

Abstract
AIM: To examine the role of E-cadherin and beta-catenin in carcinogenesis and to assess their prognostic implication in Epstein-Barr virus-associated gastric carcinomas (EBV-GCs).
METHODS: We compared the frequency of E-cadherin and beta-catenin expression in 59 EBV-GCs and 120 non-EBV-GCs, and examined the association between patients' prognosis and the expressions of these proteins.
RESULTS: Neither the cellular-membranous nor the cytoplasmic E-cadherin expression showed any difference between EBV-GCs and non-EBV-GCs. On the other hand, loss of membranous expression of beta-catenin occurred more frequently in non-EBV-GCs than EBV-GCs [odds ratio = 0.41; 95% confidence interval (CI), 0.19-0.90]. Furthermore, the nuclear and/or cytoplosmic expression of beta-catenin was seen more frequently in EBV-GCs than non-EBV-GCs (odds ratio = 2.23; 95% CI, 0.97-5.09), and was observed in a larger proportion of carcinoma cells of EBV-GCs than non-EBV-GCs (P = 0.024). Survival analysis for non-EBV-GC revealed that lymph node metastasis was significantly associated with poor prognosis (P < 0.001). Among EBV-GCs, the depth of invasion (P = 0.005), lymph node metastasis (P = 0.004) and an intestinal type by Lauren classification (hazard ratio = 9.47; 95% CI, 2.67-33.6) were significantly associated with poor prognosis. On the other hand, nuclear and/or cytoplasmic expression of beta-catenin was associated with a better prognosis in patients with EBV-GC (hazard ratio = 0.32; 95% CI, 0.11-0.93).
CONCLUSION: We observed more frequent preservation of beta-catenin in cell membrane and accumulation in nuclei and/or cytoplasm in EBV-GCs than in non-EBV-GCs. Factors involved in the prognosis of EBV-GCs and non-EBV-GCs are different in the two conditions.
Keywords: Epstein-Barr virus, Gastric carcinoma, E-cadherin, Beta-catenin, Prognosis
INTRODUCTION
In situ hybridization (ISH) of Epstein-Barr virus-encoded small RNA (EBER), which became available in the early 1990s, showed that EBER can be detected in 1%-17% of gastric carcinomas[1-5]. Such Epstein-Barr virus-associated gastric carcinomas (EBV-GC) have a uniform expression of EBNA-1 and EBERs in all carcinoma cells, in addition to the episomal monoclonality of the EBV genome[6], elevated serum antibodies against EBV-related antigens[7,8], and the unique 'lace pattern' morphology in some early-stage-EBV-GCs[9]. These features strongly suggest an important etiological role of EBV in the development of EBV-GCs. However, the EBV genome(s) responsible for gastric carcinoma development has yet to be elucidated.
The EBV-gene expression in EBV-GC resembles type-Ipattern of latent infection. In this type of latency, EBERs and EBNA1 are always expressed but EBV LMP1, which is well known for its oncogenic potential, is not produced. Recently, it has been observed that EBERs activate insulin-like growth factor (IGF)-1 in EBV-GCs[10]. In Burkett’s lymphoma, another EBV-related malignancy showing the type-IEBV latency, EBERs induce not only interleukin (IL)-10 expression but also confer resistance to interferon (IFN)-α-induced apoptosis via binding to double strand RNA-activated protein kinase (dsPKR) and inhibiting its activation by IFN[11].
BARF1 and BARTs are other EBV-encoded transcripts expressed in EBV-GC[12]. The BARF1 gene is able to immortalize primary monkey and human epithelial cells in vitro[13,14], and furthermore, transfection of BARF1 gene into the rodent fibroblast cell line BALB/c 3T3 or into the EBV-negative B cell line Louckes, resulted in tumorigenic transformation[15,16]. Recent studies regarding BARTs reported that one of the products of BARTs, RK-BARF0, may activate the Notch signal[17] and affect Wnt and JNK pathways by binding to cellular humanI-mfa domain-containing proteins, which interact with Axin and are related to the regulation of the free levels of cytosolic beta-catenin[18,19].
Another feature related to EBV’s oncogenic potential is its ability to cause CpG-island hypermethylation of host genomes[20,21]. Recent studies have shown that EBV-GCs were associated with down-regulation of p16[20-23], E-cadherin[24] and other proteins[20,21], whose expression is frequently suppressed by hypermethylation of the host genomes in EBV-GCs.
EBV-GC is known for its male predominance, predisposition to the gastric fundic-gland region, and relatively high frequency of moderately-differentiated type and poorly-differentiated solid type compared to other histological types of gastric cancer[1-5]. Another interesting but not well-known feature of EBV-GC is the macroscopic finding of a clear demarcation from the surrounding mucosa when compared to non-EBV-GC[25]. Inspired by these clinical findings, we compared the expression of E-cadherin and beta-catenin in EBV-GCs and non-EBV-GCs. Furthermore, the prognostic significance of these proteins in EBV-GCs and non-EBV-GCs was also examined since they are related to carcinogenesis and prognosis of gastric cancer.
MATERIALS AND METHODS
Subjects
Most of the subjects included in the present study have been reported in our previous studies[23,26]. In brief, we examined all the gastric carcinomas diagnosed at Kagoshima City hospital during the period 1976-1992, and identified 74 cases of EBV-GC. After excluding 10 cases without information on follow-up or other clinical details necessary for the present study, a total of 64 EBV-GC cases were enrolled in the present study. Two non-EBV-GC cases for each EBV-GC case were selected, matched with respect to age, sex, tumor location, and depth of invasion. Because of insufficient tissue specimens for immunostaining, E-cadherin and beta-catenin could be examined in 59 EBV-GC and 120 non-EBV-GC cases. The present study was approved by the Institutional Review Board of the Faculty of Medicine, Kagoshima University, Japan.
Histology
Histologically, all cases were classified as intestinal- or diffuse-type carcinoma based on the classification of Lauren[27]. Tumor location, defined as the predominant location of the tumor, was divided into three sites: cardia or upper one-third, middle portion and antrum or lower one-third of the stomach according to the classification of the Japanese Research Society for Gastric Cancer[28]. The histological subtypes were: well-differentiated tubular adenocarcinoma (tub1), moderately-differentiated tubular adenocarcinoma (tub2), and mucinous carcinoma (muc), regarded as Lauren’s intestinal type. The subtypes were: solid poorly-differentiated adenocarcinoma (por1), non-solid poorly-differentiated adenocarcinoma (por2), and signet-ring cell carcinoma (sig), regarded as Lauren’s diffuse type. The depth of invasion was classified as mucosal, submucosal, propia muscularis and subserosal involvement. The tumor stage was determined by the TNM classification of UICC[29].
ISH assay to detect EBER
ISH assay of paraffin-embedded tissue samples obtained from the main tumor was conducted using a digoxigenin-labeled EBER-1 oligonucleotide probe as described in our previous study[23,26]. EBER-1 positive diagnosis was made based on a positive signal at microscopic examination. A lymph node section from a patient with infectious mononucleosis was used as positive control, and a sense probe for EBER-1 was used as negative control in each assay. More precisely, the presence of EBER-1-positive signal in the tumor cells but not in the surrounding normal epithelial cells was defined as EBV-GC, while presence of EBER-1-negative tumor cells was defined as non EBV-GC. All EBV-GCs had the uniform presence of EBER in tumor cells but not in the surrounding normal epithelial cells.
Immunostaining for E-cadherin and beta-catenin
Paraffin sections were dewaxed with xylene and rehydrated with alcohol. After the eradication of endogenous peroxidase activity with methanol and H2O2, sections were autoclaved for 5 min in citric acid buffer to retrieve the antigen, and then a primary monoclonal antibody recognizing human E-cadherin (TaKaRa Biomedicals, Kyoto, Japan) or beta-catenin (Transduction Laboratories, Lexington, KY, USA) was applied and the avidin-biotin-peroxidase complex method was used for immunohistochemical staining (VECTASTAIN ABC KIT, Vector Laboratories, Inc, USA). Without knowledge of the EBV status, the expression of each protein was graded as follows: (1) 0%, (2) less than 10%, (3) 10%-49%, (4) 50%-90%, and (5) more than 90% according to the proportion of tumor cells with a positive staining. Nuclear and membranous signals were evaluated independently. Normal epithelial mucosa in each section was used as a positive control. Since these proteins are expressed intercellularly at the cell membranes of normal cells, two abnormal staining patterns of E-cadherin and beta-catenin were observed: (1) loss of membranous expression, and (2) nuclear and/or cytoplasmic pattern with expression in more than 10% of the tumor cell cytoplasm or tumor cell nuclei. Since no EBV-GC case showed membranous expression of E-cadherin in more than 90% of the tumor cells, we decided to use 50% as cut-off level for the loss of membranous E-cadherin expression. With regard to membranous beta-catenin expression, we used 10% as cut-off level for the loss of beta-catenin[30].
Statistical analysis
Conditional logistic regression analysis was performed to examine the proportion of EBV-GCs and other parameters. To permit the inclusion of as many subjects as possible with E-cadherin and beta-catenin expression, we re-matched EBV-GC cases and non-EBV-GC cases by gender, age (± 5 years), tumor location (upper-, middle-, and lower one-third), and tumor stage (early and advanced). Two non-EBV-GC cases were excluded from the conditional logistic regression analysis because they did not match any of the EBV-GC cases with respect to E-cadherin and beta-catenin expression. Maximum likelihood estimates of odds ratios (ORs) and their corresponding 95% confidence intervals (CIs) were calculated. Tests for trend used consecutive integers for levels of ordered categories.
The Cox proportional hazards model was used in the multivariate survival analysis. Maximum likelihood estimates of hazard ratios (HRs), 95% CIs, and likelihood ratio statistics in Cox proportional hazards models were obtained with the use of a statistical package, EPICURE (HiroSoft International Corp., USA). Since we found a significant association between p53 expression and prognosis in patients with intestinal type of GC in our previous study[26], the results of p53 expression was also considered in the survival analyses. We calculated Wald-type confidence intervals. Test for statistical interaction was conducted by including a cross-product term of the two variables of interest in the model.
RESULTS
We compared the expression patterns of E-cadherin and beta-catenin in 59 EBV-GCs and 120 non-EBV-GCs. The average age at diagnosis was 62.4 years (SD = 10.4) in EBV-GC cases and 61.3 years (SD = 10.1) in non-EBV-GC cases. The results of E-cadherin-expression in EBV-GCs and non-EBV-GCs are summarized in Table 1. There was no significant difference in the membranous E-cadherin-expression between EBV-GC and non-EBV-GC (P = 0.985). Three non-EBV-GCs showed membranous expression of E-cadherin in > 90% of the tumor cells but this was not seen in any of the EBV-GC cases. E-cadherin membranous-expression showed no obvious difference between EBV-GCs and non-EBV-GCs in any of the histological types (data not shown). With regard to the nuclear and/or cytoplasmic expression, there was no difference between EBV-GCs and non-EBV-GCs (P = 0.475). Although 33 EBV-GCs and 61 non-EBV-GCs were positive for E-cadherin expression in the cytoplasm, only one non-EBV-GC showed detectable nuclear E-cadherin expression. Among the cases with cytoplasmic staining of E-cadherin, 3 EBV-GCs (9%) and 3 non-EBV-GCs (5%) showed a granular staining pattern with complete loss of membranous expression. Typical membranous and nuclear and/or cytoplasmic expressions of E-cadherin are shown in Figure 1.
Table 1.
Membranous and cytoplasmic expression of E-cadherin in EBV-GCs and non EBV-GCs
| Location of E-cadherin expression |
Number of the cases (%) |
OR1 | 95% CI1 | |
| EBV-GCs | non EBV-GCs | |||
| Total | 59 | 120 | ||
| Membranous | ||||
| Preserved (> 50%) | 14 (24) | 29 (24) | 1 | Referent |
| Lost (< 50%) | 45 (76) | 91 (76) | 0.99 | 0.48-2.05 |
| Cytoplasm | ||||
| Negative | 26 (44) | 59 (49) | 1 | Referent |
| Positive (subtotal) | 33 (56) | 61 (51) | 1.28 | 0.65-2.50 |
| 10%- | 17 (29) | 29 (24) | 1.37 | 0.62-3.01 |
| 50%- | 11 (19) | 29 (24) | 0.92 | 0.39-2.18 |
| 90%- | 5 (8) | 3 (3) | 3.98 | 0.88-18.0 |
| P = 0.374 | ||||
ORs and 95% CIs were obtained from conditional logistic regression analysis with the objective variable being EBV-GC (EBV-GC = 1, and non EBV-GC = 0).
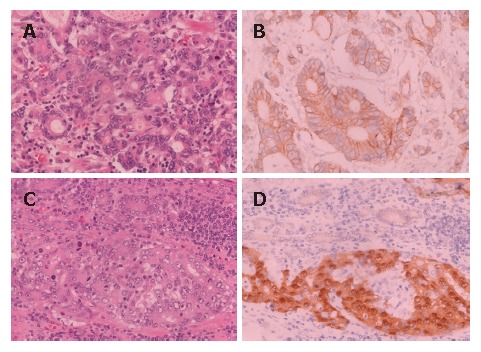
Figure 1.
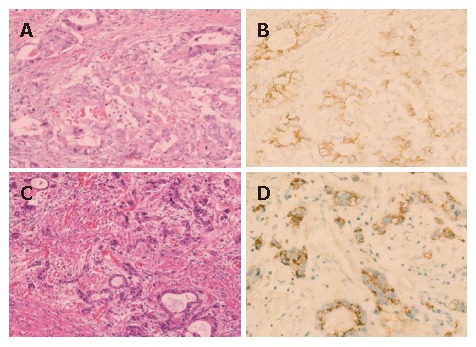
E-cadherin expression. A and B: an E-cadherin-positive case with membranous pattern; C and D: an E-cadherin-positive case with cytoplasmic pattern. E-cadherin expression is observed in cytoplasm but not in nuclei of carcinoma cells (D). A and C: hematoxylin and eosin staining; B and D: Immunohistochemical staining.
We also examined the beta-catenin expression (Table 2). There was more frequent loss of membranous beta-catenin expression in non-EBV-GCs than EBV-GCs (P = 0.019). The membranous expression showed no difference between EBV-GCs and non-EBV-GCs in any of the histological types (data not shown). Furthermore, the frequency of positive cases for nuclear and/or cytoplasmic beta-catenin expression in EBV-GCs (22%) was higher compared to non-EBV-GCs (12%), although this difference was statistically not significant (P = 0.059). In addition, the nuclear and/or cytoplasmic beta-catenin expression was observed in a larger proportion of carcinoma cells of EBV-GCs than non-EBV-GCs (P = 0.024). All but one non-EBV-GC with nuclear and/or cytoplasmic expression of beta-catenin showed detectable nuclear expression. Typical membranous and nuclear and/or cytoplasmic expression of beta-catenin is shown in Figure 2.
Table 2.
Membranous and nuclear and/or cytoplasmic expression of beta-catenin in EBV-GCs and non EBV-GCs
| Location of beta-catenin expression |
Number of the cases (%) |
OR1 | 95% CI1 | |
| EBV-GCs | non EBV-GCs | |||
| Total | 59 | 120 | ||
| Membranous | ||||
| Preserved (> 10%) | 49 (83) | 78 (65) | 1 | Referent |
| Lost (< 10%) | 10 (17) | 42 (35) | 0.41 | 0.19-0.90 |
| Nuclei and cytoplasm | ||||
| Negative | 46 (78) | 106 (88) | 1 | Referent |
| Positive (subtotal) | 13 (22) | 14 (12) | 2.23 | 0.97-5.09 |
| 10%- | 5 (8) | 8 (7) | 1.48 | 0.47-4.66 |
| 50%- | 6 (10) | 6 (5) | 2.34 | 0.74-7.36 |
| 90%- | 2 (3) | 0 (0) | - | |
| P = 0.024 | ||||
ORs and 95% CIs were obtained from conditional logistic regression analysis with the objective variable being EBV-GC (EBV-GC = 1, and non EBV-GC = 0).
Figure 2.
Beta-catenin expression. A and B: A beta-catenin-positive case with membranous pattern; C and D: Beta-catenin expression is observed in both nuclei and cytoplasm of carcinoma cells (D). A and C: hematoxylin and eosin staining; B and D: Immunohistochemical staining.
We examined the prognostic significance of E-cadherin and beta-catenin in EBV-GC and non-EBV-GC cases using Cox’s proportional hazard model taking into account the effect of p53 (Table 3). Details of the p53 overexpression among EBV-GCs and non-EBV-GCs have been reported elsewhere[26]. In brief, cases with both EBER expression and p53 over-expression showed the worst prognosis in patients with intestinal-type tumors. In diffuse-type tumor, however, only EBER was an important prognostic factor. For the assessment of survival analysis, only those subjects who underwent curative surgery were included in the analysis, thus the number of EBV-GC and non-EBV-GC cases was reduced to 49 and 100 respectively (Table 3). The average follow-up period was 70.1 months (SD = 62.5) in EBV-GC cases and 65.3 months (SD = 60.3) in non-EBV-GC cases. The survival analysis was first conducted separately for non-EBV-GCs and for EBV-GCs so that the factors involved in prognosis could be compared between the two groups. Survival analysis for non-EBV-GC cases revealed that only lymph node metastasis was significantly related to poor prognosis (P < 0.001). Cases with loss of membranous E-cadherin tended to have a poor prognosis, but the association was not significant (HR = 1.50; 95% CI, 0.70-3.22).
Table 3.
Results of survival analysis in EBV-GC and non EBV-GC cases
|
Non EBV-GCs |
EBV GCs |
|||||
| Death1/Total (%) | HR2 | 95% CI2 | Death1/Total (%) | HR2 | 95% CI2 | |
| Depth | ||||||
| m, sm | 6/29 (21) | 1 | Referent | 3/13 (23) | 1 | Referent |
| pm | 3/13 (23) | 0.76 | 0.19-3.07 | 2/8 (25) | 1.64 | 0.23-11.9 |
| s, ss, se | 37/58 (64) | 2.03 | 0.74-5.53 | 18/28 (64) | 6.03 | 1.39-26.2 |
| P = 0.112 | P = 0.005 | |||||
| Lymph node metastasis | ||||||
| 0 | 10/51 (20) | 1 | Referent | 13/32 (41) | 1 | Referent |
| 1 | 16/26 (62) | 3.55 | 1.44-8.77 | 4/9 (44) | 2.13 | 0.57-7.94 |
| 2+ | 20/23 (87) | 6.83 | 2.75-17.0 | 6/8 (75) | 7.77 | 1.93-31.3 |
| P < 0.001 | P = 0.004 | |||||
| Lauren classification | ||||||
| Diffuse | 14/30 (47) | 1 | Referent | 6/20 (30) | 1 | Referent |
| Intestinal | 32/70 (46) | 1.10 | 0.55-2.02 | 17/29 (59) | 9.47 | 2.67-33.6 |
| P = 0.865 | P < 0.001 | |||||
| p53 | ||||||
| Negative | 21/52 (40) | 1 | Referent | 13/26 (50) | 1 | Referent |
| Positive | 25/48 (52) | 1.60 | 0.87-2.91 | 10/23 (43) | 1.29 | 0.50-3.30 |
| P = 0.127 | P = 0.595 | |||||
| E-cadherin in membrane | ||||||
| Preserved | 9/28 (32) | 1 | Referent | 6/12 (50) | 1 | Referent |
| Lost | 37/72 (51) | 1.50 | 0.70-3.22 | 17/37 (46) | 0.73 | 0.25-2.12 |
| P = 0.276 | P = 0.568 | |||||
| E-cadherin in nuclei and/or cytoplasm | ||||||
| Negative | 30/62 (48) | 1 | Referent | 12/26 (46) | 1 | Referent |
| Positive | 16/38 (42) | 0.56 | 0.30-1.06 | 11/23 (48) | 1.37 | 0.56-3.34 |
| P = 0.071 | P = 0.481 | |||||
| Beta-catenin in membrane | ||||||
| Preserved | 28/64 (44) | 1 | Referent | 19/43 (44) | 1 | Referent |
| Lost | 18/36 (50) | 1.10 | 0.57-2.11 | 4/6 (67) | 1.39 | 0.42-4.60 |
| P = 0.778 | P = 0.601 | |||||
| Beta-catenin in nuclei and/or cytoplasm | ||||||
| Negative | 41/89 (20) | 1 | Referent | 18/37 (49) | 1 | Referent |
| Positive | 5/11 (45) | 0.81 | 0.31-2.09 | 5/12 (42) | 0.32 | 0.11-0.93 |
| P = 0.654 | P = 0.028 | |||||
Gastric-cancer related death.
HRs, 95% CIs, and P values were obtained by Cox proportional hazard models. Age, depth, lymph node metastasis, Lauren classification, and p53 expression were included in the models as covariates. The coefficient of age was 0.029 (SE = 0.018, P = 0.114) in non EBV-GCs, and 0.076 (SE = 0.026, P = 0.004) in EBV-GCs. m: mucosa, sm: submucosa, pm: propia muscularis, s: serosa, ss: subserosa, se: serosa exposed.
Among EBV-GCs, the depth of invasion (P = 0.005), lymph node metastasis (P = 0.004) and intestinal type by Lauren classification (HR = 9.47; 95% CI, 2.67-33.6) were related to poor prognosis. On the other hand, nuclear and/or cytoplasmic expression of beta-catenin showed a significant association with better prognosis of patients with EBV-GC (HR = 0.32; 95% CI, 0.11-0.93). All cases with beta-catenin expression in nuclei and/or cytoplasm (n = 10) who died were male patients and had intestinal type of tumors.
We next conducted survival analysis by combining EBV-GC and non-EBV-GC data and adjusting for the effects of age, depth, lymph node metastasis, Lauren classification, and p53 expression. The presence of EBER expression was not related to prognosis (HR = 1.40; 95% CI, 0.81-2.42). The apparent difference between EBV-GCs and non-EBV-GCs with respect to the prognostic effects of Lauren classification was examined by adding an interactive term between EBER and Lauren classification to a Cox regression model. The addition of this interactive term significantly improved the model (P = 0.002), indicating that the prognostic effect of EBER expression was different in the two histological types of Lauren classification. On the other hand, EBER expression did not significantly affect the prognostic effects of p53, E-cadherin, and beta-catenin status (data not shown).
There were 5 lympho-epithlioma-like carcinomas and 4 of them were EBER positive. Lympho-epithelioma-like carcinomas showed similar prognosis to the other histological types of gastric cancer.
DISCUSSION
The present study shows that membranous expression of beta-catenin was preserved more frequently in EBV-GCs than non-EBV-GCs. In addition, EBV-GCs tended to show more frequent beta-catenin expression in nuclei and/or cytoplasm compared to non-EBV-GCs. Nuclear beta-catenin expression is a hallmark of cancers with aberrant activation of the Wnt pathway, and accumulation of beta-catenin in nuclei has been observed in various tumors including gastric carcinoma[31-34]. In the nucleus, beta-catenin interacts with members of the T-cell factor/lymphoid enhancer factor family of transcription factors and stimulates the expression of target genes such as c-myc and cyclin D1[35-37]. However, it is possible that the up-regulation of beta-catenin is caused not by EBV but by the cofactors that are necessary for the development of EBV-GCs. One of the candidate cofactor is H pylori infection, although an association between H pylori and EBV infection has not been well established[38]. Since several studies have suggested that H pylori also dysregulates beta-catenin-dependent pathway[39,40], it is important to examine the interaction between H pylori and EBV infections, and beta-catenin expression.
Beta-catenin tyrosine-phosphorylation, which is caused mainly by epidermal growth factor receptor, and by two growth factors, c-met, and c-erbB2, disrupts adherent complex, consisting of E-cadherin, and, alpha and beta-catenins[41]. The resultant free beta-catenin is imported into the cytoplasm and is subjected to degradation through its binding to an APC/AXIN/GSK complex[42]. According to our previous studies, the expression levels of c-met[43] and c-erbB2 (unpublished data), did not show any signifi-cant difference between EBV-GCs and non-EBV-GCs; therefore, the increased beta-catenin expression, particularly cytoplasmic and nuclear expression, in EBV-GCs observed in the present study is unlikely to be explained by the decreased expression of these growth factors.
Among EBV-related proteins expressed in EBV-GCs, LMP2A may be involved in beta-catenin preservation, since the LMP2A is reported to inhibit glycogen synthase kinase (GSK) activity through AKT activation[44]. LMP2A may also act as decoy receptor for some tyrosine kinase, and inhibit beta-catenin phosphorylation[45]. Recently, Morrison et al[46], have reported that LMP2A activates the phosphatidylinositol 3-kinase (PI3K) and beta-catenin signaling pathways in epithelial cells. Interestingly, this phenomenon was observed in NPC but not in Hodgkin lymphoma although both these malignancies are classified as type II latency and express LMP2A[47,48], suggesting that EBV acts differently in the different tumor sites. However, this aberrant activation of the Wnt pathway can only be observed in a subset of EBV-GCs because some EBV-GCs do not show LMP2A expression at the RNA level[12].
We did not observe any difference in the frequency of E-cadherin expression between EBV-GCs and non-EBV-GCs. There are 4 possible mechanisms of abnormal expression of E-cadherin: (1) gene mutation, (2) posttranslational truncation or modification, (3) transcriptional repressor, and (4) promoter hypermethylation. With regard to the promoter hypermethylation, two conflicting results in EBV-GCs have been reported[20,24]. Further investigations are required to clarify the regulation of E-cadherin expression in gastric carcinomas.
The accumulation of beta-catenin in the nucleus of gastric-carcinoma cells is reported to have no significant effect on disease outcome[30,32,49]. On the other hand, survival analysis in the present study showed a significant association between nuclear and/or cytoplasmic expression of beta-catenin and a better prognosis in patients with EBV-GC (HR = 0.32; 95% CI, 0.11-0.93) but not in those with non-EBV-GC (HR = 0.81; 95% CI, 0.31-2.09). These observations do not conflict with each other since most gastric carcinomas are non-EBV-GCs. Recently, Nam et al[50] reported a significant association between Akt activation and better prognosis in gastric cancer patients, which may support our observation because EBV-LMP2A also activates Akt[46]. Although there are several reports suggesting that the loss of membranous E-cadherin and beta-catenin complex is associated with an unfavorable cancer progression and disease outcome, neither mem-branous expression of E-cadherin nor beta-catenin was related to disease outcome in our study.
The prognostic significance of EBV-encoded genome expression remains to be established. Until now, most studies have not shown any difference in prognosis between EBV-GCs and non-EBV-GCs. Our previous study confirmed this notion[26]. However, histology-specific analysis in the present study provided interesting findings; EBV-GCs had a poorer prognosis in intestinal type-tumors but a better prognosis in diffuse-type tumors when compared to non-EBV-GCs[26]. The poor prognosis of intestinal type EBV-GCs when compared to diffuse type EBV-GCs corresponds to histology-specific prognostic effect of EBV observed in our previous study. At the present time, it is difficult to explain the mechanism underlying these observations. However, it is apparent that factors related to the prognosis of EBV-GCs are different from those of EBV-negative tumors, suggesting that EBV may change the process of carcinogenesis in gastric cancer.
In summary, the present study shows greater preser-vation of beta-catenin in cell membrane and accumulation in nuclei and/or cytoplasm in EBV-GCs than in non-EBV-GCs. Survival analysis suggested that different factors are involved in the prognosis of EBV-GCs and non- EBV-GCs.
Footnotes
Supported by Grants-in-Aid for Scientific Research on Priority Areas (12218231, 17015037, and 18014024) of the Ministry of Education, Culture, Sports, Science and Technology, Japan
S- Editor Liu Y L- Editor Anand BS E- Editor Wang HF
References
- 1.Shibata D, Weiss LM. Epstein-Barr virus-associated gastric adenocarcinoma. Am J Pathol. 1992;140:769–774. [PMC free article] [PubMed] [Google Scholar]
- 2.Tokunaga M, Uemura Y, Tokudome T, Ishidate T, Masuda H, Okazaki E, Kaneko K, Naoe S, Ito M, Okamura A. Epstein-Barr virus related gastric cancer in Japan: a molecular patho-epidemiological study. Acta Pathol Jpn. 1993;43:574–581. doi: 10.1111/j.1440-1827.1993.tb03233.x. [DOI] [PubMed] [Google Scholar]
- 3.Corvalan A, Koriyama C, Akiba S, Eizuru Y, Backhouse C, Palma M, Argandoña J, Tokunaga M. Epstein-Barr virus in gastric carcinoma is associated with location in the cardia and with a diffuse histology: a study in one area of Chile. Int J Cancer. 2001;94:527–530. doi: 10.1002/ijc.1510. [DOI] [PubMed] [Google Scholar]
- 4.Morewaya J, Koriyama C, Akiba S, Shan D, Itoh T, Eizuru Y. Epstein-Barr virus-associated gastric carcinoma in Papua New Guinea. Oncol Rep. 2004;12:1093–1098. [PubMed] [Google Scholar]
- 5.Burgess DE, Woodman CB, Flavell KJ, Rowlands DC, Crocker J, Scott K, Biddulph JP, Young LS, Murray PG. Low prevalence of Epstein-Barr virus in incident gastric adenocarcinomas from the United Kingdom. Br J Cancer. 2002;86:702–704. doi: 10.1038/sj.bjc.6600107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Imai S, Koizumi S, Sugiura M, Tokunaga M, Uemura Y, Yamamoto N, Tanaka S, Sato E, Osato T. Gastric carcinoma: monoclonal epithelial malignant cells expressing Epstein-Barr virus latent infection protein. Proc Natl Acad Sci USA. 1994;91:9131–9135. doi: 10.1073/pnas.91.19.9131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Levine PH, Stemmermann G, Lennette ET, Hildesheim A, Shibata D, Nomura A. Elevated antibody titers to Epstein-Barr virus prior to the diagnosis of Epstein-Barr-virus-associated gastric adenocarcinoma. Int J Cancer. 1995;60:642–644. doi: 10.1002/ijc.2910600513. [DOI] [PubMed] [Google Scholar]
- 8.Shinkura R, Yamamoto N, Koriyama C, Shinmura Y, Eizuru Y, Tokunaga M. Epstein-Barr virus-specific antibodies in Epstein-Barr virus-positive and -negative gastric carcinoma cases in Japan. J Med Virol. 2000;60:411–416. doi: 10.1002/(sici)1096-9071(200004)60:4<411::aid-jmv8>3.0.co;2-8. [DOI] [PubMed] [Google Scholar]
- 9.Uemura Y, Tokunaga M, Arikawa J, Yamamoto N, Hamasaki Y, Tanaka S, Sato E, Land CE. A unique morphology of Epstein-Barr virus-related early gastric carcinoma. Cancer Epidemiol Biomarkers Prev. 1994;3:607–611. [PubMed] [Google Scholar]
- 10.Iwakiri D, Eizuru Y, Tokunaga M, Takada K. Autocrine growth of Epstein-Barr virus-positive gastric carcinoma cells mediated by an Epstein-Barr virus-encoded small RNA. Cancer Res. 2003;63:7062–7067. [PubMed] [Google Scholar]
- 11.Nanbo A, Takada K. The role of Epstein-Barr virus-encoded small RNAs (EBERs) in oncogenesis. Rev Med Virol. 2002;12:321–326. doi: 10.1002/rmv.363. [DOI] [PubMed] [Google Scholar]
- 12.zur Hausen A, Brink AA, Craanen ME, Middeldorp JM, Meijer CJ, van den Brule AJ. Unique transcription pattern of Epstein-Barr virus (EBV) in EBV-carrying gastric adenocarcinomas: expression of the transforming BARF1 gene. Cancer Res. 2000;60:2745–2748. [PubMed] [Google Scholar]
- 13.Griffin BE, Karran L. Immortalization of monkey epithelial cells by specific fragments of Epstein-Barr virus DNA. Nature. 1984;309:78–82. doi: 10.1038/309078a0. [DOI] [PubMed] [Google Scholar]
- 14.Karran L, Teo CG, King D, Hitt MM, Gao YN, Wedderburn N, Griffin BE. Establishment of immortalized primate epithelial cells with sub-genomic EBV DNA. Int J Cancer. 1990;45:763–772. doi: 10.1002/ijc.2910450432. [DOI] [PubMed] [Google Scholar]
- 15.Wei MX, Ooka T. A transforming function of the BARF1 gene encoded by Epstein-Barr virus. EMBO J. 1989;8:2897–2903. doi: 10.1002/j.1460-2075.1989.tb08438.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wei MX, Moulin JC, Decaussin G, Berger F, Ooka T. Expression and tumorigenicity of the Epstein-Barr virus BARF1 gene in human Louckes B-lymphocyte cell line. Cancer Res. 1994;54:1843–1848. [PubMed] [Google Scholar]
- 17.Kusano S, Raab-Traub N. An Epstein-Barr virus protein interacts with Notch. J Virol. 2001;75:384–395. doi: 10.1128/JVI.75.1.384-395.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kusano S, Raab-Traub N. I-mfa domain proteins interact with Axin and affect its regulation of the Wnt and c-Jun N-terminal kinase signaling pathways. Mol Cell Biol. 2002;22:6393–6405. doi: 10.1128/MCB.22.18.6393-6405.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Thornburg NJ, Kusano S, Raab-Traub N. Identification of Epstein-Barr virus RK-BARF0-interacting proteins and characterization of expression pattern. J Virol. 2004;78:12848–12856. doi: 10.1128/JVI.78.23.12848-12856.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kang GH, Lee S, Kim WH, Lee HW, Kim JC, Rhyu MG, Ro JY. Epstein-barr virus-positive gastric carcinoma demonstrates frequent aberrant methylation of multiple genes and constitutes CpG island methylator phenotype-positive gastric carcinoma. Am J Pathol. 2002;160:787–794. doi: 10.1016/S0002-9440(10)64901-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chong JM, Sakuma K, Sudo M, Ushiku T, Uozaki H, Shibahara J, Nagai H, Funata N, Taniguchi H, Aburatani H, et al. Global and non-random CpG-island methylation in gastric carcinoma associated with Epstein-Barr virus. Cancer Sci. 2003;94:76–80. doi: 10.1111/j.1349-7006.2003.tb01355.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Vo QN, Geradts J, Gulley ML, Boudreau DA, Bravo JC, Schneider BG. Epstein-Barr virus in gastric adenocarcinomas: association with ethnicity and CDKN2A promoter methylation. J Clin Pathol. 2002;55:669–675. doi: 10.1136/jcp.55.9.669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Koriyama C, Kitajima S, Nomoto M, Sueyoshi K, Itoh T, Eizuru Y, Yonezawa S, Akiba S. Loss of p16/CDKN2A protein in Epstein-Barr virus-associated gastric carcinoma. Oncology. 2004;67:81–87. doi: 10.1159/000080290. [DOI] [PubMed] [Google Scholar]
- 24.Sudo M, Chong JM, Sakuma K, Ushiku T, Uozaki H, Nagai H, Funata N, Matsumoto Y, Fukayama M. Promoter hypermethylation of E-cadherin and its abnormal expression in Epstein-Barr virus-associated gastric carcinoma. Int J Cancer. 2004;109:194–199. doi: 10.1002/ijc.11701. [DOI] [PubMed] [Google Scholar]
- 25.Itoh T, Tokunaga M. Clinicopathology of Epstein-Barr virus-related gastric carcinoma. Nihon Rinsho. 1997;55:363–367. [PubMed] [Google Scholar]
- 26.Koriyama C, Akiba S, Itoh T, Kijima Y, Sueyoshi K, Corvalan A, Herrera-Goepfer R, Eizuru Y. Prognostic significance of Epstein-Barr virus involvement in gastric carcinoma in Japan. Int J Mol Med. 2002;10:635–639. [PubMed] [Google Scholar]
- 27.Laurén P. The two histological main types of gastric carcinoma: diffuse and so called intestinal type Carcinoma. An attempt at histo-clinical classification. Acta Pathol Microbiol Scand. 1965;64:31–49. doi: 10.1111/apm.1965.64.1.31. [DOI] [PubMed] [Google Scholar]
- 28.Japanese Research Society for Gastric Cancer. Macroscopic Findings. In: Japanese classification of Gastric Carcinoma., editor. First English Edition. Tokyo: Kanehara Press; 1995. pp. 3–13. [Google Scholar]
- 29.Sobin LH, Wittekind C, editors . TNM Classification of Malignant Tumors. 5th ed. New York: Wiley-Liss; 1997. [Google Scholar]
- 30.Woo DK, Kim HS, Lee HS, Kang YH, Yang HK, Kim WH. Altered expression and mutation of beta-catenin gene in gastric carcinomas and cell lines. Int J Cancer. 2001;95:108–113. doi: 10.1002/1097-0215(20010320)95:2<108::aid-ijc1019>3.0.co;2-#. [DOI] [PubMed] [Google Scholar]
- 31.Iwamoto M, Ahnen DJ, Franklin WA, Maltzman TH. Expression of beta-catenin and full-length APC protein in normal and neoplastic colonic tissues. Carcinogenesis. 2000;21:1935–1940. doi: 10.1093/carcin/21.11.1935. [DOI] [PubMed] [Google Scholar]
- 32.Grabsch H, Takeno S, Noguchi T, Hommel G, Gabbert HE, Mueller W. Different patterns of beta-catenin expression in gastric carcinomas: relationship with clinicopathological parameters and prognostic outcome. Histopathology. 2001;39:141–149. doi: 10.1046/j.1365-2559.2001.01177.x. [DOI] [PubMed] [Google Scholar]
- 33.Kotsinas A, Evangelou K, Zacharatos P, Kittas C, Gorgoulis VG. Proliferation, but not apoptosis, is associated with distinct beta-catenin expression patterns in non-small-cell lung carcinomas: relationship with adenomatous polyposis coli and G(1)-to S-phase cell-cycle regulators. Am J Pathol. 2002;161:1619–1634. doi: 10.1016/s0002-9440(10)64440-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Horvath LG, Henshall SM, Lee CS, Kench JG, Golovsky D, Brenner PC, O'Neill GF, Kooner R, Stricker PD, Grygiel JJ, et al. Lower levels of nuclear beta-catenin predict for a poorer prognosis in localized prostate cancer. Int J Cancer. 2005;113:415–422. doi: 10.1002/ijc.20599. [DOI] [PubMed] [Google Scholar]
- 35.He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT, Morin PJ, Vogelstein B, Kinzler KW. Identification of c-MYC as a target of the APC pathway. Science. 1998;281:1509–1512. doi: 10.1126/science.281.5382.1509. [DOI] [PubMed] [Google Scholar]
- 36.Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'Amico M, Pestell R, Ben-Ze'ev A. The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad Sci USA. 1999;96:5522–5527. doi: 10.1073/pnas.96.10.5522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tetsu O, McCormick F. Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature. 1999;398:422–426. doi: 10.1038/18884. [DOI] [PubMed] [Google Scholar]
- 38.Luo B, Wang Y, Wang XF, Gao Y, Huang BH, Zhao P. Correlation of Epstein-Barr virus and its encoded proteins with Helicobacter pylori and expression of c-met and c-myc in gastric carcinoma. World J Gastroenterol. 2006;12:1842–1848. doi: 10.3748/wjg.v12.i12.1842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Murata-Kamiya N, Kurashima Y, Teishikata Y, Yamahashi Y, Saito Y, Higashi H, Aburatani H, Akiyama T, Peek RM, Azuma T, et al. Helicobacter pylori CagA interacts with E-cadherin and deregulates the beta-catenin signal that promotes intestinal transdifferentiation in gastric epithelial cells. Oncogene. 2007;26:4617–4626. doi: 10.1038/sj.onc.1210251. [DOI] [PubMed] [Google Scholar]
- 40.Franco AT, Israel DA, Washington MK, Krishna U, Fox JG, Rogers AB, Neish AS, Collier-Hyams L, Perez-Perez GI, Hatakeyama M, et al. Activation of beta-catenin by carcinogenic Helicobacter pylori. Proc Natl Acad Sci USA. 2005;102:10646–10651. doi: 10.1073/pnas.0504927102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wong NA, Pignatelli M. Beta-catenin--a linchpin in colorectal carcinogenesis? Am J Pathol. 2002;160:389–401. doi: 10.1016/s0002-9440(10)64856-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kolligs FT, Bommer G, Göke B. Wnt/beta-catenin/tcf signaling: a critical pathway in gastrointestinal tumorigenesis. Digestion. 2002;66:131–144. doi: 10.1159/000066755. [DOI] [PubMed] [Google Scholar]
- 43.Kijima Y, Hokita S, Yoshinaka H, Itoh T, Koriyama C, Eizuru Y, Akiba S, Aikou T. Amplification and overexpression of c-met gene in Epstein-Barr virus-associated gastric carcinomas. Oncology. 2002;62:60–65. doi: 10.1159/000048248. [DOI] [PubMed] [Google Scholar]
- 44.Swart R, Ruf IK, Sample J, Longnecker R. Latent membrane protein 2A-mediated effects on the phosphatidylinositol 3-Kinase/Akt pathway. J Virol. 2000;74:10838–10845. doi: 10.1128/jvi.74.22.10838-10845.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Scholle F, Longnecker R, Raab-Traub N. Epithelial cell adhesion to extracellular matrix proteins induces tyrosine phosphorylation of the Epstein-Barr virus latent membrane protein 2: a role for C-terminal Src kinase. J Virol. 1999;73:4767–4775. doi: 10.1128/jvi.73.6.4767-4775.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Morrison JA, Klingelhutz AJ, Raab-Traub N. Epstein-Barr virus latent membrane protein 2A activates beta-catenin signaling in epithelial cells. J Virol. 2003;77:12276–12284. doi: 10.1128/JVI.77.22.12276-12284.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Morrison JA, Gulley ML, Pathmanathan R, Raab-Traub N. Differential signaling pathways are activated in the Epstein-Barr virus-associated malignancies nasopharyngeal carcinoma and Hodgkin lymphoma. Cancer Res. 2004;64:5251–5260. doi: 10.1158/0008-5472.CAN-04-0538. [DOI] [PubMed] [Google Scholar]
- 48.Everly DN, Kusano S, Raab-Traub N. Accumulation of cytoplasmic beta-catenin and nuclear glycogen synthase kinase 3beta in Epstein-Barr virus-infected cells. J Virol. 2004;78:11648–11655. doi: 10.1128/JVI.78.21.11648-11655.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Miyazawa K, Iwaya K, Kuroda M, Harada M, Serizawa H, Koyanagi Y, Sato Y, Mizokami Y, Matsuoka T, Mukai K. Nuclear accumulation of beta-catenin in intestinal-type gastric carcinoma: correlation with early tumor invasion. Virchows Arch. 2000;437:508–513. doi: 10.1007/s004280000283. [DOI] [PubMed] [Google Scholar]
- 50.Nam SY, Lee HS, Jung GA, Choi J, Cho SJ, Kim MK, Kim WH, Lee BL. Akt/PKB activation in gastric carcinomas correlates with clinicopathologic variables and prognosis. APMIS. 2003;111:1105–1113. doi: 10.1111/j.1600-0463.2003.apm1111205.x. [DOI] [PubMed] [Google Scholar]