

Abstract
Lactobacillus colonization of the lower female genital tract provides protection from the acquisition of sexually transmitted diseases, including human immunodeficiency virus, and from adverse pregnancy outcomes. While glycogen in vaginal epithelium is thought to support Lactobacillus colonization in vivo, many Lactobacillus isolates cannot utilize glycogen in vitro. This study investigated how glycogen could be utilized by vaginal lactobacilli in the genital tract. Several Lactobacillus isolates were confirmed to not grow in glycogen, but did grow in glycogen-breakdown products, including maltose, maltotriose, maltopentaose, maltodextrins, and glycogen treated with salivary α-amylase. A temperature-dependent glycogen-degrading activity was detected in genital fluids that correlated with levels of α-amylase. Treatment of glycogen with genital fluids resulted in production of maltose, maltotriose, and maltotetraose, the major products of α-amylase digestion. These studies show that human α-amylase is present in the female lower genital tract and elucidates how epithelial glycogen can support Lactobacillus colonization in the genital tract.
Keywords: α-amylase, female genital tract, glycogen, Lactobacillus, maltose
Colonization of the lower genital tract with Lactobacillus is an important component of the reproductive health of women. Lactobacillus inhibits other microbes in the genital tract, primarily by creating a low genital pH through the production of lactic acid [1–4]. This inhibition results in a genital microbiota that is predominantly composed of Lactobacillus in many women.
Factors that disrupt Lactobacillus colonization can lead to a shift of the genital microbiota to bacterial vaginosis (BV) in which Lactobacillus is no longer the predominant type of bacteria [1, 5–7]. Epidemiologic studies show that a genital microbiota dominated by Lactobacillus is protective from the acquisition of human immunodeficiency virus (HIV) when compared to BV (reviewed in [8]). In HIV-infected women, a Lactobacillus-dominated microbiota is also associated with decreased shedding of virus, decreased risk of heterosexual transmission to men, and vertical transmission to the fetus during birth [9–14]. Further, a genital microbiota dominated by Lactobacillus is associated with a lowered risk of acquiring other sexually transmitted diseases (STDs), such as herpes simplex virus type 2, chlamydia, and gonorrhea [15, 16]. Finally, Lactobacillus also protects against developing BV, pelvic inflammatory disease, and adverse fetal outcomes [17–21]. Certain species of Lactobacillus have been found to be more protective [4, 22].
Although Lactobacillus is recognized as important for genital tract health, the factors responsible for its colonization are not well understood. For example, while BV can be treated with antibiotics, Lactobacillus does not always become reestablished as the dominant bacteria type, and BV recurs in 60%–80% of cases after 1 year [23, 24]. However, a relationship between vaginal epithelial glycogen and Lactobacillus colonization in the female lower genital tract has been recognized for many decades. Thus, Cruickshank and others [25, 26] observed that at puberty, glycogen becomes expressed in the vaginal epithelium and at the same time colonization with Lactobacillus becomes apparent. Those studies showed that both glycogen expression and colonization by Lactobacillus persist through life until at menopause, when both Lactobacillus colonization and glycogen expression decline.
Despite the evidence that glycogen promotes colonization by lactobacilli, most isolates of Lactobacillus do not grow in vitro in media containing glycogen as the carbohydrate source. Thus, Stewart-Tull [27] reported in 1964 that a number of isolates of Lactobacillus did not ferment glycogen. Later in that decade, Wylie and Henderson [28] showed that 39 of 42 isolates of Lactobacillus did not ferment glycogen, even when the glycogen was isolated from the genital tract of women. More recently, Martin et al [29] showed 45 strains of vaginal Lactobacillus could not use glycogen. The lack of Lactobacillus growth in vitro in the presence of glycogen leaves open the question of how glycogen could be utilized in vivo by Lactobacillus. The goal of this study was to investigate how glycogen could be utilized by genital lactobacilli in the environment of the lower female genital tract.
MATERIALS AND METHODS
Subjects
Genital cervical-vaginal lavage (CVL) samples were obtained from women who had provided written informed consent. All procedures followed Department of Health and Human Service guidelines. The study was approved by the Cook County Health and Hospitals System Institutional Review Board. Samples were collected weekly over a 1–3 month period. All women reported not douching and not engaging in sex within the 48 hours before sample collection. Trichomonas, yeast, and BV were not detected at the time of sample collection as determined by wet mount, potassium hydroxide, and Amsel criteria [30]. The CVL samples were obtained by irrigation of the cervix with 10 mL of nonbacteriostatic sterile saline, followed by aspiration from the posterior fornix.
Bacterial Cultures
All reagents were purchased from Sigma (Sigma-Aldrich, St Louis, MO) unless noted. A basal de Man, Rogosa, and Sharpe (MRS) medium contained proteose peptone #3 (10.0 g, Remel, Thermo Fisher Scientific, Lenexa, KS), beef extract (10.0 g, Remel), yeast extract (5.0 g, Fisher), sorbitan monooleate (1.0 g), ammonium citrate (2.0 g), sodium acetate (5.0 g), MnSo4xH2O (0.05 g), and Na2HPO4 (2.0 g) per liter. Medium was supplemented with 2% glucose or other carbohydrates, including glycogen (from oyster), maltodextrins, maltose, maltotriose, and maltopentaose. In some experiments, normal saliva collected from 1 donor was immediately filtered (0.45 µm), and aliquots frozen. Upon thawing, saliva was added to glycogen-containing MRS medium immediately before the addition of bacteria to give concentrations of 2.5%, or 0.25% of the culture volume. Most cultures were in ambient air, except some run under anaerobic conditions using the Anaero Pack System (Mitsubishi Gas Chemical Co, Tokyo).
Bacteria tested included Lactobacillus jensenii (ATCC #25258), L. gasseri (ATCC #9857), and L. johnsonii (ATCC # 33200). Cultures (4 mL) were initiated from frozen stocks with 50 µL phosphate-buffered saline–washed bacteria at time 0 and the optical density (OD) at 600 nm was taken at 24-hour intervals.
Glycogen Degradation by Genital Fluids
Genital fluid samples collected by CVL were centrifuged at 4°C in a microfuge for 10 minutes to pellet bacteria. The supernatants (25 µL) were added to oyster glycogen (75 µL of 5 mg/mL) dissolved in phenol-red-free Roswell Park Memorial Institute 1640 medium. Aliquots were incubated at either 37°C or 4°C for 90 minutes and the amount of glycogen then measured using a colorimetric assay [31] by adding 50 µL of each of the following solutions: 10% potassium iodide, 0.01 N potassium iodate, and 1 N acetate. The mixture was incubated for 10 minutes and read at 565 nm. In some experiments, no extra glycogen was added to genital fluid samples so that degradation of endogenous glycogen could be observed.
Analysis of Oyster Glycogen Degradation Products
Oyster glycogen (20 mg/mL, weight per volume [w/v]) was hydrolyzed by adding saliva (see above) at 2.5% or 0.5% of the final volume for 10, 30, and 120 minutes at 37°C . Hydrolyzed samples were diluted 10× with deionized water. Molecular weight (MW) distributions of salivary α-amylase-treated glycogen samples were obtained by high-pressure size exclusion chromatography (HPSEC) using a Varian model 9012 high-performance liquid chromatography pump system connected to a Varian Star 9040 refractive index detector (Agilent Technologies, Santa Clara, CA) at room temperature. Injected samples (100 µL, 0.45 µm filtered) were separated using Sephacryl TM S-500 HR gel filtration (GE Healthcare, Piscataway, NJ). The eluent was purified water (18.2 MΩ) with 0.02% sodium azide [32]. Pullulan standards (Polymer Laboratories Inc, Amherst, MA) were used as a standard curve.
Oligosaccharide Analysis by High-Performance Anion-Exchange Chromatography
Hydrolysis products from oyster glycogen treated with either the collected saliva or the vaginal fluids were determined by a high-performance anion-exchange chromatography (HPAEC) system equipped with an electrochemical detector (Dionex, Sunnyvale, CA). The filtered (0.45 µm) samples were separated using a CarboPac PA-1 pellicular anion-exchange column (Dionex) with gradient elution from 100% eluent A (150 mM NaOH) to 100% eluent B (600 mM NaOAc in 150 mM NaOH) [33].
Bacterial Polymerase Chain Reactions
Bacteria specific quantitative polymerase chain reaction polymerase chain reaction (qPCR) assays were performed on isolated CVL genomic DNA [34]. Each 20 µL qPCR reaction contained Supermix (Bio-Rad Hercules, CA), primers, probes (IDT, Coralville, IA), and 1–10 ng template DNA. Primers and probes for the bacteria were described previously [34, 35]. Known quantities of 16S rRNA plasmid targets were used as standards [34, 35].
Enzyme Detection
Enzyme-linked immunosorbent assay (ELISA) was used to quantify human pancreatic α-amylase (AbCam, Cambridge, UK) and acidic α-glucosidase (MyBiosource, San Diego, CA).
RESULTS
Growth of Lactobacillus in Glucose, Glycogen, and Other Glucose Polymers
We first performed experiments to confirm previous reports that some genital Lactobacillus isolates do not grow in media with glycogen as the source of carbohydrate [27–29]. L. gasseri (ATCC #9857) was cultured in medium containing either 2% glucose, 2% glycogen, or no added carbohydrate (Figure 1). Over a 72-hour period, a relatively large increase in absorbance was observed for glucose cultures, while the ODs of cultures with either no carbohydrate or glycogen were similar. A similar lack of growth in glycogen when compared with no carbohydrate was observed with L. jensenii (ATCC #25258) and L. johnsonii (ATCC # 33200) (Figure 2). The lack of growth in glycogen was seen under both aerobic and anaerobic conditions (data not shown). Soluble starch (2% w/v) was also tested because its structure is similar to that of glycogen. No growth was observed using starch as the sole carbohydrate (Figure 1).
Figure 1.

Lack of L. gasseri growth in glycogen. L. gasseri was cultured in MRS medium that contained either no added carbohydrates (No Carb), 2% glucose, 2% glycogen, or 2% starch for 72 hours. The OD at 600 was read each day. Each culture condition was run in duplicate and the duplicates averaged. Each culture experiment was performed at least 3 times with similar results. Abbreviations: MRS, de Man, Rogosa, and Sharpe; OD, optical density.
Figure 2.

Growth of lactobacilli in glucose polymers. L. gasseri (A), L. jensenii (B and D), and L. johnsonii (C) were cultured in MRS medium that contained either no added carbohydrates (No Carb) or 2% of either glucose, maltose, maltodextrin 4–7, maltodextrin 13–17, maltodextrin 16–19, maltotriose, or maltopentaose for 72 hours. The OD at 600 was read each day. Abbreviations: MD, maltodextrin; MRS, de Man, Rogosa, and Sharpe; OD, optical density.
The lactobacilli tested above were able to utilize glucose for growth, but not glycogen, a high-molecular-weight polymer of glucose. Smaller polymers of glucose were also tested for their ability to support growth of lactobacilli. All 3 isolates of Lactobacillus were able to use 2% maltose for growth (Figure 2A–C). L. jensenii grew to relatively high levels by 48 hours in both maltotriose (polymer of 3 glucose units) and maltopentaose (5 glucose units) (Figure 2D).
Maltodextrins are derived from the partial hydrolysis of starch, and maltodextrins that are the least hydrolyzed have the lowest dextrose equivalent (DE) values. Growth of L. gasseri, L. jensenii, and L. johnsonii was also assessed in media containing 2% of 1 of 3 different maltodextrin preparations; DE 4–7, 13–17, and 16–19. All 3 of the species grew in the maltodextrins as a source of carbohydrates (Figure 2A–C), although the level of growth was generally less than that for glucose.
Growth of Lactobacilli in α-Amylase-Treated Glycogen
The above experiments indicated that some genital lactobacilli cannot utilize glycogen or starch for growth but can utilize smaller glucose polymers (dimers, trimers, pentamers). Therefore, the effect of glycogen breakdown by α-amylase on Lactobacillus growth over 72 hours was tested by adding a small amount of filtered saliva (2.5% or 0.25%) to the culture medium as a source of α-amylase. While L. gasseri did not grow in glycogen alone, it grew at a rate similar to cultures with glucose when cultured in glycogen with 2.5% filtered saliva added (Figure 3A). No growth above the control was seen with 2.5% saliva only. Similar rapid growth of L. gasseri was seen when 0.25% saliva was added to glycogen in cultures (data not shown).
Figure 3.
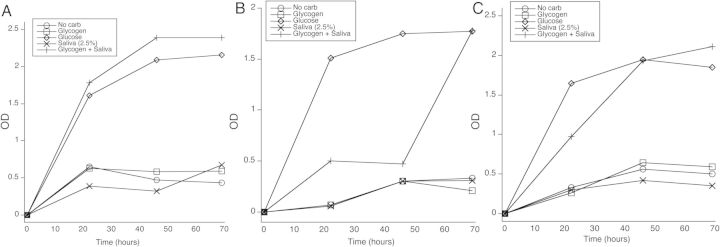
Growth of lactobacilli in salivary α-amylase-treated glycogen. L. gasseri (A), L. jensenii (B), or L. johnsonii (C) were cultured in MRS broth that contained either no added carbohydrates (No Carb), 2% glucose, 2% glycogen, 2.5% saliva alone, or glycogen plus saliva for 72 hours. The OD at 600 was read each day. Abbreviations: MRS, de Man, Rogosa, and Sharpe; OD, optical density.
Growth of L. johnsonii in glycogen with 2.5% saliva was also similar to growth in glucose (Figure 3B). In contrast, L. jensenii grew slowly in glycogen with saliva for the first 48 hours when compared to glucose, but growth increased by 72 hours to a level similar to glucose (Figure 3C).
The characteristics of the saliva-treated and control-treated glycogen were analyzed (Figure 4). HPSEC showed a peak at approximately 45 minutes that corresponded to undegraded glycogen. As little as 10 minutes of incubation with 2.5% saliva resulted in the appearance of a peak at 69 minutes, which was not present in untreated glycogen (Figure 4A). Longer incubation times (30 minutes, 120 minutes) led to an increase in the size of this peak. Analysis of the smaller carbohydrates in the 120-minute tube by HPAEC (Figure 4B) showed the appearance of peaks corresponding to maltose (G2) and maltotriose (G3) with some maltotetraose (G4), which are known major products of the α-amylase reaction [36, 37].
Figure 4.

Degradation of glycogen by salivary α-amylase. Glycogen (2% oyster) was incubated for 10 minutes with no saliva (Oyster) or for 10, 30, or 120 minutes with 2.5% saliva. The size of the degraded glycogen was determined by HPSEC (A) and the products analyzed by HPAEC (B). Abbreviations: HPAEC, high-performance anion-exchange chromatography; HPSEC, high-pressure size exclusion chromatography.
Vaginal Fluid From Normal Women Contains an Activity That Degrades Glycogen
The above experiments showed that salivary α-amylase could process glycogen into products that could be used by vaginal lactobacilli for growth. Experiments were next performed to determine if any α-amylase-like activity was present in the genital fluid of women [31]. Samples were obtained from women that were all premenopausal with ages ranging from 34 to 48. Ten were African American and the race of 1 classified as “other.” Aliquots of 2 genital mucosal fluid samples (collected by lavage) with detectable glycogen were incubated at either 4°C or 37°C for 90 minutes, and a colorimetric iodine assay was then used to detect glycogen [31]. Sample A had more detected glycogen than sample B after incubation at 4°C (Figure 5A). For both of the samples, less glycogen was detected in the aliquot incubated at 37°C than in the portion incubated at 4°C, indicating temperature-dependent degradation of glycogen.
Figure 5.

Temperature-dependent degradation of glycogen in genital fluid. A, Aliquots of genital fluid collected from 2 subjects (A and B) were incubated at either 37°C or 4°C for 90 minutes and the level of endogenous glycogen was measured with a colorimetric iodine assay. B, Glycogen was added to genital fluids collected from 4 other women (C–F) or saline, and aliquots were incubated at either 4°C or 37°C for 90 minutes. At the end of the incubation, the amount of glycogen in the samples was measured with the colorimetric iodine assay. C, Degradation in 21 genital fluid samples was measured as in B and the percent degradation was calculated by the following formula: 1 – glycogen 37/glycogen 4 × 100.
Several other genital-fluid samples were tested (not shown) for glycogen breakdown in a similar manner. However, many of the samples had very low or no detectable glycogen and so glycogen degradation could not be assessed. Therefore, in further experiments, exogenous glycogen was added to genital fluids collected from women to ensure that degradation could be detected. Figure 5B shows the results from incubating genital fluid from 4 different individuals with exogenous glycogen at 4°C and 37°C. Less glycogen was detected for all 4 samples after incubation at 37°C compared to incubation at 4°C, with sample F exhibiting the most glycogen degradation and C having the least. Twenty-one samples from a total of 11 women were tested in this manner and the percent degradation was calculated using the following formula: 1 – glycogen 37/glycogen 4 × 100. The median degradation for the 21 samples was 73%, with only 3 samples having values less than 20% (Figure 5C).
To investigate whether the source of the glycogen-degrading activity could be bacterial, PCR was used to quantify several types of bacteria in the same samples. No significant relationships were observed between either L. crispatus, L. iners, or Gardnerella vaginalis and the glycogen degradation activity in women (P >.05, Spearman rank correlation, Figure 6A–C). Similarly, no relationships were observed between either L. jensenii, Mycoplasma hominis, or Sneathia (not shown). Further, the vaginal pH that was measured for diagnosing bacterial vaginosis was not significantly associated with α-amylase activity (P > .05, Figure 6D). These experiments therefore showed no obvious relationship between the types of microbiota in genital tract samples and the levels of the glycogen degradation activity.
Figure 6.

Relationship of glycogen degradation with genital bacteria, vaginal pH, α-amylase and α-glucosidase. The percent of glycogen degradation (Figure 5C) in 21 samples was plotted along with either genital-tract levels of L. iners (A), L. crispatus (B), G. vaginalis (C), vaginal pH (D), pancreatic α-amylase (E), or α-glucosidase (F).
To investigate a possible human source of the glycogen degradation activity, the genital fluid samples were tested for human α-amylase using an ELISA for pancreatic α-amylase. All tested samples had ELISA-detectable α-amylase, and the median α-amylase detected in genital fluids was 1.86 mU/mL (range, 0.02–12.7 mU/mL). The pancreatic α-amylase levels were strongly associated (Spearman r = 0.73; P = .0002) with the amount of glycogen degradation (Figure 6E). Acidic α-glucosidase was also detected in some of the samples (Figure 6F). However, its presence was not significantly associated with glycogen degradation (P > .05).
Glycogen Breakdown Products Generated by Genital Fluids
Because the above experiments suggested that glycogen produced by the genital epithelium could be degraded into smaller glucose polymers such as maltose and maltotriose in genital fluids, genital fluids from several women were analyzed by HPAEC for small carbohydrates. However, only trace amounts of small sugars were detected, and these levels were too low to determine the characteristics of these sugars. Therefore, in further experiments, exogenous glycogen was added to genital fluids and the types of carbohydrates that were generated were analyzed by HPAEC. The glycogen used for this experiment had only trace peaks corresponding to glucose, maltose, maltotriose, and maltotetraose (Figure 7A). After incubation for 48 hours in buffer with no added genital fluids, there was essentially no change in these peaks (Figure 7B). However, incubation for 48 hours with genital fluid from 2 different subjects resulted in substantial increases in peaks corresponding to maltose (G2), maltotriose (G3), and maltotetraose (G4), the major known α-amylase digestion products (Figure 7C and 7D). Trace peaks corresponding to glucose and maltopentaose were also observed. Similar results were observed after 24 hours of incubation, although the size of each of the peaks was reduced when compared with the 48-hour peaks (not shown). No peaks were detected in genital fluids with no added glycogen (not shown).
Figure 7.

Breakdown of glycogen by genital fluids. Glycogen was incubated for 0 hours (A) or 48 hours (B) with saline or 48 hours with genital fluid collected in saline from 2 different subjects (C and D). After incubation the size of small carbohydrates was determined by HPAEC. Abbreviation: HPAEC, high-performance anion-exchange chromatography.
DISCUSSION
Genital colonization with Lactobacillus has been associated with protection from STDs, BV, and adverse pregnancy outcomes [1, 5, 6, 8–21]. However, the factors that promote and maintain Lactobacillus colonization are not well understood. Glycogen is synthesized in the vaginal epithelium and has long been postulated to provide an energy source for genital Lactobacillus [25, 26]. However, it has not been clear how Lactobacillus uses glycogen because most genital isolates do not grow on glycogen in vitro [27, 28]. This study shows that (1) isolates of Lactobacillus that cannot use glycogen can grow in glycogen breakdown products, including maltose and maltotriose; (2) an activity that breaks down glycogen to maltose and maltotriose is present in the lower-genital-tract fluid of women; (3) human α-amylase is present in the genital fluids of women; (4) the glycogen breakdown activity in genital fluid correlates with genital levels of α-amylase; and (5) incubation of genital fluids with glycogen generated products such as maltose, maltotriose, and maltotetraose. This study therefore supports a model where α-amylase present in the genital tract breaks down glycogen into smaller polymers, such as maltose and maltotriose, that are then utilized by Lactobacillus to aid its colonization.
Glycogen is composed mainly of α1,4-linked chains of glucose but also has α1,6-linked branches. Human α-amylase (both salivary and pancreatic) preferentially processes glycogen to α1,4-linked multimers, such as maltose and maltotriose, but also results in production of α-dextrins that contain α1,6 branches and α1,4 linkages [36, 37]. This study shows that genital lactobacilli can grow well in the small unbranched glucose polymers. It is not clear, however, if lactobacilli can utilize branched carbohydrates.
There are some previous studies of utilization of maltose by Lactobacillus in the food and beverage industries, such as L. casei, and L. reuteri [38, 39]. However, those Lactobacillus species are different than the ones found in the genital tract, including L. crispatus, L. jensenii, L. iners, and L. gasseri [4, 20, 40, 41]. There is a paucity of studies on the utilization of small glucose polymers by genital tract lactobacilli. This study suggests that the small glucose polymers can support growth by genital Lactobacillus in vivo, and the consequent acid production suppresses growth of nonlactobacilli. The low genital pH found may vary somewhat depending on the species of Lactobacillus that is dominant [4]. The pH can be neutralized by factors such as sexual activity due to semen or by douching [42–44]. During periods of increased pH, other bacteria may utilize the glycogen breakdown products, and due to competition for this food source and production of pH buffering amines [45], prevent Lactobacillus from acidifying the genital environment.
Starch and glycogen have similar structures and there are reports of nongenital lactobacilli, such as L. fermentum and L. plantarum, degrading starch [46]. While there have been some reports of genital Lactobacillus isolates breaking down glycogen, this was later discovered in some cases to have been due to serum added to the culture medium that likely was a source of α-amylase [27, 47]. However, some genital Lactobacillus isolates can directly utilize glycogen [28].
We observed in this study that α-amylase and Lactobacillus levels were not correlated (Figure 6). However, we recently observed that there are variable levels of glycogen in the genital fluids of different women and that the levels of glycogen correlate strongly with the level of colonization by Lactobacillus (manuscript in preparation). Based on that observation, we postulate that women must have 2 conditions met to be dominantly colonized by Lactobacillus: (1) an adequate amount of glycogen in genital fluid; and (2) α-amylase to cleave the glycogen into smaller carbohydrates. If glycogen is low, then Lactobacillus levels will also be low. Potentially, if α-amylase levels were low but glycogen high, this could also lead to low Lactobacillus. Our studies do not rule out the possibility that other enzymes, including bacteria-derived enzymes, could process glycogen in some women. Also, because these experiments were conducted with women consisting mostly of 1 ethnic group, it is possible that women from other groups could have differing genital amylase activity.
This study has implications for the measurement of glycogen in genital secretions because α-amylase acts on glycogen, resulting in a mixture of glycogen and smaller glucose polymers. Glycogen is made in the epithelium, but it has not been clearly established how glycogen in epithelial cells becomes available for Lactobacillus utilization. Factors such as the amount of glycogen in cells, the rate of cell release, and the rate of glycogen breakdown are unknown. In our study, samples were collected on ice so that glycogen breakdown was arrested relatively quickly, but even so, many women had glycogen levels too low to measure with the iodine method. Further, glycogen breakdown products were too low to measure, possibly due to rapid uptake by bacteria.
While this study showed that human α-amylase was in genital fluid, the source for it was not established. As mentioned above, α-amylase is produced in both the pancreas and salivary glands, and essentially all of the extracellular α-amylase activity in the body is produced by those 2 glands [47, 48]. Both pancreatic and salivary α-amylase are in serum and urine at relatively high levels that can become elevated during certain clinical conditions such as alcoholism or trauma. The salivary and pancreatic enzymes are closely related, with only about 3% difference in the protein sequence [47]. Because the protein sequences are so similar, the ELISA for pancreatic α-amylase may have cross-reactivity with salivary α-amylase. Therefore, it is possible that the α-amylase in genital fluids is from 1 or both of these glands.
In conclusion, this study provides evidence that human α-amylase in the genital tract processes epithelial glycogen. This, in turn, suggests a mechanism for how glycogen can support genital colonization by strains of Lactobacillus that cannot directly use glycogen. Further studies are needed to clarify the source of the enzyme and whether too much or too little α-amylase could affect levels of Lactobacillus.
Notes
Financial support. This work was supported by National Institutes of Health (NIH) grants P01 AI08297 and P30 AI 082151.
Potential conflicts of interest. All authors: No reported conflicts.
All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
References
- 1.Lamont RF, Sobel JD, Akins RA, et al. The vaginal microbiome: new information about genital tract flora using molecular based techniques. BJOG. 2011;118:533–49. doi: 10.1111/j.1471-0528.2010.02840.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.O'Hanlon DE, Moench TR, Cone RA. Vaginal pH and microbicidal lactic acid when lactobacilli dominate the microbiota. PLOS ONE. 2013;8:e80074. doi: 10.1371/journal.pone.0080074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aldunate M, Tyssen D, Johnson A, et al. Vaginal concentrations of lactic acid potently inactivate HIV. J Antimicrob Chemother. 2013;68:2015–25. doi: 10.1093/jac/dkt156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ravel J, Gajer P, Abdo Z, et al. Microbes and Health Sackler Colloquium: vaginal microbiome of reproductive-age women. Proc Natl Acad Sci USA. 2010;108(Suppl 1):4680–7. doi: 10.1073/pnas.1002611107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Marrazzo JM. Interpreting the epidemiology and natural history of bacterial vaginosis: are we still confused? Anaerobe. 2011;17:186–90. doi: 10.1016/j.anaerobe.2011.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li J, McCormick J, Bocking A, et al. Importance of vaginal microbes in reproductive health. Reprod Sci. 2012;19:235–42. doi: 10.1177/1933719111418379. [DOI] [PubMed] [Google Scholar]
- 7.Macklaim JM, Cohen CR, Donders G, et al. Exploring a road map to counter misconceptions about the cervicovaginal microbiome and disease. Reprod Sci. 2012;19:1154–62. doi: 10.1177/1933719112446075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Atashili J, Poole C, Ndumbe PM, et al. Bacterial vaginosis and HIV acquisition: a meta-analysis of published studies. AIDS. 2008;22:1493–501. doi: 10.1097/QAD.0b013e3283021a37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Coleman JS, Hitti J, Bukusi EA, et al. Infectious correlates of HIV-1 shedding in the female upper and lower genital tracts. Aids. 2007;21:755–9. doi: 10.1097/QAD.0b013e328012b838. [DOI] [PubMed] [Google Scholar]
- 10.Sha BE, Zariffard MR, Wang QJ, et al. Female genital-tract HIV load correlates inversely with Lactobacillus species but positively with bacterial vaginosis and Mycoplasma hominis. J Infect Dis. 2005;191:25–32. doi: 10.1086/426394. [DOI] [PubMed] [Google Scholar]
- 11.Cu-Uvin S, Hogan JW, Caliendo AM, et al. Association between bacterial vaginosis and expression of human immunodeficiency virus type 1 RNA in the female genital tract. Clin Infect Dis. 2001;33:894–6. doi: 10.1086/322613. [DOI] [PubMed] [Google Scholar]
- 12.Mitchell C, Balkus JE, Fredricks D, et al. Interaction between lactobacilli, bacterial vaginosis–associated bacteria, and HIV type 1 RNA and DNA genital shedding in U.S. and Kenyan women. AIDS Res Hum Retroviruses. 2013;29:13–9. doi: 10.1089/aid.2012.0187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cohen CR, Lingappa JR, Baeten JM, et al. Bacterial vaginosis associated with increased risk of female-to-male HIV-1 transmission: a prospective cohort analysis among African couples. PLOS Med. 2012;9:e1001251. doi: 10.1371/journal.pmed.1001251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Watts DH. Mother to child transmission of HIV—another complication of bacterial vaginosis? J Acquir Immune Defic Syndr. 2012;60:221–4. doi: 10.1097/QAI.0b013e318256941c. [DOI] [PubMed] [Google Scholar]
- 15.Cherpes TL, Meyn LA, Krohn MA, et al. Association between acquisition of herpes simplex virus type 2 in women and bacterial vaginosis. Clin Infect Dis. 2003;37:319–25. doi: 10.1086/375819. [DOI] [PubMed] [Google Scholar]
- 16.Wiesenfeld HC, Hillier SL, Krohn MA, et al. Bacterial vaginosis is a strong predictor of Neisseria gonorrhoeae and Chlamydia trachomatis infection. Clin Infect Dis. 2003;36:663–8. doi: 10.1086/367658. [DOI] [PubMed] [Google Scholar]
- 17.Wiesenfeld HC, Hillier SL, Krohn MA, et al. Lower genital tract infection and endometritis: insight into subclinical pelvic inflammatory disease. Obstet Gynecol. 2002;100:456–63. doi: 10.1016/s0029-7844(02)02118-x. [DOI] [PubMed] [Google Scholar]
- 18.Hillier SL, Nugent RP, Eschenbach DA, et al. Association between bacterial vaginosis and preterm delivery of a low-birth-weight infant. The Vaginal Infections and Prematurity Study Group [see comments] N Engl J Med. 1995;333:1737–42. doi: 10.1056/NEJM199512283332604. [DOI] [PubMed] [Google Scholar]
- 19.Goldenberg RL, Klebanoff MA, Nugent R, et al. Bacterial colonization of the vagina during pregnancy in four ethnic groups. Vaginal Infections and Prematurity Study Group. Am J Obstet Gynecol. 1996;174:1618–21. doi: 10.1016/s0002-9378(96)70617-8. [DOI] [PubMed] [Google Scholar]
- 20.Antonio MA, Hawes SE, Hillier SL. The identification of vaginal Lactobacillus species and the demographic and microbiologic characteristics of women colonized by these species. J Infect Dis. 1999;180:1950–6. doi: 10.1086/315109. [DOI] [PubMed] [Google Scholar]
- 21.Eschenbach DA, Davick PR, Williams BL, et al. Prevalence of hydrogen peroxide–producing Lactobacillus species in normal women and women with bacterial vaginosis. J Clin Microbiol. 1989;27:251–6. doi: 10.1128/jcm.27.2.251-256.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Verstraelen H, Verhelst R, Claeys G, et al. Longitudinal analysis of the vaginal microflora in pregnancy suggests that L. crispatus promotes the stability of the normal vaginal microflora and that L. gasseri and/or L. iners are more conducive to the occurrence of abnormal vaginal microflora. BMC Microbiol. 2009;9:116. doi: 10.1186/1471-2180-9-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Larsson PG, Forsum U. Bacterial vaginosis—a disturbed bacterial flora and treatment enigma. APMIS. 2005;113:305–16. doi: 10.1111/j.1600-0463.2005.apm_113501.x. [DOI] [PubMed] [Google Scholar]
- 24.Mehta SD. Systematic review of randomized trials of treatment of male sexual partners for improved bacteria vaginosis outcomes in women. Sex Transm Dis. 2012;39:822–30. doi: 10.1097/OLQ.0b013e3182631d89. [DOI] [PubMed] [Google Scholar]
- 25.Rogosa M, Sharpe ME. Species differentiation of human vaginal lactobacilli. J Gen Microbiol. 1960;23:197–201. doi: 10.1099/00221287-23-1-197. [DOI] [PubMed] [Google Scholar]
- 26.Cruickshank R, Sharman A. The biology of the vagina in the human subject. II. The bacterial flora and secretion of the vagina at various age-periods and their relation to glycogen in the vagina epithelium. J Obstet Gynaec Brit Emp. 1934;41:208. [Google Scholar]
- 27.Stewart-Tull DE. Evidence that vaginal lactobacilli do not ferment glycogen. Am J Obstet Gynecol. 1964;88:676–9. doi: 10.1016/0002-9378(64)90898-1. [DOI] [PubMed] [Google Scholar]
- 28.Wylie JG, Henderson A. Identity and glycogen-fermenting ability of lactobacilli isolated from the vagina of pregnant women. J Med Microbiol. 1969;2:363–6. doi: 10.1099/00222615-2-3-363. [DOI] [PubMed] [Google Scholar]
- 29.Martin R, Soberon N, Vaneechoutte M, et al. Characterization of indigenous vaginal lactobacilli from healthy women as probiotic candidates. Int Microbiol. 2008;11:261–6. doi: 10.2436/20.1501.01.70. [DOI] [PubMed] [Google Scholar]
- 30.Amsel R, Totten PA, Spiegel CA, et al. Nonspecific vaginitis. Diagnostic criteria and microbial and epidemiologic associations. Am J Med. 1983;74:14–22. doi: 10.1016/0002-9343(83)91112-9. [DOI] [PubMed] [Google Scholar]
- 31.Morris DL. Colorimetric determination of glycogen; disadvantages of the iodine method. J Biol Chem. 1946;166:199–203. [PubMed] [Google Scholar]
- 32.Ao Z, Quezada-Calvillo R, Sim L, et al. Evidence of native starch degradation with human small intestinal maltase-glucoamylase (recombinant) FEBS Lett. 2007;581:2381–8. doi: 10.1016/j.febslet.2007.04.035. [DOI] [PubMed] [Google Scholar]
- 33.Lee CK, Le QT, Kim YH, et al. Enzymatic synthesis and properties of highly branched rice starch amylose and amylopectin cluster. J Agric Food Chem. 2008;56:126–31. doi: 10.1021/jf072508s. [DOI] [PubMed] [Google Scholar]
- 34.Fredricks DN, Fiedler TL, Thomas KK, et al. Changes in vaginal bacterial concentrations with intravaginal metronidazole therapy for bacterial vaginosis as assessed by quantitative PCR. J Clin Microbiol. 2009;47:721–6. doi: 10.1128/JCM.01384-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Srinivasan S, Liu C, Mitchell CM, et al. Temporal variability of human vaginal bacteria and relationship with bacterial vaginosis. PLOS ONE. 2010;5:e10197. doi: 10.1371/journal.pone.0010197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gray GM. Starch digestion and absorption in nonruminants. J Nutr. 1992;122:172–7. doi: 10.1093/jn/122.1.172. [DOI] [PubMed] [Google Scholar]
- 37.Larson SB, Day JS, McPherson A. X-ray crystallographic analyses of pig pancreatic alpha-amylase with limit dextrin, oligosaccharide, and alpha-cyclodextrin. Biochemistry. 2010;49:3101–15. doi: 10.1021/bi902183w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ganzle MG, Zhang C, Monang BS, et al. Novel metabolites from cereal-associated lactobacilli—novel functionalities for cereal products? Food Microbiol. 2009;26:712–9. doi: 10.1016/j.fm.2009.07.009. [DOI] [PubMed] [Google Scholar]
- 39.Meroth CB, Walter J, Hertel C, et al. Monitoring the bacterial population dynamics in sourdough fermentation processes by using PCR-denaturing gradient gel electrophoresis. Appl Environ Microbiol. 2003;69:475–82. doi: 10.1128/AEM.69.1.475-482.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Fredricks DN, Fiedler TL, Marrazzo JM. Molecular identification of bacteria associated with bacterial vaginosis. N Engl J Med. 2005;353:1899–911. doi: 10.1056/NEJMoa043802. [DOI] [PubMed] [Google Scholar]
- 41.Gajer P, Brotman RM, Bai G, et al. Temporal dynamics of the human vaginal microbiota. Sci Transl Med. 2012;4:132ra52. doi: 10.1126/scitranslmed.3003605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.O'Hanlon DE, Lanier BR, Moench TR, et al. Cervicovaginal fluid and semen block the microbicidal activity of hydrogen peroxide produced by vaginal lactobacilli. BMC Infect Dis. 2010;10:120. doi: 10.1186/1471-2334-10-120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Boskey ER, Telsch KM, Whaley KJ, et al. Acid production by vaginal flora in vitro is consistent with the rate and extent of vaginal acidification. Infect Immun. 1999;67:5170–5. doi: 10.1128/iai.67.10.5170-5175.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cherpes TL, Meyn LA, Krohn MA, et al. Risk factors for infection with herpes simplex virus type 2: role of smoking, douching, uncircumcised males, and vaginal flora. Sex Transm Dis. 2003;30:405–10. doi: 10.1097/00007435-200305000-00006. [DOI] [PubMed] [Google Scholar]
- 45.Hillier SL. Diagnostic microbiology of bacterial vaginosis. Am J Obstet Gynecol. 1993;169:455–9. doi: 10.1016/0002-9378(93)90340-o. [DOI] [PubMed] [Google Scholar]
- 46.Ganzle MG, Follador R. Metabolism of oligosaccharides and starch in lactobacilli: a review. Front Microbiol. 2012;3:340. doi: 10.3389/fmicb.2012.00340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Pieper-Bigelow C, Strocchi A, Levitt MD. Where does serum amylase come from and where does it go? Gastroenterol Clin North Am. 1990;19:793–810. [PubMed] [Google Scholar]
- 48.Vissers RJ, Abu-Laban RB, McHugh DF. Amylase and lipase in the emergency department evaluation of acute pancreatitis. J Emerg Med. 1999;17:1027–37. doi: 10.1016/s0736-4679(99)00136-5. [DOI] [PubMed] [Google Scholar]