

Abstract
Epithelial ovarian carcinoma (EOC), the major cause of gynaecological cancer death, is a heterogeneous disease classified into five subtypes. Each subtype has distinct clinical characteristics and is associated with different genetic risk factors and molecular events, but all are treated with surgery and platinum/taxane regimes. Tumour progression and chemoresistance is generally associated with major metabolic alterations, notably altered mitochondrial function(s). Here, we report for the first time that the expression of the mitochondrial regulators PGC1α and TFAM varies between EOC subtypes; furthermore, we have identified a profile in clear-cell carcinoma consisting of undetectability of PGC1α/TFAM, and low ERα/Ki-67. By contrast, high-grade serous carcinomas were characterised by a converse state of PGC1α/TFAM, ERα positivity and a high Ki-67 index. Interestingly, loss of PGC1α/TFAM and ERα was found also in a non-clear cell EOC cell line made highly resistant to platinum in vitro. Similar to clear-cell carcinomas, these resistant cells also showed accumulation of glycogen. Altogether, our data provide mechanistic insights into the chemoresistant nature of ovarian clear-cell carcinomas. Furthermore, these findings corroborate the need to take into account the diversity of EOC and to develop subtype specific treatment strategies.
Introduction
Epithelial ovarian carcinoma (EOC) is the major cause of gynaecological cancer death in women [1]. It is treated with surgery and platinum/taxane regimens as first-line therapy, but despite initial responsiveness, >75% of patients relapse into metastatic and essentially incurable chemoresistant disease [1], [2]. EOC is a heterogeneous disease that is broadly classified as serous (high- and low-grade serous carcinoma (HGSC and LGSC, respectively)), or non-serous (endometrioid, EC; mucinous, MC; clear-cell, CCC) histology. Each subtype has distinct clinical characteristics and is associated with different genetic risk factors and molecular events [3], [4]. Sensitivity to chemotherapy also varies between them, and survival correlates with subtype [5]. In CCC, both response rates and five-year survival are lower than in HGSC [5]–[7] and CCC has been generally accepted as an aggressive subtype that is poorly responsive to existing chemotherapy regimes.
To support the increased demand for energy and building blocks during tumour progression, cancer cells undergo major metabolic alterations [8], [9]. Chemoresistance and metastasis are associated with such alterations, notably altered mitochondrial function(s) [8]. Whereas normal cells depend on mitochondrial oxidative phosphorylation (OxPhos) for energy, tumour cell metabolism includes, among other pathways, decreased OxPhos and increased glycolysis (the Warburg effect) and glutaminolysis, both of which require mitochondria [9], [10]. Tumour cells also show survival-promoting alterations e.g. in regulation of autophagy in response to nutritional and other conditions of cellular/metabolic stress; these processes involve the AMP-activated protein kinase AMPK [11].
In normal cells, the transcriptional coactivator peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1α) is a major coordinator of metabolism and mitochondrial function [12] and together with SIRT1 and AMPK constitutes a sensitive network responding to metabolic stress [13]. Together with SIRT1, PGC1α has been reported to reside inside mitochondria with direct functions on mitochondrial biofunction [14]. PGC1α also regulates mitochondrial biogenesis through regulation of NRF1-2 [15], which in turn regulates mitochondrial transcription factor A (TFAM) [16]. TFAM plays a critical role in maintaining copy number and structure of mtDNA [17], [18], and is hence crucial for efficient transcription of mtDNA genes such as cytochrome c oxidase subunit 2 (MT-CO2) and other OxPhos proteins [17]. TFAM binding to mtDNA is regulated via phosphorylation [19]; unbound TFAM may be degraded by the Lon protease [19], [20]. Although well studied in normal cells, for instance regarding aging and neurodegeneration [21], [22], diabetes and obesity [23], it remains unknown what roles PGC1α/TFAM play in tumour progression.
Mitochondrial biogenesis has, in addition to PGC1α, in several studies been linked to expression and localisation of oestrogen (E2) and oestrogen receptor alpha (ERα) [24], [25]. The EOC subtypes express ERα at different levels, and expression is lowest in CCC [26].
The aim of this study was to examine the role of mitochondrial regulation in EOC by examining and correlating the expression of the mitochondrial and metabolic markers PGC1α, TFAM and ERα in clinical samples, and to relate the expression to EOC subtypes. Furthermore, we examined these markers in an EOC cell line and a multiresistant subline thereof.
Materials and Methods
Cells and cell culture
The non-CCC EOC cell line SKOV-3 and a multiresistant subline, SKOV-3-R (previously described in [27]) were cultured at +37°C, in 5% CO2 in RPMI-1640 medium containing 2 mM L-glutamine, 5% fetal bovine serum and 1% penicillin–streptomycin (all from Nordic Biolabs). SKOV-3 identity was validated using short tandem repeat analysis AmpFlSTR Identifiler kit (Applied Biosystems/Life Technologies) and cultures were routinely tested for mycoplasma contamination.
Quantitative real-time PCR
RNA was extracted using Qiagen RNeasy mini kit (Qiagen) and 500 ng/sample was used for the generation of cDNA using High Capacity cDNA Reverse Transcription Kit (Life Technologies) according to manufacturer's instructions. Quantitative real-time PCR (qRT-PCR) was performed on a 7900 HT Fast Real-Time system using TaqMan Gene expression assays (PPARC1A: Hs01016719_m1, TFAM: Hs01082775_m1, ESR1: Hs00174860_m1) and TaqMan Fast Universal PCR Master Mix (all from Applied Biosystems/Life Technologies) with the following cycling conditions: 95°C for 20 sec, 40 cycles of 95°C for 1 sec and 60°C for 20 sec. Expression of ACTB (Hs99999903_m1) was used as an endogenous control and the fold gene expression was calculated using the 2−ΔΔCT-method [28]. Raw data are shown in file Raw Data S1.
Western blot
Cells were lysed in RIPA lysis buffer (1% NP-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulphate, 0.004% sodium azide) containing protease and phosphatase inhibitor cocktails (all from Sigma-Aldrich). Samples were loaded for gel electrophoresis at 20 µg/sample and blotted onto PVDF membranes. Primary rabbit antibodies were to: PGC1α, β-tubulin (both from Abcam, 1∶1,000, 1∶5,000 respectively) and TFAM (1∶1,000, Sigma-Aldrich). Primary mouse antibodies were to ERα (1∶1,000, Santa Cruz Biotechnology) and Ki-67 (1∶500, Dako). Secondary HRP-conjugated antibodies used were donkey anti-rabbit IgG (1∶2,500, Abcam) and sheep anti-mouse IgG (1∶5,000, GE Healthcare). Images were developed with Western Lightning Plus-ECL (PerkinElmer) and captured using Protec Optimax X-Ray film processor.
Study population and ethical considerations
The study set (n = 53 cases) was drawn from cases originally diagnosed at the Karolinska University Hospital (Stockholm, Sweden). Participants provided written informed consent and the study was carried out with the approval of the Regional Ethics Committee of Stockholm (EPN 2013/318-31/2). Two pathologists reviewed all cases. The diagnoses included in the study set included 21 HGSC (40%), 14 CCC (26%), ten EC (19%), 7 MC (13%) and one LGSC (2%). Clinicopathological features of the study population can be found in Table 1. A tissue microarray (TMA) was constructed using paired 1 mm cores from representative tumour material. Representativity was confirmed using a hematoxylin and eosin control section.
Table 1. Study population.
| Clinical Variable | All | High-grade serous | Clear cell | Endometrioid | Mucinous | Low-grade serous | p-valuea |
| Number of cases, n (%) | 53 (100) | 21 (40) | 14 (26) | 10 (19) | 7 (13) | 1 (2) | |
| Age in years, median (range) | 64.0 (25–90) | 66.0 (36–83) | 63.5 (40–87) | 55.5 (43–88) | 64.0 (25–90) | n.d. | n.s. |
| Ki-67, median % (range) | 31.2 (1–90) | 38.8 (12–90) | 20.2 (1–46) | 19.3 (1–43) | 31.2 (12–49) | n.d. | 0.003 |
| ERα | <0.001 | ||||||
| Negative, n (%) | 23 (43) | 2 (10) | 12 (86) | 2 (20) | 7 (100) | 0 (0) | |
| Positive, n (%) | 30 (57) | 19 (90) | 2 (14) | 8 (80) | 0 (0) | 1 (100) | |
| PR | 0.038 | ||||||
| Negative, n (%) | 36 (68) | 14 (67) | 11 (79) | 4 (40) | 7 (100) | 0 (0) | |
| Positive, n (%) | 17 (32) | 7 (33) | 3 (21) | 6 (60) | 0 (0) | 1 (100) | |
| Histological Stage | 0.001 | ||||||
| I | 22 (42) | 2 (10) | 7 (50) | 8 (80) | 5 (71) | 0 (0) | |
| II | 5 (9) | 2 (10) | 2 (14) | 1 (10) | 0 (0) | 0 (0) | |
| III | 26 (49) | 17 (81) | 5 (36) | 1 (10) | 2 (29) | 1 (100) |
Abbreviations: ERα; oestrogen receptor alpha, nd; not determined, ns; not significant, PR; progesterone receptor.
Across variable; tested with Fisher's exact test for subtype, expression of ERα and PR, and Kruskal-Wallis test for distribution of age and Ki-67.
Immunohistochemistry and antibodies
Immunohistochemical staining (IHC) was performed on whole TMA using the EnVision + System (Dako). Antigen retrieval was achieved by immersing the sections in citrate buffer at pH 6.0 and boiling for 20 minutes in a water bath. The sections were then incubated with primary rabbit polyclonal antibodies against human voltage-dependent anion channel (VDAC) and PGC1α (both from Abcam, 1∶800 and 1∶100, respectively), TFAM and Lon (both from Sigma-Aldrich, 1∶10 and 1∶800, respectively) and rabbit monoclonal antibodies against human MT-CO2 (1∶100, Abcam) for 30 min at room temperature. A positive reaction was detected using the EnVision +System (Dako) and slides were counterstained with hematoxylin (Dako). Staining of Ki-67, ERα and progesterone receptor (PR) was performed at the routine laboratory at the Department of Pathology-Cytology, Karolinska University Hospital (Stockholm, Sweden), using rabbit monoclonal antibodies from Ventana Medical Systems.
Evaluation of immunohistochemistry
For evaluation of immunoreactivity and histological appearance, all TMA slides were examined independently by two investigators (MG and MS). Positive cytoplasmic expression of VDAC, ERα, PGC1α, TFAM, MT-CO2 and Lon was defined as immunoreactivity in >1% of epithelial cells, while immunoreactivity in <1% of epithelial cells defined negative protein expression. The scoring for each individual sample can be seen in the Raw Data S1. Ki-67 index was assessed as a continuous variable as percentage of positive tumour cells.
Glycogen assay
Cells were grown to confluency prior to harvesting. Intracellular levels of glycogen were then detected in 1×106 cells using the colorimetric Glycogen Assay Kit II (Abcam) according to manufacturer's instructions.
Sulforhodamine B assay
Growth rates in SKOV-3 and SKOV-3-R cells were evaluated based on total cellular protein in the samples, in turn assessed using the sulforhodamine B-based TOX6 kit (SRB assay, Sigma-Aldrich) according to manufacturer's instructions. Briefly, SKOV-3 and SKOV-3-R cells were seeded in 96-well plates at 4×103 cells/well in quadruplicates for each time point. After overnight incubation the wells for t = 0 h were fixed while the remaining wells were incubated additional 72 h before fixation. Resulting absorbances (i.e. total cellular protein) were compared with that at t = 0 h.
siRNA transfection
SKOV-3 cells seeded at 7.5×104 cells/well (12-well plates) or 15×104 cells/well (6-well plates) were transfected with 50 nM Silencer siRNA specific for PPARGC1A or with 50 nM Silencer Negative Control No. 1 siRNA (Life Technologies, id: 114747 and AM4611, respectively) after overnight incubation. siRNA, diluted in Opti-MEM (Life Technologies), was mixed by vortexing with 3 µl HiPerFect Transfection Reagent (Qiagen) per 100 µl Opti-MEM/siRNA mixture followed by 10 min incubation at RT. Subsequently, transfection complexes were added to the cells and incubated for 5 h under normal growth conditions, whereafter these complexes were replaced by antibiotic-free growth medium. At 72 h post-transfection, mRNA and protein levels were analysed by qRT-PCR and western blot, respectively. In parallel experiments, at 24 h post-transfected cells were treated with cisplatin (Hospira) for 48 h. Response to treatment was evaluated using the SRB assay.
Statistical analysis
To assess the relationship between protein expression levels and the variables in the clinical material, a non-parametric Chi2 test or Fisher's exact test was used as indicated in the tables. Comparisons between groups were assessed using the independent t-test (in vitro data), Mann-Whitney U test or Kruskal-Wallis test as indicated (IHC data). A two-tailed P value <0.05 was considered significant, and p values <0.05, <0.01 or <0.001 are represented with one, two or three asterisks, respectively, in the figures. IBM SPSS 20.0 statistical software (SPSS Inc.) was used for all analyses. For IHC analysis of MT-CO2 and Lon, two and five samples, respectively, were unavailable for scoring, which was taken into account in the statistical analysis.
Results
PGC1α and TFAM expression in multiresistant EOC cells
The EOC cell line SKOV-3 was subjected to repeated long-term treatment with cisplatin generating the multiresistant subline SKOV-3-R, as previously described in [27]. Since mitochondrial content was previously found to be higher in the resistant subline [27], expression of mitochondrial regulators PGC1α and TFAM was investigated here. mRNA expression of the genes coding for PGC1α and TFAM (PPARGC1A and TFAM, respectively) was significantly decreased in SKOV-3-R compared to parental SKOV-3 (p<0.001 for both) (Fig. 1A). Accordingly, protein expression of PGC1α and TFAM was reduced in SKOV-3-R cells (Fig. 1B).
Figure 1. PGC1α and TFAM expression in EOC SKOV-3 cells and the multiresistant subline SKOV-3-R.
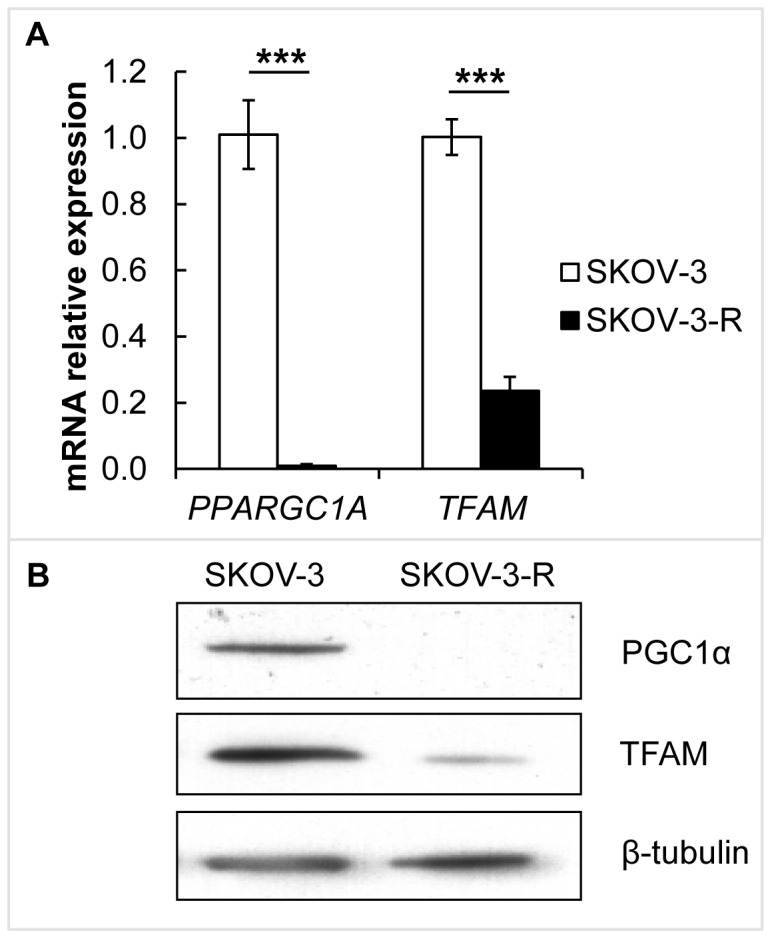
(A) Gene expression of PPARGC1A and TFAM in SKOV-3 and SKOV-3-R cells was evaluated by qRT-PCR (n = 3). The decrease in expression in SKOV-3-R was statistically significant (both p<0.001, using independent t-test). Expression levels were normalised to ACTB. Error bars represent S.E.M. (B) Representative western blot showing protein expression of PGC1α and TFAM. β-tubulin was used as loading control.
Clinicopathological features of the cohort
Immunohistochemistry was used to analyse the expression of selected proteins in 53 cases of EOC, including 21 HGSC (40%), 14 CCC (26%), ten EC (19%), seven MC (13%) and one LGSC (2%). Patient and tumour characteristics are described in Table 1. Representative images of IHC staining for Ki-67, MT-CO2 and Lon are shown in Fig. S1.
No significant difference in median age was observed between the subtypes. Significant differences between subtypes were found for the biomarkers Ki-67 (p = 0.003) Figs. S1–S2), ERα (p<0.001), PR, (p = 0.038) as well as for the distribution across histological stages (p = 0.001) (Table 1). Within HGSC, 19 tumours (90%) were positive and 2 (10%) were negative for ERα (p<0.001), whereas in CCC 2 tumours (14%) were positive and 12 (86%) were negative (p = 0.008).
The mitochondrial outer membrane VDAC was used as a positive control for presence of mitochondria. Expression of VDAC was found in all samples and was equal across all EOC subtypes (not shown).
Expression of PGC1α and TFAM in EOC tumours
Protein expression of PGC1α was cytoplasmic and seen in 42 tumours (79%); conversely, eleven tumours (21%) lacked PGC1α. The expression varied significantly across all EOC subtypes (p = 0.002). In HGSC, 18 tumours (86%) were positive and three (14%) were negative for PGC1α (p<0.001). In CCC, six tumours (43%) were positive whereas eight (57%) were negative. All EC, MC and LGSC tumours were positive for PGC1α. In the total cohort, there was no significant correlation between PGC1α, Ki-67 index, PR, age or histological stage. By contrast, there was a significant positive correlation between ERα and PGC1α (p<0.001) (Table 2).
Table 2. Expression status of PGC1α and TFAM in the EOC study population.
| PGC1α | TFAM | |||||||
| Clinical Variable | Negative | Positive | p-valuea | p-valueb | Negative | Positive | p-valuea | p-valueb |
| Number of cases, n (%) | 11 (21) | 42 (79) | 18 (34) | 35 (66) | ||||
| Subtype | 0.002 | 0.001 | ||||||
| High-grade serous, n (%) | 3 (14) | 18 (86) | 0.001 | 3 (14) | 18 (86) | 0.001 | ||
| Clear cell, n (%) | 8 (57) | 6 (43) | n.s. | 11 (79) | 3 (21) | 0.033 | ||
| Endometrioid, n (%) | 0 (0) | 10 (100) | n.d. | 3 (30) | 7 (70) | n.s. | ||
| Mucinous, n (%) | 0 (0) | 7 (100) | n.d. | 1 (34) | 6 (66) | n.s. | ||
| Low-grade serous, n (%) | 0 (0) | 1 (100) | n.d. | 0 (0) | 1 (100) | n.d. | ||
| Age in years, median (range) | 61.0 (40–87) | 64.5 (25–90) | n.s. | 65.5 (40–87) | 63.0 (25–90) | ns | ||
| Ki-67, median % (range) | 32.5 (1–52) | 30.0 (1–90) | n.s. | 20.2 (1–68) | 33.2 (1–90) | 0.014 | ||
| ERα | <0.001 | 0.014 | ||||||
| Negative, n (%) | 10 (44) | 13 (57) | 12 (52) | 11 (48) | ||||
| Positive, n (%) | 1 (3) | 29 (97) | 6 (20) | 24 (80) | ||||
| PR | n.s. | n.s. | ||||||
| Negative, n (%) | 10 (28) | 26 (72) | 15 (41) | 21 (58) | ||||
| Positive, n (%) | 1 (6) | 16 (94) | 3 (18) | 14 (82) | ||||
Abbreviations: EOC; epithelial ovarian carcinoma, ERα; oestrogen receptor alpha, nd; not determined, ns; not significant, PR; progesterone receptor, PGC1α; Peroxisome proliferator-activated receptor gamma co-activator 1-alpha, TFAM; mitochondrial transcription factor A.
Across variable; tested with Fisher's exact test for subtypes, expression of ERα and PR, and Mann-Whitney U test for distribution of age and Ki-67.
Within variable; tested with non-parametric Chi2 test.
Expression of TFAM was cytoplasmic, and varied significantly across all EOC subtypes (p = 0.001). In total, TFAM was expressed in 35 tumours (66%) and undetectable in 18 tumours (34%). In HGSC, 18 tumours were positive and three were negative for TFAM (86% and 14%, respectively) (p = 0.001). In CCC, three tumours were positive whereas eleven were negative (21% and 79%, respectively) (p = 0.033). In EC, seven tumours were positive and three were negative (70% and 30%, respectively). In MC, six tumours were positive and one was negative (66% and 34%, respectively), and in LGSC, the only tumour included expressed TFAM (100%).
Across the EOC subtypes, there was no significant correlation between TFAM, PR, age or histological stage. However, tumours expressing TFAM displayed a significantly higher Ki-67 index compared to TFAM negative tumours (median score 33% and 20%, respectively) (p = 0.014) (Fig. 2). There was also a significant positive correlation between TFAM and expression of ERα (p = 0.014). In order to investigate if lack of TFAM expression affected the expression of mtDNA-encoded genes, protein expression of MT-CO2 was assessed. Of 51 tumours stained for MT-CO2, 47 (92%) were positive and four (8%) were negative. Three of the four MT-CO2 negative tumours were also TFAM negative; however, there was no significant relation between expression of MT-CO2 and TFAM (Table S1 in File S1).
Figure 2. Ki-67 index distribution in TFAM+ and TFAM- EOC tumours.

EOC tumours with negative expression of TFAM (n = 18) showed significantly lower Ki-67 index compared to tumours with positive expression of TFAM (n = 35) (Mann-Whitney U, p = 0.014).
TFAM protein levels are in part regulated through degradation by the Lon protease [19], [20]. To investigate if increased levels of Lon might explain the reduced levels of TFAM found in CCC, expression of Lon was assessed. Lon was localised to the cytoplasm, and was found in 38 tumours (79%) which were differently distributed across all EOC subtypes (p = 0.032). There was no significant correlation between expression of TFAM and Lon (Table S2 in File S1).
Co-expression of PGC1α and TFAM in EOC tumours
As PGC1α is a major coordinator of mitochondrial functions including regulation of transcription of TFAM [12], [15], [16], we investigated the co-expression of these two proteins. The cohort was categorised into four different groups depending on their expression of PGC1α and TFAM (PGC1α-/TFAM-; PGC1α-/TFAM+; PGC1α+/TFAM- and PGC1α+/TFAM+). The expression of PGC1α/TFAM varied significantly across the EOC subtypes (p = 0.001) (Table 3, Fig. 3). There was also a significant difference in median Ki-67 index between the four groups, where positive expression of TFAM associated with increased Ki-67 index independently of PGC1α expression (p = 0.048) (Table 3, and Fig. S3). Furthermore, there was a positive association between ERα expression and co-expression of PGC1α/TFAM (p = 0.002) (Table 3), whereas no significant association with PR was observed.
Table 3. Expression status of PGC1α/TFAM in EOC.
| Clinical Variable | PGC1α-/TFAM- | PGC1α-/TFAM+ | PGC1α/TFAM- | PGC1α/TFAM+ | p-valuea |
| Number of cases, n (%) | 9 (17) | 2 (4) | 9 (17) | 33 (62) | |
| Subtype | 0.001 | ||||
| High-grade serous, n (%) | 1 (5) | 2 (10) | 2 (10) | 16 (76) | |
| Clear cell, n (%) | 8 (57) | 0 (0) | 3 (21) | 3 (21) | |
| Endometrioid, n (%) | 0 (0) | 0 (0) | 3 (30) | 7 (70) | |
| Mucinous, n (%) | 0 (0) | 0 (0) | 1 (14) | 6 (86) | |
| Low-grade serous, n (%) | 0 (0) | 0 (0) | 0 (0) | 1 (100) | |
| Age in years, median (range) | 61.0 (40–87) | 55.0 (44–66) | 70.0 (53–80) | 63.0 (25–90) | n.s |
| Ki-67, median % (range) | 24.7 (1–43) | 49.5 (48–52) | 17.9 (5–68) | 32.8 (1–90) | 0.048 |
| ERα | 0.002 | ||||
| Negative, n (%) | 8 (35) | 2 (9) | 4 (17) | 9 (39) | |
| Positive, n (%) | 1 (3) | 0 (0) | 5 (17) | 24 (80) | |
| PR | n.s. | ||||
| Negative, n (%) | 8 (22) | 2 (6) | 7 (19) | 19 (53) | |
| Positive, n (%) | 1 (6) | 0 (0) | 2 (12) | 14 (82) |
Abbreviations: EOC; epithelial ovarian carcinoma, ERα; oestrogen receptor alpha, nd; not determined, ns; not significant, PR; progesterone receptor, PGC1α; Peroxisome proliferator-activated receptor gamma co-activator 1-alpha, TFAM; mitochondrial transcription factor A.
Across variable; tested with Fisher's exact test for subtype, expression of ERα and PR, and Kruskal-Wallis test for distribution of age and Ki-67.
Figure 3. Expression of PGC1α and TFAM in EOC subtypes.

(A) Representative immunohistochemical staining of VDAC, ERα, PGC1α and TFAM in HGSC and CCC. VDAC indicates presence of mitochondria. ERα, PGC1α and TFAM were all expressed in HGSC, whereas in CCC expression of the proteins was lost. Each HGSC and CCC quadruplicate is from one and the same patient (magnification: 400x, scale bar shows 500 µm). (B) Histogram representing number of cases with expression of PGC1α/TFAM across EOC subtypes.
Co-expression of PGC1α/TFAM was found in 33 samples (62% of the total cohort) of which 16 (48%) were HGSC. Within the HGSC subtype, 76% of the tumours were double-positive for PGC1α/TFAM (p<0.001). Lack of both PGCα and TFAM was observed in nine tumours (17% of total cohort). Of these nine, all but one were CCC (89% of all double negative) (p = 0.020). Within the CCC subtype, double-negative expression occurred in 57% of the tumours and all of these were ERα negative (Table 3).
CCC profile of multiresistant EOC cell line
SKOV-3 cells have been reported to have a 32-basepair deletion in exon 1 of ERα transcript and are thus ERα positive but oestrogen-insensitive [29], [30]. To further investigate the link between regulation of ERα expression, as such, and PGC1α/TFAM, levels of ERα were analysed on mRNA and protein levels. Compared to parental SKOV-3, the multiresistant SKOV-3-R cells show significantly decreased mRNA expression of the ESR1 gene (p<0.001) (Fig. 4A), and accordingly decreased ERα protein expression (Fig. 4B). Raw data can be found under S_PCR in the file Raw data S1s.
Figure 4. ERα expression and intracellular levels of glycogen in SKOV-3 and SKOV-3-R.
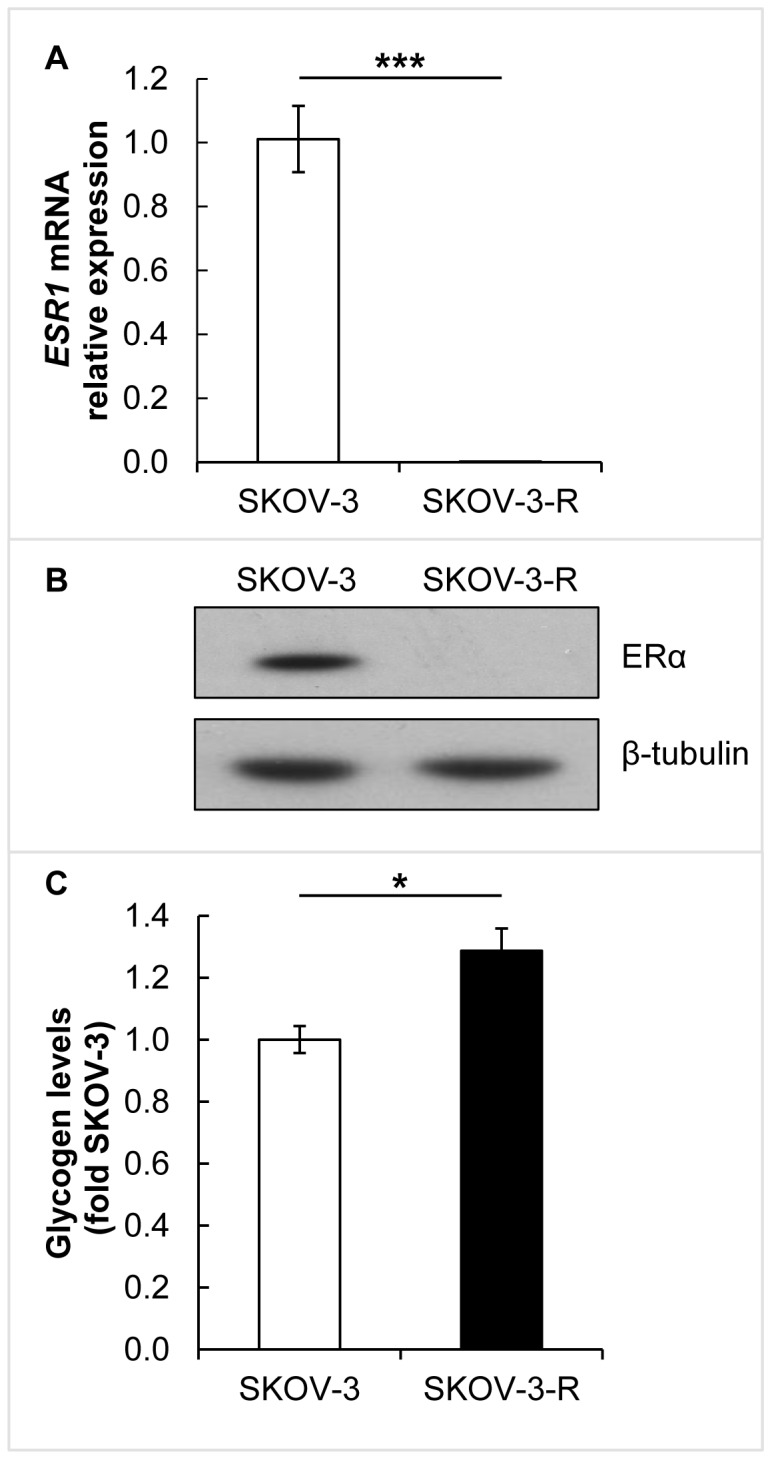
(A) Gene expression of ESR1 evaluated by qRT-PCR (n = 3). The decrease in expression in SKOV-3-R was statistically significant (p<0.001, using independent t-test). Expression levels were normalised to ACTB. Error bars represent S.E.M. (B) Representative western blot showing protein expression of ERα. β-tubulin was used as a loading control. (C) Glycogen levels were evaluated using a colorimetric assay (n = 3). The increased level in SKOV-3-R was statistically significant (p = 0.027, using independent t-test). Error bars represent S.E.M.
Accumulation of glycogen is characteristic of CCC [31], wherefore we investigated intracellular glycogen levels in SKOV-3 and SKOV-3-R cells. The SKOV-3-R were found to accumulate 25% more intracellular glycogen (p = 0.027) compared to parental cells (Fig. 4C).
Discussion
We have here shown that the expression of mitochondrial regulators PGC1α and TFAM varies significantly between HGSC and CCC. Furthermore, we identified a profile in CCC consisting of undetectability of PGC1α/TFAM, and low ERα/Ki-67. By contrast, HGSC samples were characterised by a converse state of PGC1α/TFAM and ERα positivity as well as a high Ki-67 index (summarized in Fig. 5). Interestingly, the CCC profile had developed also in an EOC cell line that is not of CCC origin and which had been made highly chemoresistant in vitro.
Figure 5. Comparison of ovarian HGSC and CCC.

Representative hematoxylin and eosin staining of ovarian HGSC and CCC. We hypothesise that these subtypes represents extreme forms of EOC; HGSC generally display positive expression of PGC1α, TFAM and ERα, have a higher Ki-67 index and are more responsive to initial platinum/taxane chemotherapy. By contrast, CCC generally do not express PGC1α, TFAM and ERα, have a lower Ki-67 index and are less responsive to initial platinum/taxane chemotherapy. Magnification 400x, scale bar shows 50 µm.
Here, out of the nine tumour samples that were double-negative for both PGC1α/TFAM, eight were CCC. In view of its characteristic histology, accumulation of intracellular glycogen and its highly chemoresistant nature, CCC represents a distinct clinicopathological subtype of requires new treatment strategies [32]. We believe that our findings support this notion; furthermore, our data also provide mechanistic insights into where this chemoresistance comes from.
Very little is reported on the role of PGC1α in cancer in general, and even less in chemoresistance. Low levels of PGC1α have been reported in ten EOC samples compared with normal ovaries, and PGC1α overexpression in vitro led to upregulation of Bax, downregulation of Bcl-2 and to apoptosis [33]. PGC1α was reported to be low also in 15 out of 17 colon cancer biopsies compared to normal [34]. There were, however, no reported results on any association with data on treatment effects or chemoresistance. Nevertheless, the role of the PGC1α/oestrogen-related receptors (ERR) axis is known to be instrumental in regulation of cancer cell metabolism (reviewed in [35]), which in turn is known to affect responses to chemotherapy [8], [36].
Low expression of PGC1α might be expected to correlate with lower mitochondrial content. Mitochondria are required for the full effect of some chemotherapeutic drugs, e.g. cisplatin, due to their role in apoptosis and a requirement for mitochondrially produced ROS [37]. However, we did not observe any correlation between PGC1α expression and mitochondrial content, based on VDAC and MT-CO2 expression. PGC1α is involved also in regulation of antioxidant defence [21], [38], and the observed low levels of PGC1α would thus be expected to not reduce ROS production and thereby the effect of platinum. Therefore, the role of PGC1α in EOC, particularly in notoriously resistant CCC [32], is likely not linked to antioxidant defence.
TFAM is a requirement for normal replication and transcription of mtDNA [17], [18]; this is evidenced as reduced mtDNA content in colon carcinoma cell lines as well as in biopsies carrying truncating mutations in TFAM [39]. Here, of the four samples that did not express mitochondrially encoded MT-CO2, three did not show detectable TFAM. This suggests the possibility that a detectable proportion of EOC tumours do not have functional respiration, as MT-CO2 provides the substrate-binding site of Complex IV in the respiratory chain [40]. Based on the literature [18], [41], we hypothesised that minute amounts of TFAM are sufficient for transcriptional activity and that the undetectability of TFAM protein might be due to Lon-mediated degradation [19], [20]. However, as there was no significant correlation between the expression of TFAM and Lon, this was not the case.
CCC generally shows low Ki-67 index [32]. A correlation between TFAM and Ki-67 has, to our knowledge, not been studied. However, in analogy with the report that in highly proliferative transformed lymphoblastoid cells, increased expression of TFAM was suggested to be associated with protection from oxidative stress induced damage [42], we speculate that highly proliferative HGSC tumours, as indicated by Ki-67, upregulate TFAM as part of an antioxidant defence. Our in vitro results are also in accordance with this, since SKOV-3-R cells showed downregulation of TFAM and grow more slowly than the parental cells (Fig. S4). Moreover, TFAM expression correlated positively with ERα, which might reflect a potential for growth as well as for an efficient oxidative phenotype. In line with previous studies [26], [43], expression of ERα varied across the EOC subtypes, with low levels in CCC.
In a mouse model, SKOV-3 tumour xenografts (non-CCC, EOC cells) treated with trastuzumab were at post-treatment found to be heterogeneous with large areas of ERα negative CCC morphology [44]. This finding is interesting in the present context, as the multiresistant SKOV-3-R cells had acquired the CCC profile, compared to parental SKOV-3 cells. Furthermore, SKOV-3-R showed accumulation of glycogen, again in accordance with a CCC phenotype. Knocking down PPARGC1A mRNA using siRNA in the SKOV-3 cells down-regulated PGC1α mRNA as well as protein levels seen at 72 h post transfection (Figs. S5A–B). TFAM protein was also reduced, although TFAM mRNA expression was restored at this time point (Figs. S5A–B). Although not significant, the knockdown cells showed a clear tendency of reduced response to cisplatin treatment compared to the negative control cells (Fig. S5C). This tendency was not due to reduced growth, as knocking down PPARGC1A did not alter the growth rate or the expression of Ki-67 positive cells. We speculate that long-term downregulation of PGC1α and/or TFAM is necessary to decrease growth rate over time.
Although our SKOV-3 cell line model represents only itself, we suggest that downregulation of PGC1α activity as part of a chemoresistant phenotype profile might not be restricted to ovarian CCC. Low PGC1α activity would be expected in tumour contexts where loss of LKB1/AMPK represents loss of tumour suppressor function [45]. In keeping with a context-dependent tumour suppressor and oncogenic role for AMPK [45], the comparatively high levels of PGC1α expression that we observed in the highly proliferative, i.e., with high Ki-67 expression, HGSC tumours could then be suggested to belong to a context of AMPK-mediated protection from proliferation-related oxidative stress.
In summary, we propose that the profile consisting of PGC1α/TFAM downregulation and decreased ERα/Ki-67, and likely also glycogen accumulation, is representative of CCC and chemoresistance. We are currently investigating an underlying mechanistic explanation for this association. Fig. 5 summarises how HGSC and CCC may represent opposites with regard to the profile. Altogether, the findings corroborate the need to take into account the diversity in EOC and to develop specific treatment strategies for each subtype.
Supporting Information
Expression of Ki-67, MT-CO2 and Lon in EOC. Representative negative and positive immunohistochemical staining of Ki-67, MT-CO2 and Lon in EOC (magnification: 400x, scale bar shows 500 µm).
(TIF)
Ki-67 index distribution in different EOC subtypes. Distribution of Ki-67 index varied significantly across the different EOC subtypes (Mann-Whitney U, p = 0.003). High-grade serous carcinoma (HGSC) (n = 21), clear cell (CC) (n = 14), endometrial carcinoma (EC) (n = 10), mucinous carcinoma (MC) (n = 7) and low-grade serous carcinoma (LGSC) (n = 1).
(TIF)
Ki-67 index distribution in EOC tumours depending on expression of PGC1α and TFAM. Distribution of Ki-67 index varied significantly across the different groups of tumours with PGC1α-/TFAM- (n = 9), PGC1α-/TFAM+ (n = 2), PGC1α+/TFAM- (n = 9) and PGC1α+/TFAM+ (n = 33) (Kruskal-Wallis test, p = 0.048).
(TIF)
Growth rates in EOC SKOV-3 cells and the multiresistant subline SKOV-3-R. Growth rates in SKOV-3 and SKOV-3-R cells assessed as cellular protein at given time points using the sulphorhodamine B assay (n = 4). Data are expressed as fold increase from t = 0 h. S.E.M. error bars were too small to be visualized, except where shown.
(TIF)
PGC1α/TFAM expression and response to cisplatin treatment in SKOV-3 PPARGC1A siRNA knockdown cells. SKOV-3 cells were treated with PPARGC1A siRNA knockdown or siRNA negative control; (A) gene expression of PPARGC1A and TFAM was evaluated at 72 h post-transfection by qRT-PCR (n = 3). Expression levels were normalised to ACTB. Error bars represent S.E.M. (B) representative western blot showing protein expression at 72 h post-transfection of PGC1α, Ki-67 and TFAM. β-tubulin was used as loading control. (C) At 24 h post-transfection, SKOV-3 PPARGC1A siRNA knockdown and siRNA negative control cells were treated with indicated doses of cisplatin for 48 h, whereafter cellular protein was measured using the sulphorhodamine B assay (n = 3). Data are expressed as percent of untreated cells. Error bars represent S.E.M.
(TIF)
Supporting tables. Table S1, Expression of MT-CO2 in EOC. Shown is the distribution of positive and negative expression of MT-CO2 in the total sample, in the five EOC subgroups and in TFAM-expressing and non-expressing samples, respectively. Table S2, Expression of Lon in EOC. Shown is the distribution of positive and negative expression of Lon in the total sample, in the five EOC subgroups and in TFAM-expressing and non-expressing samples, respectively.
(DOCX)
S_PCR: Gene expression raw data given as Ct, delta-Ct (DCt) and delta-delta-Ct (DDCt) values, respectively, for results shown in Figs. 1 and 4, and Fig. S5. S_Spect: Spectrophotometry raw data given as absorbances and calculations based thereupon, for results shown in Figs. 4 and S5. S_IHC: The finalized scoring of immunohistochemical stainings.
(ZIP)
Acknowledgments
We thank Inger Bodin, Cancer Center Karolinska, Karolinska Institutet, and Elin Edsbäcker, for technical assistance and fruitful discussions.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by Swedish Cancer Society, contracts 100476 and 120814; research Council of Sweden, contract A0360901; and Radiumhemmet Research Foundation, contract 124182. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Lengyel E (2010) Ovarian cancer development and metastasis. The American journal of pathology 177: 1053–1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Coleman RL, Monk BJ, Sood AK, Herzog TJ (2013) Latest research and treatment of advanced-stage epithelial ovarian cancer. Nat Rev Clin Oncol 10: 211–224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Shih Ie M, Kurman RJ (2004) Ovarian tumorigenesis: a proposed model based on morphological and molecular genetic analysis. Am J Pathol 164: 1511–1518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Gurung A, Hung T, Morin J, Gilks CB (2013) Molecular abnormalities in ovarian carcinoma: clinical, morphological and therapeutic correlates. Histopathology 62: 59–70. [DOI] [PubMed] [Google Scholar]
- 5. Winter WE 3rd, Maxwell GL, Tian C, Carlson JW, Ozols RF, et al. (2007) Prognostic factors for stage III epithelial ovarian cancer: a Gynecologic Oncology Group Study. J Clin Oncol 25: 3621–3627. [DOI] [PubMed] [Google Scholar]
- 6. du Bois A, Luck HJ, Meier W, Adams HP, Mobus V, et al. (2003) A randomized clinical trial of cisplatin/paclitaxel versus carboplatin/paclitaxel as first-line treatment of ovarian cancer. J Natl Cancer Inst 95: 1320–1329. [DOI] [PubMed] [Google Scholar]
- 7. Takano M, Kikuchi Y, Yaegashi N, Kuzuya K, Ueki M, et al. (2006) Clear cell carcinoma of the ovary: a retrospective multicentre experience of 254 patients with complete surgical staging. Br J Cancer 94: 1369–1374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144: 646–674. [DOI] [PubMed] [Google Scholar]
- 9. Ward PS, Thompson CB (2012) Metabolic reprogramming: a cancer hallmark even warburg did not anticipate. Cancer cell 21: 297–308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. DeBerardinis RJ, Mancuso A, Daikhin E, Nissim I, Yudkoff M, et al. (2007) Beyond aerobic glycolysis: transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. Proc Natl Acad Sci U S A 104: 19345–19350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Hardie DG (2011) AMP-activated protein kinase: an energy sensor that regulates all aspects of cell function. Genes Dev 25: 1895–1908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Finck BN, Kelly DP (2006) PGC-1 coactivators: inducible regulators of energy metabolism in health and disease. J Clin Invest 116: 615–622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Canto C, Auwerx J (2009) PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Current opinion in lipidology 20: 98–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Aquilano K, Vigilanza P, Baldelli S, Pagliei B, Rotilio G, et al. (2010) Peroxisome proliferator-activated receptor gamma co-activator 1alpha (PGC-1alpha) and sirtuin 1 (SIRT1) reside in mitochondria: possible direct function in mitochondrial biogenesis. J Biol Chem 285: 21590–21599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Ventura-Clapier R, Garnier A, Veksler V (2008) Transcriptional control of mitochondrial biogenesis: the central role of PGC-1alpha. Cardiovasc Res 79: 208–217. [DOI] [PubMed] [Google Scholar]
- 16. Virbasius JV, Scarpulla RC (1994) Activation of the human mitochondrial transcription factor A gene by nuclear respiratory factors: a potential regulatory link between nuclear and mitochondrial gene expression in organelle biogenesis. Proc Natl Acad Sci U S A 91: 1309–1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Kaufman BA, Durisic N, Mativetsky JM, Costantino S, Hancock MA, et al. (2007) The mitochondrial transcription factor TFAM coordinates the assembly of multiple DNA molecules into nucleoid-like structures. Mol Biol Cell 18: 3225–3236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Kanki T, Ohgaki K, Gaspari M, Gustafsson CM, Fukuoh A, et al. (2004) Architectural role of mitochondrial transcription factor A in maintenance of human mitochondrial DNA. Mol Cell Biol 24: 9823–9834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Lu B, Lee J, Nie X, Li M, Morozov YI, et al. (2013) Phosphorylation of human TFAM in mitochondria impairs DNA binding and promotes degradation by the AAA+ Lon protease. Mol Cell 49: 121–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Matsushima Y, Goto Y, Kaguni LS (2010) Mitochondrial Lon protease regulates mitochondrial DNA copy number and transcription by selective degradation of mitochondrial transcription factor A (TFAM). Proc Natl Acad Sci U S A 107: 18410–18415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Austin S, St-Pierre J (2012) PGC1alpha and mitochondrial metabolism–emerging concepts and relevance in ageing and neurodegenerative disorders. J Cell Sci 125: 4963–4971. [DOI] [PubMed] [Google Scholar]
- 22. Lettieri Barbato D, Baldelli S, Pagliei B, Aquilano K, Ciriolo MR (2012) Caloric Restriction and the Nutrient-Sensing PGC-1alpha in Mitochondrial Homeostasis: New Perspectives in Neurodegeneration. Int J Cell Biol 2012: 759583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Handschin C, Spiegelman BM (2006) Peroxisome proliferator-activated receptor gamma coactivator 1 coactivators, energy homeostasis, and metabolism. Endocr Rev 27: 728–735. [DOI] [PubMed] [Google Scholar]
- 24. Klinge CM (2008) Estrogenic control of mitochondrial function and biogenesis. J Cell Biochem 105: 1342–1351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Tcherepanova I, Puigserver P, Norris JD, Spiegelman BM, McDonnell DP (2000) Modulation of estrogen receptor-alpha transcriptional activity by the coactivator PGC-1. J Biol Chem 275: 16302–16308. [DOI] [PubMed] [Google Scholar]
- 26. Kobel M, Kalloger SE, Boyd N, McKinney S, Mehl E, et al. (2008) Ovarian carcinoma subtypes are different diseases: implications for biomarker studies. PLoS Med 5: e232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Wintzell M, Lofstedt L, Johansson J, Pedersen AB, Fuxe J, et al. (2012) Repeated cisplatin treatment can lead to a multiresistant tumor cell population with stem cell features and sensitivity to 3-bromopyruvate. Cancer Biol Ther 13: 1454–1462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402–408. [DOI] [PubMed] [Google Scholar]
- 29. Lau KM, Mok SC, Ho SM (1999) Expression of human estrogen receptor-alpha and -beta, progesterone receptor, and androgen receptor mRNA in normal and malignant ovarian epithelial cells. Proc Natl Acad Sci U S A 96: 5722–5727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Hua W, Christianson T, Rougeot C, Rochefort H, Clinton GM (1995) SKOV3 ovarian carcinoma cells have functional estrogen receptor but are growth-resistant to estrogen and antiestrogens. J Steroid Biochem Mol Biol 55: 279–289. [DOI] [PubMed] [Google Scholar]
- 31. Ohkawa K, Amasaki H, Terashima Y, Aizawa S, Ishikawa E (1977) Clear cell carcinoma of the ovary: light and electron microscopic studies. Cancer 40: 3019–3029. [DOI] [PubMed] [Google Scholar]
- 32. Anglesio MS, Carey MS, Kobel M, Mackay H, Huntsman DG, et al. (2011) Clear cell carcinoma of the ovary: a report from the first Ovarian Clear Cell Symposium, June 24th, 2010. Gynecol Oncol 121: 407–415. [DOI] [PubMed] [Google Scholar]
- 33. Zhang Y, Ba Y, Liu C, Sun G, Ding L, et al. (2007) PGC-1alpha induces apoptosis in human epithelial ovarian cancer cells through a PPARgamma-dependent pathway. Cell Res 17: 363–373. [DOI] [PubMed] [Google Scholar]
- 34. Feilchenfeldt J, Brundler MA, Soravia C, Totsch M, Meier CA (2004) Peroxisome proliferator-activated receptors (PPARs) and associated transcription factors in colon cancer: reduced expression of PPARgamma-coactivator 1 (PGC-1). Cancer Lett 203: 25–33. [DOI] [PubMed] [Google Scholar]
- 35. Deblois G, St-Pierre J, Giguere V (2013) The PGC-1/ERR signaling axis in cancer. Oncogene 32: 3483–3490. [DOI] [PubMed] [Google Scholar]
- 36. Zhao Y, Butler EB, Tan M (2013) Targeting cellular metabolism to improve cancer therapeutics. Cell Death Dis 4: e532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Berndtsson M, Hagg M, Panaretakis T, Havelka AM, Shoshan MC, et al. (2007) Acute apoptosis by cisplatin requires induction of reactive oxygen species but is not associated with damage to nuclear DNA. Int J Cancer 120: 175–180. [DOI] [PubMed] [Google Scholar]
- 38. Ray PD, Huang BW, Tsuji Y (2012) Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal 24: 981–990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Guo J, Zheng L, Liu W, Wang X, Wang Z, et al. (2011) Frequent truncating mutation of TFAM induces mitochondrial DNA depletion and apoptotic resistance in microsatellite-unstable colorectal cancer. Cancer research 71: 2978–2987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Cooper CE, Nicholls P, Freedman JA (1991) Cytochrome c oxidase: structure, function, and membrane topology of the polypeptide subunits. Biochem Cell Biol 69: 586–607. [DOI] [PubMed] [Google Scholar]
- 41. Goto A, Matsushima Y, Kadowaki T, Kitagawa Y (2001) Drosophila mitochondrial transcription factor A (d-TFAM) is dispensable for the transcription of mitochondrial DNA in Kc167 cells. Biochem J 354: 243–248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chakrabarty S, D'Souza RR, Kabekkodu SP, Gopinath PM, Rossignol R, et al.. (2014) Upregulation of TFAM and mitochondria copy number in human lymphoblastoid cells. Mitochondrion. [DOI] [PubMed]
- 43. Sieh W, Kobel M, Longacre TA, Bowtell DD, deFazio A, et al. (2013) Hormone-receptor expression and ovarian cancer survival: an Ovarian Tumor Tissue Analysis consortium study. Lancet Oncol 14: 853–862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Faratian D, Zweemer AJ, Nagumo Y, Sims AH, Muir M, et al. (2011) Trastuzumab and pertuzumab produce changes in morphology and estrogen receptor signaling in ovarian cancer xenografts revealing new treatment strategies. Clin Cancer Res 17: 4451–4461. [DOI] [PubMed] [Google Scholar]
- 45. Hardie DG, Alessi DR (2013) LKB1 and AMPK and the cancer-metabolism link - ten years after. BMC Biol 11: 36. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Expression of Ki-67, MT-CO2 and Lon in EOC. Representative negative and positive immunohistochemical staining of Ki-67, MT-CO2 and Lon in EOC (magnification: 400x, scale bar shows 500 µm).
(TIF)
Ki-67 index distribution in different EOC subtypes. Distribution of Ki-67 index varied significantly across the different EOC subtypes (Mann-Whitney U, p = 0.003). High-grade serous carcinoma (HGSC) (n = 21), clear cell (CC) (n = 14), endometrial carcinoma (EC) (n = 10), mucinous carcinoma (MC) (n = 7) and low-grade serous carcinoma (LGSC) (n = 1).
(TIF)
Ki-67 index distribution in EOC tumours depending on expression of PGC1α and TFAM. Distribution of Ki-67 index varied significantly across the different groups of tumours with PGC1α-/TFAM- (n = 9), PGC1α-/TFAM+ (n = 2), PGC1α+/TFAM- (n = 9) and PGC1α+/TFAM+ (n = 33) (Kruskal-Wallis test, p = 0.048).
(TIF)
Growth rates in EOC SKOV-3 cells and the multiresistant subline SKOV-3-R. Growth rates in SKOV-3 and SKOV-3-R cells assessed as cellular protein at given time points using the sulphorhodamine B assay (n = 4). Data are expressed as fold increase from t = 0 h. S.E.M. error bars were too small to be visualized, except where shown.
(TIF)
PGC1α/TFAM expression and response to cisplatin treatment in SKOV-3 PPARGC1A siRNA knockdown cells. SKOV-3 cells were treated with PPARGC1A siRNA knockdown or siRNA negative control; (A) gene expression of PPARGC1A and TFAM was evaluated at 72 h post-transfection by qRT-PCR (n = 3). Expression levels were normalised to ACTB. Error bars represent S.E.M. (B) representative western blot showing protein expression at 72 h post-transfection of PGC1α, Ki-67 and TFAM. β-tubulin was used as loading control. (C) At 24 h post-transfection, SKOV-3 PPARGC1A siRNA knockdown and siRNA negative control cells were treated with indicated doses of cisplatin for 48 h, whereafter cellular protein was measured using the sulphorhodamine B assay (n = 3). Data are expressed as percent of untreated cells. Error bars represent S.E.M.
(TIF)
Supporting tables. Table S1, Expression of MT-CO2 in EOC. Shown is the distribution of positive and negative expression of MT-CO2 in the total sample, in the five EOC subgroups and in TFAM-expressing and non-expressing samples, respectively. Table S2, Expression of Lon in EOC. Shown is the distribution of positive and negative expression of Lon in the total sample, in the five EOC subgroups and in TFAM-expressing and non-expressing samples, respectively.
(DOCX)
S_PCR: Gene expression raw data given as Ct, delta-Ct (DCt) and delta-delta-Ct (DDCt) values, respectively, for results shown in Figs. 1 and 4, and Fig. S5. S_Spect: Spectrophotometry raw data given as absorbances and calculations based thereupon, for results shown in Figs. 4 and S5. S_IHC: The finalized scoring of immunohistochemical stainings.
(ZIP)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.