

Abstract
Prosociality can be defined as any behaviour performed to alleviate the needs of others or to improve their welfare. Prosociality has probably played an essential role in the evolution of cooperative behaviour and several studies have already investigated it in primates to understand the evolutionary origins of human prosociality. Two main tasks have been used to test prosociality in a food context. In the Platforms task, subjects can prosocially provide food to a partner by selecting a prosocial platform over a selfish one. In the Tokens task, subjects can prosocially provide food to a partner by selecting a prosocial token over a selfish one. As these tasks have provided mixed results, we used both tasks to test prosociality in great apes, capuchin monkeys and spider monkeys. Our results provided no compelling evidence of prosociality in a food context in any of the species tested. Additionally, our study revealed serious limitations of the Tokens task as it has been previously used. These results highlight the importance of controlling for confounding variables and of using multiple tasks to address inconsistencies present in the literature.
Keywords: prosocial behaviour, primates, other-regarding preferences, apes, monkeys
1. Introduction
Prosociality can be defined as any behaviour performed by one individual to alleviate the needs of other individuals or to improve their welfare, without the actor necessarily incurring extra costs to provide these benefits [1]. In evolutionary terms, prosociality might have played an essential role in the evolution of mutually beneficial cooperative behaviour by providing individuals with the psychological predisposition to be concerned for the welfare of others (e.g. [2–4]; but see [5]). Although prosocial behaviour is well documented among primates in feeding and agonistic contexts [6–8], pinpointing the precise motivations underlying this behaviour has been difficult. Whereas several studies have reported chimpanzees helping others to obtain objects (including tools) in both mutualistic and altruistic settings [9–13], studies involving food distribution have produced mixed results [14–29].
Two types of task (Platforms and Tokens) have commonly been used to measure primates' prosocial tendency to give food to partners. In the Platforms task, subjects are presented with sliding platforms and two options [14]. The subject always obtains the same amount of food regardless of the option chosen, but one of the options also provides food to the partner. By choosing this option, the subject can therefore benefit a partner without incurring any extra cost. Versions of this task have been used with several primate species. Chimpanzees (Pan troglodytes), for example, generally failed to give food to their partners [14–17]. By contrast, dominant long-tailed macaques (Macaca fascicularis) behaved prosocially towards subordinates [18]. However, the experimental set-up used in this last study makes the interpretation of the results not entirely straightforward because making a prosocial choice required the subject to sit closer to the partner, whereas making a non-prosocial choice required the subject to sit next to an empty room. It is conceivable that dominants might have preferred to sit next to the subordinate (to elicit grooming), whereas subordinates might have avoided the dominant's proximity (to avoid aggression). Further studies would be necessary to rule out this alternative interpretation.
Tufted capuchin monkeys (Sapajus apella) also gave food to their partners in two independent studies [19–20]. Similarly, common marmosets (Callithrix jacchus) prosocially and altruistically gave food to their partners [21]. Interestingly, cotton-top tamarins (Saguinus oedipus), despite also being a cooperative breeding species, did not prosocially provide food to other group members in two different studies [22–23]. Additionally, in a new version of this task, subjects could pull a platform to provide food rewards to all the other group members [24]. Capuchin monkeys and Japanese macaques (Macaca fuscata) failed to be prosocial, but marmosets, again, gave food to their group members [24]. In general, subjects in the studies above did not make more prosocial choices when the partner showed interest in the food reward, by for example reaching out for it (chimpanzees [16], cotton-top tamarins [22], common marmosets [21]; see [25]), suggesting that subjects may not have been responding to the needs of their partner. Despite its frequent use, the Platforms task has been criticized, first for being too complex from a technical and a social perspective, and second for the presence of visible food rewards which may elicit selfish and competitive behaviour rather than prosocial attitudes ([26–27]; but see [16,25])
In the Tokens task, subjects learn to associate different stimuli with different food distribution outcomes (e.g. food for the partner versus food for nobody, food for both versus food for the subject). Then subjects have the opportunity to choose between stimuli, thus determining who will receive food. Chang et al. [28] trained rhesus macaques (Macaca mulatta) to associate unique visual cues with specific food distributions and found that macaques preferred the prosocial visual stimulus (providing food to the partner) over the selfish one (in this case, providing food to nobody). However, this preference did not hold when subjects also received food rewards (i.e. they did not prefer the stimulus providing food to the partner and the subject over the one providing food to the subject). These results do not allow us to draw definitive conclusions, as the number of subjects was too small. More importantly, this study cannot rule out that subjects were not behaving prosocially, but were simply preferring the prosocial option when receiving no food reward, as the prosocial option was the only one associated with food. de Waal et al. [29] trained female capuchin monkeys to discriminate between tokens providing food only to the subject and tokens providing food also to their partners. Subjects significantly preferred the prosocial token over the selfish one. Also female chimpanzees preferred the prosocial token when tested with a similar set-up [27]. These studies, however, are problematic because they failed to rule out alternative explanations. In the study by de Waal et al. [29], for example, there were no control conditions investigating the behaviour of capuchin monkeys when subjects were provided with the same two tokens, without a partner present in the adjacent cage. Without this control condition, it is impossible to know whether subjects' choice reflected a prosocial tendency to give food to their partners, or a general preference for the choice associated with an overall higher amount of food rewards. Horner et al. [27] introduced no-partner control conditions, but they were always conducted after the experimental condition with the partner, and used different pairs of tokens to prevent subjects from learning that all tokens produced the same outcome. Therefore, it is impossible to rule out the possibility that the order in which conditions were administered and the type of tokens used affected the results. Moreover, both studies [27,29] failed to test whether subjects understood the task. Only recently, Suchak & de Waal [30] introduced a condition to test subjects' understanding of the task, as well as a no-partner control condition, but unfortunately the no-partner control always took place after the experimental condition with the partner, to try to avoid that subjects expected to receive both rewards. Therefore, no definitive conclusions can be drawn from these studies.
The main aim of this study was to use both the Platforms and the Tokens tasks to test prosociality in six primate species: chimpanzees (P. troglodytes), bonobos (Pan paniscus), gorillas (Gorilla gorilla), orangutans (Pongo abelii), tufted capuchin monkeys (S. apella) and Geoffroyi's spider monkeys (Ateles geoffroyi). Although prosociality has already been tested in some of these species, as far as we know, no previous study has investigated multiple species of monkeys and apes with the two most frequently used food providing prosociality tests. In the Platforms task, subjects could choose the prosocial platform over the selfish one to provide food to their partners, and in the Tokens task they could select a prosocial token over a selfish one to provide food to their partners. If the use of both tasks (with appropriate controls) on the same population leads to the same outcome, then results become more compelling. All the species included are good candidates for prosociality owing to their complex cognitive skills (great apes [31]), high levels of allo-maternal care (capuchin monkeys, which might thus show enhanced motivation to be prosocial [32,33]) and high levels of social tolerance (Pan, Pongo and spider monkeys [34]).
2. Material and methods
(a). Subjects
We tested seven spider monkeys (combined in 26 pairs) at the Animaya Zoo in Merida, Mexico, 10 tufted capuchin monkeys (combined in 18 pairs) at the ISTC-CNR Primate Centre in Rome, Italy, and 12 chimpanzees (combined in 34 pairs), nine bonobos (combined in 26 pairs), five orangutans (combined in 18 pairs) and seven gorillas (combined in 22 pairs) at the Wolfgang Koehler Primate Research Center in the Leipzig Zoo, Germany. Subjects were of both sexes and various age classes (adults, subadults and juveniles, according to [35,36]). They were all born in captivity, except for one gorilla (Bebe) and all the spider monkeys, who were born in the wild but were raised as pets before being rescued and brought to the zoo.
Subjects were all housed in groups with their group members, in enclosures with outdoor and indoor areas, and they were never deprived of food or water before or during the experiment. All of them were used to being temporarily isolated in testing rooms (with their infant if present) and were tested by the same familiar experimenter only after they were comfortable with the set-up and the testing room. All subjects had previously participated in experimental tasks, but they had not been previously tested in the same tasks administered in this study, with the exception of six chimpanzees tested by Jensen et al. [15] with a task similar to the Platforms task (also see the electronic supplementary material, table S1). For different reasons, not all dyads could be tested with both tasks. Some subjects were moved to/from different facilities during the study, other subjects were not motivated to exchange tokens, or could not be paired with the same partner when the second experiment started because the quality of their relationship worsened for no reason the experimenters could discern. Importantly, only pairs with a good quality of relationship were included in this study, consequently enhancing the possibility to detect prosocial behaviour (see [1]). Quality of relationship was assessed by the keepers working at the different facilities, excluding pairs that would have shown distress if being tested in two adjacent rooms. Finally, spider monkeys were only tested with the Platforms task. They were the only species with no previous experience at exchanging tokens (see the electronic supplementary material, table S1) and time constraints meant that training was not possible.
(b). Materials and procedure
(i). Platforms task
In this task, the subject was located in the testing room and faced an apparatus consisting of a table with three sliding platforms baited with different food rewards (figure 1). Food rewards were out of the subject's reach, but the subject could use handles to pull in two of the three platforms (the prosocial one, or P, and the selfish one, or S). After selecting either S or P by pulling the corresponding handle, the subject could reach the food on the chosen platform, while the other platform moved back, out of the subject's reach. The third platform could be pulled by neither the subject nor the partner, but it was connected to P: if the subject pulled P, the third platform moved towards the adjacent room, where the food could be retrieved by the partner. If the subject selected S, the third platform moved further away from the partner, who then received no food. In contrast to other studies [14,21], we did not use stacked platforms, and the third platform was physically separated from P and S. Each dyad received one training condition and three experimental conditions: Equal, Unequal and Equidistant (with three corresponding control conditions).
Figure 1.
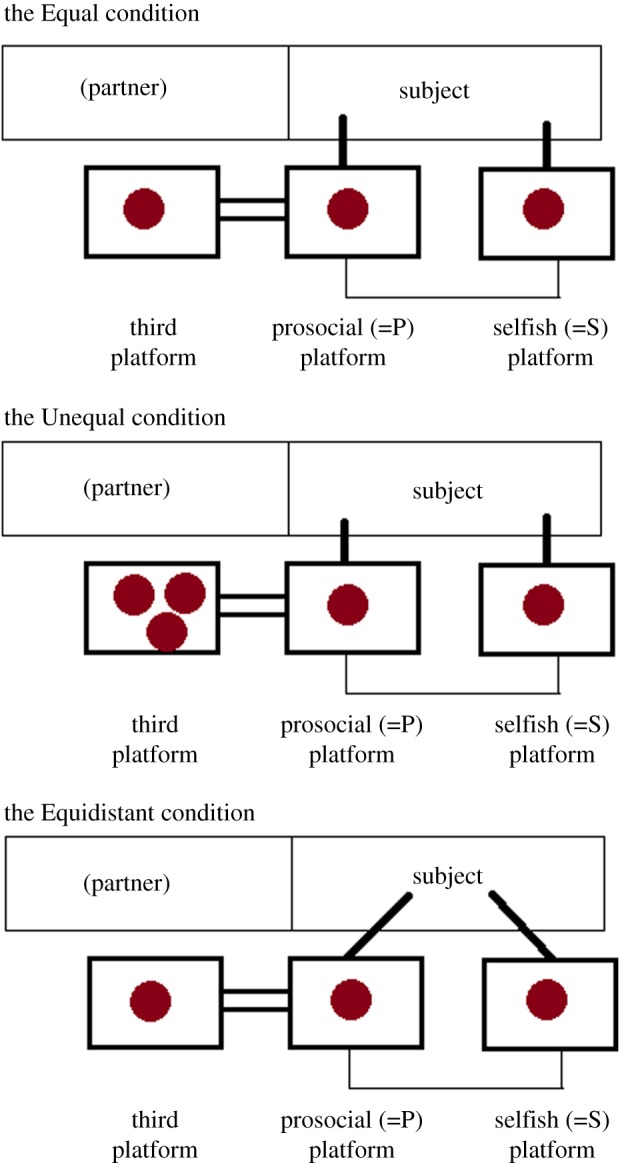
Set-up in the three conditions of the Platforms task. (Online version in colour.)
In the Training condition, the subject was tested alone. Sessions were alternated so that the subject had access exclusively to her testing room (and thus S and P), or also to the adjacent room (and thus also to the third platform). Each session was composed of six trials, in which only one of the three platforms was baited (randomizing the order and baiting each platform twice in each session). Once the platforms were baited, the subject had 30 s to make a selection. Subjects were considered to have knowledge of the set-up and could therefore move to the following condition if: (i) they always pulled S when S was baited and always pulled P when P was baited in two consecutive sessions in which they had only access to S and P; and if (ii) they always pulled S when S was baited and P when P or the third platform were baited in two consecutive sessions in which they had access to all three platforms. Subjects required a mean of 3.4 sessions (with two sessions being the minimal number of sessions required to master the Training condition) to advance to the next condition.
In the Equal condition, the subject was in the testing room and the partner in the adjacent room. In full view of the subject, the Experimenter (E) baited all three platforms with an equal food reward (a slice of banana, a raisin or a pellet, depending on the subject's preference). As soon as the last platform was positioned and baited, the subject had 30 s to select one of the two platforms (the prosocial one, P, or the selfish one, S) by pulling it and retrieving the food. In this condition, subjects had therefore to get closer to their partner to make the prosocial choice, something that we controlled for in the Equidistant condition (see below). In the corresponding control condition, the subject was in the testing room and the partner in a room other than the adjacent one, which was empty and visible to the subject. E followed exactly the same procedure as before, but when the subject pulled P, E retrieved the food on the third platform and put it back in the food bucket.
In the Unequal and Equidistant conditions, E followed exactly the same procedure as the Equal condition, with the exception that: (i) in the Unequal condition the third platform was baited with three food rewards instead of one, and (ii) in the Equidistant condition the handles of S and P did not point straight to the subject, but 45° towards each other, so that the subject did not have to get closer to the partner's adjacent room in order to pull P. The Unequal condition allowed us to understand whether prosocial choices were facilitated by an equal/unequal food distribution between subject and partner. The Equidistant condition, instead, allowed us to control whether prosocial/selfish choices depended on whether the subject preferred to approach or to avoid the side of the partner's room. The corresponding control conditions were identical, but the partner was in a room other than the adjacent one, which was empty and visible to the subject, and E retrieved the food on the third platform and put it back in the food bucket in case the subject chose P.
After mastering the training, dyads were administered the Equal condition (randomizing the order in which the three experimental and the three control sessions were administered), followed by the Unequal condition (randomizing the order of the three experimental and control sessions) and by the Equidistant condition (with half of the subjects starting with the experimental and half with the control session). In all conditions, sessions consisted of 12 trials. The inclusion of the three conditions allowed us to explore the potential effects of food inequality and inter-individual distance when making choices.
(ii). Tokens task
Each subject was tested with up to three partners, and with each partner a different pair of tokens was used (see the electronic supplementary material, table S1). Great apes were tested with the same three pairs of tokens, while capuchin monkeys were tested with three different pairs of tokens, to ensure that no species was tested with tokens similar to those already used in the past. Tokens were little Plexiglas cylinders or cuboids (approx. 4–6 cm in size) painted in different colours. If subjects were paired with more than one partner, a different pair of tokens was used for each of the partners. Following Horner et al. [27], subjects were tested with a second partner only after all the conditions with the first partner had been run, and only after subject and partner had also switched their roles (if this dyad was also planned). For each dyad, we administered five conditions.
Unless otherwise stated, we followed the procedure by Horner et al. [27]. In the Preference condition, we tested whether subjects preferred one of the two tokens, in each pair they had been assigned. Subjects were tested alone and underwent one session of 10 trials. In each trial, subjects were presented with the two different tokens and could choose one of the two by pointing or trying to reach for it. The position of the tokens (right/left) was randomized and counterbalanced across trials. Regardless of the choice made, subjects were rewarded with one pellet wrapped in paper. In this task, food rewards were always wrapped in paper so that subjects were not distracted by visible food and could rely on both sound and sight to know whether the partner had been rewarded, as unwrapping the paper made noise and attracted the subject's attention (see [27]). Subjects were considered to have a token preference when one of the two tokens was selected in more than or equal to 80% of the trials. As this only happened in four out of 55 dyads, and in each case the preferred token was a different one, we did not change any pair of tokens, as done in [27,29].
In the Training condition, one of the two tokens within each pair was considered the prosocial token (P) and the other one the selfish token (S). For each species, the same token was considered to be P in approximately half of the dyads tested with that pair of tokens, and it was considered to be S in the other half of the dyads, to avoid that our results were affected by any intrinsic token properties. The Training condition consisted of one session of 10 trials. In this condition, the subject was in the testing room and the partner in an adjacent room. In each trial, E placed one of the two tokens in a bin hanging in the subject's room. The tokens were randomized and counterbalanced across trials, so that the same token was not given more than three times in a row. After the token was placed in the bin, E asked the subject to give it back. The subject had 4 min to hand it back to E, who then placed it in a visible position between the subject and the partner. If E received P, E took two food rewards from a bucket, held them up briefly, one in each hand, to make them visible to the subject and partner, and then gave one food reward to the subject and then one to the partner. If E received S, E took one food reward from the bucket, held it briefly in one hand to make it visible to the subject and partner, and then only rewarded the subject. Food rewards were always wrapped in paper. By the end of this condition, subjects and partners had experienced five trials with a selfish outcome and five trials with a prosocial one.
In the Experimental condition, as in the Training, the subject was in the testing room and the partner in an adjacent room. This condition consisted of one session of 30 trials. At the beginning of the session, E placed 15 S and 15 P tokens in the bin and then asked the subject to hand one back within 4 min. E placed the returned token in a visible place between the subject and the partner, before rewarding them with wrapped food rewards like in the Training condition. At the end of the trial, E placed the token back in the bin and started another trial. If subjects did not give back any token, the session was interrupted and continued on the next possible day. If subjects selected more than one token from the bin, each returned token was considered as a choice, tokens were rewarded sequentially, with each token displayed as the appropriate reward was offered and then placed out of the subjects' view, and all the tokens were placed back together in the bin.
In the Control condition, the subject was in the testing room but the partner was in a room other than the adjacent one (so that the adjacent room was visibly empty to the subject). E followed exactly the same procedure as in the Experimental condition, but when she was given the prosocial token P, E rewarded the subject and then pretended to reward a partner in the adjacent testing room, which was empty, by pretending to put the wrapped food reward through the mesh, hiding it in the hand and eventually placing it back in the bucket out of the subject's view (see [27]). In contrast to the study from Horner et al. [27], dyads were tested with the same pair of tokens as in the Experimental condition, in order to rule out the possibility that different performance across conditions might depend on the different pair of tokens used. Also differing from Horner et al. [27], subjects did not always run the Control condition after the Experimental condition. After completing the Preference and Training conditions, instead, half of the subjects underwent the Experimental before the Control condition, and half of the subjects underwent the Control condition before the Experimental one (see the electronic supplementary material, table S1). In this way, we ensured that the order in which conditions were administered could not account for any differences in performance between the two conditions.
In the Solitary condition, the subject was in the testing room but also had access to the adjacent room. E followed the same procedure as in the Experimental condition, but when the subject gave her P back, E placed the second wrapped food reward in the adjacent testing room, so that the subject could obtain it by simply moving to that room. If subjects had knowledge of the set-up, they should select P significantly more than chance to obtain double the amount of food. This condition was always administered after the Experimental and Control conditions, to avoid subjects expecting both rewards in all conditions and thus fostering competitive attitudes and hindering prosocial behaviour in the Experimental condition [27].
(c). Scoring and data analysis
In each condition of the two tasks, E coded live whether subjects selected the prosocial (P) or the selfish (S) platform or token. We videotaped all the trials (1.7% of the trials were not recorded due to technical problems) and later checked the live-scored behaviour against the videotapes for accuracy. The videos are digitally archived at the Max Planck Institute for Evolutionary Anthropology in Leipzig and are available on request. A second observer coded 19% of all the trials performed (3589/18 984 trials in the Platforms task, and 983/4950 trials in the Tokens task) to assess the inter-observer reliability, which was very good (Cohen's k = 0.94 in both cases).
In both tasks, our dependent variable was the percentage of P choices. Non-parametric Wilcoxon tests were used to compare performance between conditions (for all species and for each species separately). In the Tokens task, we repeated our analyses only including those dyads in which the Experimental condition was performed before the Control condition (as in [27]), in order to assess the impact of condition order on the subjects' performance. Additionally, we used binomial tests to investigate subjects' knowledge of the set-up, by analysing whether subjects selected P significantly more than chance in the Solitary condition, for each dyad separately. When this was the case, Wilcoxon tests for each dyad were run to compare performance between Experimental and Control conditions. No analyses were done for the Training condition in the Platforms task, as almost all subjects immediately mastered the task and could move to the following condition. All tests were exact and two-tailed, and the α level was set at 0.05. However, when analyses were conducted separately for each dyad, Bonferroni–Holm corrections were added to control for multiple comparisons. Finally, to control that our conclusions were robust to different forms of analyses, we also repeated our analyses using linear mixed models, which provided the same results (see the electronic supplementary material).
3. Results
(a). Platforms task
Overall subjects did not choose the prosocial platform (P) significantly more often in the Experimental than in the Control sessions. In fact, they selected P more often in the Control than in the Experimental sessions in the Equal condition (n = 52, T = 2915, p = 0.031; P choices in Experimental: 27.0%) and in the Unequal condition (n = 62, T = 2881, p = 0.004; P choices in Experimental: 24.0%), but showed no significant preference in the Equidistant condition (n = 37, T = 1195, p = 0.201; P choices in Experimental: 22.9%).
Analysing each species separately in the Equal condition revealed that orangutans, capuchin monkeys and spider monkeys selected P more often in the Control than in the Experimental sessions (orangutans: n = 11, T = 95.5, p = 0.043; capuchin monkeys: n = 10, T = 96.5, p = 0.037; spider monkeys: n = 16, T = 224, p = 0.033; figure 2a). By contrast, gorillas selected P more often in Experimental than Control sessions (n = 10, T = 55.5, p = 0.048). Neither chimpanzees nor bonobos showed a significant difference between conditions (chimpanzees: n = 10, T = 120, p = 0.326; bonobos: n = 8, T = 52, p = 0.334).
Figure 2.

Mean percentage of prosocial choices (+s.e.) in the (a) Equal condition, (b) Unequal condition and (c) Equidistant condition of the Platforms task, as a function of species. Only gorillas made the prosocial choice significantly more in the experimental (grey bars) than in the control (white bars) Equal condition (p = 0.048). The black line represents chance performance (at 50%).
In the Unequal condition, chimpanzees and spider monkeys selected P more often in the Control than in the Experimental sessions (chimpanzees: n = 15, T = 176, p = 0.006; spider monkeys: n = 19, T = 262.5, p = 0.006). All other species showed no significant difference between conditions in their choice of P (bonobos: n = 9, T = 70, p = 0.094; gorillas: n = 9, T = 59, p = 0.124; orangutans: n = 7, T = 29, p = 0.117; capuchin monkeys: n = 8, T = 66, p = 0.419; figure 2b).
Finally, in the Equidistant condition, no species selected P more often in one condition than the other (chimpanzees: n = 7, T = 56, p = 0.480; bonobos: n = 4, T = 11, p = 0.375; gorillas: n = 8, T = 48, p = 0.194; orangutans: n = 4, T = 14, p = 0.563; capuchin monkeys: n = 6, T = 40, p = 0.571; spider monkeys: n = 13, T = 106, p = 0.165; figure 2c). Bonobos were the only species that selected P more often in the Experimental than in the Control sessions, in all conditions, but this difference was never significant.
(b). Tokens task
Overall subjects did not choose the prosocial token (P) significantly more often in the Experimental over the Control condition (n = 26, T = 564, p = 0.801; P choices in Experimental: 49.5%). This was true also when analysing each species separately (chimpanzees: n = 8, T = 74, p = 0.445; bonobos: n = 7, T = 29.5, p = 0.422; gorillas: n = 6, T = 23, p = 0.156; orangutans: n = 4, T = 15, p = 0.953; capuchin monkeys: n = 6, T = 24.5, p = 0.414; figure 3).
Figure 3.
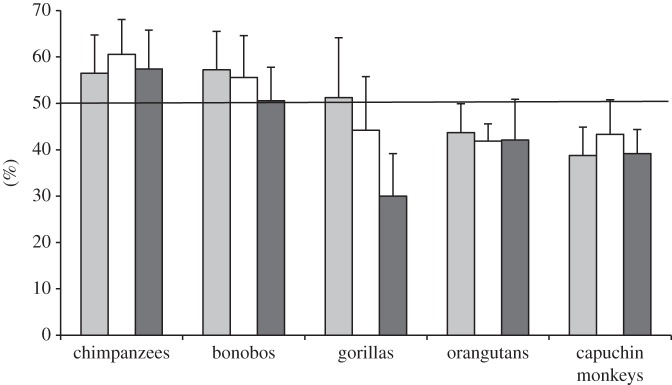
Mean percentage of prosocial choices (+s.e.) in the Experimental (grey bars), Control (white bars) and Solitary (black bars) conditions of the Tokens task, as a function of species. The black line represents chance performance (at 50%).
However, subjects that received the Experimental before the Control condition selected P significantly more often in the Experimental over the Control condition (n = 16, T = 187.5, p = 0.046; P choices in Experimental: 56.2%).
Crucially, subjects selected P at chance levels in the Solitary condition (n = 31, T = 803.5, p = 0.300; P choices in Solitary: 43.8%). This was also true when analysing each species separately (chimpanzees: n = 10, T = 106.5, p = 0.374; bonobos: n = 6, T = 41, p = 0.894; gorillas: n = 6, T = 30.5, p = 0.094; orangutans: n = 5, T = 16, p = 0.313; capuchin monkeys: n = 6, T = 31, p = 0.086; figure 3). When separately analysing each of the 55 dyads, however, 10 subjects chose P significantly more often than expected by chance in the Solitary condition. Within these dyads, however, none of the subjects chose P significantly more in the Experimental than in the Control condition (table 1).
Table 1.
For each species, list of the dyads in which the subject chose the prosocial token significantly more than chance in the Solitary condition of the Tokens task (Binomial test), with the percentage of prosocial choices made, the p-values for the Binomial test and the results for the Wilcoxon test when comparing the subject's percentage of prosocial choices in the Experimental and Control conditions.
| species | subject | partner | percentage of prosocial choices in solitary condition | binomial test (p-value) | Wilcoxon test (experimental versus control condition) |
|---|---|---|---|---|---|
| chimpanzees | Fifi | Jahaga | 90 | <0.001 | n = 9, T = 76.5, p = 0.804 |
| Fifi | Trudi | 93 | <0.001 | n = 7, T = 49, p = 1.000 | |
| Jahaga | Fifi | 97 | <0.001 | n = 1, T = 1.5, p = 1.000 | |
| Jahaga | Trudi | 100 | <0.001 | n = 0, T = 0, p = 1.000 | |
| Lome | Fraukje | 100 | <0.001 | n = 5, T = 20, p = 0.453 | |
| Tai | Fraukje | 87 | <0.001 | n = 5, T = 22.5, p = 0.727 | |
| Trudi | Fifi | 100 | <0.001 | n = 0, T = 0, p = 1.000 | |
| bonobos | Fimi | Yasa | 90 | <0.001 | n = 3, T = 7.5, p = 0.625 |
| Gimena | Yasa | 93 | <0.001 | n = 6, T = 24, p = 0.125 | |
| orangutans | Pini | Dokana | 90 | <0.001 | n = 10, T = 90, p = 0.629 |
4. Discussion
Overall, our results provided no compelling evidence of prosocial behaviour in any of the primate species tested with the two different tasks. None of the species, regardless of their socio-ecological characteristics and cognitive capacities, showed a consistent preference for selecting the prosocial option when a partner was present (Experimental sessions) compared with when the partner was absent (Control condition). In the Equal condition of the Platforms task, no species but gorillas chose the prosocial option significantly more in the Experimental than in the Control condition. However, gorillas' preference for the prosocial option in the presence of conspecifics was not replicated in any other condition. It is therefore difficult to conclude that gorillas were behaving prosocially towards their conspecifics. In the Unequal condition, no species showed evidence of prosocial behaviour, although the lack of prosocial behaviour might have been a consequence of an aversion to disadvantageous inequity (e.g. [37,38]; but see [19]). Finally, no species showed prosocial behaviour in the Equidistant condition. These results are consistent with previous studies that failed to detect prosocial behaviour in chimpanzees using a similar task [14–17].
Our results contrast with studies showing prosocial behaviour in capuchin monkeys tested with the Platforms task [19,20]. One reason why results may differ for capuchin monkeys is that a high variability might exist across populations in terms of prosociality and other behaviours potentially linked to prosociality, like social tolerance. One might speculate that the capuchin monkeys tested in our study were less socially tolerant than other capuchin monkey populations [39], and might therefore fail to behave prosocially towards each other (see [34] for a test of social tolerance including some of the same individuals). This explanation, however, is weakened by the fact that in our study, we only tested tolerant dyads (see Material and methods). Moreover, even species showing high social tolerance levels in [34] showed no evidence of prosociality in this study. Unfortunately, no data are currently available comparing the levels of social tolerance across different populations of capuchin monkeys. This means that explaining our findings in terms of a lack of social tolerance in the tested populations is conceivable, but it awaits empirical verification.
Our results extend previous findings on chimpanzees to other primate species, suggesting that prosocial behaviour as assessed with food tasks may not be a robust phenomenon, although groups and/or populations might differ in their levels of prosociality, as mentioned above. One surprising finding was that some species were not simply indifferent to the partner in the Platforms task, but made even fewer prosocial choices when the partner was present. In the Equal condition, orangutans, capuchin monkeys and spider monkeys preferred the prosocial option less often when their partner was present than when absent, and the same was true for chimpanzees and spider monkeys in the Unequal condition. Although these differences were not fully consistent across conditions and not present for each species, one could argue that some species behaved antisocially towards their group members. Alternatively, these species might simply be more reluctant to approach the partner in the testing situation. Recall that in both the Equal and Unequal conditions, subjects had to get closer to the partner to make the prosocial choice. In these conditions, therefore, subjects might have failed to behave prosocially simply to avoid approaching the partner. In line with this interpretation, all species ceased being antisocial in the Equidistant condition (i.e. they became indifferent), when subjects did not need to get closer to the partner to select the prosocial option. Therefore, partner's proximity is an important factor to consider not only when studying prosociality, but also in social competition tasks [40]. Consequently, the stacked platforms set-up [14,21,41] seems a better option to study food-distribution-based prosociality than the side-by-side set-up used in our study. Note, however, that even in the Equidistant condition, no species was prosocial, which means that lack of prosociality in our Platforms task cannot be entirely ascribed to our set-up.
Prosocial behaviour was not elicited by the Tokens task either. In contrast to previous studies highlighting the advantages of this procedure [27,29,30], our study revealed several important limitations, at least in the way that this procedure has been used so far. The most important limitation was that the vast majority of the subjects failed to understand the contingencies governing the task, in contrast to what happened in the Platforms task, where subjects mastered the procedure after a mean of only 3.4 sessions. This was clearly evidenced by the fact that 45 out of 55 dyads having access to both testing rooms (Solitary condition) failed to select the prosocial token significantly above chance level, thus missing the opportunity to double the amount of food obtained. Moreover, none of the subjects of these remaining 10 dyads that selected the prosocial token significantly more often than chance in the Solitary condition chose the prosocial option significantly more often when the partner was present. In short, most of the subjects did not understand the task, and those who did showed no evidence of prosocial behaviour. Note that exchanging objects for food per se was not the problem here. Following previous studies [27,29,30], we did not train subjects to reach a criterion. However, most of the subjects and species included in our study had already exchanged objects for food in previous studies [42–46] and all subjects readily exchanged tokens in this study (see the electronic supplementary material, table S1). In retrospect, our results highlight the importance of assessing whether the subjects understand the task before drawing conclusions from the resulting data.
Our study also demonstrated that the order in which conditions are administered significantly affects the results. Although subjects did not select the prosocial token more often when a partner was present as opposed to absent, such a preference was detected when analysing only the dyads in which the Experimental condition (partner present) was administered before the Control condition (partner absent). Randomizing the order of conditions is therefore essential to obtain unbiased data on prosocial behaviour, something that previous studies did not fully achieve.
Our results based on two tasks contrast with those previously reported for capuchin monkeys [29,30] and chimpanzees [27]. However, the caveats that we have identified above require that those results are interpreted with great caution. In particular, we still failed to obtain consistent evidence of prosocial behaviour in the tested species, despite our best attempts of eliciting prosocial behaviour, for instance by following Horner et al.'s [27] procedure and ensuring the inclusion of all their procedural improvements (e.g. use of wrapped food rewards not to distract subjects with visible food, use of transparent panels or mesh between subject and partner to allow communication, avoidance of competitive attitudes by always allowing subjects access to the partner's room in the Solitary condition only after administering the Experimental condition).
Finally, as de Waal et al. [29] and Horner et al. [27] only tested females, one could argue that sex differences could explain the discrepancies between the current and previous studies. However, none of the females who responded appropriately in the Solitary condition of the Tokens task behaved prosocially towards their partners. This means that the inclusion of males in our sample cannot explain the differences with previous studies. Although our findings with the Tokens task contrast with those in the three studies that have so far been done with the same task, our results are quite consistent with the data from the Platforms task.
In conclusion, the results of both the Platforms and the Tokens tasks (with their respective control conditions) did not provide compelling evidence of prosocial behaviour for any of the six primate species tested. When food distribution was involved, great apes, spider monkeys and capuchin monkeys failed to show prosocial behaviour towards group members, despite being paired with different partners and having the chance to provide them with food at no extra cost to themselves. These results highlight the importance of implementing tasks that control for confounding variables and of using different experimental paradigms when previous results contradict one another. Although it is still possible that other populations perform differently when tested with the same task (because of differences in diet quality, housing or personality, for example [47]), administering multiple tasks (with control conditions) is necessary to demonstrate prosociality. Finally, it is important to stress that our conclusions are limited to prosocial behaviour in an active food distribution context and do not allow us to generalize our findings to other contexts, like helping behaviour [9–13] or passive food transfers [7,8], as prosocial behaviour might be restricted to specific contexts and not generalize to active food-sharing [10].
Supplementary Material
Acknowledgements
We thank Alejandro Sánchez Amaro, Jessica Ferretti and Abraham Mesa Barroso for help during data collection, as well as Roberto Pacheco Mendez, Fernando Victoria Arceo, Iber Rodriguez Castillo and all the animal keepers at the different facilities for endless support and cooperation. Thanks to Elisa Felsche for coding data for inter-observer reliability purposes, to Shona Duguid for revising a previous version of the manuscript, to Roger Mundry for implementing the statistical models in the electronic supplementary material and to Hanna Petschauer for helping with organizing data collection.
Ethics statement
Animal husbandry and research comply with the “EAZA Minimum Standards for the Accommodation and Care of Animals in Zoos and Aquaria”, the “WAZA Ethical Guidelines for the Conduct of Research on Animals by Zoos and Aquariums” and the “Guidelines for the Treatment of Animals in Behavioural Research and Teaching” of the Association for the Study of Animal Behaviour (ASAB). Subjects were housed in large enclosures and lived in well-established groups with conspecifics. They participated in the tasks on a completely voluntary basis and were never food or water deprived. Data collection consisted in the administration of simple cognitive tasks, which were not invasive and strictly adhered to the legal requirements of Germany, Mexico and Italy, the countries were the primates were housed. Subjects received extra food when correctly solving the tasks and were never punished for incorrect performance. No medical, toxicological or neurobiological research of any kind was conducted on the subjects tested. All the protocols used in this study were ethically approved by an internal committee at the Max Planck Institute for Evolutionary Anthropology, Germany. Permission to conduct research was also provided by all the other facilities in which the tested primates were housed (the Animaya Zoo in Merida, Mexico for the spider monkeys, and the ISTC-CNR Primate Centre in Rome, Italy for the capuchin monkeys; no permission IDs were given). Because our study was purely behavioural/observational, no application to other ethic committees was required.
Funding statement
This work was conducted while the first author held a Humboldt Research Fellowship for Postdoctoral Researchers (Humboldt ID no. 1138999).
References
- 1.Cronin KA. 2012. Prosocial behaviour in animals: the influence of social relationships, communication and rewards. Anim. Behav. 84, 1085–1093. ( 10.1016/j.anbehav.2012.08.009) [DOI] [Google Scholar]
- 2.Henrich J, Boyd R, Bowles S, Camerer C, Fehr E, Gintis H, McElreath R. 2001. Cooperation, reciprocity and punishment in fifteen small-scale societies. Am. Econ. Rev. 91, 73–78. ( 10.1257/aer.91.2.73) [DOI] [Google Scholar]
- 3.Fehr E, Fischbacher U. 2003. The nature of human altruism. Nature 423, 785–791. ( 10.1038/nature02043) [DOI] [PubMed] [Google Scholar]
- 4.Fehr E, Fischbacher U, Gächter S. 2002. Strong reciprocity, human cooperation, and the enforcement of social norms. Hum. Nat. 13, 1–25. ( 10.1007/s12110-002-1012-7) [DOI] [PubMed] [Google Scholar]
- 5.Burton-Chellew MN, West SA. 2013. Prosocial preferences do not explain human cooperation in public-goods games. Proc. Natl Acad. Sci. USA 110, 216–221. ( 10.1073/pnas.1210960110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.de Waal FBM, Suchak M. 2010. Prosocial primates: selfish and unselfish motivations. Phil. Trans. R. Soc. B 365, 2711–2722. ( 10.1098/rstb.2010.0119) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sabbatini G, de Bortoli Vizioli A, Visalberghi E, Schino G. 2012. Food transfers in capuchin monkeys: an experiment on partner choice. Biol. Lett. 8, 757–759. ( 10.1098/rsbl.2012.0534) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hattori Y, Leimgruber K, Fujita K, de Waal FBM. 2012. Food-related tolerance in capuchin monkeys (Cebus apella) varies with knowledge of the partner's previous food-consumption. Behaviour 149, 171–185. ( 10.1163/156853912X634124) [DOI] [Google Scholar]
- 9.Warneken F, Tomasello M. 2006. Altruistic helping in human infants and young chimpanzees. Science 31, 1301–1303. ( 10.1126/science.1121448) [DOI] [PubMed] [Google Scholar]
- 10.Melis AP, Warneken F, Jensen K, Schneider A, Call J, Tomasello M. 2011. Chimpanzees help conspecifics obtain food and non-food items. Proc. R. Soc. B 278, 1405–1413. ( 10.1098/rspb.2010.1735) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Warneken F, Hare B, Melis AP, Hanus D, Tomasello M. 2007. Spontaneous altruism by chimpanzees and young children. PLoS Biol. 5, 181–187. ( 10.1371/journal.pbio.0050184) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Melis A, Tomasello M. 2013. Chimpanzees’ strategic helping in a collaborative task. Biol. Lett. 9, 1744–1748. ( 10.1098/rsbl.2013.0009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yamamoto S, Humle T, Tanaka M. 2012. Chimpanzees’ flexible targeted helping based on an understanding of conspecifics’ goals. Proc. Natl Acad. Sci. USA 109, 3588–3592. ( 10.1073/pnas.1108517109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Silk JB, Brosnan SF, Vonk J, Henrich J, Povinelli DJ, Richardson AS, Lambeth SP, Mascaro J, Schapiro SJ. 2005. Chimpanzees are indifferent to the welfare of unrelated group members. Nature 437, 1357–1359. ( 10.1038/nature04243) [DOI] [PubMed] [Google Scholar]
- 15.Jensen K, Hare B, Call J, Tomasello M. 2006. What's in it for me? Self-regard precludes altruism and spite in chimpanzees. Proc. R. Soc. B 273, 1013–1021. ( 10.1098/rspb.2005.3417) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vonk J, Brosnan SF, Silk JB, Henrich J, Richardson AS, Lambeth SP, Schapiro SJ, Povinelli DJ. 2008. Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members. Anim. Behav. 75, 1757–1770. ( 10.1016/j.anbehav.2007.09.036) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yamamoto S, Tanaka M. 2010. The influence of kin relationship and reciprocal context on chimpanzees’ other-regarding preferences. Anim. Behav. 79, 595–602. ( 10.1016/j.anbehav.2009.11.034) [DOI] [Google Scholar]
- 18.Massen J, van den Berg L, Spruijt B, Sterck EHM. 2010. Generous leaders and selfish underdogs: pro-sociality in despotic macaques. PLoS ONE 5, e9734 ( 10.1371/journal.pone.0009734) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Laskshminarayanan VR, Santos LR. 2008. Capuchin monkeys are sensitive to others’ welfare. Curr. Biol. 18, R999–R1000. ( 10.1016/j.cub.2008.08.057) [DOI] [PubMed] [Google Scholar]
- 20.Takimoto A, Kuroshima H, Fujita K. 2010. Capuchin monkeys (Cebus apella) are sensitive to others’ reward: an experimental analysis of food-choice for conspecifics. Anim. Cogn. 13, 249–261. ( 10.1007/s10071-009-0262-8) [DOI] [PubMed] [Google Scholar]
- 21.Burkart JM, Fehr E, Efferson C, van Schaik CP. 2007. Other-regarding preferences in a non-human primate: common marmosets provision food altruistically. Proc. Natl Acad. Sci. USA 104, 19 762–19 766. ( 10.1073/pnas.0710310104) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cronin KA, Schroeder KKE, Rothwell ES, Silk JB, Snowdon CT. 2009. Cottontop tamarins do not provide rewards to their long-term mates. J. Comp. Psychol. 123, 231–241. ( 10.1037/a0015094) [DOI] [PubMed] [Google Scholar]
- 23.Stevens JR. 2010. Donor payoffs and other-regarding preferences in cotton-top tamarins (Saguinus oedipus). Anim. Cogn. 13, 663–670. ( 10.1007/s10071-010-0309-x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Burkart JM, van Schaik C. 2013. Group service in macaques (Macaca fuscata), capuchins (Cebus apella) and marmosets (Callithrix jacchus): a comparative approach to identifying proactive prosocial motivations. J. Comp. Psychol. 127, 212–225. ( 10.1037/a0026392) [DOI] [PubMed] [Google Scholar]
- 25.Jaeggi AV, Burkart JM, van Schaik CP. 2010. On the psychology of cooperation in humans and other primates: combining the natural history and experimental evidence of prosociality. Phil. Trans. R. Soc. B 365, 2723–2735. ( 10.1098/rstb.2010.0118) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hirata S. 2007. Competitive and cooperative aspects of social intelligence in chimpanzees. Jpn J. Anim. Psychol. 57, 29–40. ( 10.2502/janip.57.1.1) [DOI] [Google Scholar]
- 27.Horner V, Carter JD, Suchak M, de Waal FBM. 2011. Spontaneous prosocial choice by chimpanzees. Proc. Natl Acad. Sci. USA 108, 13 847–13 851. ( 10.1073/pnas.1111088108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chang SWC, Winecoff AA, Platt ML. 2011. Vicarious reinforcement in rhesus macaques (Macaca mulatta). Front. Neurosci. 5, 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.de Waal FBM, Leimgruber K, Greenberg AR. 2008. Giving is self-rewarding for monkeys. Proc. Natl Acad. Sci. USA 105, 13 685–13 689. ( 10.1073/pnas.0807060105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Suchak M, de Waal FBM. 2012. Monkeys benefit from reciprocity without the cognitive burden. Proc. Natl Acad. Sci. USA 109, 15 191–15 196. ( 10.1073/pnas.1213173109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Deaner RO, van Schaik CP, Johnson V. 2006. Do some taxa have better domain-general cognition than others? A meta-analysis of nonhuman primate studies. Evol. Psychol. 4, 149–196. [Google Scholar]
- 32.Burkart JM, Hrdy SB, van Schaik CP. 2009. Cooperative breeding and human cognitive evolution. Evol. Anthropol. 18, 175–186. ( 10.1002/evan.20222) [DOI] [Google Scholar]
- 33.Burkart JM, van Schaik CP. 2010. Cognitive consequences of cooperative breeding in primates? Anim. Cogn. 13, 1–19. ( 10.1007/s10071-009-0263-7) [DOI] [PubMed] [Google Scholar]
- 34.Amici F, Call J, Aureli F. 2012. Aversion to violation of expectations of food distribution: the role of social tolerance and relative dominance in seven primate species. Behaviour 149, 345–368. ( 10.1163/156853912X637833) [DOI] [Google Scholar]
- 35.Kappeler PM, Pereira ME. 2003. Primate life histories and socioecology. Chicago, IL: University of Chicago Press. [Google Scholar]
- 36.Smuts BB, Cheney DL, Seyfarth RM, Wrangham RW, Struhsaker TT. 1987. Primate societies. Chicago, IL: University of Chicago Press. [Google Scholar]
- 37.Brosnan SF, de Waal FBM. 2003. Monkeys reject unequal pay. Nature 425, 297–299. ( 10.1038/nature01963) [DOI] [PubMed] [Google Scholar]
- 38.Fletcher GE. 2008. Attending to the outcome of others: disadvantageous inequity aversion in male capuchin monkeys (Cebus apella). Am. J. Primatol. 70, 901–905. ( 10.1002/ajp.20576) [DOI] [PubMed] [Google Scholar]
- 39.de Waal FBM. 1997. Food transfers through mesh in brown capuchins. J. Comp. Psychol. 111, 370–378. ( 10.1037/0735-7036.111.4.370) [DOI] [PubMed] [Google Scholar]
- 40.Braeuer J, Call J, Tomasello M. 2007. Chimpanzees really know what others can see in a competitive situation. Anim. Cogn. 10, 439–448. ( 10.1007/s10071-007-0088-1) [DOI] [PubMed] [Google Scholar]
- 41.Cronin KA, Schroeder KKE, Rothwell ES, Silk JB, Snowdon C. 2009. Cooperatively breeding cottontop tamarins (Saguinus oedipus) do not donate rewards to their long-term mates. J. Comp. Psychol. 123, 231–241. ( 10.1037/a0015094) [DOI] [PubMed] [Google Scholar]
- 42.Braeuer J, Call J, Tomasello M. 2009. Are apes inequity averse? New data on the token exchange paradigm. Am. J. Primatol. 7, 175–181. ( 10.1002/ajp.20639) [DOI] [PubMed] [Google Scholar]
- 43.Addessi E, Paglieri F, Focaroli V. 2011. The ecological rationality of delay tolerance: insights from capuchin monkeys. Cognition 119, 142–147. ( 10.1016/j.cognition.2010.10.021) [DOI] [PubMed] [Google Scholar]
- 44.Addessi E, Rossi S. 2011. Tokens improve capuchin performance in the reverse-reward contingency task. Proc. R. Soc. B 278, 849–854. ( 10.1098/rspb.2010.1602) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dufour V, Pelé M, Neumann M, Thierry B, Call J. 2009. Calculated reciprocity after all: computation behind token transfers in orangutans. Biol. Lett. 5, 172–175. ( 10.1098/rsbl.2008.0644) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pelé M, Dufour V, Thierry B, Call J. 2009. Token transfers among great apes: species differences, gestural requests and reciprocal exchange. J. Comp. Psychol. 123, 375–384. ( 10.1037/a0017253) [DOI] [PubMed] [Google Scholar]
- 47.Morton FB, Lee P, Buchanan-Smith HM. 2013. Taking personality selection bias seriously in animal cognition research: a case study in monkeys (Sapajus apella). Anim. Cogn. 16, 677–684. ( 10.1007/s10071-013-0603-5) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.