoinositide signaling. Plant Physiol 128: 1390–1401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Shen C, Wang S, Zhang S, Xu Y, Qian Q, et al. (2013) OsARF16, a transcription factor, is required for auxin and phosphate starvation response in rice (Oryza sativa L.). Plant Cell Environ 36: 607–620. [DOI] [PubMed] [Google Scholar]
- 41. Vision TJ, Brown DG, Tanksley SD (2000) The origins of genomic duplications in Arabidopsis. Science 290: 2114–2117. [DOI] [PubMed] [Google Scholar]
- 42. Bowers JE, Chapman BA, Rong J, Paterson AH (2003) Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 422: 433–438. [DOI] [PubMed] [Google Scholar]
- 43. Raikhel N (1992) Nuclear targeting in plants. Plant Physiol 100: 1627–1632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Remington DL, Vision TJ, Guilfoyle TJ, Reed JW (2004) Contrasting modes of diversification in the Aux/IAA and ARF gene families. Plant Physiol 135: 1738–1752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Quint M, Gray WM (2006) Auxin signaling. Curr Opin Plant Biol 9: 448–453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Molesini B, Cecconi D, Pii Y, Pandolfini T (2014) Local and systemic proteomic changes in medicago truncatula at an early phase of Sinorhizobium meliloti infection. J Proteome Res 13: 408–421. [DOI] [PubMed] [Google Scholar]
- 47.Kim J, Lee HW (2013) Direct activation of EXPANSIN14 by LBD18 in the gene regulatory network of lateral root formation in Arabidopsis. Plant Signal Behav 8. [DOI] [PMC free article] [PubMed]
- 48. Cho H, Ryu H, Rho S, Hill K, Smith S, et al. (2014) A secreted peptide acts on BIN2-mediated phosphorylation of ARFs to potentiate auxin response during lateral root development. Nat Cell Biol 16: 66–76. [DOI] [PubMed] [Google Scholar]
- 49. Wasson AP, Pellerone FI, Mathesius U (2006) Silencing the flavonoid pathway in Medicago truncatula inhibits root nodule formation and prevents auxin transport regulation by rhizobia. Plant Cell 18: 1617–1629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Plet J, Wasson A, Ariel F, Le Signor C, Baker D, et al. (2011) MtCRE1-dependent cytokinin signaling integrates bacterial and plant cues to coordinate symbiotic nodule organogenesis in Medicago truncatula. Plant J 65: 622–633. [DOI] [PubMed] [Google Scholar]
- 51. Reed JW (2001) Roles and activities of Aux/IAA proteins in Arabidopsis. Trends Plant Sci 6: 420–425. [DOI] [PubMed] [Google Scholar]
- 52. Young ND, Debelle F, Oldroyd GE, Geurts R, Cannon SB, et al. (2011) The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 480: 520–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Paponov IA, Teale W, Lang D, Paponov M, Reski R, et al. (2009) The evolution of nuclear auxin signalling. BMC Evol Biol 9: 126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Dreher KA, Brown J, Saw RE, Callis J (2006) The Arabidopsis Aux/IAA protein family has diversified in degradation and auxin responsiveness. Plant Cell 18: 699–714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Berendzen KW, Weiste C, Wanke D, Kilian J, Harter K, et al. (2012) Bioinformatic cis-element analyses performed in Arabidopsis and rice disclose bZIP- and MYB-related binding sites as potential AuxRE-coupling elements in auxin-mediated transcription. BMC Plant Biol 12: 125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Li X, Lei M, Yan Z, Wang Q, Chen A, et al. (2013) The REL3-mediated TAS3 ta-siRNA pathway integrates auxin and ethylene signaling to regulate nodulation in Lotus japonicus. New Phytol. [DOI] [PubMed]
- 57. Madsen LH, Tirichine L, Jurkiewicz A, Sullivan JT, Heckmann AB, et al. (2010) The molecular network governing nodule organogenesis and infection in the model legume Lotus japonicus. Nat Commun 1: 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Mathesius U, Schlaman HR, Spaink HP, Of Sautter C, Rolfe BG, et al. (1998) Auxin transport inhibition precedes root nodule formation in white clover roots and is regulated by flavonoids and derivatives of chitin oligosaccharides. Plant J 14: 23–34. [DOI] [PubMed] [Google Scholar]
- 59. Han X, Xu X, Fang DD, Zhang T, Guo W (2012) Cloning and expression analysis of novel Aux/IAA family genes in Gossypium hirsutum. Gene 503: 83–91. [DOI] [PubMed] [Google Scholar]
- 60. van Noorden GE, Ross JJ, Reid JB, Rolfe BG, Mathesius U (2006) Defective long-distance auxin transport regulation in the Medicago truncatula super numeric nodules mutant. Plant Physiol 140: 1494–1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Takanashi K, Sugiyama A, Yazaki K (2011) Involvement of auxin distribution in root nodule development of Lotus japonicus. Planta 234: 73–81. [DOI] [PubMed] [Google Scholar]
- 62. Parry G, Delbarre A, Marchant A, Swarup R, Napier R, et al. (2001) Novel auxin transport inhibitors phenocopy the auxin influx carrier mutation aux1. Plant J 25: 399–406. [DOI] [PubMed] [Google Scholar]
- 63. Tang H, Krishnakumar V, Bidwell S, Rosen B, Chan A, et al. (2014) An improved genome release (version Mt4.0) for the model legume Medicago truncatula. BMC Genomics 15: 312. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The qRT-PCR primers for MtIAA family genes.
(XLS)
The location information of AuxRE cis -elements in MtIAA family gene promoters.
(XLS)
The information of AtIAA gene family.
(XLSX)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.